
Antibacterial and Sporicidal Activity Evaluation of Theaflavin-3,3′-digallate


Abstract
:1. Introduction
2. Results
2.1. Determination of MIC and Half-Maximal Inhibitory Concentration (IC50)
2.2. Colony Forming Unit (CFU) Assay
2.3. BacTiter-GloTM Microbial Cell Viability Assay
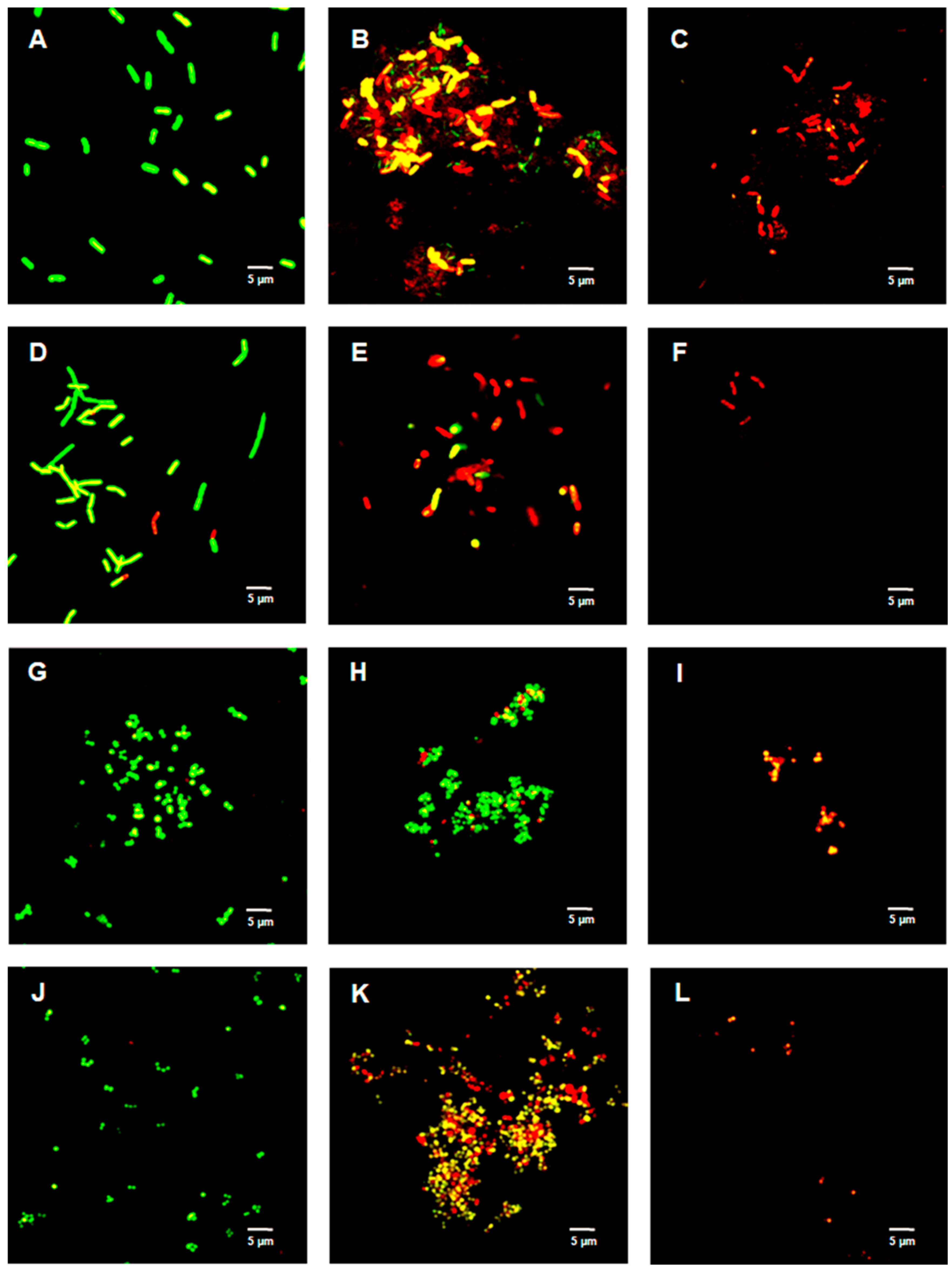
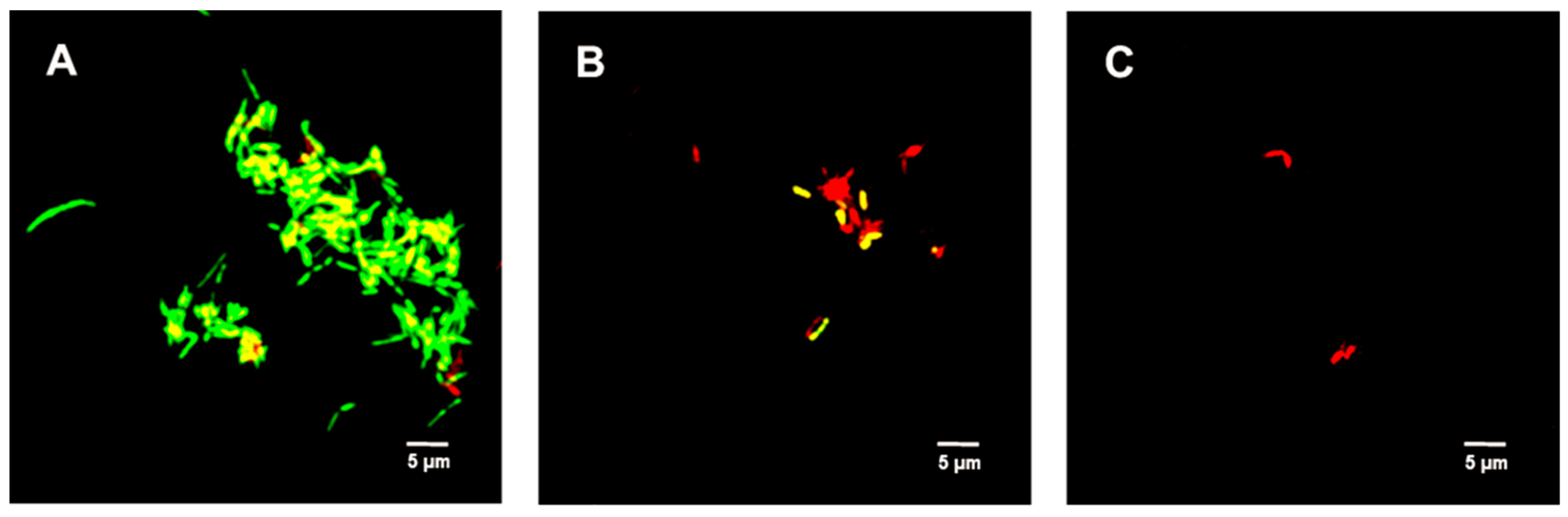
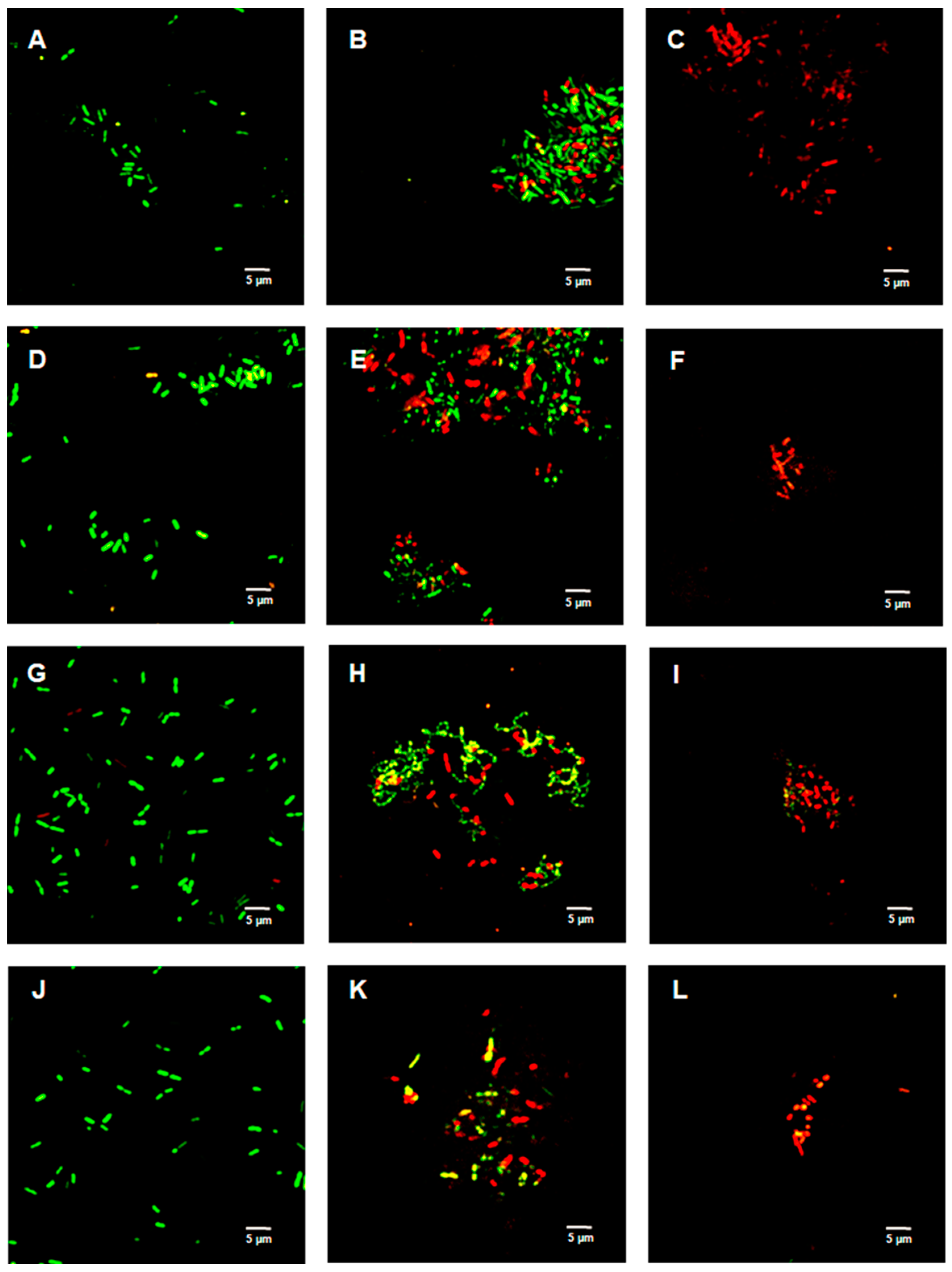
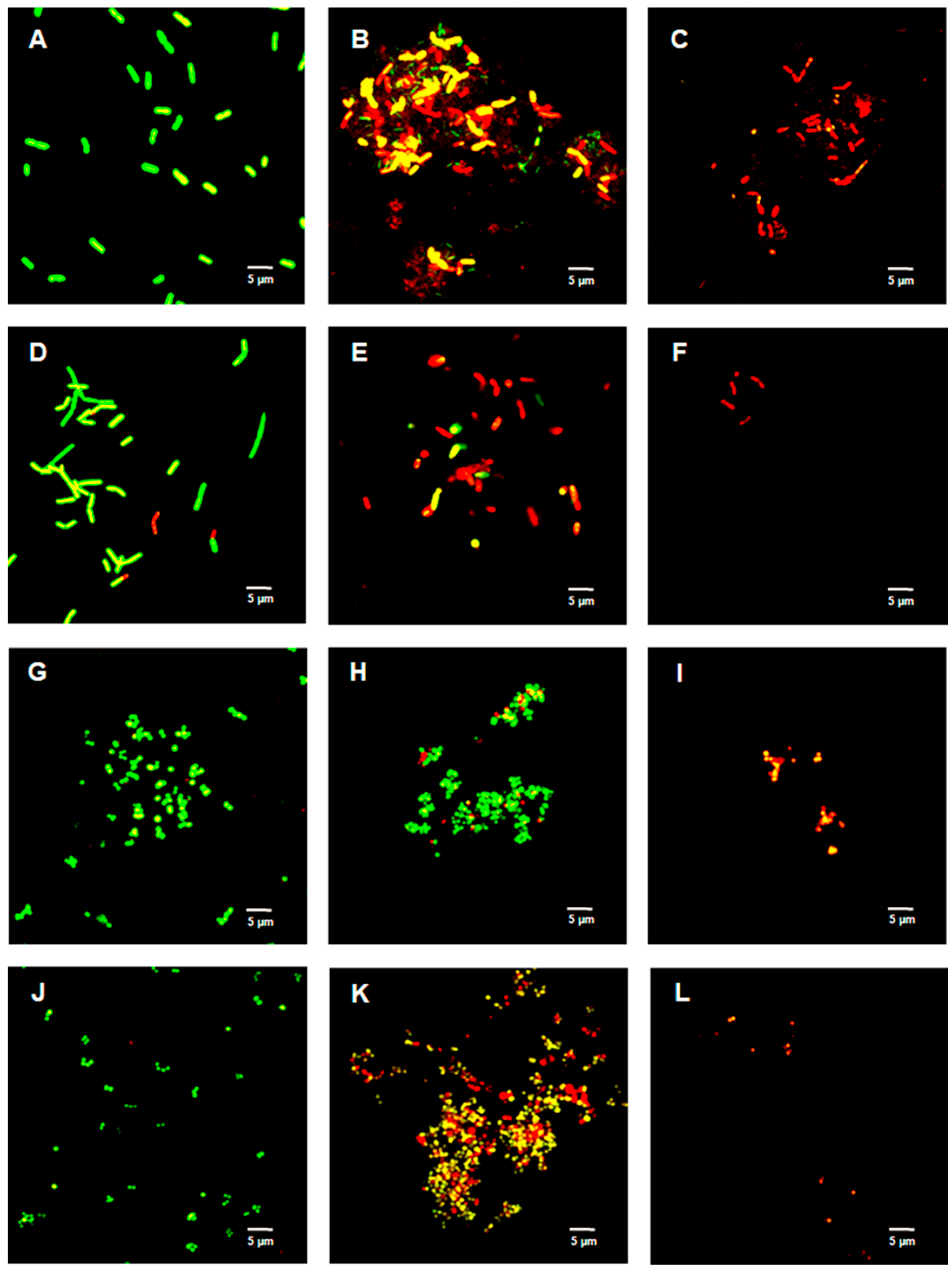
2.4. Live/Dead Bacterial Viability Assay
2.5. Germination Inhibition via CFU Assay
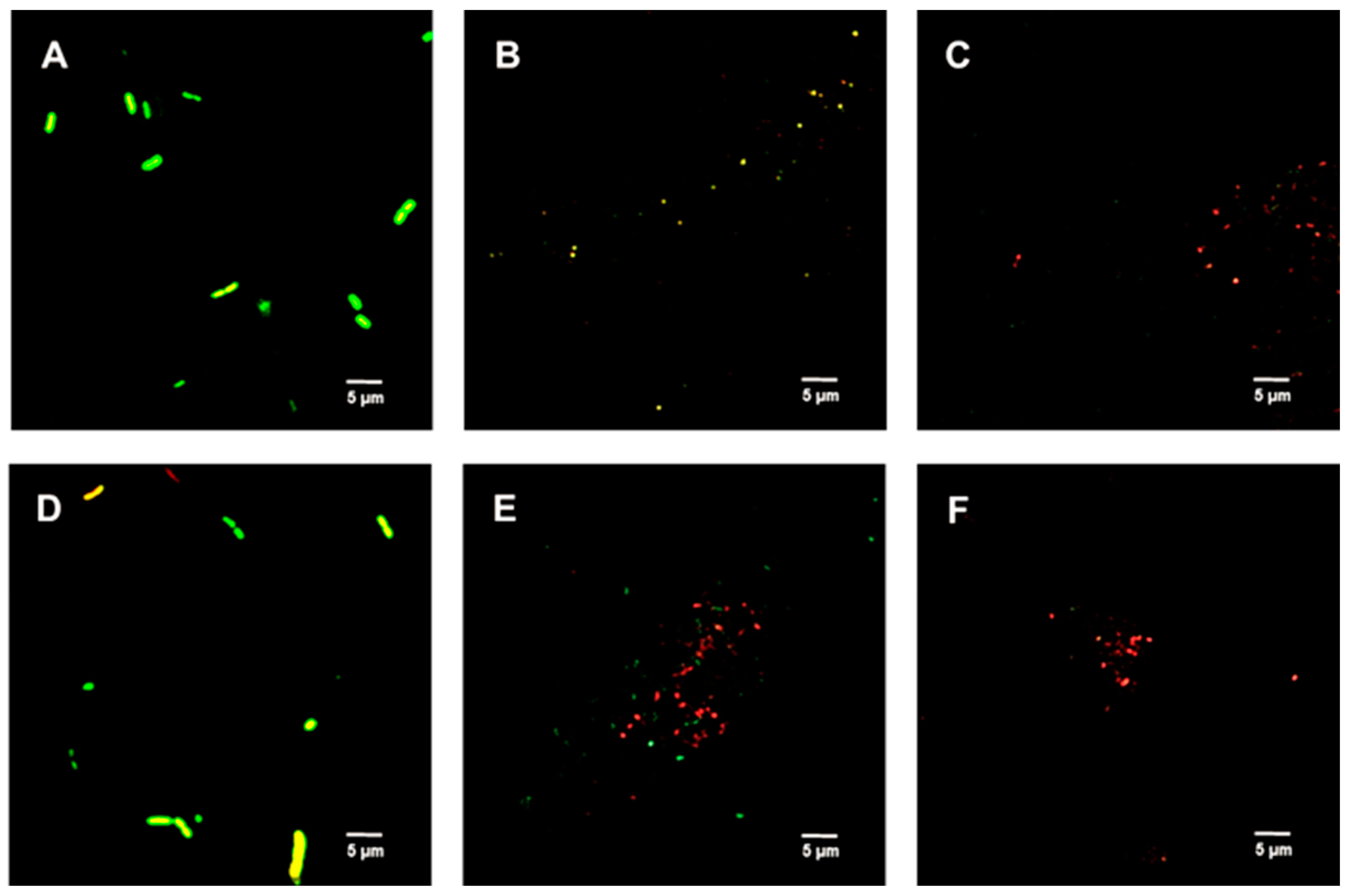
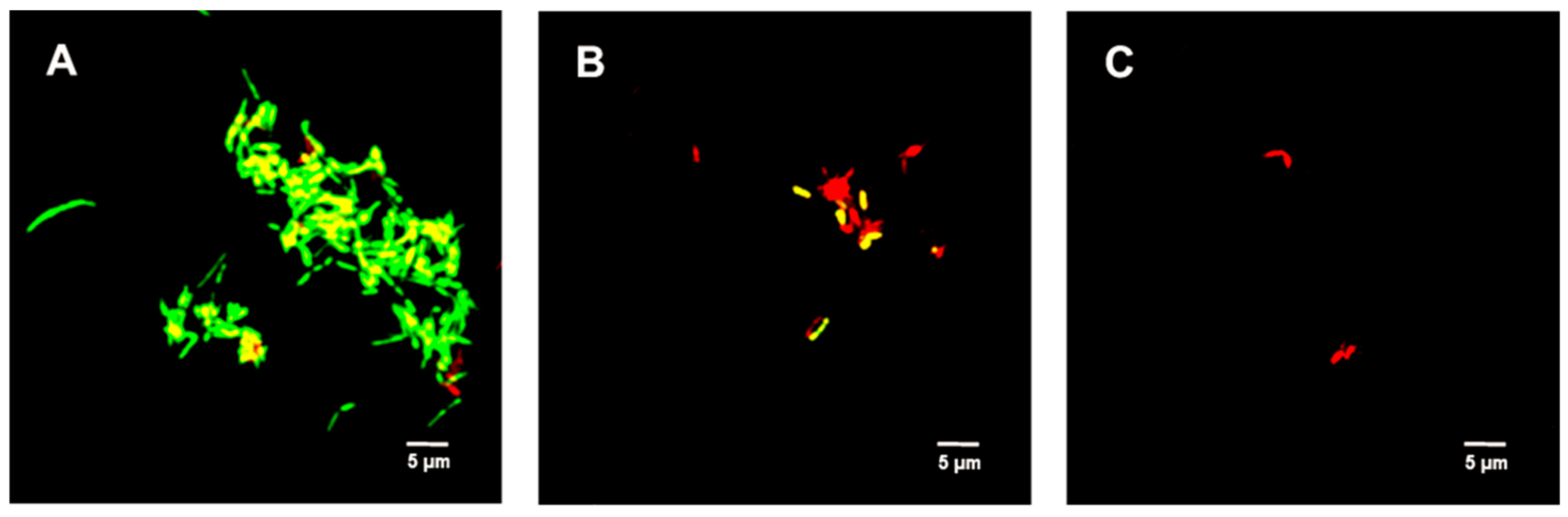
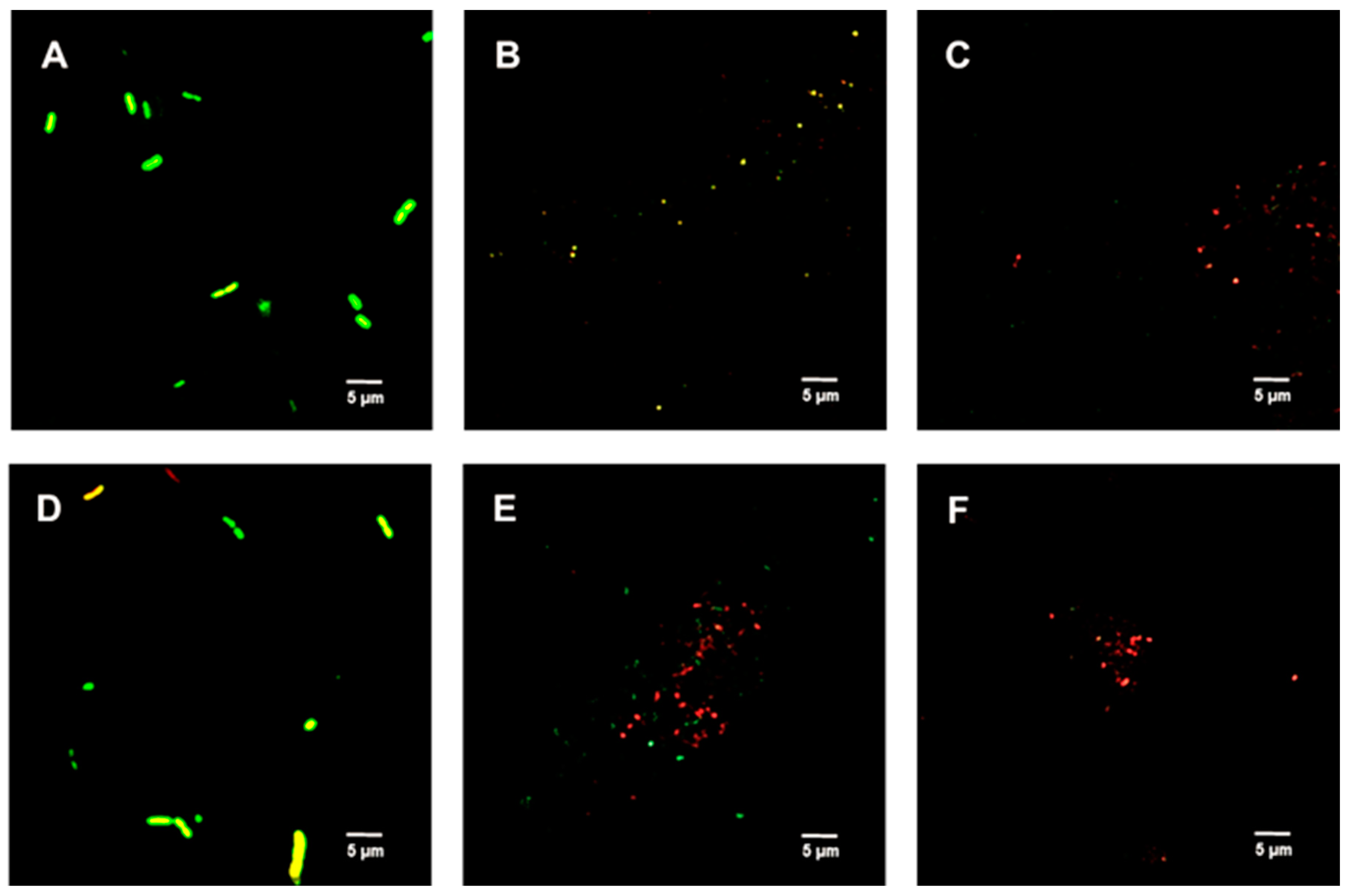
2.6. Live/Dead Spore Viability Assay
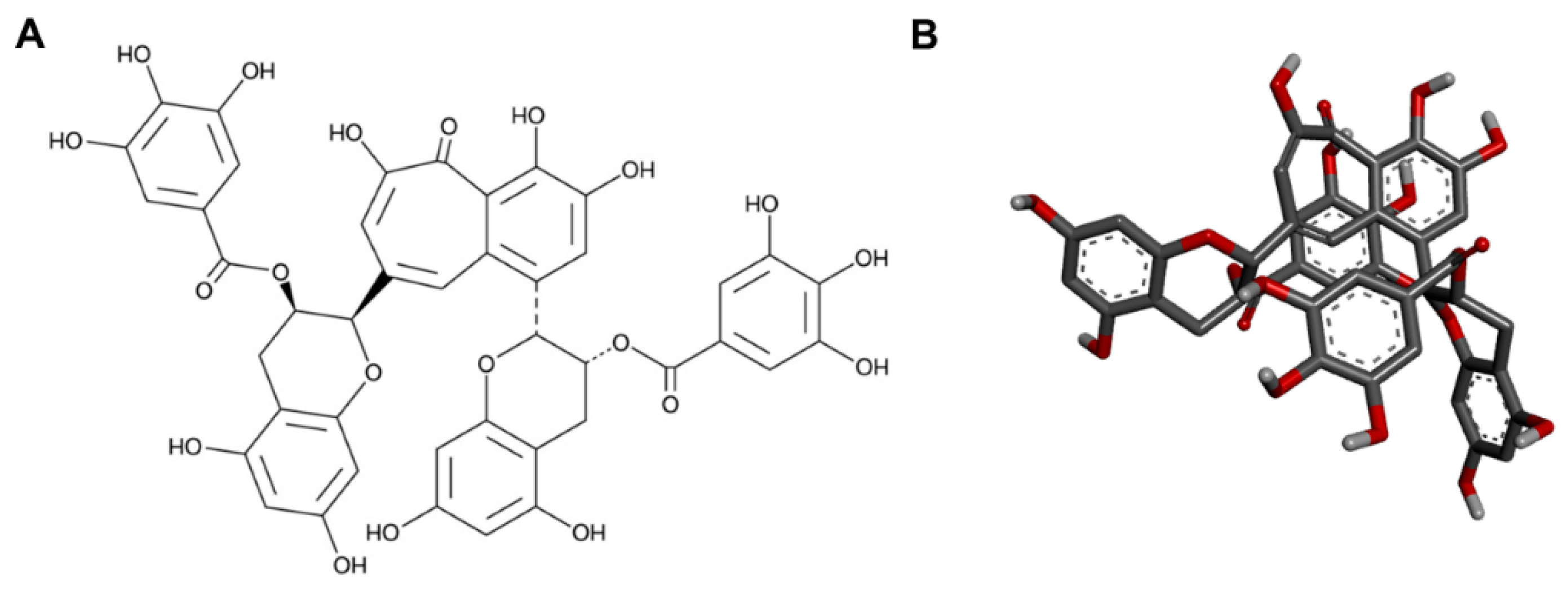
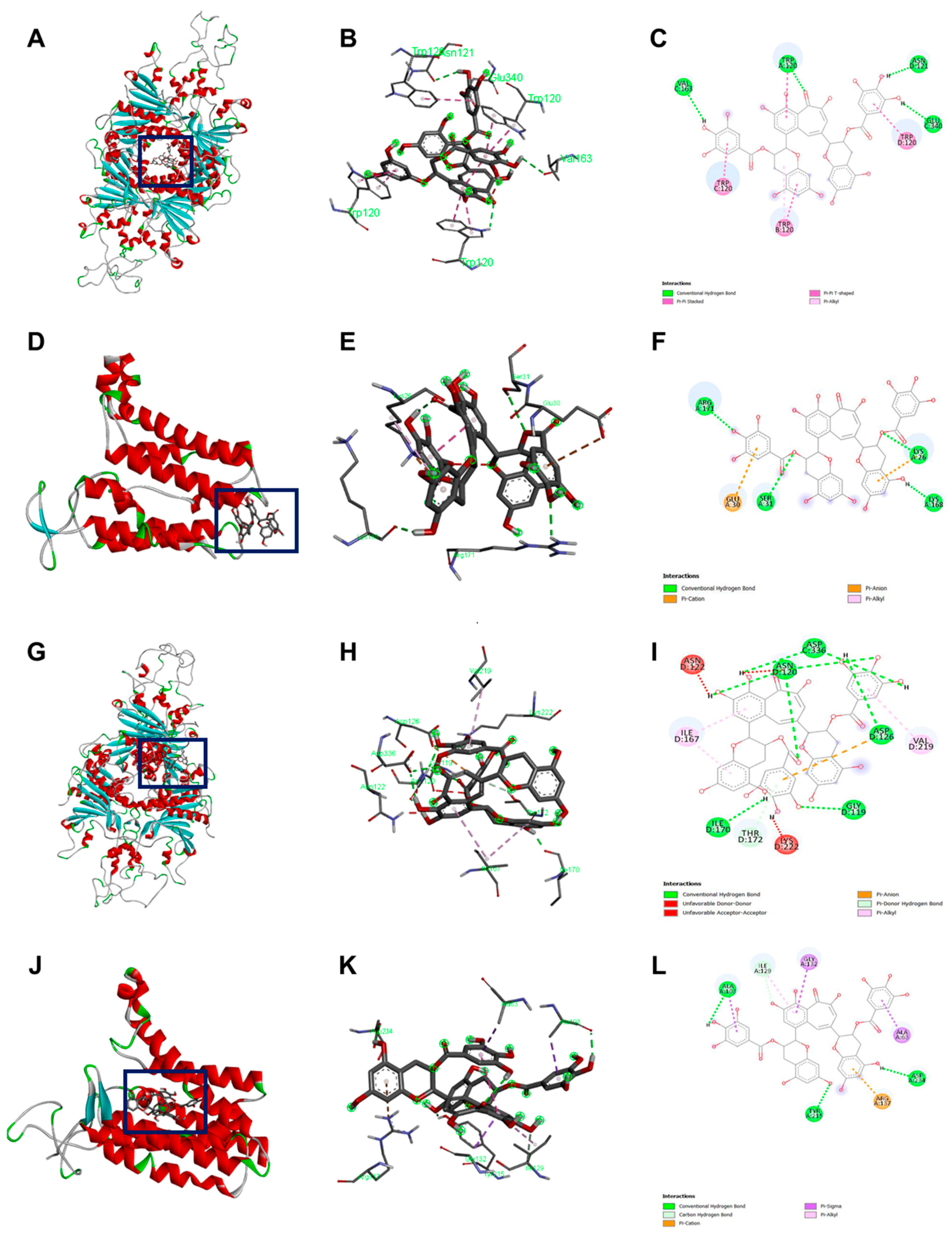
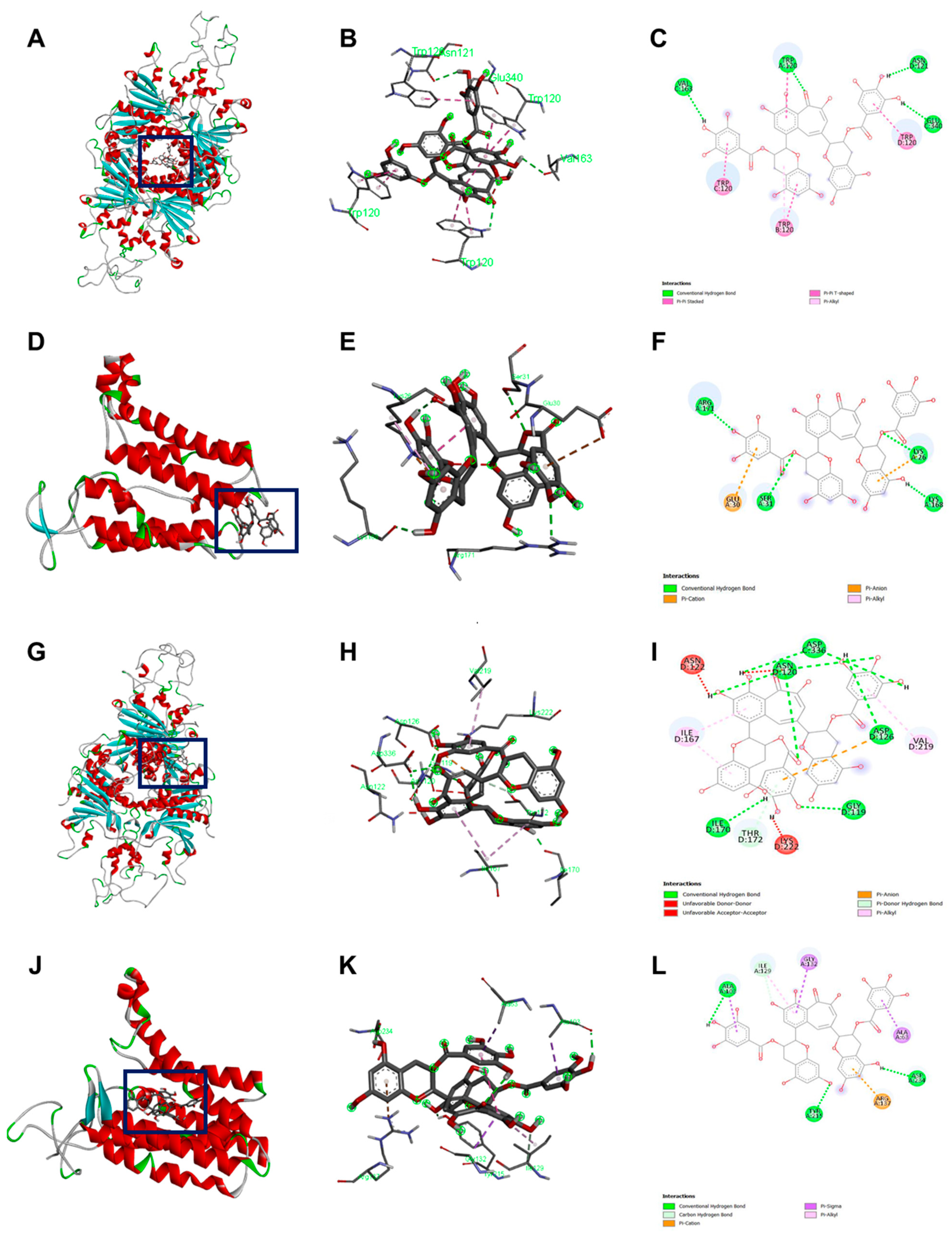
2.7. Binding Pocket
2.8. In Silico Docking Analysis
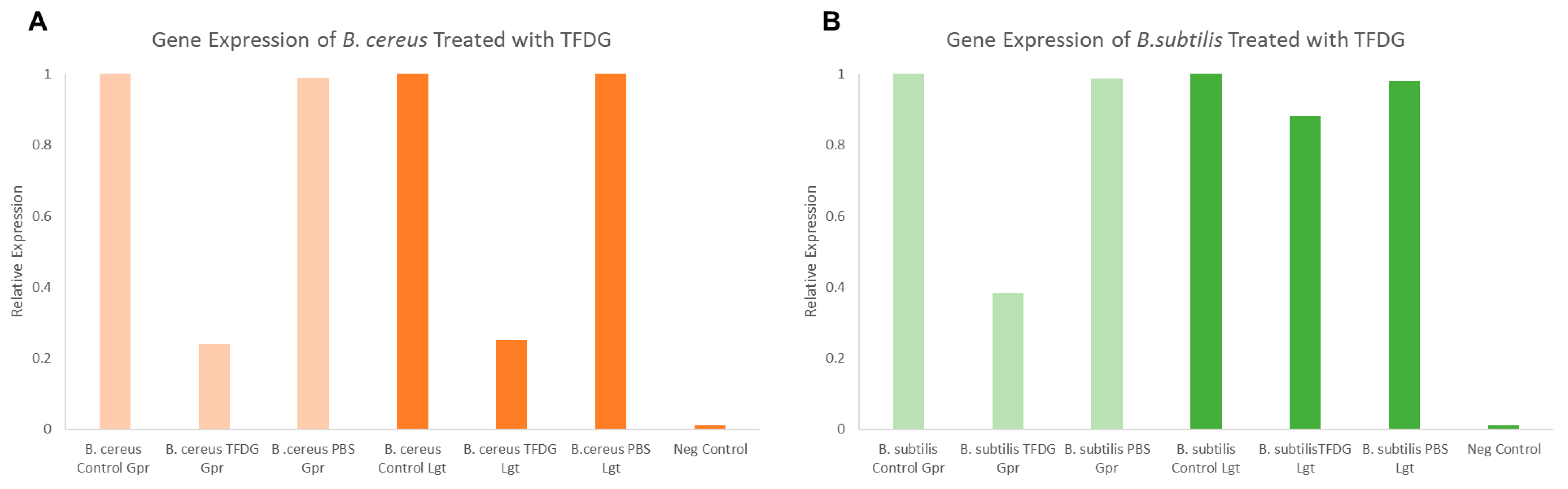
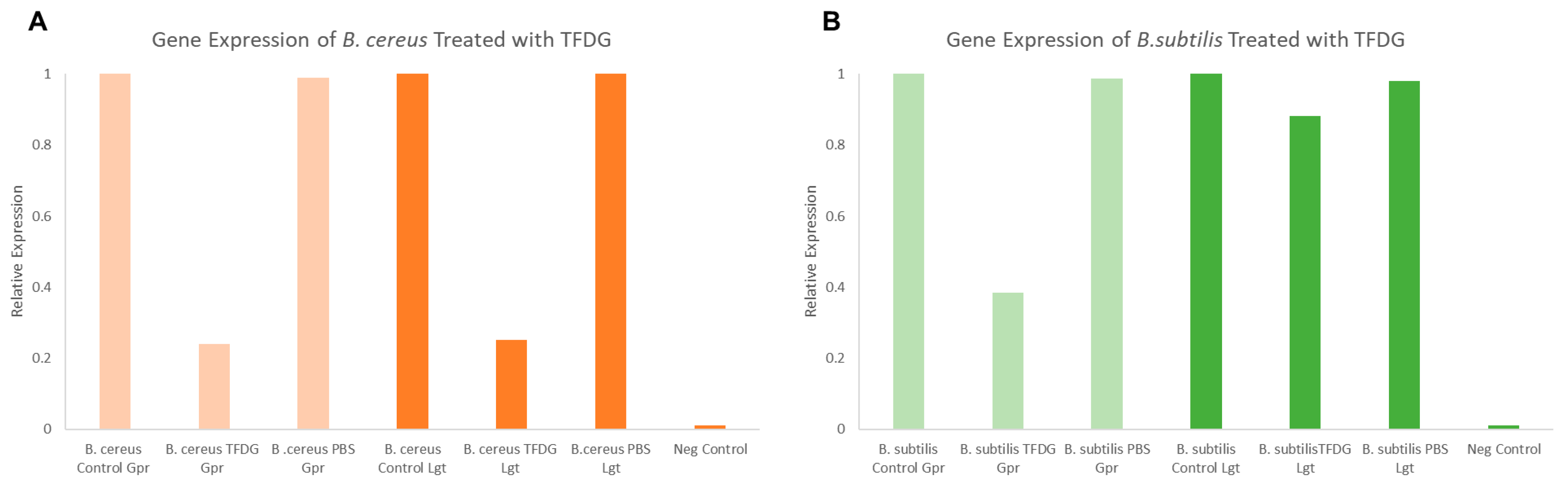
2.9. Semi-Quantitative RT-PCR
3. Discussion
4. Materials and Methods
4.1. Bacteria Culture
4.2. Culture Maintenance
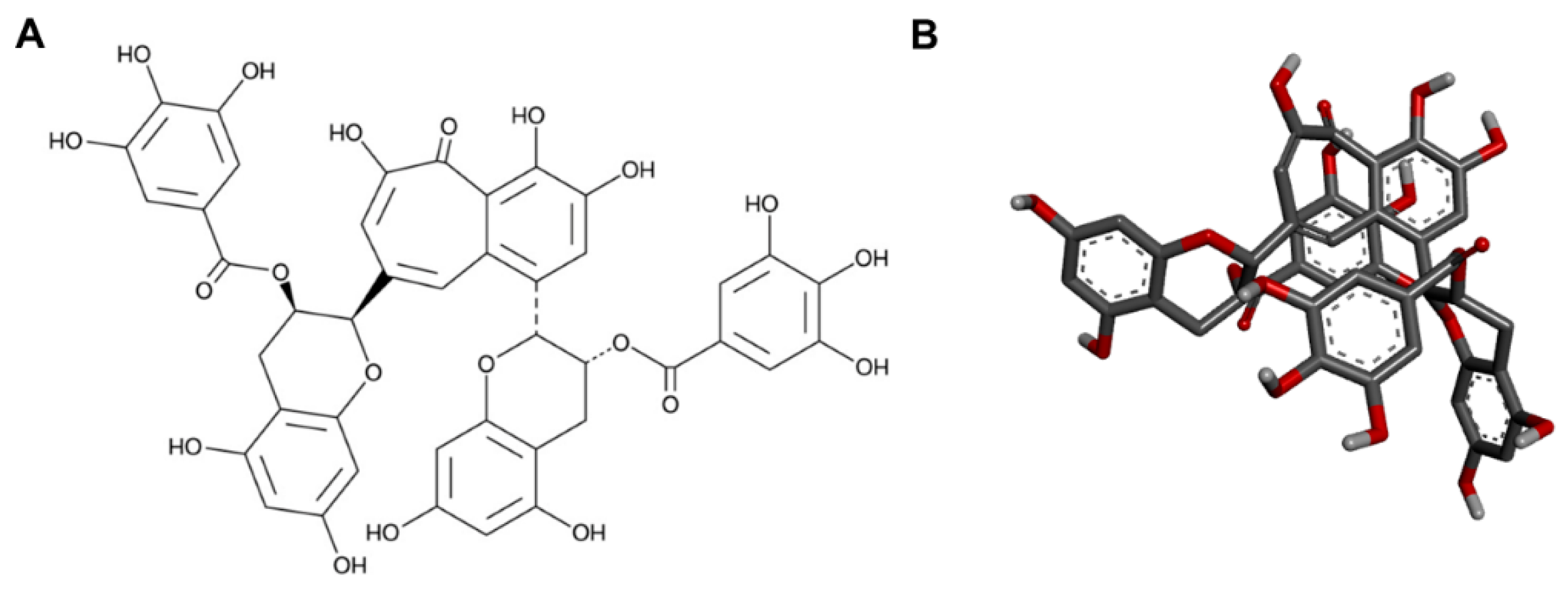
4.3. Theaflavin Preparation
4.4. Microplate Assay
4.5. Colony Forming Unit (CFU) Assay
4.6. BacTiter-GloTM Microbial Cell Viability Assay
4.7. LIVE/DEADTM BacLightTM Bacterial Viability Assay
4.8. Spore Preparation
4.9. Spore Germination Inhibition Assay
4.10. Ligand Preparation
4.11. Gene and Protein Selection
4.12. In Silico Docking Analysis and Visualization
4.13. Total RNA Extraction, cDNA Synthesis, and Semi-Quantitative RT-PCR
4.14. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Piddock, L.J.V. The crisis of no new antibiotics—What is the way forward? Lancet Infect. Dis. 2012, 12, 249–253. [Google Scholar] [CrossRef]
- Ventola, C.L. The Antibiotic Resistance Crisis Part 1: Causes and Threats. Pharm. Ther. 2015, 40, 277–283. [Google Scholar]
- CDC. Antibiotic Resistance Threats in the United States; US Department of Health and Human Services, CDC: Atlanta, GA, USA, 2019. [CrossRef] [Green Version]
- Martens, E.; Demain, A.L. The antibiotic resistance crisis, with a focus on the United States. J. Antibiot. 2017, 70, 520–526. [Google Scholar] [CrossRef] [Green Version]
- Hilal, Y.; Engelhardt, U. Characterisation of white tea—Comparison to green and black tea. J. Verbrauch. Lebensm. 2007, 2, 414–421. [Google Scholar] [CrossRef]
- Babich, H.; Gottesman, R.T.; Liebling, E.J.; Schuck, A.G. Theaflavin-3-gallate and theaflavin-3′-gallate, polyphenols in black tea with prooxidant properties. Basic Clin. Pharmacol. Toxicol. 2008, 103, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Pereira-Caro, G.; Moreno-Rojas, J.M.; Brindani, N.; Del Rio, D.; Lean, M.E.J.; Hara, Y.; Crozier, A. Bioavailability of Black Tea Theaflavins: Absorption, Metabolism, and Colonic Catabolism. J. Agric. Food Chem. 2017, 65, 5365–5374. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, A.; Prince, D.; Lo, C.Y.; Lee, L.H.; Chu, T.C. Antiviral activity of theaflavin digallate against herpes simplex virus type 1. Antivir. Res. 2015, 118, 56–67. [Google Scholar] [CrossRef]
- Wang, S.; Wang, Y.; Wang, Y.; Duan, Z.; Ling, Z.; Wu, W.; Tong, S.; Wang, H.; Deng, S. Theaflavin-3,3′-Digallate Suppresses Biofilm Formation, Acid Production, and Acid Tolerance in Streptococcus mutans by Targeting Virulence Factors. Front. Microbiol. 2019, 10, 1705. [Google Scholar] [CrossRef] [Green Version]
- Engelhardt, U.H. Chemistry of Tea. In Comprehensive Natural Products II; Elsevier Science: Amsterdam, The Netherlands, 2010; pp. 999–1032. [Google Scholar]
- Peterson, J.; Dwyer, J.; Jacques, P.; Rand, W.; Prior, R.; Chui, K. Tea variety and brewing techniques influence flavonoid content of black tea. J. Food Compos. Anal. 2004, 17, 397–405. [Google Scholar] [CrossRef]
- Leung, L.K.; Su, Y.; Chen, R.; Zhang, Z.; Huang, Y.; Chen, Z.-Y. Theaflavins in Black Tea and Catechins in Green Tea Are Equally Effective Antioxidants. J. Nutr. 2001, 131, 2248–2251. [Google Scholar] [CrossRef]
- Noor Mohammadi, T.; Maung, A.T.; Sato, J.; Sonoda, T.; Masuda, Y.; Honjoh, K.; Miyamoto, T. Mechanism for antibacterial action of epigallocatechin gallate and theaflavin-3,3′-digallate on Clostridium perfringens. J. Appl. Microbiol. 2019, 126, 633–640. [Google Scholar] [CrossRef] [PubMed]
- Hui, X.; Yue, Q.; Zhang, D.D.; Li, H.; Yang, S.Q.; Gao, W.Y. Antimicrobial mechanism of theaflavins: They target 1-deoxy-D-xylulose 5-phosphate reductoisomerase, the key enzyme of the MEP terpenoid biosynthetic pathway. Sci. Rep. 2016, 6, 38945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teng, Z.; Guo, Y.; Liu, X.; Zhang, J.; Niu, X.; Yu, Q.; Deng, X.; Wang, J. Theaflavin-3,3-digallate increases the antibacterial activity of β-lactam antibiotics by inhibiting metallo-beta-lactamase activity. J. Cell Mol. Med. 2019, 23, 6955–6964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, B.; Lee, L.H.; Laskar, N.; Shaikh, N.; Tahir, H.; Hsu, S.D.; Newby, R., Jr.; Valsechi-Diaz, J.; Chu, T. Modified Green Tea Polyphenols, EGCG-S and LTP, Inhibit Endospore in Three Bacillus spp. Adv. Microbiol. 2017, 7, 175–187. [Google Scholar] [CrossRef] [Green Version]
- Renzetti, A.; Betts, J.W.; Fukumoto, K.; Rutherford, R.N. Antibacterial green tea catechins from a molecular perspective: Mechanisms of action and structure-activity relationships. Food Funct. 2020, 11, 9370–9396. [Google Scholar] [CrossRef]
- Ben Lagha, A.; Grenier, D. Black tea theaflavins attenuate Porphyromonas gingivalis virulence properties, modulate gingival keratinocyte tight junction integrity and exert anti-inflammatory activity. J. Periodontal Res. 2017, 52, 458–470. [Google Scholar] [CrossRef]
- Chowdhury, P.; Sahuc, M.E.; Rouille, Y.; Riviere, C.; Bonneau, N.; Vandeputte, A.; Brodin, P.; Goswami, M.; Bandyopadhyay, T.; Dubuisson, J.; et al. Theaflavins, polyphenols of black tea, inhibit entry of hepatitis C virus in cell culture. PLoS ONE 2018, 13, e0198226. [Google Scholar] [CrossRef] [Green Version]
- Kong, J.; Zhang, G.; Xia, K.; Diao, C.; Yang, X.; Zuo, X.; Li, Y.; Liang, X. Tooth brushing using toothpaste containing theaflavins reduces the oral pathogenic bacteria in healthy adults. 3 Biotech 2021, 11, 150. [Google Scholar] [CrossRef]
- Wang, M.; Li, J.; Hu, T.; Zhao, H. Metabolic fate of tea polyphenols and their crosstalk with gut microbiota. Food Sci. Hum. Wellness 2022, 11, 455–466. [Google Scholar] [CrossRef]
- Passarelli-Araujo, H.; Palmeiro, J.K.; Moharana, K.C.; Pedrosa-Silva, F.; Dalla-Costa, L.M.; Venancio, T.M. Genomic analysis unveils important aspects of population structure, virulence, and antimicrobial resistance in Klebsiella aerogenes. FEBS J. 2019, 286, 3797–3810. [Google Scholar] [CrossRef]
- Kaper, J.B.; Nataro, J.P.; Mobley, H.L. Pathogenic Escherichia coli. Nat. Rev. Microbiol. 2004, 2, 123–140. [Google Scholar] [CrossRef] [PubMed]
- Sondi, I.; Salopek-Sondi, B. Silver nanoparticles as antimicrobial agent: A case study on E. coli as a model for Gram-negative bacteria. J. Colloid Interface Sci. 2004, 275, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Blount, Z.D. The Natural History of Model Organisms: The unexhausted potential of E. coli. elife 2015, 4, e05826. [Google Scholar] [CrossRef] [PubMed]
- Iakovides, I.C.; Michael-Kordatou, I.; Moreira, N.F.F.; Ribeiro, A.R.; Fernandes, T.; Pereira, M.F.R.; Nunes, O.C.; Manaia, C.M.; Silva, A.M.T.; Fatta-Kassinos, D. Continuous ozonation of urban wastewater: Removal of antibiotics, antibiotic-resistant Escherichia coli and antibiotic resistance genes and phytotoxicity. Water Res. 2019, 159, 333–347. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.M.; Xu, L.M.; Wang, X.C.; Zhuang, K.; Liu, Q.Q. Effects of ultraviolet disinfection on antibiotic-resistant Escherichia coli from wastewater: Inactivation, antibiotic resistance profiles and antibiotic resistance genes. J. Appl. Microbiol. 2017, 123, 295–306. [Google Scholar] [CrossRef] [PubMed]
- Roth, N.; Kasbohrer, A.; Mayrhofer, S.; Zitz, U.; Hofacre, C.; Domig, K.J. The application of antibiotics in broiler production and the resulting antibiotic resistance in Escherichia coli: A global overview. Poult. Sci. 2019, 98, 1791–1804. [Google Scholar] [CrossRef]
- Davis, G.S.; Waits, K.; Nordstrom, L.; Grande, H.; Weaver, B.; Papp, K.; Horwinski, J.; Koch, B.; Hungate, B.A.; Liu, C.M.; et al. Antibiotic-resistant Escherichia coli from retail poultry meat with different antibiotic use claims. BMC Microbiol. 2018, 18, 174. [Google Scholar] [CrossRef]
- Poole, K. Pseudomonas aeruginosa: Resistance to the max. Front. Microbiol. 2011, 2, 65. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, A.L.; Kamm, M.A.; Ng, S.C.; Morrison, M. Proteus spp. as Putative Gastrointestinal Pathogens. Clin. Microbiol. Rev. 2018, 31, e00085-17. [Google Scholar] [CrossRef] [Green Version]
- Cohen-Nahum, K.; Saidel-Odes, L.; Riesenberg, K.; Schlaeffer, F.; Borer, A. Urinary tract infections caused by multi-drug resistant Proteus mirabilis: Risk factors and clinical outcomes. Infection 2010, 38, 41–46. [Google Scholar] [CrossRef]
- Ehling-Schulz, M.; Lereclus, D.; Koehler, T.M. The Bacillus cereus Group: Bacillus Species with Pathogenic Potential. Microbiol. Spectr. 2019, 7. [Google Scholar] [CrossRef] [PubMed]
- Errington, J.; Aart, L.T.V. Microbe Profile: Bacillus subtilis: Model organism for cellular development, and industrial workhorse. Microbiology 2020, 166, 425–427. [Google Scholar] [CrossRef] [PubMed]
- Pogmore, A.R.; Seistrup, K.H.; Strahl, H. The Gram-positive model organism Bacillus subtilis does not form microscopically detectable cardiolipin-specific lipid domains. Microbiology 2018, 164, 475–482. [Google Scholar] [CrossRef] [PubMed]
- Setlow, P. Germination of Spores of Bacillus Species: What We Know and Do Not Know. J. Bacteriol. 2014, 196, 1297–1305. [Google Scholar] [CrossRef] [Green Version]
- Keijser, B.J.; Ter Beek, A.; Rauwerda, H.; Schuren, F.; Montijn, R.; van der Spek, H.; Brul, S. Analysis of temporal gene expression during Bacillus subtilis spore germination and outgrowth. J. Bacteriol. 2007, 189, 3624–3634. [Google Scholar] [CrossRef] [Green Version]
- Levinson, H.S.; Hyatt, M. Effects of Temperature on Activation, Germination, and Outgrowth of Bacillus megaterium Spores. J. Bacteriol. 1970, 101, 56–64. [Google Scholar] [CrossRef] [Green Version]
- Hornstra, L.M.; de Vries, Y.P.; Wells-Bennik, M.H.; de Vos, W.M.; Abee, T. Characterization of germination receptors of Bacillus cereus ATCC 14579. Appl. Environ. Microbiol. 2006, 72, 44–53. [Google Scholar] [CrossRef] [Green Version]
- Traag, B.A.; Pugliese, A.; Eisen, J.A.; Losick, R. Gene Conservation among Endospore-Forming Bacteria Reveals Additional Sporulation Genes in Bacillus subtilis. J. Bacteriol. 2013, 195, 253–260. [Google Scholar] [CrossRef] [Green Version]
- Seo, Y.S.; Lee, D.Y.; Rayamahji, N.; Kang, M.L.; Yoo, H.S. Biofilm-forming associated genotypic and phenotypic characteristics of Staphylococcus spp. isolated from animals and air. Res. Vet. Sci. 2008, 85, 433–438. [Google Scholar] [CrossRef]
- Sriskandan, S.; Faulkner, L.; Hopkins, P. Streptococcus pyogenes: Insight into the function of the streptococcal superantigens. Int. J. Biochem. Cell Biol. 2007, 39, 12–19. [Google Scholar] [CrossRef]
- Zhu, L.; Olsen, R.J.; Horstmann, N.; Shelburne, S.A.; Fan, J.; Hu, Y.; Musser, J.M. Intergenic Variable-Number Tandem-Repeat Polymorphism Upstream of rocA Alters Toxin Production and Enhances Virulence in Streptococcus pyogenes. Infect. Immun. 2016, 84, 2086–2093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forbes, B.A. Mycobacterial Taxonomy. J. Clin. Microbiol. 2017, 55, 380–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO. W.H.O. Global Tuberculosis Report; World Health Organization: Geneva, Switzerland, 2020; pp. 1–232. [Google Scholar]
- Lelovic, N.; Mitachi, K.; Yang, J.; Lemieux, M.R.; Ji, Y.; Kurosu, M. Application of Mycobacterium smegmatis as a surrogate to evaluate drug leads against Mycobacterium tuberculosis. J. Antibiot. 2020, 73, 780–789. [Google Scholar] [CrossRef] [PubMed]
- Altaf, M.; Miller, C.H.; Bellows, D.S.; O’Toole, R. Evaluation of the Mycobacterium smegmatis and BCG models for the discovery of Mycobacterium tuberculosis inhibitors. Tuberculosis 2010, 90, 333–337. [Google Scholar] [CrossRef]
- Kwofie, S.K.; Broni, E.; Asiedu, S.O.; Kwarko, G.B.; Dankwa, B.; Enninful, K.S.; Tiburu, E.K.; Wilson, M.D. Cheminformatics-Based Identification of Potential Novel Anti-SARS-CoV-2 Natural Compounds of African Origin. Molecules 2021, 26, 406. [Google Scholar] [CrossRef]
- Chang, M.W.; Lindstrom, W.; Olson, A.J.; Belew, R.K. Analysis of HIV Wild-Type and Mutant Structures via in Silico Docking against Diverse Ligand Libraries. J. Chem. Inf. Model. 2007, 47, 1258–1262. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; Oezguen, N.; Urvil, P.; Ferguson, C.; Dann, S.M.; Savidge, T.C. Regulation of protein-ligand binding affinity by hydrogen bond pairing. Sci. Adv. 2016, 2, e1501240. [Google Scholar] [CrossRef] [Green Version]
- Ardal, C.; Balasegaram, M.; Laxminarayan, R.; McAdams, D.; Outterson, K.; Rex, J.H.; Sumpradit, N. Antibiotic development—Economic, regulatory and societal challenges. Nat. Rev. Microbiol. 2020, 18, 267–274. [Google Scholar] [CrossRef]
- Mempin, R.; Tran, H.; Chen, C.; Gong, H.; Kim Ho, K.; Lu, S. Release of extracellular ATP by bacteria during growth. BMC Microbiol. 2013, 13, 301. [Google Scholar] [CrossRef] [Green Version]
- Reyneke, B.; Dobrowsky, P.H.; Ndlovu, T.; Khan, S.; Khan, W. EMA-qPCR to monitor the efficiency of a closed-coupled solar pasteurization system in reducing Legionella contamination of roof-harvested rainwater. Sci. Total Environ. 2016, 553, 662–670. [Google Scholar] [CrossRef]
- Ignasimuthu, K.; Prakash, R.; Murthy, P.S.; Subban, N. Enhanced bioaccessibility of green tea polyphenols and lipophilic activity of EGCG octaacetate on gram-negative bacteria. LWT 2019, 105, 103–109. [Google Scholar] [CrossRef]
- Santajit, S.; Indrawattana, N. Mechanisms of Antimicrobial Resistance in ESKAPE Pathogens. Biomed. Res. Int. 2016, 2016, 2475067. [Google Scholar] [CrossRef] [Green Version]
- Feng, S.; Eucker, T.P.; Holly, M.K.; Konkel, M.E.; Lu, X.; Wang, S. Investigating the responses of Cronobacter sakazakii to garlic-drived organosulfur compounds: A systematic study of pathogenic-bacterium injury by use of high-throughput whole-transcriptome sequencing and confocal micro-raman spectroscopy. Appl. Environ. Microbiol. 2014, 80, 959–971. [Google Scholar] [CrossRef] [Green Version]
- Shigemune, N.; Nakayama, M.; Tsugukuni, T.; Hitomi, J.; Yoshizawa, C.; Mekada, Y.; Kurahachi, M.; Miyamoto, T. The mechanisms and effect of epigallocatechin gallate (EGCg) on the germination and proliferation of bacterial spores. Food Control 2012, 27, 269–274. [Google Scholar] [CrossRef]
- Tian, W.; Chen, C.; Lei, X.; Zhao, J.; Liang, J. CASTp 3.0: Computed atlas of surface topography of proteins. Nucleic Acids Res. 2018, 46, W363–W367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kukol, A. Consensus virtual screening approaches to predict protein ligands. Eur. J. Med. Chem. 2011, 46, 4661–4664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marone, M.; Mozzetti, S.; Ritis, D.D.; Pierelli, L.; Scambia, G. Semiquantitative RT-PCR analysis to assess the expression levels of multiple transcripts from the same sample. Biol. Proced. Online 2001, 3, 19–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, Y.; Francke, C.; Abee, T.; Wells-Bennik, M.H. Clostridial spore germination versus bacilli: Genome mining and current insights. Food Microbiol. 2011, 28, 266–274. [Google Scholar] [CrossRef]
- Norsigian, C.J.; Danhof, H.A.; Brand, C.K.; Oezguen, N.; Midani, F.S.; Palsson, B.O.; Savidge, T.C.; Britton, R.A.; Spinler, J.K.; Monk, J.M. Systems biology analysis of the Clostridioides difficile core-genome contextualizes microenvironmental evolutionary pressures leading to genotypic and phenotypic divergence. NPJ Syst. Biol. Appl. 2020, 6, 31. [Google Scholar] [CrossRef]
- Igarashi, T.; Setlow, B.; Paidhungat, M.; Setlow, P. Effects of a gerF (lgt) mutation on the germination of spores of Bacillus subtilis. J. Bacteriol. 2004, 186, 2984–2991. [Google Scholar] [CrossRef] [Green Version]
- Okugawa, S.; Moayeri, M.; Pomerantsev, A.P.; Sastalla, I.; Crown, D.; Gupta, P.K.; Leppla, S.H. Lipoprotein biosynthesis by prolipoprotein diacylglyceryl transferase is required for efficient spore germination and full virulence of Bacillus anthracis. Mol. Microbiol. 2012, 83, 96–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Davis, A.; Korza, G.; Zhang, P.; Li, Y.Q.; Setlow, B.; Setlow, P.; Hao, B. Role of a SpoVA protein in dipicolinic acid uptake into developing spores of Bacillus subtilis. J. Bacteriol. 2012, 194, 1875–1884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trunk, T.; Khalil, H.S.; Leo, J.C. Bacterial autoaggregation. AIMS Microbiol. 2018, 4, 140–164. [Google Scholar] [CrossRef]
- Wang, L.; Qin, W.; Yang, S.; Zhai, R.; Zhou, L.; Sun, C.; Pan, F.; Ji, Q.; Wang, Y.; Gu, J.; et al. The Adh adhesin domain is required for trimeric autotransporter Apa1-mediated Actinobacillus pleuropneumoniae adhesion, autoaggregation, biofilm formation and pathogenicity. Vet. Microbiol. 2015, 177, 175–183. [Google Scholar] [CrossRef]
- Yonezawa, H.; Osaki, T.; Kurata, S.; Zaman, C.; Hanawa, T.; Kamiya, S. Assessment of in vitro biofilm formation by Helicobacter pylori. J. Gastroenterol. Hepatol. 2010, 25 (Suppl. 1), S90–S94. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Sato, K.; Yukitake, H.; Noiri, Y.; Ebisu, S.; Nakayama, K. A Porphyromonas gingivalis mutant defective in a putative glycosyltransferase exhibits defective biosynthesis of the polysaccharide portions of lipopolysaccharide, decreased gingipain activities, strong autoaggregation, and increased biofilm formation. Infect. Immun. 2010, 78, 3801–3812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziebuhr, W.; Heilmann, C.; Götz, F.; Meyer, P.; Wilms, K.; Straube, E.; Hacker, J. Detection of the intercellular adhesion gene cluster (ica) and phase variation in Staphylococcus epidermidis blood culture strains and mucosal isolates. Infect. Immun. 1997, 65, 890–896. [Google Scholar] [CrossRef] [Green Version]
- Lo, C.Y.; Li, S.; Tan, D.; Pan, M.H.; Sang, S.; Ho, C.T. Trapping reactions of reactive carbonyl species with tea polyphenols in simulated physiological conditions. Mol. Nutr. Food Res. 2006, 50, 1118–1128. [Google Scholar] [CrossRef]
- Gray, D.A.; Dugar, G.; Gamba, P.; Strahl, H.; Jonker, M.J.; Hamoen, L.W. Extreme slow growth as alternative strategy to survive deep starvation in bacteria. Nat. Commun. 2019, 10, 890. [Google Scholar] [CrossRef] [Green Version]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Galperin, M.Y.; Mekhedov, S.L.; Puigbo, P.; Smirnov, S.; Wolf, Y.I.; Rigden, D.J. Genomic determinants of sporulation in Bacilli and Clostridia: Towards the minimal set of sporulation-specific genes. Environ. Microbiol. 2012, 14, 2870–2890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alambra, J.R.; Alenton, R.R.R.; Gulpeo, P.C.R.; Mecenas, C.L.; Miranda, A.P.; Thomas, R.C.; Velando, M.K.S.; Vitug, L.D.; Maningas, M.B.B. Immunomodulatory effects of turmeric, Curcuma longa (Magnoliophyta, Zingiberaceae) on Macrobrachium rosenbergii (Crustacea, Palaemonidae) against Vibrio alginolyticus (Proteobacteria, Vibrionaceae). Aquac. Aquar. Conserv. Legis. Int. J. Bioflux Soc. 2012, 5, 13–17. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacteria | TFDG (µg/mL) | CFU/mL (Mean ± SD) | Log Reduction (Mean ± SD) | % Inhibition (Mean ± SD) | ||
|---|---|---|---|---|---|---|
| K. aerogenes | 0 | (9.86 ± 0.09) × 108 | 0 | 0 | ||
| Gram-negative | 62.5 | (4.40 ± 0.12) × 108 | 0.35 ± 0.02 | 55.37 ± 1.59% | ||
| 250 | (6.07 ± 0.48) × 107 | 1.17 ± 0.03 | 93.12 ± 0.46% | |||
| E. coli | 0 | (1.09 ± 0.06) × 109 | 0 | 0 | ||
| 62.5 | (6.20 ± 0.42) × 108 | 0.25 ± 0.01 | 43.20 ± 0.83% | |||
| 250 | (2.36 ± 0.66) × 107 | 1.69 ± 0.12 | 97.87 ± 0.52% | |||
| P. aeruginosa | 0 | (7.01 ± 0.07) × 108 | 0 | 0 | ||
| 62.5 | (3.68 ± 0.11) × 108 | 0.28 ± 0.01 | 47.50 ± 1.15% | |||
| 250 | (1.57 ± 0.60) × 105 | 3.69 ± 0.21 | 99.98 ± 0.01% | |||
| P. mirabilis | 0 | (6.27 ± 0.08) × 109 | 0 | 0 | ||
| 62.5 | (3.17 ± 0.05) × 109 | 0.30 ± 0.00 | 49.39 ± 0.23% | |||
| 250 | (1.57 ± 0.39) × 107 | 2.62 ± 0.13 | 99.75 ± 0.07% | |||
| Gram-positive | Spore former | B. cereus | 0 | (6.54 ± 0.13) × 108 | 0 | 0 |
| 62.5 | (3.32 ± 0.08) × 108 | 0.29 ± 0.01 | 49. 16 ± 1.65% | |||
| 250 | (6.00 ± 1.63) × 105 | 3.05 ± 0.13 | 99.91 ± 0.03% | |||
| B. subtilis | 0 | (6.89 ± 0.09) × 108 | 0 | 0 | ||
| 62.5 | (3.35 ± 0.08) × 108 | 0.31 ± 0.01 | 51.40 ± 0.72% | |||
| 250 | (3.33 ± 0.82) × 105 | 3.34 ± 0.13 | 99.95 ± 0.01% | |||
| Non-spore former | S. aureus | 0 | (4.69 ± 0.12) × 109 | 0 | 0 | |
| 62.5 | (2.10 ± 0.18) × 109 | 0.35 ± 0.04 | 55.20 ± 4.62% | |||
| 250 | (3.03 ± 1.23) × 106 | 3.24 ± 0.24 | 99.93 ± 0.03% | |||
| S. pyogenes | 0 | (4.33 ± 0.02) × 109 | 0 | 0 | ||
| 62.5 | (2.24 ± 0.05) × 109 | 0.29 ± 0.01 | 48.28 ± 0.85% | |||
| 250 | (3.47 ± 0.09) × 106 | 3.10 ± 0.01 | 99.92 ± 0.05% | |||
| Acid-fast | M. smegmatis | 0 | (4.11 ± 0.07) × 109 | 0 | 0 | |
| 62.5 | (2.02 ± 0.05) × 109 | 0.31 ± 0.02 | 50.88 ± 1.92% | |||
| 250 | (1.33 ± 0.34) × 106 | 3.50 ± 0.11 | 99.97 ± 0.01% | |||
| Bacteria | TFDG (µg/mL) | RFU (Mean ± SD) | Log Reduction (Mean ± SD) | % Inhibition (Mean ± SD) | ||
|---|---|---|---|---|---|---|
| K. aerogenes | 0 | (1.90 ± 0.46) × 106 | 0 | 0 | ||
| Gram-negative | 62.5 | (9.17 ± 2.01) × 105 | 0.31 ± 0.01 | 51.52 ± 1.63% | ||
| 250 | (1.51 ± 0.73) × 104 | 2.14 ± 0.12 | 99.25 ± 0.22% | |||
| E. coli | 0 | (1.67 ± 0.09) × 106 | 0 | 0 | ||
| 62.5 | (7.89 ± 1.08) × 105 | 0.33 ± 0.04 | 53.00 ± 4.48% | |||
| 250 | (1.15 ± 0.19) × 104 | 2.17 ± 0.09 | 99.31 ± 0.14% | |||
| P. aeruginosa | 0 | (1.67 ± 0.36) × 106 | 0 | 0 | ||
| 62.5 | (8.07 ± 1.91) × 105 | 0.32 ± 0.02 | 51.97 ± 2.74% | |||
| 250 | (6.16 ± 2.84) × 104 | 1.48 ± 0.30 | 95.87 ± 2.39% | |||
| P. mirabilis | 0 | (1.52 ± 0.05) × 106 | 0 | 0 | ||
| 62.5 | (7.05 ± 0.36) × 105 | 0.34 ± 0.02 | 53.77 ± 1.69% | |||
| 250 | (3.52 ± 2.45) × 104 | 1.78 ± 0.37 | 97.71 ± 1.58% | |||
| Gram-positive | Spore former | B. cereus | 0 | (2.03 ± 1.00) × 106 | 0 | 0 |
| 62.5 | (7.58 ± 2.80) × 105 | 0.40 ± 0.07 | 59. 82 ± 6.19% | |||
| 250 | (1.28 ± 0.48) × 105 | 1.18 ± 0.08 | 93.24 ± 1.26% | |||
| B. subtilis | 0 | (2.30 ± 0.32) × 106 | 0 | 0 | ||
| 62.5 | (1.09 ± 0.21) × 105 | 0.33 ± 0.03 | 52.81 ± 3.47% | |||
| 250 | (4.82 ± 1.41) × 104 | 1.69 ± 0.07 | 97.94 ± 0.35% | |||
| Non-spore former | S. aureus | 0 | (7.74 ± 2.02) × 106 | 0 | 0 | |
| 62.5 | (3.66 ± 1.08) × 106 | 0.33 ± 0.03 | 53.12 ± 3.11% | |||
| 250 | (8.55 ± 3.73) × 104 | 1.98 ± 0.29 | 98.71 ± 0.76% | |||
| S. pyogenes | 0 | (4.43 ± 0.99) × 106 | 0 | 0 | ||
| 62.5 | (2.15 ± 0.42) × 106 | 0.31 ± 0.05 | 50.81 ± 5.81% | |||
| 250 | (1.01 ± 0.80) × 105 | 1.83 ± 0.50 | 97.72 ± 1.41% | |||
| Acid-fast | M. smegmatis | 0 | (1.83 ± 0.25) × 106 | 0 | 0 | |
| 62.5 | (9.09 ± 0.59) × 105 | 0.30 ± 0.04 | 49.82 ± 4.42% | |||
| 250 | (1.24 ± 0.42) × 104 | 2.19 ± 0.12 | 99.33 ± 0.16% | |||
| Bacteria | TFDG (µg/mL) | CFU/mL (Mean ± SD) | Log Reduction (Mean ± SD) | % Inhibition (Mean ± SD) | ||
|---|---|---|---|---|---|---|
| Gram-positive | Spore former | B. cereus | 0 | (1.11 ± 0.85) × 1010 | 0 | 0 |
| 312.5 | (4.79 ± 3.55) × 109 | 0.34 ± 0.03 | 54. 13 ± 3.51% | |||
| 625 | (6.55 ± 4.57) × 105 | 2.22 ± 0.12 | 99.37 ± 0.17% | |||
| B. subtilis | 0 | (7.24 ± 4.90) × 109 | 0 | 0 | ||
| 312.5 | (3.25 ± 2.33) × 109 | 0.41 ± 0.09 | 60.49 ± 7.91% | |||
| 625 | (7.75 ± 5.54) × 106 | 3.40 ± 0.64 | 99.92 ± 0.05% | |||
| Bacteria | Protein | Pocket Area (Å2) | Volume (Å3) | Residue Lining Pocket |
|---|---|---|---|---|
| B. cereus | GPR | 616.45 | 290.23 | Arg196, Ser197, Ile198, Thr252, Ile253, Asp254, Phe255, Ile256, Leu257, Lys258, Phe260, Gly261, Arg262, Met264, Lys265, Ala304, Gly306, Glu309, Leu316, Leu319, Val320, Leu321, Ser322, Val330 |
| Lgt (GerF) | 1284.21 | 1087.71 | Trp3, Ile4, Val5, Arg6, Gln8, Pro9, Ser11, Leu12, Ile13, Gly15, Ser16, Gly19, Met23, Val41, Ala44, Phe45, 1le48, Ala49, Trp52, Lys53, Ile75, Lys79, His80, Val82, Cys85, Ile86, Ser89, Val90. Ile92, Leu107, Leu110, Pro111, Ile112, Ala113, Leu114, Cys115, Met116, Ser117, Ile118, Ile119, Phe120, Tyr 121, Glu154, Ala158, Leu159, Val162, Gly163, Leu165, Trp166, Ile178, Phe181, Leu182, Glu185, Gly186, His189, Phe203, Gly204, Met207, Gln208, Leu211, Ser212, Cys214, Val215, Leu218 | |
| B. subtilis | GPR | 433.40 | 225.27 | Arg44, His46, Lys50, Thr53, Asp55, Val56, Thr57, Glu59, Leu74, Ala76, Gln77, Gly78, Val90, Val93, Phe94, Glu96, Glu97, Ser99, Ala100, Phe101, Glu103, Asn104, Lys109, Ile206, Ile208, His355, Lys357, Val58, Ser359, Gln360, Asn362 Lys363, Gly364, Ser365, Tyr366,Asn367 |
| Lgt (GerF) | 1565.20 | 1484.55 | Leu18, Ala 19, His21, Tyr23, Gly24, Ile26, Ile27, Gly30, Ala31, Gly34, Ile37, Ala38, Arg40, Glu41, Lys44, Arg45, Gly46, Leu47, Phe52, Val56, Ala59, Ile60, Ala63, Ile64, Ala67, Ile89, Trp90, Gly93, Ile94, Gly99, Leu100, Ala103, Ile104, Thr106, Gly107, Leu116, Phe118, Lys120, Leu121, Ala122, Asp123, Ile124, Ala125, Ala126, Pro127, Ser128, Ile129, Leu 131, Gly132, Gln 133, Ile135, Gly136, Arg137, Gly139, Asn140, Glu145, Phe180, Glu183, Ser187, Ile192, Leu196, Arg198, Arg199, Ala200, Asn201, Leu202, Arg203, Arg204, Glu206, Met207, Phe208, Tyr211, Ile212, Tyr215, Arg219, Arg226, Thr233, Asp234, Ser235, Leu236 |
| Bacteria | Protein | Docking Score (kcal/mol) | Interacting Residue | Distance | Category |
|---|---|---|---|---|---|
| B. cereus | GPR | −9.7 | Trp120 | 2.51 | Conventional H-bond |
| Trp120 | 3.98, 3.88 | Pi-Pi Stacked | |||
| Asn121 | 2.59 | Conventional H-bond | |||
| Val163 | 2.78 | Conventional H-bond | |||
| Glu340 | 2.37 | Conventional H-bond | |||
| Lgt (GerF) | −7.6 | Lys26 | 1.85 | Conventional H-bond | |
| Lys26 | 3.73 | Pi-Cation | |||
| Ser31 | 2.99 | Conventional H-bond | |||
| Lys168 | 2.00 | Conventional H-bond | |||
| Arg171 | 2.51 | Conventional H-bond | |||
| B. subtilis | GPR | −10.3 | Gly119 | 2.06, 2.94 | Conventional H-bond |
| ASN120 | 2.34 | Conventional H-bond | |||
| ASN120 | 2.92 | Conventional H-bond | |||
| ASN120 | 2.03 | Conventional H-bond | |||
| ASN120 | 1.97 | Conventional H-bond | |||
| ASN122 | 1.68 | Unfavorable Donor-Donor | |||
| ASP126 | 2.97 | Conventional H-bond | |||
| Ile170 | 1.93 | Conventional H-bond | |||
| Thr172 | 3.05 | Pi Donor H-bond | |||
| Lys222 | 1.22 | Unfavorable Donor-Donor | |||
| Asp336 | 2.49 | Conventional H-bond | |||
| Asp336 | 2.23 | Conventional H-bond | |||
| Lgt (GerF) | −10.3 | Ala63 | 3.78 | Hydrophobic Pi-Sigma | |
| Ala103 | 3.62 | Hydrophobic Pi-Sigma | |||
| Ala103 | 2.07 | Conventional H-Bond | |||
| Ile129 | 3.29 | Carbon Hydrogen Bond | |||
| Gly132 | 3.96 | Hydrophobic Pi-Sigma | |||
| Tyr215 | 1.92 | Conventional H-bond | |||
| Asp234 | 2.38 | Conventional H-bond |
| Bacteria | Gene | NCBI Gene ID | Crystal Structure | PDB Reference | % Sequence Identity |
|---|---|---|---|---|---|
| B. cereus | gpr | 56320307 | None | IC8B | 71.47% |
| lgt (gerF) | 56322894 | None | 5AZC | 16.74% | |
| B. subtilis | gpr | 937838 | None | IC8B | 68.68% |
| lgt (gerF) | 12085459 | None | 5AZC | 35.21% |
| Bacteria | Protein | Center X | Center Y | Center Z | Size X | Size Y | Size Z |
|---|---|---|---|---|---|---|---|
| B. cereus | GPR | −2 | 10 | 110 | 126 | 126 | 126 |
| Lgt (GerF) | −20 | 0 | 0 | 92 | 120 | 104 | |
| B. subtilis | GPR | −20 | 40 | 50 | 126 | 126 | 126 |
| Lgt (GerF) | −15 | 0 | −8 | 90 | 114 | 126 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yussof, A.; Cammalleri, B.; Fayemiwo, O.; Lopez, S.; Chu, T. Antibacterial and Sporicidal Activity Evaluation of Theaflavin-3,3′-digallate. Int. J. Mol. Sci. 2022, 23, 2153. https://doi.org/10.3390/ijms23042153
Yussof A, Cammalleri B, Fayemiwo O, Lopez S, Chu T. Antibacterial and Sporicidal Activity Evaluation of Theaflavin-3,3′-digallate. International Journal of Molecular Sciences. 2022; 23(4):2153. https://doi.org/10.3390/ijms23042153
Chicago/Turabian StyleYussof, Ayuni, Brian Cammalleri, Oluwanifemi Fayemiwo, Sabrina Lopez, and Tinchun Chu. 2022. "Antibacterial and Sporicidal Activity Evaluation of Theaflavin-3,3′-digallate" International Journal of Molecular Sciences 23, no. 4: 2153. https://doi.org/10.3390/ijms23042153
APA StyleYussof, A., Cammalleri, B., Fayemiwo, O., Lopez, S., & Chu, T. (2022). Antibacterial and Sporicidal Activity Evaluation of Theaflavin-3,3′-digallate. International Journal of Molecular Sciences, 23(4), 2153. https://doi.org/10.3390/ijms23042153