
Remodeling of Lipid A in Pseudomonas syringae pv. phaseolicola In Vitro

 , , , , and
, , , , and
Abstract
1. Introduction
2. Results
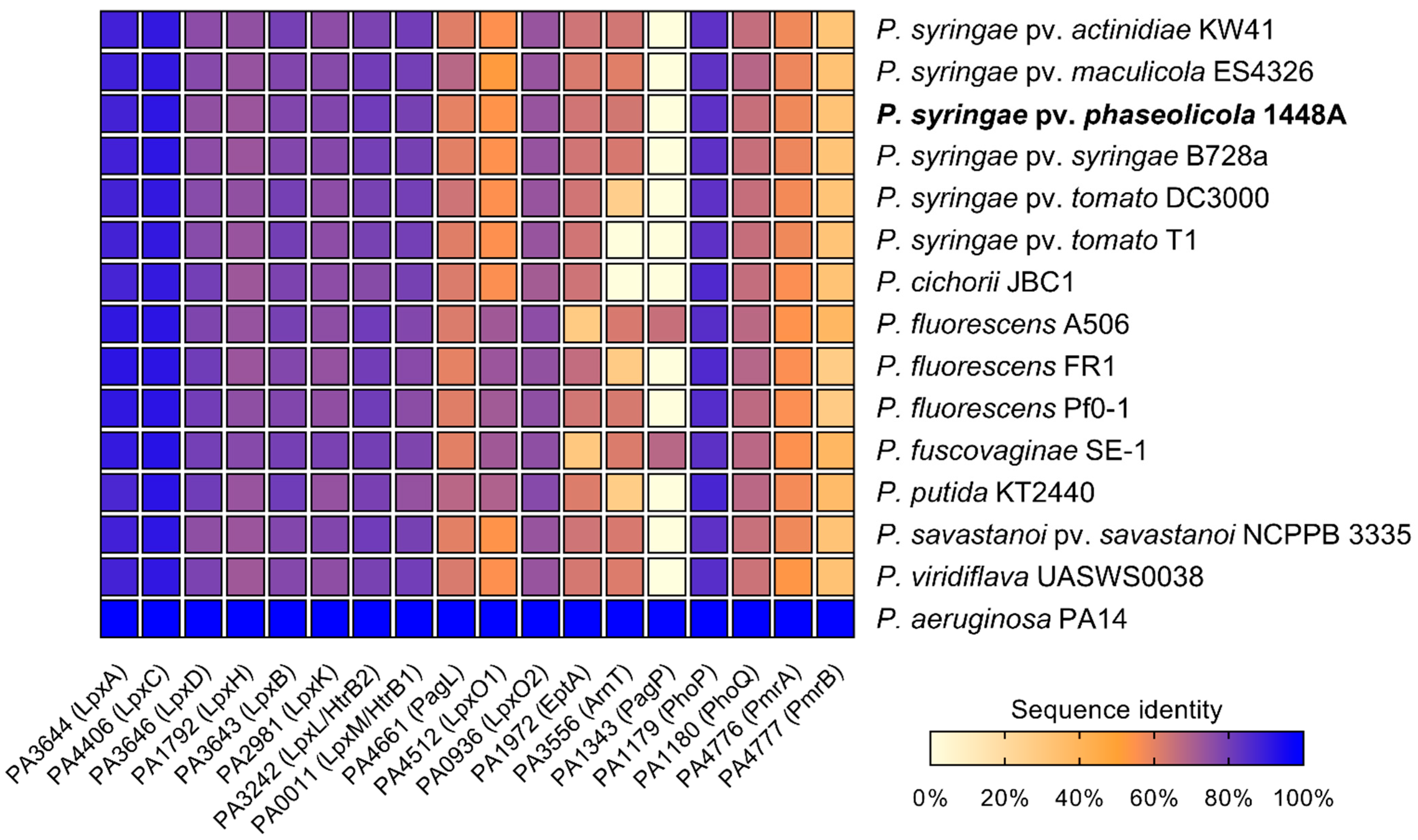
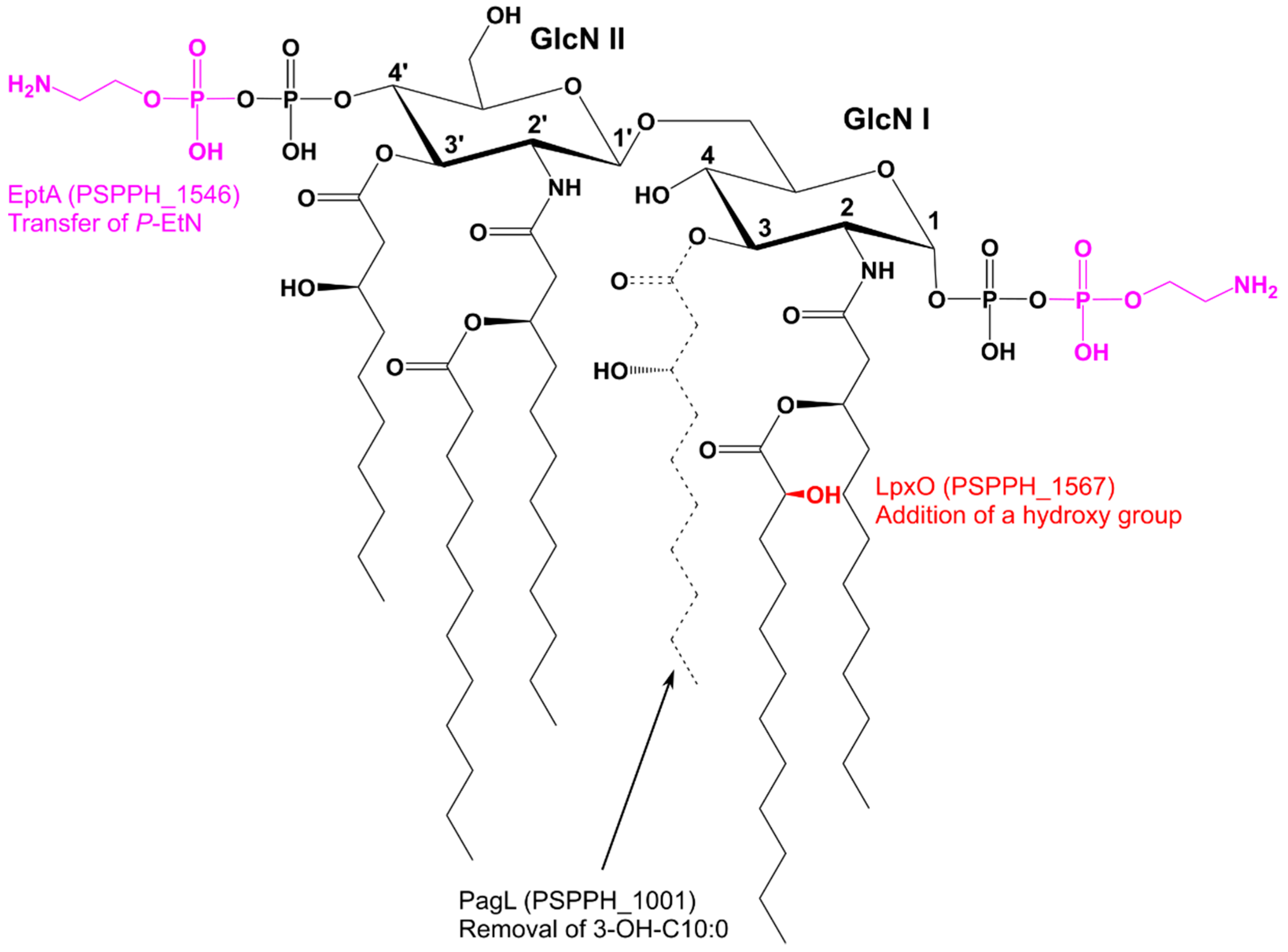
2.1. Orthologues of Lipid A-Modifying Genes Can Be Found in Pseudomonas spp.
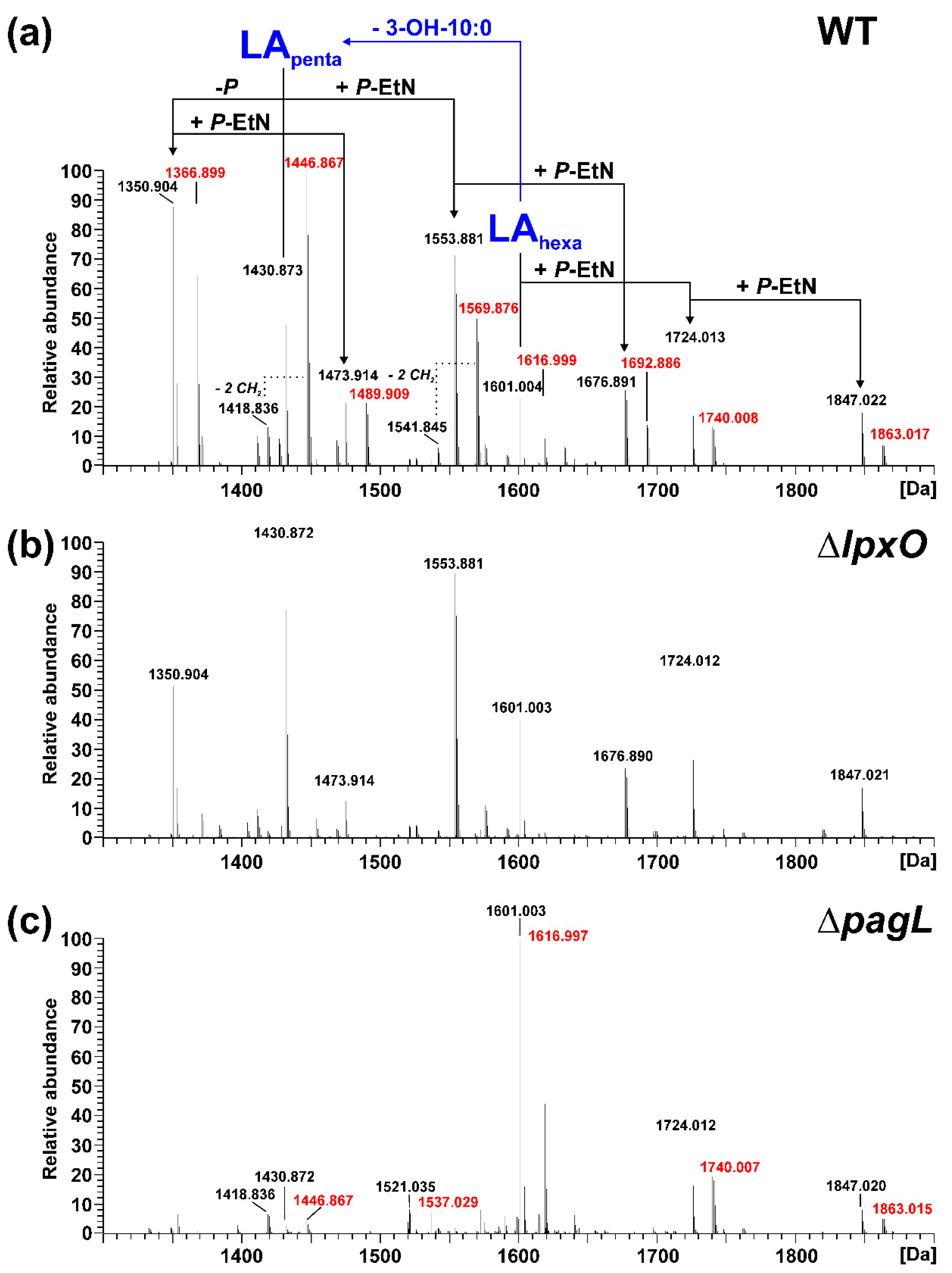
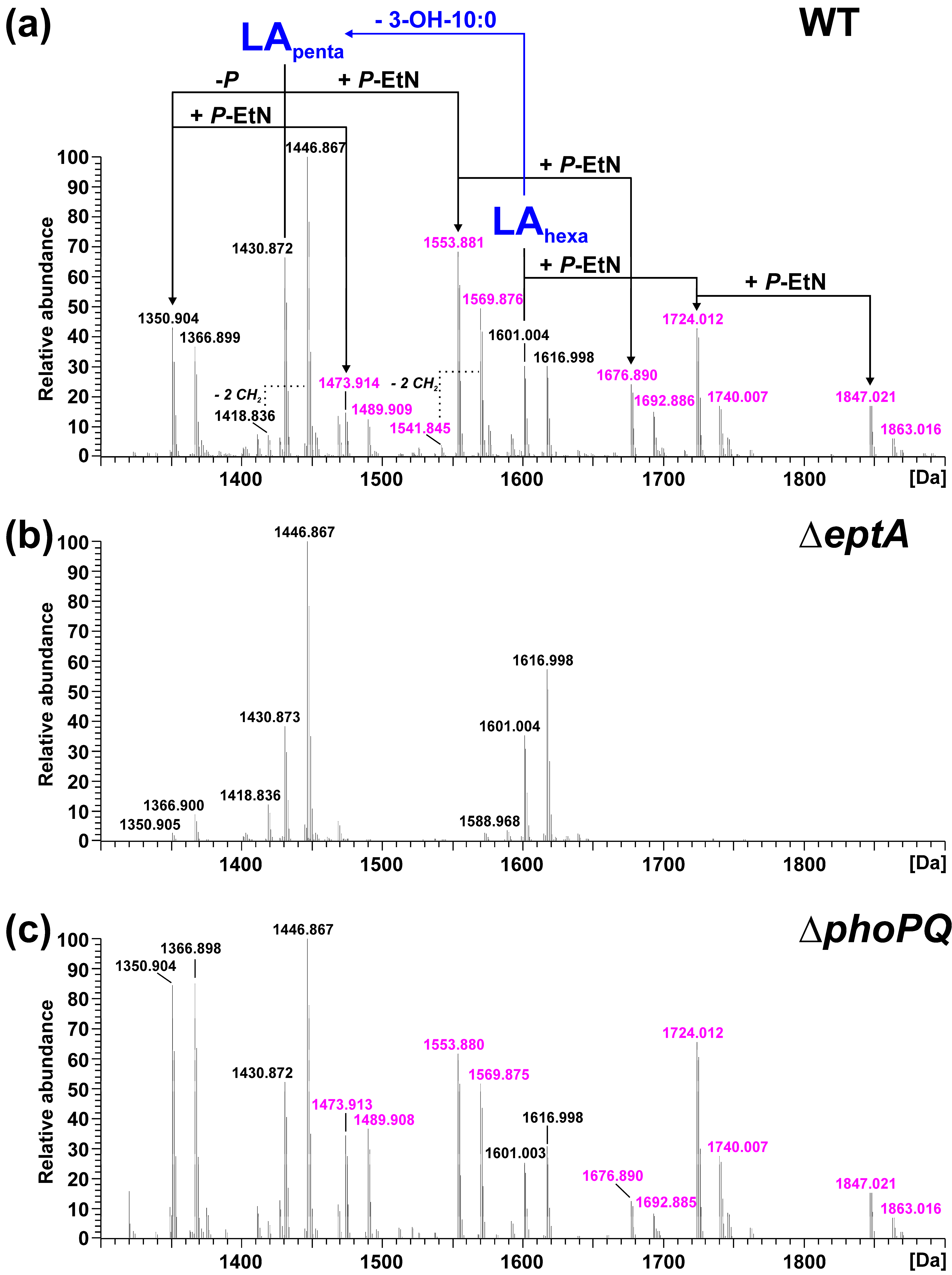
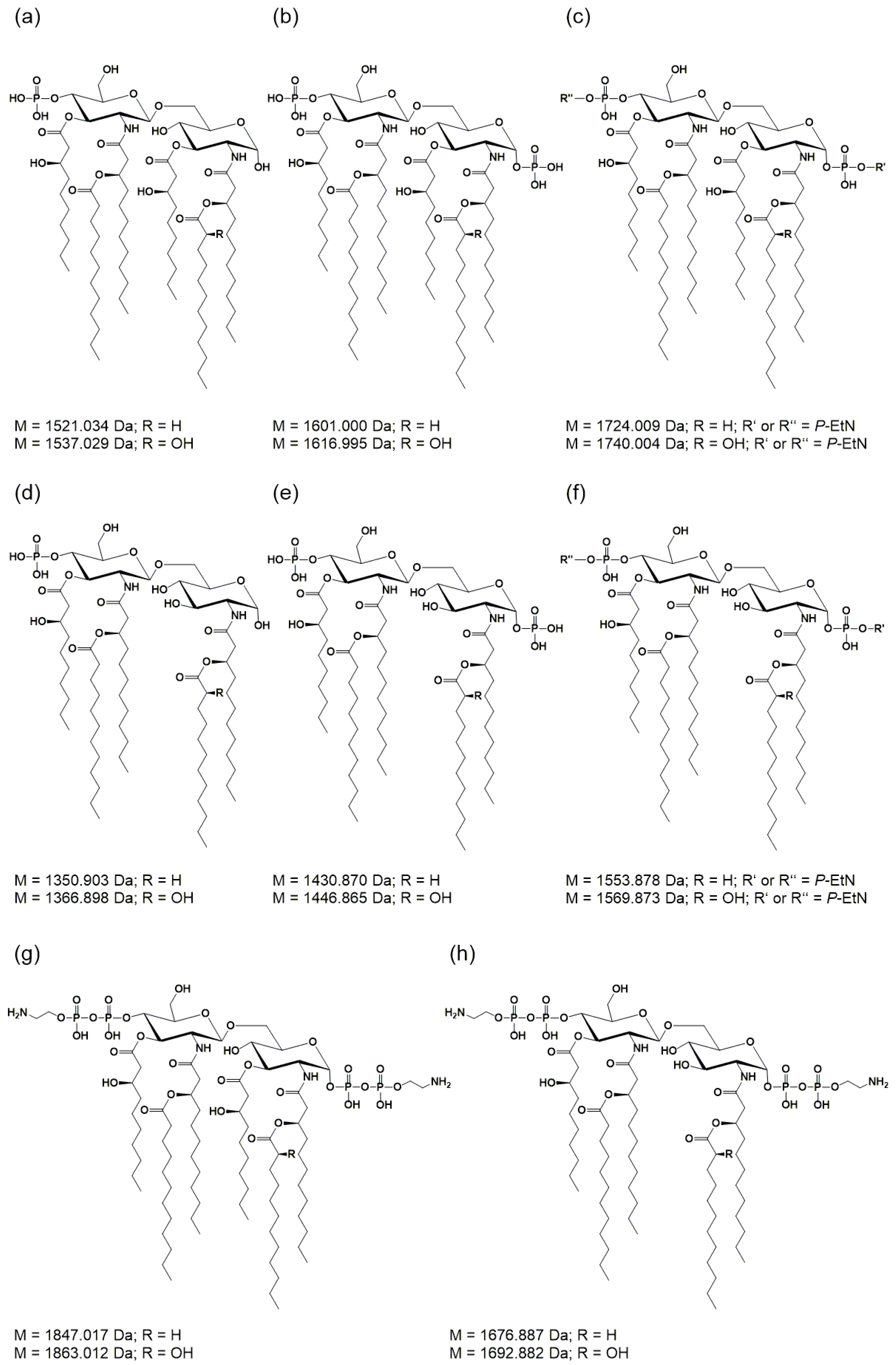
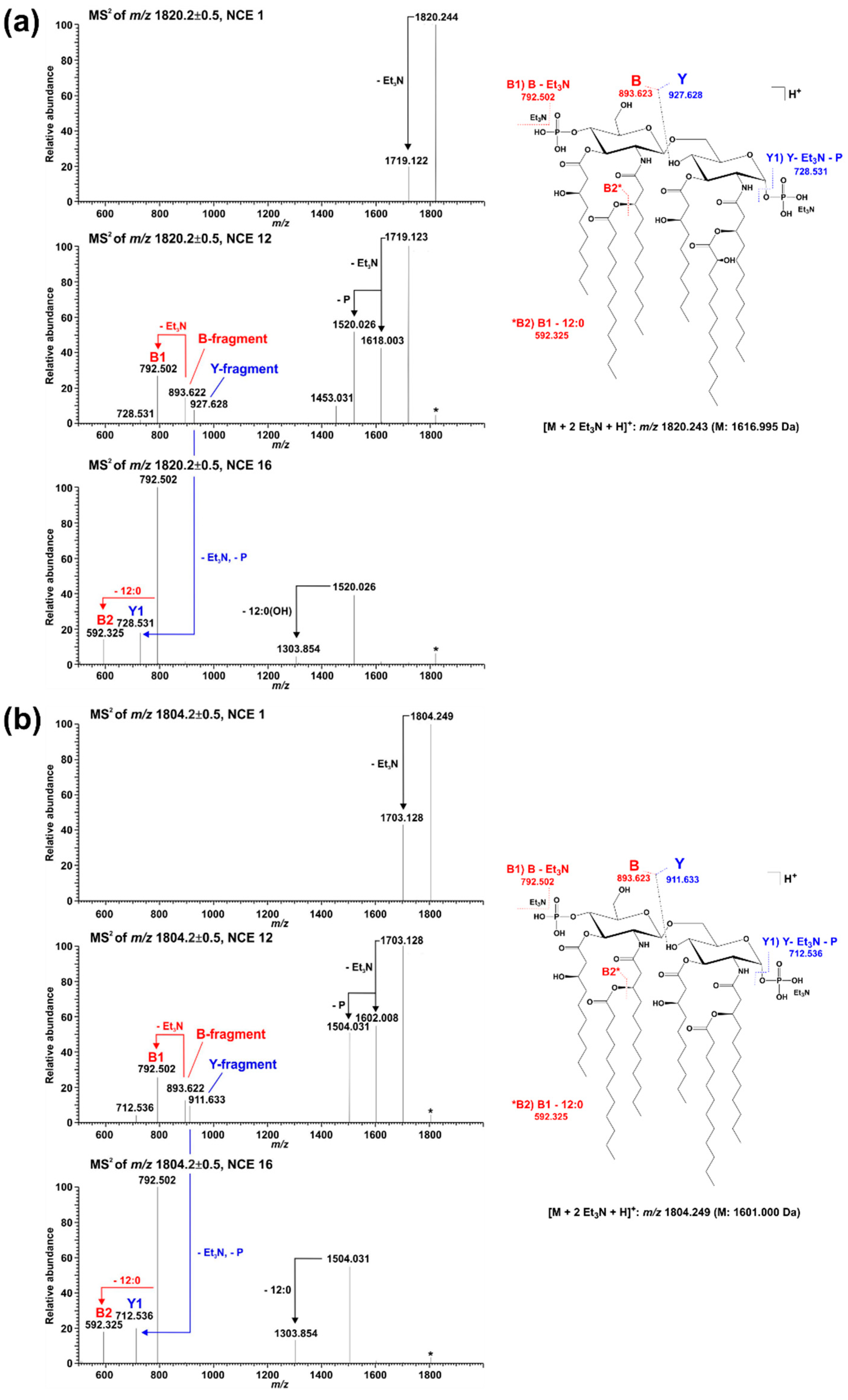
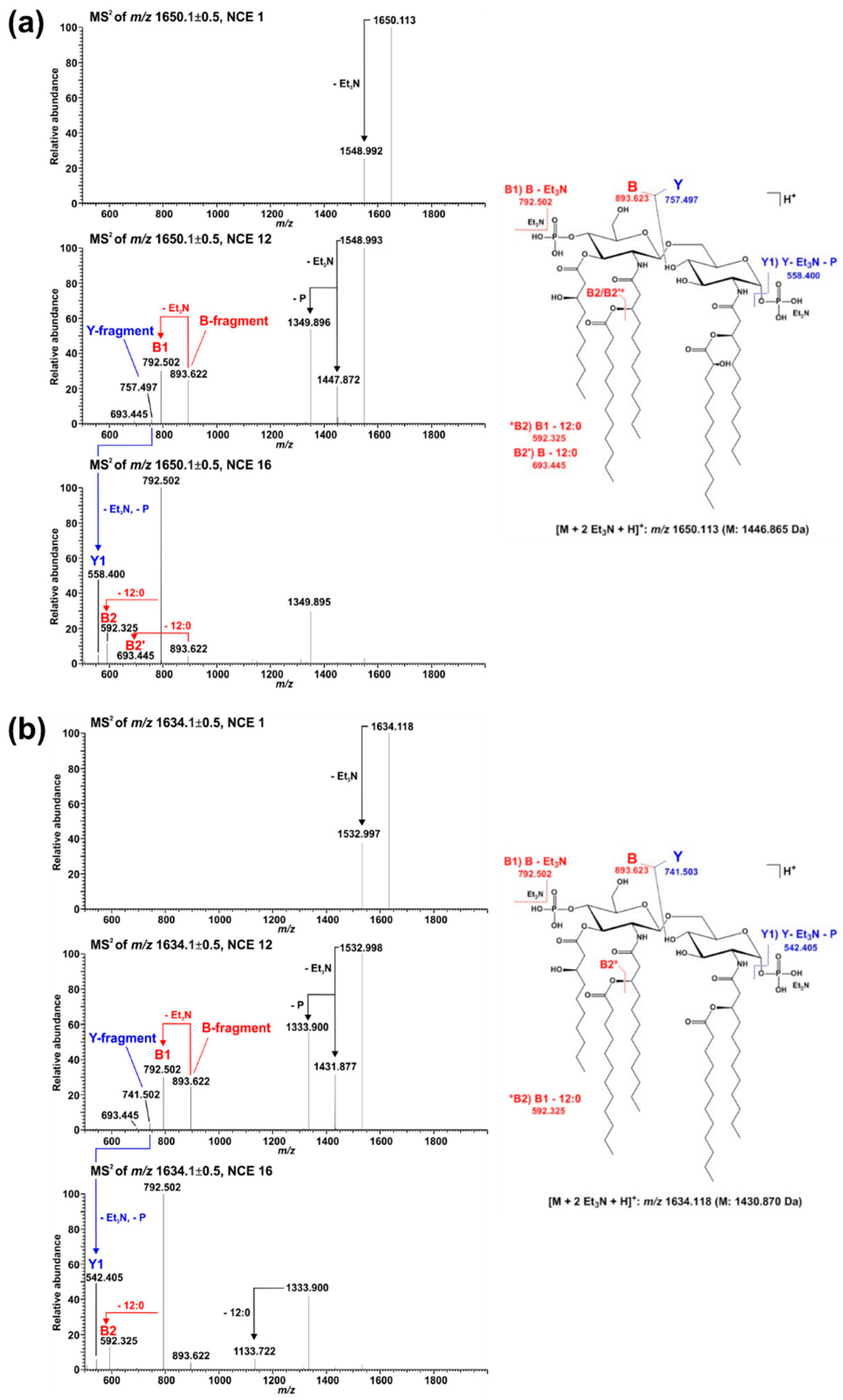
2.2. Mass Spectrometric Analysis of Pph 1448A Lipid A Reveals the Activity of PagL, EptA, and LpxO
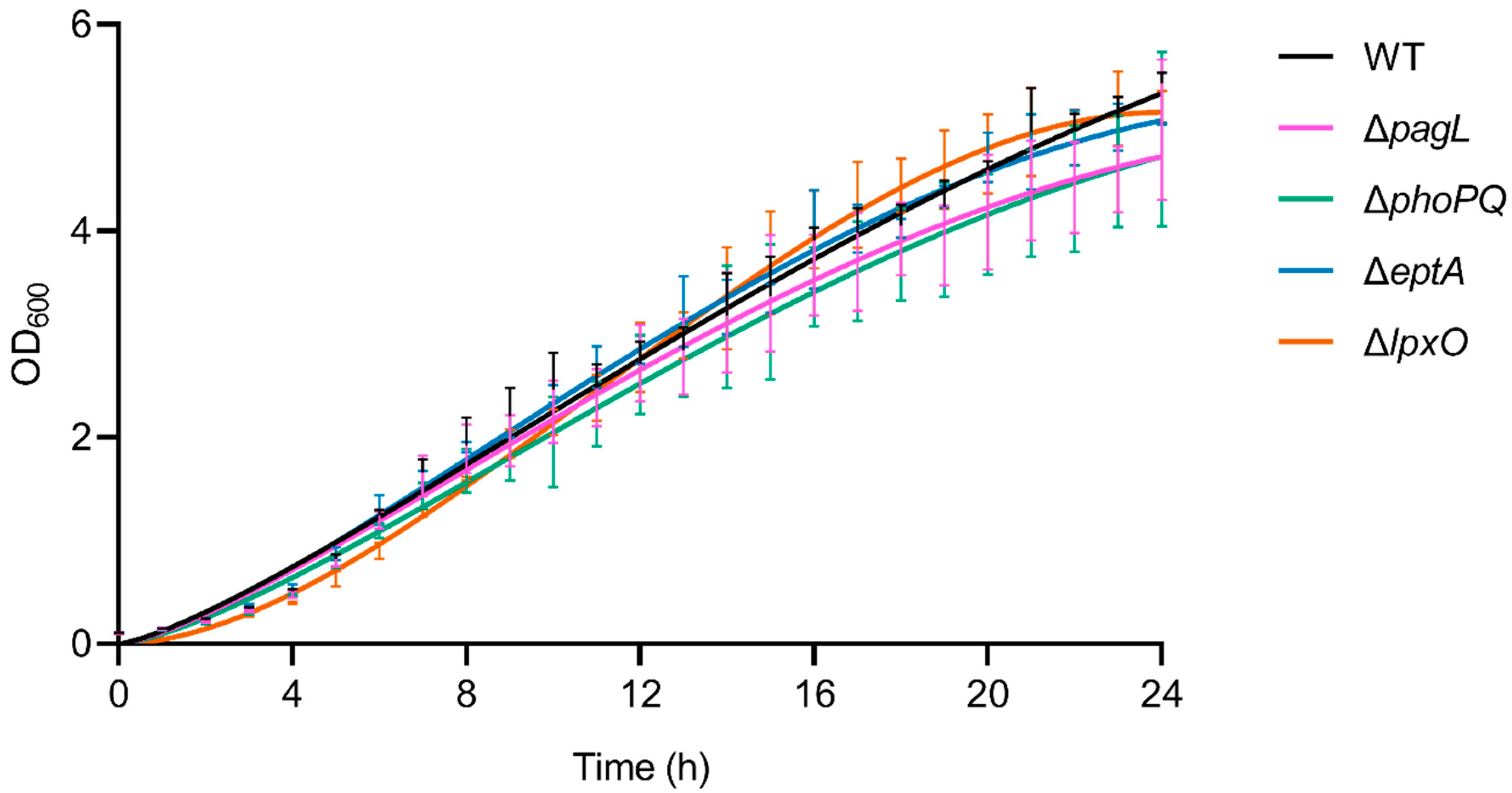
2.3. Lack of Lipid A-Modifying Enzymes Does Not Affect Growth and LPS Formation in Pph 1448A
3. Discussion
4. Materials and Methods
4.1. Strains and Growth Conditions
4.2. Analysis of Predicted Proteomes of Pseudomonas spp.
4.3. Gene Knockout in Pph 1448A
4.4. Preparation of Lipopolysaccharide from Pph 1448A Strains
4.5. Lipid A Preparation
4.6. Mass Spectrometric Analysis of Lipid A via ESI-MS
4.7. Analysis of Fatty Acids via GC-MS
4.8. Crude LPS Extraction
4.9. Mass Spectrometric Analysis of Crude Lipid A Extracts via MALDI-TOF
4.10. SDS-PAGE and Silver Staining of Bacterial LPS
4.11. Bacterial Growth Curve
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xin, X.F.; Kvitko, B.; He, S.Y. Pseudomonas syringae: What it takes to be a pathogen. Nat. Rev. Microbiol. 2018, 16, 316–328. [Google Scholar] [CrossRef] [PubMed]
- Taylor, J.D.; Teverson, D.M.; Allen, D.J.; Pastor-Corrales, M.A. Identification and origin of races of Pseudomonas syringae pv. phaseolicola from Africa and other bean growing areas. Plant Pathol. 1996, 45, 469–478. [Google Scholar] [CrossRef]
- Abdullah, A.S.; Douglas, C. Halo blight of mungbean in Australia. Crops 2021, 1, 3–7. [Google Scholar] [CrossRef]
- Sun, S.; Zhi, Y.; Zhu, Z.; Jin, J.; Duan, C.; Wu, X.; Wang, X. An emerging disease caused by Pseudomonas syringae pv. phaseolicola threatens mung bean production in China. Plant Dis. 2017, 101, 95–102. [Google Scholar] [CrossRef]
- Kabanov, D.S.; Prokhorenko, I.R. Structural analysis of lipopolysaccharides from Gram-negative bacteria. Biochemistry 2010, 75, 383–404. [Google Scholar] [CrossRef]
- Fahy, E.; Subramaniam, S.; Brown, H.A.; Glass, C.K.; Merrill, A.H.; Murphy, R.C.; Raetz, C.R.H.; Russell, D.W.; Seyama, Y.; Shaw, W.; et al. A comprehensive classification system for lipids. J. Lipid Res. 2005, 46, 839–861. [Google Scholar] [CrossRef]
- Raetz, C.R.H.; Whitfield, C. Lipopolysaccharide endotoxins. Annu. Rev. Biochem. 2002, 71, 635–700. [Google Scholar] [CrossRef]
- Carpenter, T.S.; Parkin, J.; Khalid, S. The free energy of small solute permeation through the Escherichia coli outer membrane has a distinctly asymmetric profile. J. Phys. Chem. Lett. 2016, 7, 3446–3451. [Google Scholar] [CrossRef]
- Whitfield, C.; Trent, M.S. Biosynthesis and export of bacterial lipopolysaccharides. Annu. Rev. Biochem. 2014, 83, 99–128. [Google Scholar] [CrossRef]
- Opiyo, S.O.; Pardy, R.L.; Moriyama, H.; Moriyama, E.N. Evolution of the Kdo2-lipid A biosynthesis in bacteria. BMC Evol. Biol. 2010, 10, 362. [Google Scholar] [CrossRef]
- King, J.D.; Kocíncová, D.; Westman, E.L.; Lam, J.S. Lipopolysaccharide biosynthesis in Pseudomonas aeruginosa. Innate Immun. 2009, 15, 261–312. [Google Scholar] [CrossRef] [PubMed]
- Clementz, T.; Bednarski, J.J.; Raetz, C.R.H. Function of the htrB high temperature requirement gene of Escherichia coli in the acylation of lipid A: HtrB catalyzed incorporation of laurate. J. Biol. Chem. 1996, 271, 12095–12102. [Google Scholar] [CrossRef]
- Clementz, T.; Zhou, Z.; Raetz, C.R.H. Function of the Escherichia coli msbB gene, a multicopy suppressor of htrB knockouts, in the acylation of lipid A. Acylation by MsbB follows laurate incorporation by HtrB. J. Biol. Chem. 1997, 272, 10353–10360. [Google Scholar] [CrossRef] [PubMed]
- Lo Sciuto, A.; Cervoni, M.; Stefanelli, R.; Spinnato, M.C.; Di Giamberardino, A.; Mancone, C.; Imperi, F. Genetic basis and physiological effects of lipid A hydroxylation in Pseudomonas aeruginosa PAO1. Pathogens 2019, 8, 291. [Google Scholar] [CrossRef] [PubMed]
- Kutschera, A.; Dawid, C.; Gisch, N.; Schmid, C.; Raasch, L.; Gerster, T.; Schäffer, M.; Smakowska-Luzan, E.; Belkhadir, Y.; Corina Vlot, A.; et al. Bacterial medium-chain 3-hydroxy fatty acid metabolites trigger immunity in Arabidopsis plants. Science 2019, 364, 178–181. [Google Scholar] [CrossRef] [PubMed]
- Raetz, C.R.H.; Reynolds, C.M.; Trent, M.S.; Bishop, R.E. Lipid A modification systems in Gram-negative bacteria. Annu. Rev. Biochem. 2007, 76, 295–329. [Google Scholar] [CrossRef]
- Anandan, A.; Vrielink, A. Structure and function of lipid A–modifying enzymes. Ann. N. Y. Acad. Sci. 2020, 1459, 19–37. [Google Scholar] [CrossRef]
- Ernst, R.K.; Adams, K.N.; Moskowitz, S.M.; Kraig, G.M.; Kawasaki, K.; Stead, C.M.; Trent, M.S.; Miller, S.I. The Pseudomonas aeruginosa lipid A deacylase: Selection for expression and loss within the cystic fibrosis airway. J. Bacteriol. 2006, 188, 191–201. [Google Scholar] [CrossRef]
- Thaipisuttikul, I.; Hittle, L.E.; Chandra, R.; Zangari, D.; Dixon, C.L.; Garrett, T.A.; Rasko, D.A.; Dasgupta, N.; Moskowitz, S.M.; Malmström, L.; et al. A divergent Pseudomonas aeruginosa palmitoyltransferase essential for cystic fibrosis-specific lipid A. Mol. Microbiol. 2014, 91, 158–174. [Google Scholar] [CrossRef]
- Bhat, R.; Marx, A.; Galanos, C.; Conrad, R.S. Structural studies of lipid A from Pseudomonas aeruginosa PAO1: Occurrence of 4-amino-4-deoxyarabinose. J. Bacteriol. 1990, 172, 6631–6636. [Google Scholar] [CrossRef]
- Trent, M.S. Biosynthesis, transport, and modification of lipid A. Biochem. Cell Biol. 2004, 82, 71–86. [Google Scholar] [CrossRef]
- Gunn, J.S. The Salmonella PmrAB regulon: Lipopolysaccharide modifications, antimicrobial peptide resistance and more. Trends Microbiol. 2008, 16, 284–290. [Google Scholar] [CrossRef]
- Trent, M.S.; Ribeiro, A.A.; Doerrler, W.T.; Lin, S.; Cotter, R.J.; Raetz, C.R.H. Accumulation of a polyisoprene-linked amino sugar in polymyxin-resistant Salmonella typhimurium and Escherichia coli: Structural characterization and transfer to lipid A in the periplasm. J. Biol. Chem. 2001, 276, 43132–43144. [Google Scholar] [CrossRef] [PubMed]
- Harper, M.; Wright, A.; Michael, F.S.; Li, J.; Lucas, D.D.; Ford, M.; Adler, B.; Cox, A.D.; Boyce, J.D. Characterization of two novel lipopolysaccharide phosphoethanolamine transferases in Pasteurella multocida and their role in resistance to cathelicidin-2. Infect. Immun. 2017, 85, e00557-17. [Google Scholar] [CrossRef] [PubMed]
- Wright, J.C.; Hood, D.W.; Randle, G.A.; Makepeace, K.; Cox, A.D.; Li, J.; Chalmers, R.; Richards, J.C.; Moxon, E.R. lpt6, a gene required for addition of phosphoethanolamine to inner-core lipopolysaccharide of Neisseria meningitidis and Haemophilus influenzae. J. Bacteriol. 2004, 186, 6970–6982. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Cox, A.D.; Wright, J.C.; Li, J.; Hood, D.W.; Moxon, E.R.; Richards, J.C. Phosphorylation of the lipid A region of meningococcal lipopolysaccharide: Identification of a family of transferases that add phosphoethanolamine to lipopolysaccharide. J. Bacteriol. 2003, 185, 3270–3277. [Google Scholar] [CrossRef] [PubMed]
- Cullen, T.W.; Madsen, J.A.; Ivanov, P.L.; Brodbelt, J.S.; Trent, M.S. Characterization of unique modification of flagellar rod protein FlgG by Campylobacter jejuni lipid A phosphoethanolamine transferase, linking bacterial locomotion and antimicrobial peptide resistance. J. Biol. Chem. 2012, 287, 3326–3336. [Google Scholar] [CrossRef] [PubMed]
- Klein, G.; Lindner, B.; Brade, H.; Raina, S. Molecular basis of lipopolysaccharide heterogeneity in Escherichia coli: Envelope stress-responsive regulators control the incorporation of glycoforms with a third 3-deoxy-α-D-manno-oct-2-ulosonic acid and rhamnose. J. Biol. Chem. 2011, 286, 42787–42807. [Google Scholar] [CrossRef]
- Klein, G.; Müller-Loennies, S.; Lindner, B.; Kobylak, N.; Brade, H.; Raina, S. Molecular and structural basis of inner core lipopolysaccharide alterations in Escherichia coli: Incorporation of glucuronic acid and phosphoethanolamine in the heptose region. J. Biol. Chem. 2013, 288, 8111–8127. [Google Scholar] [CrossRef]
- Groisman, E.A. Feedback control of two-component regulatory systems. Annu. Rev. Microbiol. 2016, 70, 103–124. [Google Scholar] [CrossRef]
- Zhou, Z.; Ribeiro, A.A.; Lin, S.; Cotter, R.J.; Miller, S.I.; Raetz, C.R.H. Lipid A modifications in polymyxin-resistant Salmonella typhimurium: PmrA-dependent 4-amino-4-deoxy-L-arabinose, and phosphoethanolamine incorporation. J. Biol. Chem. 2001, 276, 43111–43121. [Google Scholar] [CrossRef] [PubMed]
- Nowicki, E.M.; O’Brien, J.P.; Brodbelt, J.S.; Trent, M.S. Extracellular zinc induces phosphoethanolamine addition to Pseudomonas aeruginosa lipid A via the ColRS two-component system. Mol. Microbiol. 2015, 97, 166–178. [Google Scholar] [CrossRef] [PubMed]
- Sinha, A.; Nyongesa, S.; Viau, C.; Gruenheid, S.; Veyrier, F.J.; Le Moual, H. PmrC (EptA) and CptA negatively affect outer membrane vesicle production in Citrobacter rodentium. J. Bacteriol. 2019, 201, e00454-18. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Lim, K.B.; Poduje, C.M.; Daniel, M.; Gunn, J.S.; Hackett, M.; Miller, S.I. Lipid A acylation and bacterial resistance against vertebrate antimicrobial peptides. Cell 1998, 95, 189–198. [Google Scholar] [CrossRef]
- Bishop, R.E.; Gibbons, H.S.; Guina, T.; Trent, M.S.; Miller, S.I.; Raetz, C.R.H. Transfer of palmitate from phospholipids to lipid A in outer membranes of Gram-negative bacteria. EMBO J. 2000, 19, 5071–5080. [Google Scholar] [CrossRef]
- Simpson, B.W.; Trent, M.S. Pushing the envelope: LPS modifications and their consequences. Nat. Rev. Microbiol. 2019, 17, 403–416. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, K.; China, K.; Nishijima, M. Release of the lipopolysaccharide deacylase PagL from latency compensates for a lack of lipopolysaccharide aminoarabinose modification-dependent resistance to the antimicrobial peptide polymyxin B in Salmonella enterica. J. Bacteriol. 2007, 189, 4911–4919. [Google Scholar] [CrossRef] [PubMed]
- Elhenawy, W.; Bording-Jorgensen, M.; Valguarnera, E.; Haurat, M.F.; Wine, E.; Feldman, M.F. LPS remodeling triggers formation of outer membrane vesicles in Salmonella. MBio 2016, 7, e00940-16. [Google Scholar] [CrossRef]
- Bartholomew, T.L.; Kidd, T.J.; Pessoa, J.S.; Álvarez, R.C.; Bengoechea, J.A. 2-hydroxylation of Acinetobacter baumannii lipid A contributes to virulence. Infect. Immun. 2019, 87, e00066-19. [Google Scholar] [CrossRef]
- Groisman, E.A. The pleiotropic two-component regulatory system PhoP-PhoQ. J. Bacteriol. 2001, 183, 1835–1842. [Google Scholar] [CrossRef]
- García Véscovi, E.; Soncini, F.C.; Groisman, E.A. Mg2+ as an extracellular signal: Environmental regulation of Salmonella virulence. Cell 1996, 84, 165–174. [Google Scholar] [CrossRef]
- Bader, M.W.; Navarre, W.W.; Shiau, W.; Nikaido, H.; Frye, J.G.; McClelland, M.; Fang, F.C.; Miller, S.I. Regulation of Salmonella typhimurium virulence gene expression by cationic antimicrobial peptides. Mol. Microbiol. 2003, 50, 219–230. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.; Groisman, E.A.; Shin, D. Activated by different signals, the PhoP/PhoQ two-component system differentially regulates metal uptake. J. Bacteriol. 2009, 191, 7174–7181. [Google Scholar] [CrossRef] [PubMed]
- Prost, L.R.; Miller, S.I. The Salmonellae PhoQ sensor: Mechanisms of detection of phagosome signals. Cell. Microbiol. 2008, 10, 576–582. [Google Scholar] [CrossRef] [PubMed]
- Pierce, B.K.; Kirkpatrick, B.C. The PhoP/Q two-component regulatory system is essential for Xylella fastidiosa survival in Vitis vinifera grapevines. Physiol. Mol. Plant Pathol. 2015, 89, 55–61. [Google Scholar] [CrossRef]
- Needham, B.D.; Trent, M.S. Fortifying the barrier: The impact of lipid A remodelling on bacterial pathogenesis. Nat. Rev. Microbiol. 2013, 11, 467–481. [Google Scholar] [CrossRef]
- Zdorovenko, G.M.; Veremeichenko, S.N. Comparative characterization of the lipopolysaccharides of different Pseudomonas fluorescens biovar I strains. Microbiology 2001, 70, 441–450. [Google Scholar] [CrossRef]
- Ernst, R.K.; Hajjar, A.M.; Tsai, J.H.; Moskowitz, S.M.; Wilson, C.B.; Miller, S.I. Pseudomonas aeruginosa lipid A diversity and its recognition by Toll-like receptor 4. J. Endotoxin Res. 2003, 9, 395–400. [Google Scholar] [CrossRef]
- El Hamidi, A.; Tirsoaga, A.; Novikov, A.; Hussein, A.; Caroff, M. Microextraction of bacterial lipid A: Easy and rapid method for mass spectrometric characterization. J. Lipid Res. 2005, 46, 1773–1778. [Google Scholar] [CrossRef]
- Moskowitz, S.M.; Ernst, R.K.; Miller, S.I. PmrAB, a two-component regulatory system of Pseudomonas aeruginosa that modulates resistance to cationic antimicrobial peptides and addition of aminoarabinose to lipid A. J. Bacteriol. 2004, 186, 575–579. [Google Scholar] [CrossRef]
- Kawasaki, K.; Ernst, R.K.; Miller, S.I. Deacylation and palmitoylation of lipid A by Salmonellae outer membrane enzymes modulate host signaling through Toll-like receptor 4. J. Endotoxin Res. 2004, 10, 439–444. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, K.; Ernst, R.K.; Miller, S.I. 3-O-Deacylation of Lipid A by PagL, a PhoP/PhoQ-regulated deacylase of Salmonella typhimurium, modulates signaling through Toll-like Receptor 4. J. Biol. Chem. 2004, 279, 20044–20048. [Google Scholar] [CrossRef] [PubMed]
- Ranf, S.; Gisch, N.; Schäffer, M.; Illig, T.; Westphal, L.; Knirel, Y.A.; Sánchez-Carballo, P.M.; Zähringer, U.; Hückelhoven, R.; Lee, J.; et al. A lectin S-domain receptor kinase mediates lipopolysaccharide sensing in Arabidopsis thaliana. Nat. Immunol. 2015, 16, 426–433. [Google Scholar] [CrossRef] [PubMed]
- Samantha, A.; Vrielink, A. Lipid A phosphoethanolamine transferase: Regulation, structure and immune response. J. Mol. Biol. 2020, 432, 5184–5196. [Google Scholar] [CrossRef]
- Nawrot, R.; Barylski, J.; Nowicki, G.; Broniarczyk, J.; Buchwald, W.; Goździcka-Józefiak, A. Plant antimicrobial peptides. Folia Microbiol. 2014, 59, 181–196. [Google Scholar] [CrossRef]
- Jack, H.W.; Tzi, B.N. Sesquin, a potent defensin-like antimicrobial peptide from ground beans with inhibitory activities toward tumor cells and HIV-1 reverse transcriptase. Peptides 2005, 26, 1120–1126. [Google Scholar] [CrossRef]
- Rybak, K.; Robatzek, S. Functions of extracellular vesicles in immunity and virulence. Plant Physiol. 2019, 179, 1236–1247. [Google Scholar] [CrossRef]
- McMillan, H.M.; Zebell, S.G.; Ristaino, J.B.; Dong, X.; Kuehn, M.J. Protective plant immune responses are elicited by bacterial outer membrane vesicles. Cell Rep. 2021, 34, 108645. [Google Scholar] [CrossRef]
- Tran, A.X.; Whittimore, J.D.; Wyrick, P.B.; McGrath, S.C.; Cotter, R.J.; Trent, M.S. The lipid A 1-phosphatase of Helicobacter pylori is required for resistance to the antimicrobial peptide polymyxin. J. Bacteriol. 2006, 188, 4531–4541. [Google Scholar] [CrossRef]
- Cullen, T.W.; Giles, D.K.; Wolf, L.N.; Ecobichon, C.; Boneca, I.G.; Trent, M.S. Helicobacter pylori versus the host: Remodeling of the bacterial outer membrane is required for survival in the gastric mucosa. PLoS Pathog. 2011, 7, e1002454. [Google Scholar] [CrossRef]
- Joardar, V.; Lindeberg, M.; Jackson, R.W.; Selengut, J.; Dodson, R.; Brinkac, L.M.; Daugherty, S.C.; DeBoy, R.; Durkin, A.S.; Giglio, M.G.; et al. Whole-genome sequence analysis of Pseudomonas syringae pv. phaseolicola 1448A reveals divergence among pathovars in genes involved in virulence and transposition. J. Bacteriol. 2005, 187, 6488–6498. [Google Scholar] [CrossRef] [PubMed]
- King, E.O.; Ward, M.K.; Raney, D.E. Two simple media for the demonstration of pyocyanin and fluorescin. J. Lab. Clin. Med. 1954, 44, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Bertani, G. Studies on lysogenesis. I. The mode of phage liberation by lysogenic Escherichia coli. J. Bacteriol. 1951, 62, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Kutschera, A.; Schombel, U.; Wröbel, M.; Gisch, N.; Ranf, S. Loss of wbpL disrupts O-polysaccharide synthesis and impairs virulence of plant-associated Pseudomonas strains. Mol. Plant Pathol. 2019, 20, 1535–1549. [Google Scholar] [CrossRef]
- Hanahan, D. Studies on transformation of Escherichia coli with plasmids. J. Mol. Biol. 1983, 166, 557–580. [Google Scholar] [CrossRef]
- Kvitko, B.H.; Collmer, A. Construction of Pseudomonas syringae pv. tomato DC3000 mutant and polymutant strains. Methods Mol. Biol. 2011, 712, 109–128. [Google Scholar] [CrossRef]
- Boyer, H.W.; Roulland-Dussoix, D. A complementation analysis of the restriction and modification of DNA in Escherichia coli. J. Mol. Biol. 1969, 41, 459–472. [Google Scholar] [CrossRef]
- Westphal, O.; Jann, K. Extraction with phenol-water and further applications of the procedure. Methods Carbohydr. Chem. 1965, 5, 83–91. [Google Scholar]
- Domon, B.; Costello, C.E. A systematic nomenclature for carbohydrate fragmentations in FAB-MS/MS spectra of glycoconjugates. Glycoconj. J. 1988, 5, 397–409. [Google Scholar] [CrossRef]
- Knirel, Y.A.; Bystrova, O.V.; Kocharova, N.A.; Zähringer, U.; Pier, G.B. Conserved and variable structural features in the lipopolysaccharide of Pseudomonas aeruginosa. J. Endotoxin Res. 2006, 12, 324–336. [Google Scholar] [CrossRef]
- Hitchcock, P.J.; Brown, T.M. Morphological heterogeneity among Salmonella lipopolysaccharide chemotypes in silver-stained polyacrylamide gels. J. Bacteriol. 1983, 154, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.Y.; Chandler, C.E.; Leung, L.M.; McElheny, C.L.; Mettus, R.T.; Shanks, R.M.Q.; Liu, J.H.; Goodlett, D.R.; Ernst, R.K.; Doi, Y. Structural modification of lipopolysaccharide conferred by mcr-1 in Gram-negative ESKAPE pathogens. Antimicrob. Agents Chemother. 2017, 61, e00580-17. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Calculated Mono-Isotopic Mass | Substituting Acyl or Acyloxy-Acyl Chains/Additional Residues on Lipid A Backbone | |||||
|---|---|---|---|---|---|---|
| [Da] | C-4′ | C-3′ | C-2′ | C-3 | C-2 | C-1 |
| 1322.872 | P | 10:0((R)-3-OH) | 12:0((R)-3-O(12:0)) # | H | 10:0((R)-3-O(12:0)) # | |
| 1338.867 | P | 10:0((R)-3-OH) | 12:0((R)-3-O(12:0)) # | H | 10:0((R)-3-O(12:0((S)-2-OH))) # | |
| 1350.903 | P | 10:0((R)-3-OH) | 12:0((R)-3-O(12:0)) | H | 12:0((R)-3-O(12:0)) | |
| 1366.898 | P | 10:0((R)-3-OH) | 12:0((R)-3-O(12:0)) | H | 12:0((R)-3-O(12:0((S)-2-OH))) | |
| 1402.838 | P | 10:0((R)-3-OH) | 12:0((R)-3-O(12:0)) # | H | 10:0((R)-3-O(12:0)) # | P |
| 1418.833 § | P | 10:0((R)-3-OH) | 12:0((R)-3-O(12:0)) # | H | 10:0((R)-3-O(12:0((S)-2-OH))) # | P |
| 1418.833 § | P | 10:0((R)-3-OH) | 12:0((R)-3-OH) ## | 10:0((R)-3-OH) | 12:0((R)-3-O(12:0)) ## | P |
| 1430.870 | P | 10:0((R)-3-OH) | 12:0((R)-3-O(12:0)) | H | 12:0((R)-3-O(12:0)) | P |
| 1446.865 | P | 10:0((R)-3-OH) | 12:0((R)-3-O(12:0)) | H | 12:0((R)-3-O(12:0((S)-2-OH))) | P |
| 1473.912 | PP-EtN | 10:0((R)-3-OH) | 12:0((R)-3-O(12:0)) | H | 12:0((R)-3-O(12:0)) | |
| 1489.907 | PP-EtN | 10:0((R)-3-OH) | 12:0((R)-3-O(12:0)) | H | 12:0((R)-3-O(12:0((S)-2-OH))) | |
| 1521.034 | P | 10:0((R)-3-OH) | 12:0((R)-3-O(12:0)) | 10:0((R)-3-OH) | 12:0((R)-3-O(12:0)) | |
| 1537.029 | P | 10:0((R)-3-OH) | 12:0((R)-3-O(12:0)) | 10:0((R)-3-OH) | 12:0((R)-3-O(12:0((S)-2-OH))) | |
| 1541.842 §§ | P * | 10:0((R)-3-OH) | 12:0((R)-3-O(12:0)) # | H | 10:0((R)-3-O(12:0((S)-2-OH))) # | PP-EtN * |
| 1541.842 §§ | P * | 10:0((R)-3-OH) | 12:0((R)-3-OH) ## | 10:0((R)-3-OH) | 12:0((R)-3-O(12:0)) ## | PP-EtN * |
| 1553.878 | P * | 10:0((R)-3-OH) | 12:0((R)-3-O(12:0)) | H | 12:0((R)-3-O(12:0)) | PP-EtN * |
| 1569.873 | P * | 10:0((R)-3-OH) | 12:0((R)-3-O(12:0)) | H | 12:0((R)-3-O(12:0((S)-2-OH))) | PP-EtN * |
| 1572.969 | P | 10:0((R)-3-OH) | 12:0((R)-3-O(12:0)) # | 10:0((R)-3-OH) | 10:0((R)-3-O(12:0)) # | P |
| 1588.964 | P | 10:0((R)-3-OH) | 12:0((R)-3-O(12:0)) # | 10:0((R)-3-OH) | 10:0((R)-3-O(12:0((S)-2-OH))) # | P |
| 1601.000 | P | 10:0((R)-3-OH) | 12:0((R)-3-O(12:0)) | 10:0((R)-3-OH) | 12:0((R)-3-O(12:0)) | P |
| 1616.995 | P | 10:0((R)-3-OH) | 12:0((R)-3-O(12:0)) | 10:0((R)-3-OH) | 12:0((R)-3-O(12:0((S)-2-OH))) | P |
| 1676.887 | PP-EtN | 10:0((R)-3-OH) | 12:0((R)-3-O(12:0)) | H | 12:0((R)-3-O(12:0)) | PP-EtN |
| 1692.882 | PP-EtN | 10:0((R)-3-OH) | 12:0((R)-3-O(12:0)) | H | 12:0((R)-3-O(12:0((S)-2-OH))) | PP-EtN |
| 1724.009 | P * | 10:0((R)-3-OH) | 12:0((R)-3-O(12:0)) | 10:0((R)-3-OH) | 12:0((R)-3-O(12:0)) | PP-EtN * |
| 1740.004 | P * | 10:0((R)-3-OH) | 12:0((R)-3-O(12:0)) | 10:0((R)-3-OH) | 12:0((R)-3-O(12:0((S)-2-OH))) | PP-EtN * |
| 1847.017 | PP-EtN | 10:0((R)-3-OH) | 12:0((R)-3-O(12:0)) | 10:0((R)-3-OH) | 12:0((R)-3-O(12:0)) | PP-EtN |
| 1863.012 | PP-EtN | 10:0((R)-3-OH) | 12:0((R)-3-O(12:0)) | 10:0((R)-3-OH) | 12:0((R)-3-O(12:0((S)-2-OH))) | PP-EtN |
| Strain (Figure Number of Respective MS1 Spectrum) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| WT (2a) | ΔlpxO (2b) | ΔpagL (2c) | WT (3a) | ΔeptA (3b) | ΔphoPQ (3c) | |||||||
| Mcal | Mexp* | Error (Δppm) | Mexp* | Error (Δppm) | Mexp* | Error (Δppm) | Mexp* | Error (Δppm) | Mexp* | Error (Δppm) | Mexp* | Error (Δppm) |
| 1322.872 | 1322.872 | 0.0 | 1322.872 | 0.0 | n.d. | - | 1322.872 | 0.0 | n.d. | - | 1322.871 | −0.8 |
| 1338.867 | 1338.867 | 0.0 | n.d. | - | 1338.866 | −0.7 | 1338.868 | 0.7 | 1338.869 | 1.5 | 1338.866 | −0.7 |
| 1350.903 | 1350.904 | 0.7 | 1350.904 | 0.7 | n.d. | - | 1350.904 | 0.7 | 1350.905 | 1.5 | 1350.904 | 0.7 |
| 1366.898 | 1366.899 | 0.7 | n.d. | - | 1366.898 | 0.0 | 1366.899 | 0.7 | 1366.900 | 1.5 | 1,366.898 | 0.0 |
| 1402.838 | n.d. ** | - | 1402.841 | 2.1 | n.d. | - | 1402.842 | 2.9 | 1402.842 | 2.9 | n.d. | - |
| 1418.833 $ | 1418.836 | 2.1 | 1418.837 | 2.8 | 1418.836 | 2.1 | 1418.836 | 2.1 | 1,418.836 | 2.1 | 1418.836 | 2.1 |
| 1430.870 | 1430.873 | 2.1 | 1430.872 | 1.4 | 1430.872 | 1.4 | 1430.872 | 1.4 | 1,430.873 | 2.1 | 1430.872 | 1.4 |
| 1446.865 | 1446.867 | 1.4 | n.d. | - | 1446.867 | 1.4 | 1446.867 | 1.4 | 1,446.867 | 1.4 | 1446.867 | 1.4 |
| 1473.912 | 1473.914 | 1.4 | 1473.914 | 1.4 | n.d. | - | 1473.914 | 1.4 | n.d. | - | 1473.913 | 0.7 |
| 1489.907 | 1489.909 | 1.3 | n.d. | - | n.d. | - | 1489.909 | 1.3 | n.d. | - | 1489.908 | 0.7 |
| 1521.034 | 1521.034 | 0.0 | 1521.035 | 0.7 | 1521.035 | 0.7 | 1521.034 | 0.0 | n.d. | - | 1521.033 | −0.7 |
| 1537.029 | 1537.029 | 0.0 | n.d. | - | 1537.029 | 0.0 | 1537.030 | 0.7 | 1537.031 | 1.3 | 1537.029 | 0.0 |
| 1541.842 $ | 1541.845 | 2.0 | 1541.846 | 2.6 | 1541.845 | 2.0 | 1541.845 | 2.0 | n.d. | - | n.d. | - |
| 1553.878 | 1553.881 | 1.9 | 1553.881 | 1.9 | 1553.882 | 2.6 | 1553.881 | 1.9 | n.d. | - | 1553.880 | 1.3 |
| 1569.873 | 1569.876 | 1.9 | n.d. | - | 1569.877 | 2.5 | 1569.876 | 1.9 | n.d. | - | 1569.875 | 1.3 |
| 1572.969 | n.d. | - | n.d. | - | 1572.972 | 1.9 | 1572.973 | 2.5 | 1572.973 | 2.5 | n.d. | - |
| 1588.964 | n.d. | - | n.d. | - | 1588.967 | 1.9 | 1588.968 | 2.5 | 1588.968 | 2.5 | n.d. | - |
| 1601.000 | 1601.004 | 2.5 | 1601.003 | 1.9 | 1601.003 | 1.9 | 1601.004 | 2.5 | 1601.004 | 2.5 | 1601.003 | 1.9 |
| 1616.995 | 1616.999 | 2.5 | n.d. | - | 1616.997 | 1.2 | 1616.998 | 1.9 | 1616.998 | 1.9 | 1616.998 | 1.9 |
| 1676.887 | 1676.891 | 2.4 | 1676.890 | 1.8 | n.d. | - | 1676.890 | 1.8 | n.d. | - | 1676.890 | 1.8 |
| 1692.882 | 1692.886 | 2.4 | n.d. | - | n.d. | - | 1692.886 | 2.4 | n.d. | - | 1692.885 | 1.8 |
| 1724.009 | 1724.013 | 2.3 | 1724.012 | 1.7 | 1724.012 | 1.7 | 1724.012 | 1.7 | n.d. | - | 1724.012 | 1.7 |
| 1740.004 | 1740.008 | 2.3 | n.d. | - | 1740.007 | 1.7 | 1740.007 | 1.7 | n.d. | - | 1740.007 | 1.7 |
| 1847.017 | 1847.022 | 2.7 | 1847.021 | 2.2 | 1847.020 | 1.6 | 1847.021 | 2.2 | n.d. | - | 1847.021 | 2.2 |
| 1863.012 | 1863.017 | 2.7 | n.d. | - | 1863.015 | 1.6 | 1863.016 | 2.1 | n.d. | - | 1863.016 | 2.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gerster, T.; Wröbel, M.; Hofstaedter, C.E.; Schwudke, D.; Ernst, R.K.; Ranf, S.; Gisch, N. Remodeling of Lipid A in Pseudomonas syringae pv. phaseolicola In Vitro. Int. J. Mol. Sci. 2022, 23, 1996. https://doi.org/10.3390/ijms23041996
Gerster T, Wröbel M, Hofstaedter CE, Schwudke D, Ernst RK, Ranf S, Gisch N. Remodeling of Lipid A in Pseudomonas syringae pv. phaseolicola In Vitro. International Journal of Molecular Sciences. 2022; 23(4):1996. https://doi.org/10.3390/ijms23041996
Chicago/Turabian StyleGerster, Tim, Michelle Wröbel, Casey E. Hofstaedter, Dominik Schwudke, Robert K. Ernst, Stefanie Ranf, and Nicolas Gisch. 2022. "Remodeling of Lipid A in Pseudomonas syringae pv. phaseolicola In Vitro" International Journal of Molecular Sciences 23, no. 4: 1996. https://doi.org/10.3390/ijms23041996
APA StyleGerster, T., Wröbel, M., Hofstaedter, C. E., Schwudke, D., Ernst, R. K., Ranf, S., & Gisch, N. (2022). Remodeling of Lipid A in Pseudomonas syringae pv. phaseolicola In Vitro. International Journal of Molecular Sciences, 23(4), 1996. https://doi.org/10.3390/ijms23041996