
The Influence of Intravaginal Gestagens Treatment on the Morphological Features and Endometrial Steroid Hormone Receptors Content during Anestrus Type II in Dairy Cattle

 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
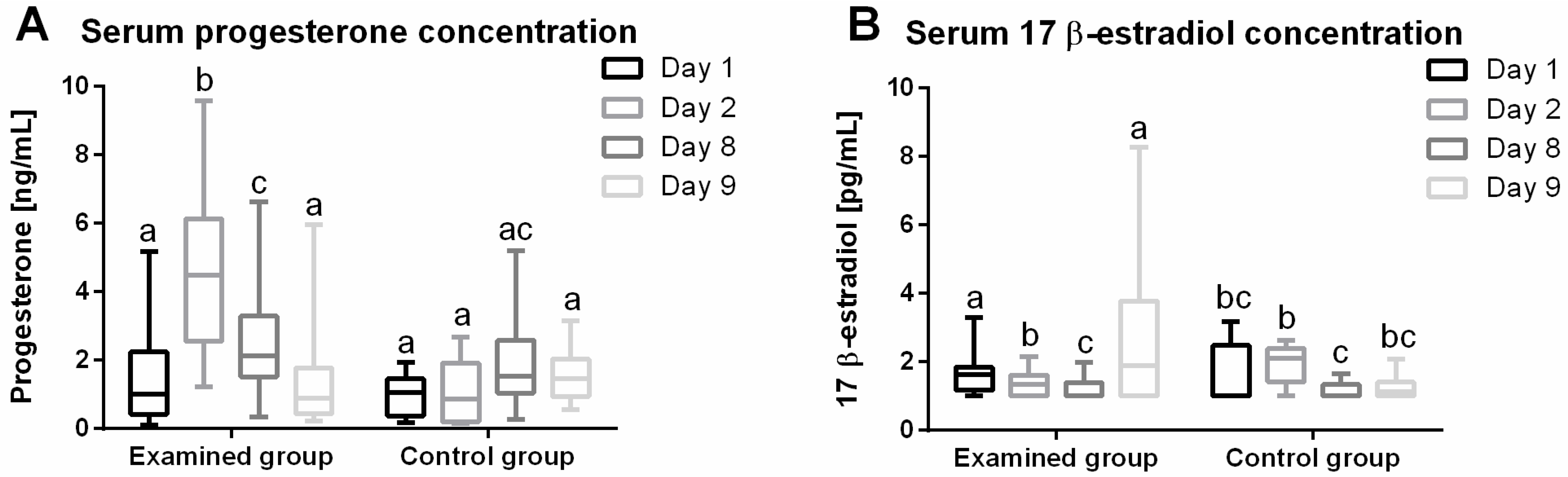
2.1. Progesterone (P4) Plasma Concentration
2.2. 17β-Estradiol (E2) Plasma Concentration
2.3. Progesterone Receptors (PR) mRNA Expression in Endometrium
2.4. Estrogen Receptors (ER) mRNA Expression in Endometrium
2.5. Progesterone Receptors (PR) Protein Expression in Endometrium
2.6. Estrogen Receptors (ER) Protein Expression in Endometrium
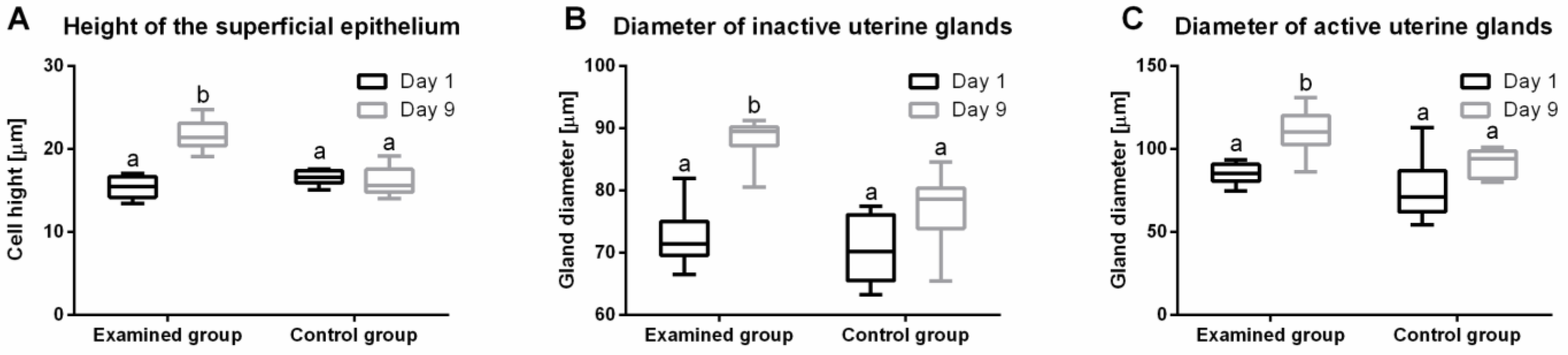
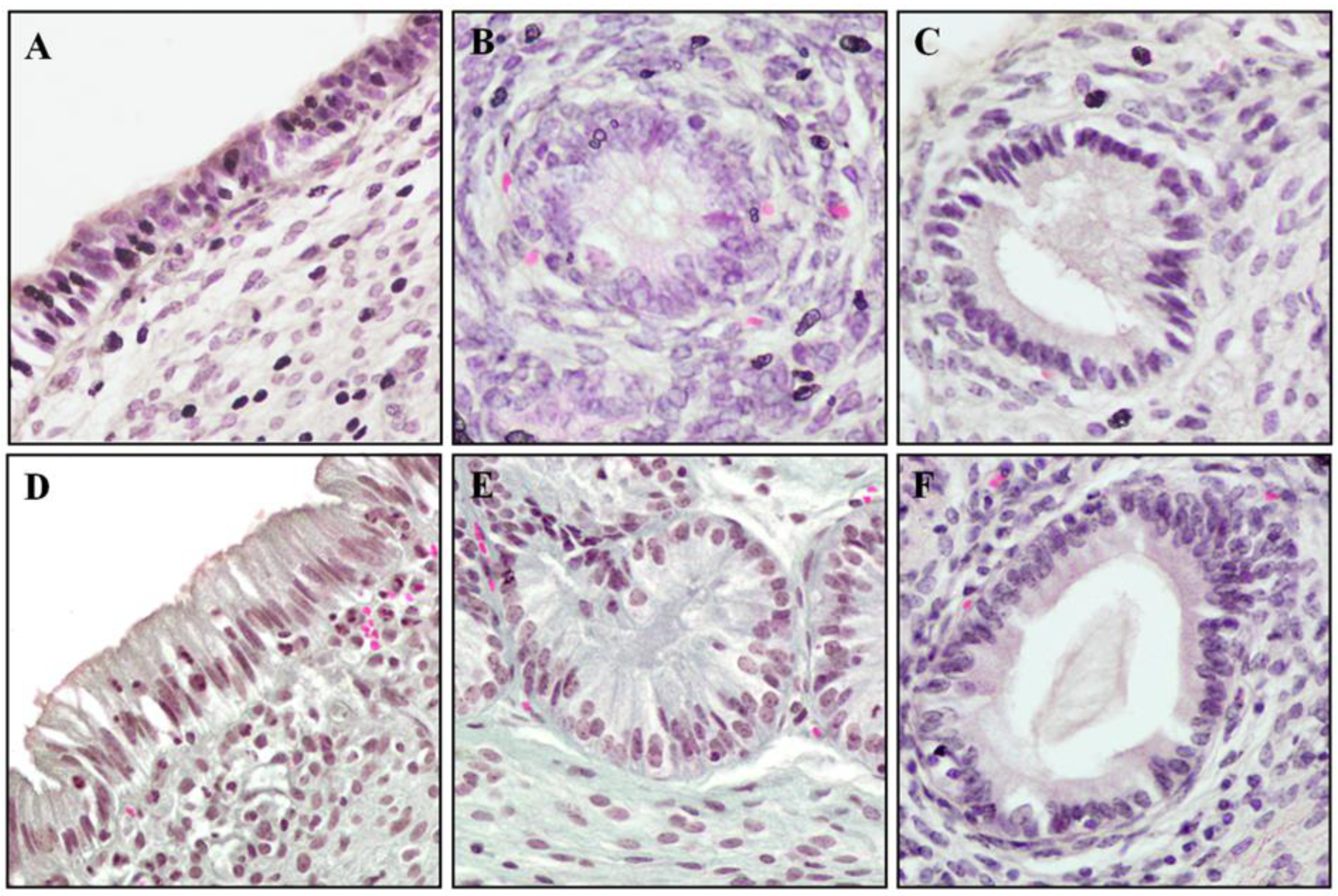
2.7. Histological Evaluation of Endometrium Reaction
2.8. Correlations
3. Discussion
4. Limitations
5. Materials and Methods
5.1. Animals
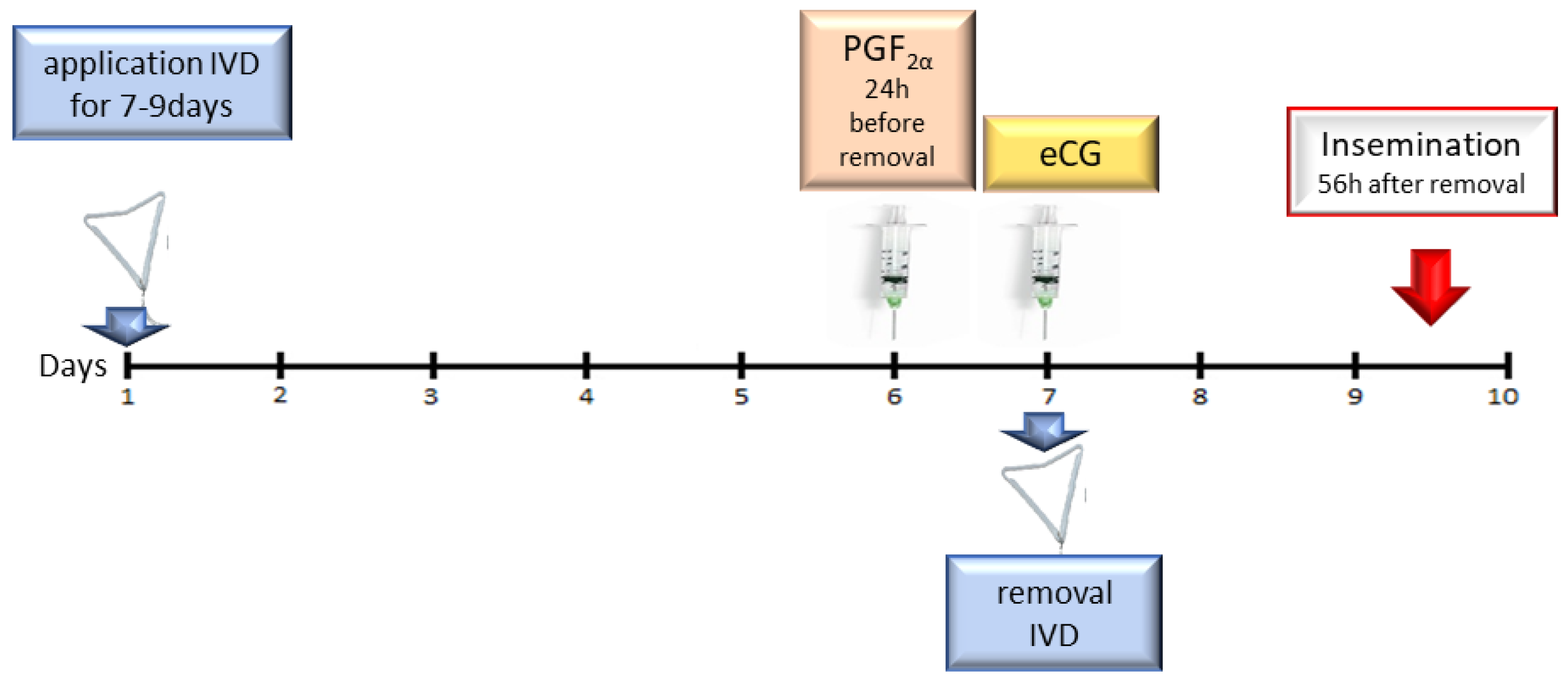
5.2. Therapy Protocol
5.3. Sampling
5.4. Analyses of mRNA Expression in Endometrium
5.5. Measurements of the Systemic Concentration of 17 ß-Estradiol and Progesterone
5.6. Endometrial Measurements
5.7. Statistical Analysis
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mwaanga, E.; Janowski, T. Anoestrus in Dairy Cows: Causes, Prevalence and Clinical Forms. Reprod. Domest. Anim. 2000, 35, 193–200. [Google Scholar] [CrossRef]
- Cerri, R.L.A.; Santos, J.E.P.; Juchem, S.O.; Galvão, K.N.; Chebel, R.C. Timed Artificial Insemination with Estradiol Cypionate or Insemination at Estrus in High-Producing Dairy Cows. J. Dairy Sci. 2004, 87, 3704–3715. [Google Scholar] [CrossRef]
- Santos, J.E.P.; Rutigliano, H.M.; Filho, M.F.S. Risk factors for resumption of postpartum estrous cycles and embryonic survival in lactating dairy cows. Anim. Reprod. Sci. 2009, 110, 207–221. [Google Scholar] [CrossRef]
- Stevenson, J.S.; Pursley, J.R.; Garverick, H.A.; Fricke, P.M.; Kesler, D.J.; Ottobre, J.S.; Wiltbank, M.C. Treatment of Cycling and Noncycling Lactating Dairy Cows with Progesterone During Ovsynch. J. Dairy Sci. 2006, 89, 2567–2578. [Google Scholar] [CrossRef]
- Bartlett, P.C.; Kirk, J.; Coe, P.; Marteniuk, J.; Mather, E.C. Descriptive epidemiology of anestrus in Michigan Holstein-Friesian cattle. Theriogenology 1987, 27, 459–476. [Google Scholar] [CrossRef]
- Martinez, J.; Thibier, M. Reproductive disorders in dairy cattle. II. Interrelationships between pre-or post-service infectious and functional disorders. Theriogenology 1984, 21, 583–590. [Google Scholar] [CrossRef]
- Nation, D.P.; Morton, J.; Cavalieri, J.; Macmillan, K.L. Factors associated with the incidence of Phantom cows in Australian dairy herds. In Proceedings of the New Zealand Society of Animal Production; New Zealand Society of Animal Production: Christchurch, New Zealand, 2001; Volume 61, pp. 180–183. [Google Scholar]
- O’Connor, M.L.; Senger, P.L. Estrus Detection. In Current Therapy in Large Animal Theriogenology, 2nd ed.; Youngquist, R.S., Ed.; W.B. Saunders Company: Philadelphia, PA, USA, 1997; p. 276. [Google Scholar]
- Cardoso Consentini, C.E.; Wiltbank, M.C.; Sartori, R. Factors That Optimize Reproductive Efficiency in Dairy Herds with an Emphasis on Timed Artificial Insemination Programs. Animals 2021, 11, 301. [Google Scholar] [CrossRef]
- Xu, C.; Xia, C.; Sun, Y.; Xiao, X.; Wang, G.; Fan, Z.; Shu, S.; Zhang, H.; Xu, C.; Yang, W. Metabolic profiles using 1H-nuclear magnetic resonance spectroscopy in postpartum dairy cows with ovarian inactivity. Theriogenology 2015, 86, 1475–1481. [Google Scholar] [CrossRef] [Green Version]
- Kumar, P.R.; Kumar, S.S.; Suresh Dinkar Kharche, C.S.G.; Behera, B.K.; Shukla, S.N.; Kumar, H.; Agarwal, S.K. Anestrus in Cattle and Buffalo: Indian Perspective. Adv. Anim. Vet. Sci. 2014, 2, 124–138. [Google Scholar] [CrossRef]
- Barański, W.; Nowicki, A.; Zduńczyk, S.; Janowski, T. Incidence of clinical form of anestrus after unsuccessful service in cows in eight dairy herds in north-east of Poland. Pol. J. Vet. Sci. 2018, 21, 377–381. [Google Scholar] [CrossRef] [PubMed]
- Senger, P.L. The Estrus Detection Problem: New Concepts, Technologies, and Possibilities. J. Dairy Sci. 1994, 77, 2745–2753. [Google Scholar] [CrossRef]
- Bekara, M.E.A.; Bareille, N. Quantification by simulation of the effect of herd management practices and cow fertility on the reproductive and economic performance of Holstein dairy herds. J. Dairy Sci. 2019, 102, 9435–9457. [Google Scholar] [CrossRef] [Green Version]
- Kotlarczyk, A.M.; Grzyb, M.; Korzekwa, A.J. Regulation of uterine function during estrous cycle, anestrus phase and pregnancy by steroids in red deer (Cervus elaphus L.). Sci. Rep. 2021, 11, 20109. [Google Scholar] [CrossRef] [PubMed]
- Núñez-Olivera, R.; Cuadro, F.; Bosolasco, D.; de Brun, V.; de la Mata, J.; Brochado, C.; Meikle, A.; Bó, G.A.; Menchaca, A. Effect of equine chorionic gonadotropin (eCG) administration and proestrus length on ovarian response, uterine functionality and pregnancy rate in beef heifers inseminated at a fixed-time. Theriogenology 2020, 151, 16–27. [Google Scholar] [CrossRef]
- Wiltbank, M.C.; Gümen, A.; Sartori, R. Physiological classification of anovulatory conditions in cattle. Theriogenology 2002, 57, 21–52. [Google Scholar] [CrossRef]
- Peter, A.T.; Levine, H.; Drost, M.; Bergfelt, D.R. Compilation of classical and contemporary terminology used to describe morphological aspects of ovarian dynamics in cattle. Theriogenology 2009, 71, 1343–1357. [Google Scholar] [CrossRef]
- Lopez, H.; Caraviello, D.Z.; Satter, L.D.; Fricke, P.M.; Wiltbank, M.C. Relationship Between Level of Milk Production and Multiple Ovulations in Lactating Dairy Cows. J. Dairy Sci. 2005, 88, 2783–2793. [Google Scholar] [CrossRef]
- Sangsritavong, S.; Combs, D.K.; Sartori, R.; Armentano, L.E.; Wiltbank, M.C. High Feed Intake Increases Liver Blood Flow and Metabolism of Progesterone and Estradiol-17β in Dairy Cattle. J. Dairy Sci. 2002, 85, 2831–2842. [Google Scholar] [CrossRef] [Green Version]
- Peter, A.T.; Vos, P.L.A.M.; Ambrose, D.J. Postpartum anestrus in dairy cattle. Theriogenology 2009, 71, 1333–1342. [Google Scholar] [CrossRef]
- Bisinotto, R.S.; Lean, I.J.; Thatcher, W.W.; Santos, J.E.P. Meta-analysis of progesterone supplementation during timed artificial insemination programs in dairy cows. J. Dairy Sci. 2015, 98, 2472–2487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- e Silva, L.O.; Valenza, A.; Alves, R.L.O.R.; da Silva, M.A.; da Silva, T.J.B.; Motta, J.C.L.; Drum, J.N.; Madureira, G.; de Souza, A.H.; Sartori, R. Progesterone release profile and follicular development in Holstein cows receiving intravaginal progesterone devices. Theriogenology 2021, 172, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Robinson, R.S.; Mann, G.E.; Lamming, G.E.; Wathes, D.C. Expression of oxytocin, oestrogen and progesterone receptors in uterine biopsy samples throughout the oestrous cycle and early pregnancy in cows. Reproduction 2001, 122, 965–979. [Google Scholar] [CrossRef]
- Kimmins, S.; Maclaren, L.A. Oestrous cycle and pregnancy effects on the distribution of eestrogen and progesterone receptors in Bovine endometrium. Placenta 2001, 22, 742–748. [Google Scholar] [CrossRef]
- Kowalik, M.K.; Slonina, D.; Rekawiecki, R.; Kotwica, J. Expression of progesterone receptor membrane component (PGRMC) 1 and 2, serpine mrnabinding protein 1 (SERBP1) and nuclear progesterone receptor (PGR) in the bovine endometrium during the estrous cycle and the first trimester of pregnancy. Reprod. Biol. 2013, 13, 15–23. [Google Scholar] [CrossRef]
- Xie, J.; Kusnadi, E.P.; Furic, L.; Selth, L.A. Regulation of mrna translation by hormone receptors in breast and prostate cancer. Cancers 2021, 13, 3254. [Google Scholar] [CrossRef]
- Witkowska-Piłaszewicz, O.; Bąska, P.; Czopowicz, M.; Żmigrodzka, M.; Szarska, E.; Szczepaniak, J.; Nowak, Z.; Winnicka, A.; Cywińska, A. Anti-Inflammatory State in Arabian Horses Introduced to the Endurance Training. Animals 2019, 9, 616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharov, A.A.; Sharova, L.V.; Shaik, N.; Nedorezov, T.; Piao, Y.; Ko, M.S.H. Database for mRNA Half-Life of 19 977 Genes Obtained by DNA Microarray Analysis of Pluripotent and Differentiating Mouse Embryonic Stem Cells. DNA Res. 2008, 16, 45–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, B.; Elguero, S.; Thakore, S.; Dahoud, W.; Bedaiwy, M.; Mesiano, S. Role of nuclear progesterone receptor isoforms in uterine pathophysiology. Hum. Reprod. Update 2015, 21, 155–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eozenou, C.; Lesage-padilla, A.; Mauffré, V.; Healey, G.D.; Camous, S.; Bolifraud, P.; Giraud-delville, C.; Vaiman, D.; Shimizu, T.; Miyamoto, A.; et al. FOXL2 is a progesterone target gene in the endometrium of ruminants. Int. J. Mol. Sci. 2020, 21, 1478. [Google Scholar] [CrossRef] [Green Version]
- Tsai, M.-J.; Clark, J.; Schrader, W.; O’Malley, B. Mechanisms of action of hormones that act as transcription-regulatory factors. In Williams Textbook of Endocrinology; Wilson, J., Foster, D., Kronenberg, H.L.P., Eds.; W.B. Saunders Company: Philadelphia, PA, USA, 1998; pp. 55–94. [Google Scholar]
- Graham, J.D.; Clarke, C.L. Physiological action of progesterone in target tissues. Endocr. Rev. 1997, 18, 502–519. [Google Scholar] [PubMed] [Green Version]
- Rękawiecki, R.; Kowalik, M.K.; Kotwica, J. Onapristone (ZK299) and mifepristone (RU486) regulate the messenger RNA and protein expression levels of the progesterone receptor isoforms A and B in the bovine endometrium. Theriogenology 2015, 84, 348–357. [Google Scholar] [CrossRef] [PubMed]
- Piotrowska-Tomala, K.K.; Jonczyk, A.W.; Skarzynski, D.J.; Szóstek-Mioduchowska, A.Z. Luteinizing hormone and ovarian steroids affect in vitro prostaglandin production in the equine myometrium and endometrium. Theriogenology 2020, 153, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Robinson, R. The expression of the IGF system in the bovine uterus throughout the oestrous cycle and early pregnancy. J. Endocrinol. 2000, 165, 231–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, R.S.; Mann, G.E.; Lamming, G.E.; Wathes, D.C. The effect of pregnancy on the expression of uterine oxytocin, oestrogen and progesterone receptors during early pregnancy in the cow. J. Endocrinol. 1999, 160, 21–33. [Google Scholar] [CrossRef]
- Paterni, I.; Granchi, C.; Katzenellenbogen, J.A.; Minutolo, F. Estrogen receptors alpha (ERα) and beta (ERβ): Subtype-selective ligands and clinical potential. Steroids 2014, 90, 13–29. [Google Scholar] [CrossRef] [Green Version]
- McDougall, S. Effects of treatment of anestrous dairy cows with gonadotropin-releasing hormone, prostaglandin, and progesterone. J. Dairy Sci. 2010, 93, 1944–1959. [Google Scholar] [CrossRef] [Green Version]
- McDougall, S.; Compton, C.W..; Anniss, F. Effect of exogenous progesterone and oestradiol on plasma progesterone concentrations and follicle wave dynamics in anovulatory anoestrous post-partum dairy cattle. Anim. Reprod. Sci. 2004, 84, 303–314. [Google Scholar] [CrossRef]
- McDougall, S.; Compton, C.W.R.; Hanlon, D.W.; Davidson, P.J.; Sullivan, D.J.; Gore, A.H.; Anniss, F.M. Reproductive performance in anestrous dairy cows following treatment with two protocols and two doses of progesterone. Theriogenology 2005, 63, 1529–1548. [Google Scholar] [CrossRef]
- Rękawiecki, R.; Kowalik, M.K.; Kotwica, J. Cloning and expression of progesterone receptor isoforms A and B in bovine corpus luteum. Reprod. Fertil. Dev. 2015, 27, 1029–1037. [Google Scholar] [CrossRef]
- Zhao, S.; Fernald, R.D. Comprehensive Algorithm for Quantitative Real-Time Polymerase Chain Reaction. J. Comput. Biol. 2005, 12, 1047–1064. [Google Scholar] [CrossRef]
- Świątek, M. Wizualizacja Immunofluorescencyjna Receptorów Estrogenowych i Progesteronowych w Endometrium krów Mlecznych w Anestrus Typu II. Engineer’s Thesis, Warsaw University of Life Sciences, Warsaw, Poland, 2017. [Google Scholar]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trela, M.; Witkowska-Piłaszewicz, O.; Domańska, D.; Kaczmarek, M.M.; Pawliński, B.; Gajewski, Z.; Domino, M. The Influence of Intravaginal Gestagens Treatment on the Morphological Features and Endometrial Steroid Hormone Receptors Content during Anestrus Type II in Dairy Cattle. Int. J. Mol. Sci. 2022, 23, 1235. https://doi.org/10.3390/ijms23031235
Trela M, Witkowska-Piłaszewicz O, Domańska D, Kaczmarek MM, Pawliński B, Gajewski Z, Domino M. The Influence of Intravaginal Gestagens Treatment on the Morphological Features and Endometrial Steroid Hormone Receptors Content during Anestrus Type II in Dairy Cattle. International Journal of Molecular Sciences. 2022; 23(3):1235. https://doi.org/10.3390/ijms23031235
Chicago/Turabian StyleTrela, Michał, Olga Witkowska-Piłaszewicz, Dominika Domańska, Monika M. Kaczmarek, Bartosz Pawliński, Zdzisław Gajewski, and Małgorzata Domino. 2022. "The Influence of Intravaginal Gestagens Treatment on the Morphological Features and Endometrial Steroid Hormone Receptors Content during Anestrus Type II in Dairy Cattle" International Journal of Molecular Sciences 23, no. 3: 1235. https://doi.org/10.3390/ijms23031235
APA StyleTrela, M., Witkowska-Piłaszewicz, O., Domańska, D., Kaczmarek, M. M., Pawliński, B., Gajewski, Z., & Domino, M. (2022). The Influence of Intravaginal Gestagens Treatment on the Morphological Features and Endometrial Steroid Hormone Receptors Content during Anestrus Type II in Dairy Cattle. International Journal of Molecular Sciences, 23(3), 1235. https://doi.org/10.3390/ijms23031235