
Fetal Undernutrition Modifies Vascular RAS Balance Enhancing Oxidative Damage and Contributing to Remodeling


 ,
, 
Abstract
1. Introduction
2. Results
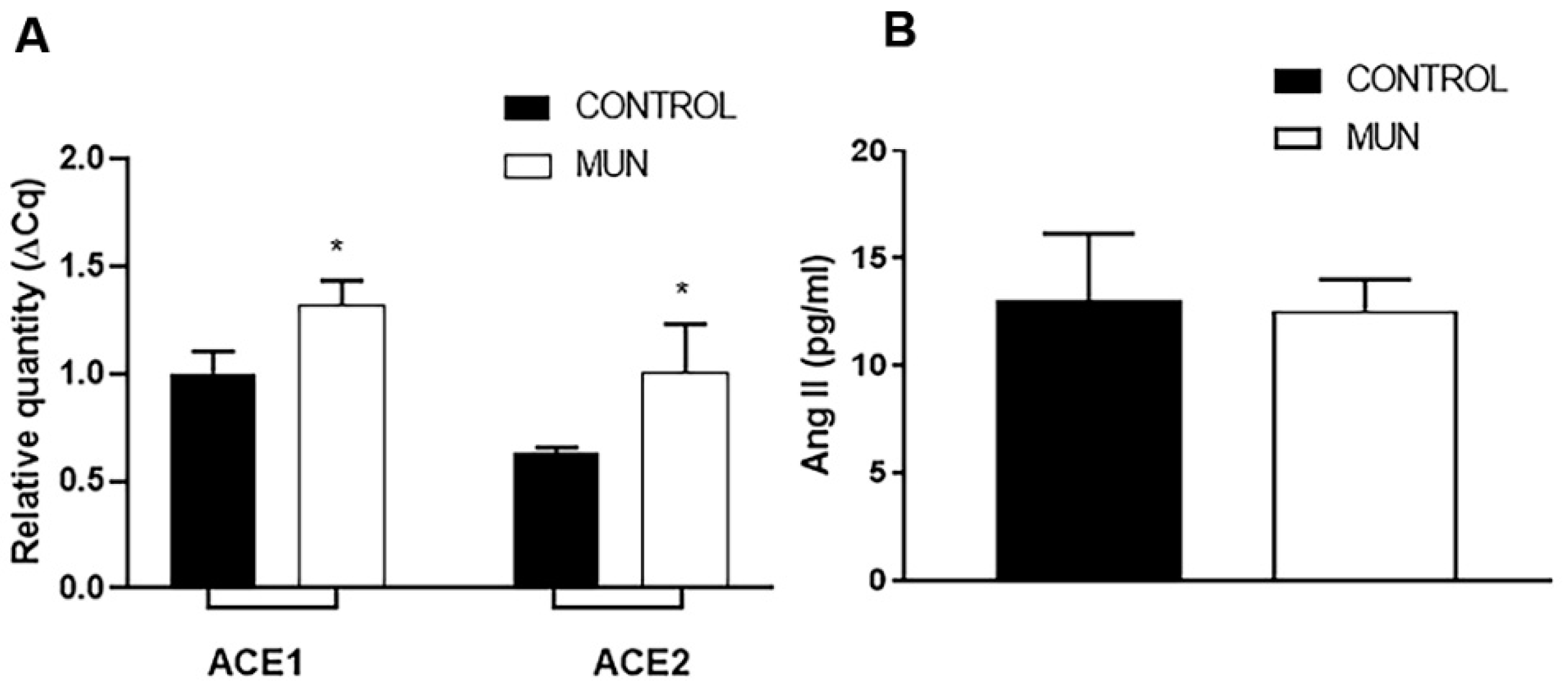
2.1. Influence of Fetal Undernutrition on Arterial ACE and ACE2 Expression and Plasma Ang II Levels
2.2. Influence of Fetal Undernutrition on Renin-Angiotensin System Receptors
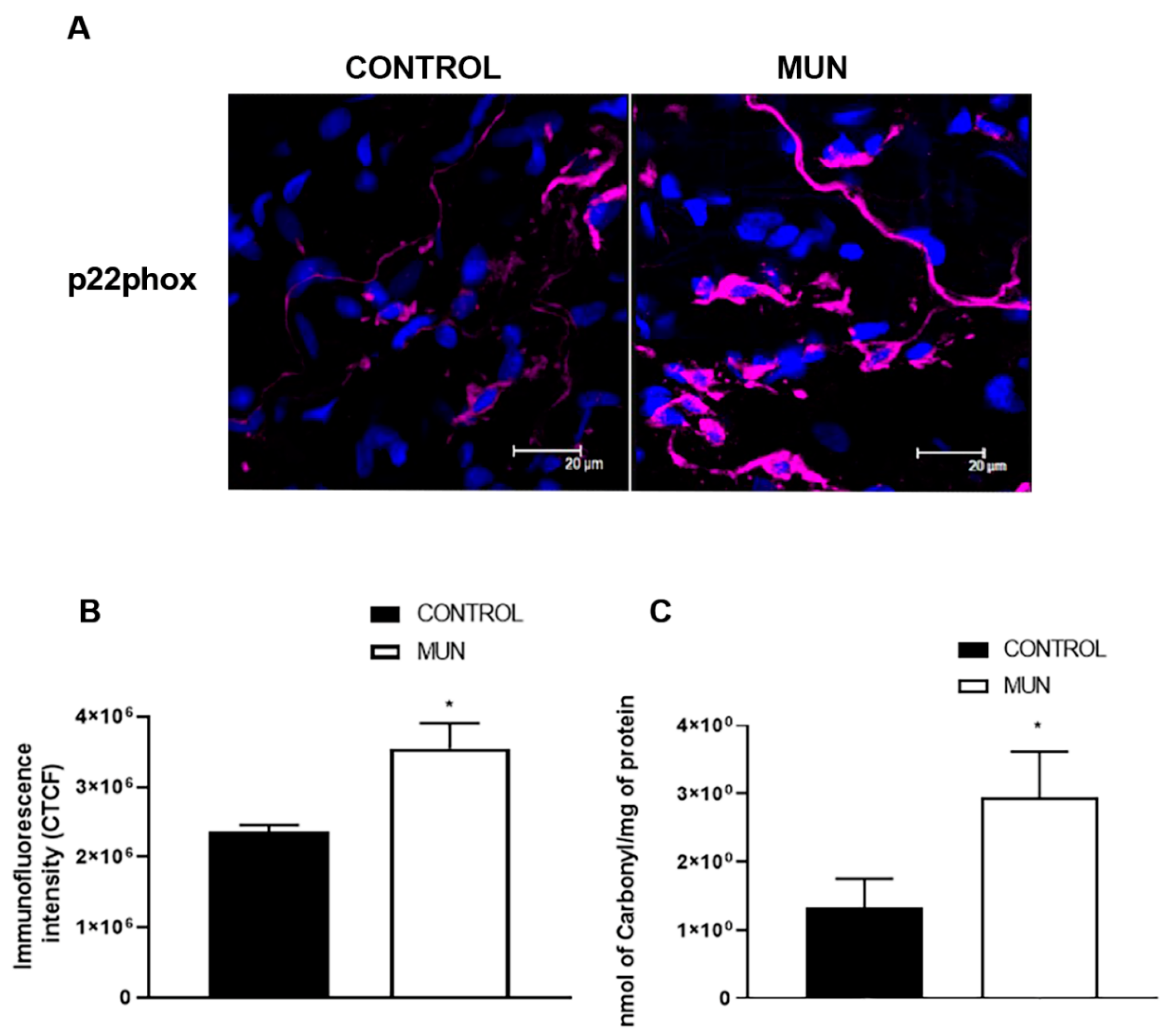
2.3. Influence of Fetal Undernutrition on NADPH Oxidase and Oxidative Damage
2.4. Influence of Fetal Undernutrition on Vascular Wall Remodelling and BP
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Experimental Model of FPH
4.3. Hemodynamic Parameters Measurement and Blood Sample Collection
4.4. Plasma Measurements
4.5. Immunohistochemistry
4.6. qPCR
4.7. Histology
4.8. Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Santos, R.A.; Ferreira, A.J.; Verano-Braga, T.; Bader, M. Angiotensin-converting enzyme 2, angiotensin-(1-7) and Mas: New players of the renin-angiotensin system. J. Endocrinol. 2013, 216, R1–R17. [Google Scholar] [CrossRef]
- Etelvino, G.M.; Peluso, A.; Santos, R.A.S. New Components of the Renin-Angiotensin System: Alamandine and the Mas-Related G Protein-Coupled Receptor D. Curr. Hypertens. Rep. 2014, 16, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Villela, D.C.; Passos-Silva, D.G.; Santos, R.A. Alamandine: A new member of the angiotensin family. Curr. Opin. Nephrol. Hypertens. 2014, 23, 130–134. [Google Scholar] [CrossRef]
- Barker, D.J.; Osmond, C. Low birth weight and hypertension. BMJ 1988, 297, 134–135. [Google Scholar] [CrossRef] [PubMed]
- Barker, D.J.; Clark, P.M. Fetal undernutrition and disease in later life. Rev. Reprod. 1997, 2, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Alexander, B.T. Fetal programming of hypertension. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006, 290, R1–R10. [Google Scholar] [CrossRef]
- Alexander, B.T.; Dasinger, J.H.; Intapad, S. Fetal Programming and Cardiovascular Pathology. Compr. Physiol. 2015, 5, 997–1025. [Google Scholar] [CrossRef]
- Chappell, M.C.; Marshall, A.A.; Alzayadneh, E.M.; Shaltout, H.A.; Diz, D.I. Update on the Angiotensin converting enzyme 2-Angiotensin (1-7)-MAS receptor axis: Fetal programing, sex differences, and intracellular pathways. Front. Endocrinol. 2014, 4, 201. [Google Scholar] [CrossRef]
- South, A.M.; Shaltout, H.; Washburn, L.K.; Hendricks, A.S.; Diz, D.I.; Chappell, M.C. Fetal programming and the angiotensin-(1-7) axis: A review of the experimental and clinical data. Clin. Sci. 2019, 133, 55–74. [Google Scholar] [CrossRef]
- Manning, J.; Vehaskari, V.M. Postnatal modulation of prenatally programmed hypertension by dietary Na and ACE inhibition. Am. J. Physiol. Integr. Comp. Physiol. 2005, 288, R80–R84. [Google Scholar] [CrossRef]
- Langley-Evans, S.C.; Jackson, A.A. Captopril normalises systolic blood pressure in rats with hypertension induced by fetal exposure to maternal low protein diets. Comp. Biochem. Physiol. Part A Physiol. 1995, 110, 223–228. [Google Scholar] [CrossRef]
- Sherman, R.C.; Langley-Evans, S.C. Antihypertensive treatment in early postnatal life modulates prenatal dietary influences upon blood pressure in the rat. Clin. Sci. 2000, 98, 269. [Google Scholar] [CrossRef]
- Rodríguez-Rodríguez, P.; De Pablo, L.L.; García-Prieto, C.F.; Somoza, B.; Quintana-Villamandos, B.; De Diego, J.J.G.; Gutierrez-Arzapalo, P.Y.; Ramiro-Cortijo, D.; Gonzalez-Garcia, M.C.; Arribas, S.M. Long term effects of fetal undernutrition on rat heart. Role of hypertension and oxidative stress. PLoS ONE 2017, 12, e0171544. [Google Scholar] [CrossRef]
- Rodríguez-Rodríguez, P.; Cortijo, D.R.; Reyes-Hernández, C.G.; De Pablo, A.L.L.; González, M.C.; Arribas, S.M. Implication of Oxidative Stress in Fetal Programming of Cardiovascular Disease. Front. Physiol. 2018, 9, 602. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Rodríguez, P.; Vieira-Rocha, M.; Quintana-Villamandos, B.; Monedero-Cobeta, I.; Prachaney, P.; de Pablo, A.L.; González, M.; Morato, M.; Diniz, C.; Arribas, S. Implication of RAS in Postnatal Cardiac Remodeling, Fibrosis and Dysfunction Induced by Fetal Undernutrition. Pathophysiology 2021, 28, 273–290. [Google Scholar] [CrossRef]
- Vieira-Rocha, M.; Rodríguez-Rodríguez, P.; Sousa, J.; González, M.; Arribas, S.; de Pablo, A.L.; Diniz, C. Vascular angiotensin AT1 receptor neuromodulation in fetal programming of hypertension. Vasc. Pharmacol. 2018, 117, 27–34. [Google Scholar] [CrossRef]
- Havelka, G.E.; Kibbe, M.R. The Vascular Adventitia: Its Role in the Arterial Injury Response. Vasc. Endovasc. Surg. 2011, 45, 381–390. [Google Scholar] [CrossRef]
- Berry, C.; Hamilton, C.A.; Brosnan, M.J.; Magill, F.G.; Berg, G.A.; McMurray, J.J.; Dominiczak, A.F. Investigation into the sources of superoxide in human blood vessels: Angiotensin II increases superoxide production in human internal mammary arteries. Circulation 2000, 101, 2206–2212. [Google Scholar] [CrossRef]
- Sirker, A.; Zhang, M.; Murdoch, C.; Shah, A.M. Involvement of NADPH Oxidases in Cardiac Remodelling and Heart Failure. Am. J. Nephrol. 2007, 27, 649–660. [Google Scholar] [CrossRef]
- Muller, D.N.; Bohlender, J.; Hilgers, K.F.; Dragun, D.; Costerousse, O.; Ménard, J.; Luft, F. Vascular Angiotensin-Converting Enzyme Expression Regulates Local Angiotensin II. Hypertension 1997, 29, 98–104. [Google Scholar] [CrossRef]
- Müller, D.; Luft, F. The renin-angiotensin system in the vessel wall. Basic Res. Cardiol. 1998, 93, s007–s014. [Google Scholar] [CrossRef] [PubMed]
- Navar, L.G.; Mitchell, K.D.; Harrison-Bernard, L.M.; Kobori, H.; Nishiyama, A. Review: Intrarenal angiotensin II levels in normal and hypertensive states. J. Renin-Angiotensin-Aldosterone Syst. 2001, 2, S176–S184. [Google Scholar] [CrossRef] [PubMed]
- Xiao, F.; Burns, K.D. Measurement of Angiotensin Converting Enzyme 2 Activity in Biological Fluid (ACE2). Methods Mol. Biol. 2017, 1527, 101–115. [Google Scholar] [CrossRef]
- Yamaleyeva, L.M.; Gilliam-Davis, S.; Almeida, I.; Brosnihan, K.B.; Lindsey, S.H.; Chappell, M.C. Differential regulation of circulating and renal ACE2 and ACE in hypertensive mRen2.Lewis rats with early-onset diabetes. Am. J. Physiol. Physiol. 2012, 302, F1374–F1384. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.-R.; Tain, Y.-L.; Tiao, M.-M.; Chen, C.-C.; Sheen, J.-M.; Lin, I.-C.; Li, S.-W.; Tsai, C.-C.; Lin, Y.-J.; Hsieh, K.-S.; et al. Prenatal dexamethasone and postnatal high-fat diet have a synergistic effect of elevating blood pressure through a distinct programming mechanism of systemic and adipose renin–angiotensin systems. Lipids Health Dis. 2018, 17, 50. [Google Scholar] [CrossRef]
- Paul, M.; Mehr, A.P.; Kreutz, R. Physiology of Local Renin-Angiotensin Systems. Physiol. Rev. 2006, 86, 747–803. [Google Scholar] [CrossRef]
- Vehaskari, V.M.; Stewart, T.; Lafont, D.; Soyez, C.; Seth, D.; Manning, J. Kidney angiotensin and angiotensin receptor expression in prenatally programmed hypertension. Am. J. Physiol. Physiol. 2004, 287, F262–F267. [Google Scholar] [CrossRef]
- Pladys, P.; Lahaie, I.; Cambonie, G.; Thibault, G.; Lê, N.L.O.; Abran, D.; Nuyt, A.M. Role of Brain and Peripheral Angiotensin II in Hypertension and Altered Arterial Baroreflex Programmed during Fetal Life in Rat. Pediatr. Res. 2004, 55, 1042–1049. [Google Scholar] [CrossRef]
- Yzydorczyk, C.; Gobeil, F.; Cambonie, G.; Lahaie, I.; Lê, N.L.O.; Samarani, S.; Ahmad, A.; Lavoie, J.C.; Oligny, L.L.; Pladys, P.; et al. Exaggerated vasomotor response to ANG II in rats with fetal programming of hypertension associated with exposure to a low-protein diet during gestation. Am. J. Physiol. Integr. Comp. Physiol. 2006, 291, R1060–R1068. [Google Scholar] [CrossRef]
- Mao, C.; Zhang, H.; Xiao, D.; Zhu, L.; Ding, Y.; Zhang, Y.; Wu, L.; Xu, Z.; Zhang, L. Perinatal nicotine exposure alters AT1 and AT2 receptor expression pattern in the brain of fetal and offspring rats. Brain Res. 2008, 1243, 47–52. [Google Scholar] [CrossRef]
- Goyal, R.; Leitzke, A.; Goyal, D.; Gheorghe, C.P.; Longo, L.D. Antenatal maternal hypoxic stress: Adaptations in fetal lung Renin-Angiotensin system. Reprod. Sci. 2011, 18, 180–189. [Google Scholar] [CrossRef] [PubMed]
- Mao, C.; Hou, J.; Ge, J.; Hu, Y.; Ding, Y.; Zhou, Y.; Zhang, H.; Xu, Z.; Zhang, L. Changes of renal AT1/AT2 receptors and structures in ovine fetuses following exposure to long-term hypoxia. Am. J. Nephrol. 2010, 31, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Vieira-Rocha, M.S.; Sousa, J.B.; Rodriguez-Rodriguez, P.; Morato, M.; Arribas, S.M.; Diniz, C. Insights into sympathetic nervous system and GPCR interplay in fetal programming of hypertension: A bridge for new pharmacological strategies. Drug Discov. Today 2020, 25, 739–747. [Google Scholar] [CrossRef]
- Mesquita, F.F.; Gontijo, J.A.R.; Boer, P.A. Expression of renin-angiotensin system signalling compounds in maternal protein-restricted rats: Effect on renal sodium excretion and blood pressure. Nephrol. Dial. Transplant. 2009, 25, 380–388. [Google Scholar] [CrossRef] [PubMed]
- Tsukuda, K.; Mogi, M.; Iwanami, J.; Min, L.-J.; Jing, F.; Ohshima, K.; Horiuchi, M. Influence of angiotensin II type 1 receptor-associated protein on prenatal development and adult hypertension after maternal dietary protein restriction during pregnancy. J. Am. Soc. Hypertens. 2012, 6, 324–330. [Google Scholar] [CrossRef] [PubMed]
- Gwathmey, T.M.; Shaltout, H.A.; Rose, J.C.; Diz, D.I.; Chappell, M.C. Glucocorticoid-Induced Fetal Programming Alters the Functional Complement of Angiotensin Receptor Subtypes Within the Kidney. Hypertension 2011, 57, 620–626. [Google Scholar] [CrossRef] [PubMed]
- Marshall, A.C.; Shaltout, H.A.; Nautiyal, M.; Rose, J.C.; Chappell, M.C.; Diz, D.I. Fetal betamethasone exposure attenuates angiotensin-(1-7)-Mas receptor expression in the dorsal medulla of adult sheep. Peptides 2013, 44, 25–31. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Sousa, J.; Vieira-Rocha, M.S.; Sá, C.; Ferreirinha, F.; Correia-De-Sá, P.; Fresco, P.; Diniz, C. Lack of Endogenous Adenosine Tonus on Sympathetic Neurotransmission in Spontaneously Hypertensive Rat Mesenteric Artery. PLoS ONE 2014, 9, e105540. [Google Scholar] [CrossRef] [PubMed]
- Sousa, J.B.; Vieira-Rocha, M.S.; Arribas, S.M.; Gonzalez-Garcia, M.C.; Fresco, P.; Diniz, C. Endothelial and Neuronal Nitric Oxide Activate Distinct Pathways on Sympathetic Neurotransmission in Rat Tail and Mesenteric Arteries. PLoS ONE 2015, 10, e0129224. [Google Scholar] [CrossRef] [PubMed]
- Stenmark, K.R.; Yeager, M.E.; El Kasmi, K.C.; Nozik-Grayck, E.; Gerasimovskaya, E.V.; Li, M.; Riddle, S.R.; Frid, M.G. The Adventitia: Essential Regulator of Vascular Wall Structure and Function. Annu. Rev. Physiol. 2013, 75, 23–47. [Google Scholar] [CrossRef]
- Csányi, G.; Taylor, W.R.; Pagano, P.J. NOX and inflammation in the vascular adventitia. Free Radic. Biol. Med. 2009, 47, 1254–1266. [Google Scholar] [CrossRef]
- Cat, A.N.D.; Montezano, A.; Burger, D.; Touyz, R.M. Angiotensin II, NADPH Oxidase, and Redox Signaling in the Vasculature. Antioxid. Redox Signal. 2013, 19, 1110–1120. [Google Scholar] [CrossRef]
- Liu, J.; Yang, F.; Yang, X.-P.; Jankowski, M.; Pagano, P.J. NAD(P)H Oxidase Mediates Angiotensin II–Induced Vascular Macrophage Infiltration and Medial Hypertrophy. Arter. Thromb. Vasc. Biol. 2003, 23, 776–782. [Google Scholar] [CrossRef] [PubMed]
- Ushio-Fukai, M.; Zafari, A.M.; Fukui, T.; Ishizaka, N.; Griendling, K.K. p22phox is a critical component of the superoxide-generating NADH/NADPH oxidase system and regulates angiotensin II-induced hypertrophy in vascular smooth muscle cells. J. Biol. Chem. 1996, 271, 23317–23321. [Google Scholar] [CrossRef] [PubMed]
- Paravicini, T.M.; Touyz, R.M. Redox signaling in hypertension. Cardiovasc. Res. 2006, 71, 247–258. [Google Scholar] [CrossRef]
- Condezo-Hoyos, L.; Arribas, S.M.; Abderrahim, F.; Somoza, B.; Gil-Ortega, M.; Díaz-Gil, J.J.; Conde, M.V.; Susin, C.; González, M.C. Liver growth factor treatment reverses vascular and plasmatic oxidative stress in spontaneously hypertensive rats. J. Hypertens. 2012, 30, 1185–1194. [Google Scholar] [CrossRef]
- Tinajero, M.G.; Gotlieb, A.I. Recent Developments in Vascular Adventitial Pathobiology. Am. J. Pathol. 2020, 190, 520–534. [Google Scholar] [CrossRef]
- Sousa, J.; Diniz, C. Vascular Sympathetic Neurotransmission and Endothelial Dysfunction. In Endothelial Dysfunction—Old Concepts and New Challenges; Lenasi, H., Ed.; InTechOpen: London, UK, 2018; ISBN 978-1-78984-254-8. Available online: https://www.intechopen.com/chapters/58245 (accessed on 22 December 2021).
- Somoza, B.; González, M.C.; González, J.M.; Abderrahim, F.; Arribas, S.M.; Fernández-Alfonso, M.S. Modulatory role of the adventitia on noradrenaline and angiotensin II responses role of endothelium and AT2 receptors. Cardiovasc. Res. 2005, 65, 478–486. [Google Scholar] [CrossRef]
- Rodríguez, P.R.; de Pablo, A.L.L.; Condezo-Hoyos, L.; Martín-Cabrejas, M.A.; Aguilera, Y.; Ruiz-Hurtado, G.; Gutierrez-Arzapalo, P.Y.; Cortijo, D.R.; Fernandez-Alfonso, M.S.; Gonzalez-Garcia, M.C.; et al. Fetal undernutrition is associated with perinatal sex-dependent alterations in oxidative status. J. Nutr. Biochem. 2015, 26, 1650–1659. [Google Scholar] [CrossRef]
- Griendling, K.K.; Camargo, L.L.; Rios, F.J.; Alves-Lopes, R.; Touyz, R.M. Oxidative Stress and Hypertension. Circ. Res. 2021, 128, 993–1020. [Google Scholar] [CrossRef]
- Guzik, T.J.; Touyz, R.M. Oxidative Stress, Inflammation, and Vascular Aging in Hypertension. Hypertension 2017, 70, 660–667. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, L.C.; Oliveira-Paula, G.H. Sources and Effects of Oxidative Stress in Hypertension. Curr. Hypertens. Rev. 2020, 16, 166–180. [Google Scholar] [CrossRef] [PubMed]
- Conde, M.V.; Gonzalez, M.C.; Quintana-Villamandos, B.; Abderrahim, F.; Briones, A.M.; Condezo-Hoyos, L.; Regadera, J.; Susin, C.; de Diego, J.J.G.; Delgado-Baeza, E.; et al. Liver growth factor treatment restores cell-extracellular matrix balance in resistance arteries and improves left ventricular hypertrophy in SHR. Am. J. Physiol. Circ. Physiol. 2011, 301, H1153–H1165. [Google Scholar] [CrossRef] [PubMed]
- González, M.C.; Arribas, S.M.; Molero, F.; Fernández-Alfonso, M.S. Effect of removal of adventitia on vascular smooth muscle contraction and relaxation. Am. J. Physiol. Circ. Physiol. 2001, 280, H2876–H2881. [Google Scholar] [CrossRef]
- Masi, S.; Uliana, M.; Virdis, A. Angiotensin II and vascular damage in hypertension: Role of oxidative stress and sympathetic activation. Vasc. Pharmacol. 2019, 115, 13–17. [Google Scholar] [CrossRef]
- Schiffrin, E.L. Vascular remodeling in hypertension: Mechanisms and treatment. Hypertension 2012, 59, 367–374. [Google Scholar] [CrossRef]
- Forrester, S.J.; Booz, G.W.; Sigmund, C.D.; Coffman, T.M.; Kawai, T.; Rizzo, V.; Scalia, R.; Eguchi, S. Angiotensin II Signal Transduction: An Update on Mechanisms of Physiology and Pathophysiology. Physiol. Rev. 2018, 98, 1627–1738. [Google Scholar] [CrossRef]
- Mathiassen, O.N.; Buus, N.H.; Sihm, I.; Thybo, N.K.; Mørn, B.; Schroeder, A.P.; Thygesen, K.; Aalkjaer, C.; Lederballe, O.; Mulvany, M.J.; et al. Small artery structure is an independent predictor of cardiovascular events in essential hypertension. J. Hypertens. 2007, 25, 1021–1026. [Google Scholar] [CrossRef]
- Rizzoni, D.; Porteri, E.; Boari, G.E.; De Ciuceis, C.; Sleiman, I.; Muiesan, M.L.; Castellano, M.; Miclini, M.; Agabiti-Rosei, E. Prognostic Significance of Small-Artery Structure in Hypertension. Circulation 2003, 108, 2230–2235. [Google Scholar] [CrossRef]
- Lehoux, S. Adventures in the Adventitia. Hypertension 2016, 67, 836–838. [Google Scholar] [CrossRef]
- Ehret, G.B.; Munroe, P.B.; Rice, K.M.; Bochud, M.; Johnson, A.D.; Chasman, D.I.; Smith, A.V.; Tobin, M.D.; Verwoert, G.C.; Hwang, S.-J.; et al. Genetic variants in novel pathways influence blood pressure and cardiovascular disease risk. Nature 2011, 478, 103–109. [Google Scholar] [PubMed]
- Percie du Sert, N.; Hurst, V.; Ahluwalia, A.; Alam, S.; Avey, M.T.; Baker, M.; Browne, W.J.; Clark, A.; Cuthill, I.C.; Howells, D.W.; et al. The ARRIVE guidelines 2.0: Updated guidelines for reporting animal research. PLoS Biol. 2020, 18, e3000410. [Google Scholar]
- Muñoz-Valverde, D.; Rodríguez, P.R.; Gutierrez-Arzapalo, P.Y.; De Pablo, A.L.L.; González, M.C.; López-Giménez, R.; Somoza, B.; Arribas, S.M. Effect of Fetal Undernutrition and Postnatal Overfeeding on Rat Adipose Tissue and Organ Growth at Early Stages of Postnatal Development. Physiol. Res. 2015, 64, 547–559. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, C.L.; Morgan, P.E.; Davies, M.J. Quantification of protein modification by oxidants. Free Radic. Biol. Med. 2009, 46, 965–988. [Google Scholar] [CrossRef] [PubMed]
- Ferreira-Duarte, M.; Rodrigues-Pinto, T.; Sousa, T.; Faria, M.; Rocha, M.; Menezes-Pinto, D.; Esteves-Monteiro, M.; Magro, F.; Dias-Pereira, P.; Duarte-Araújo, M.; et al. Interaction between the Renin–Angiotensin System and Enteric Neurotransmission Contributes to Colonic Dysmotility in the TNBS-Induced Model of Colitis. Int. J. Mol. Sci. 2021, 22, 4836. [Google Scholar] [CrossRef]
- Rocha-Pereira, C.; Sousa, J.; Rocha, M.S.V.; Fresco, P.; Gonçalves, J.; Diniz, C. Differential inhibition of noradrenaline release mediated by inhibitory A1-adenosine receptors in the mesenteric vein and artery from normotensive and hypertensive rats. Neurochem. Int. 2013, 62, 399–405. [Google Scholar] [CrossRef]
- Arribas, S.M.; Hillier, C.; González, C.; McGrory, S.; Dominiczak, A.; McGrath, J. Cellular Aspects of Vascular Remodeling in Hypertension Revealed by Confocal Microscopy. Hypertension 1997, 30, 1455–1464. [Google Scholar] [CrossRef]
- Sousa, J.B.P.F.a.C.D. Imaging receptors with Laser Scanning Confocal Microscopy: Qualitative and quantitative analysis. In Microscopy: Advances in Scientific Research and Education; Méndez-Villas, A., Ed.; Formatex Research Center: Badajoz, Spain, 2014; pp. 201–208. ISBN 978-84-942134-3-4. [Google Scholar]
- Sabatino, L.; Costagli, C.; Lapi, D.; Del Seppia, C.; Federighi, G.; Balzan, S.; Colantuoni, A.; Iervasi, G.; Scuri, R. Renin-Angiotensin System Responds to Prolonged Hypotensive Effect Induced by Mandibular Extension in Spontaneously Hypertensive Rats. Front. Physiol. 2018, 9, 1613. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, 0034.1–0034.11. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibish, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Body Weight (g) | Systolic Pressure (mmHg) | Diastolic Pressure (mmHg) | Heart Rate (BPM) | N | |
|---|---|---|---|---|---|
| CONTROL (n = 7) | 486.6 ± 18.6 | 125.6 ± 5.1 | 68.7 ± 4.2 | 258 ± 8 | 6 |
| MUN (n = 7) | 460.18 ± 8.1 | 151.0 ± 3.9 * | 90.3 ± 4.6 * | 254 ± 10 | 6 |
| Gene | Primer Sequence (5′→3′) | Length (bp) | GenBank No. |
|---|---|---|---|
| Ren | F: CTAAAGCATCTCGCCAAGG | AB188298 | |
| R: ACCAGGGATGTGTCGAATGA | |||
| Ace | F: TCCTATTCCCGCTCATCT | 127 | NM_012544.1 |
| R: CCAGCCCTTCTGTACCATT | |||
| Ace 2 | F: GAATGCGACCATCAAGCG | 228 | AY881244 |
| R: CAAGCCCAGAGCCTACGA | |||
| Agtr1 | F: TCTGGATAAATCACACAACCCTC | 77 | NM_030985.4 |
| R: GAGTTGGTCTCAGACACTATTCG | |||
| Agtr2 | F: CTGGCAAGCATCTTATGTAGTTC | 115 | U22663.1 |
| R: ACAAGCATTCACACCTAAGTATTC | |||
| Mas1 | F: ACTGTCGGGCGGTCATCATC | 272 | NM_012757.2 |
| R: GGTGGAGAAAAGCAAGGAGA | |||
| Mrgprd | F: CAGCCTCGGCGGCTCTA | 293648 | |
| R: CCAACGGCAGAGAACAGGTAAG | |||
| Hprt-1 | F: CCCAGCGTCGTGATTAGTGATG | 110 | NM_012583 |
| R: TTCAGTCCTGTCCATAATCAGTCC | |||
| Hmbs | F: TCTAGATGGCTCAGATAGCATGCA | 76 | NM_013168 |
| R: TGGACCATCTTCTTGCTGAACA | |||
| Papbn-1 | F: TATGGTGCGACAGCAGAAGA | 110 | 116697 |
| R: TATGCAAACCCTTTGGGATG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vieira-Rocha, M.S.; Rodriguez-Rodriguez, P.; Ferreira-Duarte, M.; Faria, M.; Sousa, J.B.; Morato, M.; Arribas, S.M.; Diniz, C. Fetal Undernutrition Modifies Vascular RAS Balance Enhancing Oxidative Damage and Contributing to Remodeling. Int. J. Mol. Sci. 2022, 23, 1233. https://doi.org/10.3390/ijms23031233
Vieira-Rocha MS, Rodriguez-Rodriguez P, Ferreira-Duarte M, Faria M, Sousa JB, Morato M, Arribas SM, Diniz C. Fetal Undernutrition Modifies Vascular RAS Balance Enhancing Oxidative Damage and Contributing to Remodeling. International Journal of Molecular Sciences. 2022; 23(3):1233. https://doi.org/10.3390/ijms23031233
Chicago/Turabian StyleVieira-Rocha, Maria Sofia, Pilar Rodriguez-Rodriguez, Mariana Ferreira-Duarte, Miguel Faria, Joana Beatriz Sousa, Manuela Morato, Silvia Magdalena Arribas, and Carmen Diniz. 2022. "Fetal Undernutrition Modifies Vascular RAS Balance Enhancing Oxidative Damage and Contributing to Remodeling" International Journal of Molecular Sciences 23, no. 3: 1233. https://doi.org/10.3390/ijms23031233
APA StyleVieira-Rocha, M. S., Rodriguez-Rodriguez, P., Ferreira-Duarte, M., Faria, M., Sousa, J. B., Morato, M., Arribas, S. M., & Diniz, C. (2022). Fetal Undernutrition Modifies Vascular RAS Balance Enhancing Oxidative Damage and Contributing to Remodeling. International Journal of Molecular Sciences, 23(3), 1233. https://doi.org/10.3390/ijms23031233