
Molecular Characteristics of Cell Pyroptosis and Its Inhibitors: A Review of Activation, Regulation, and Inhibitors


Abstract
1. Introduction
2. Characteristics of Pyroptosis
2.1. Morphological Characteristics of Pyroptosis
2.2. Molecular Characteristics of Pyroptosis
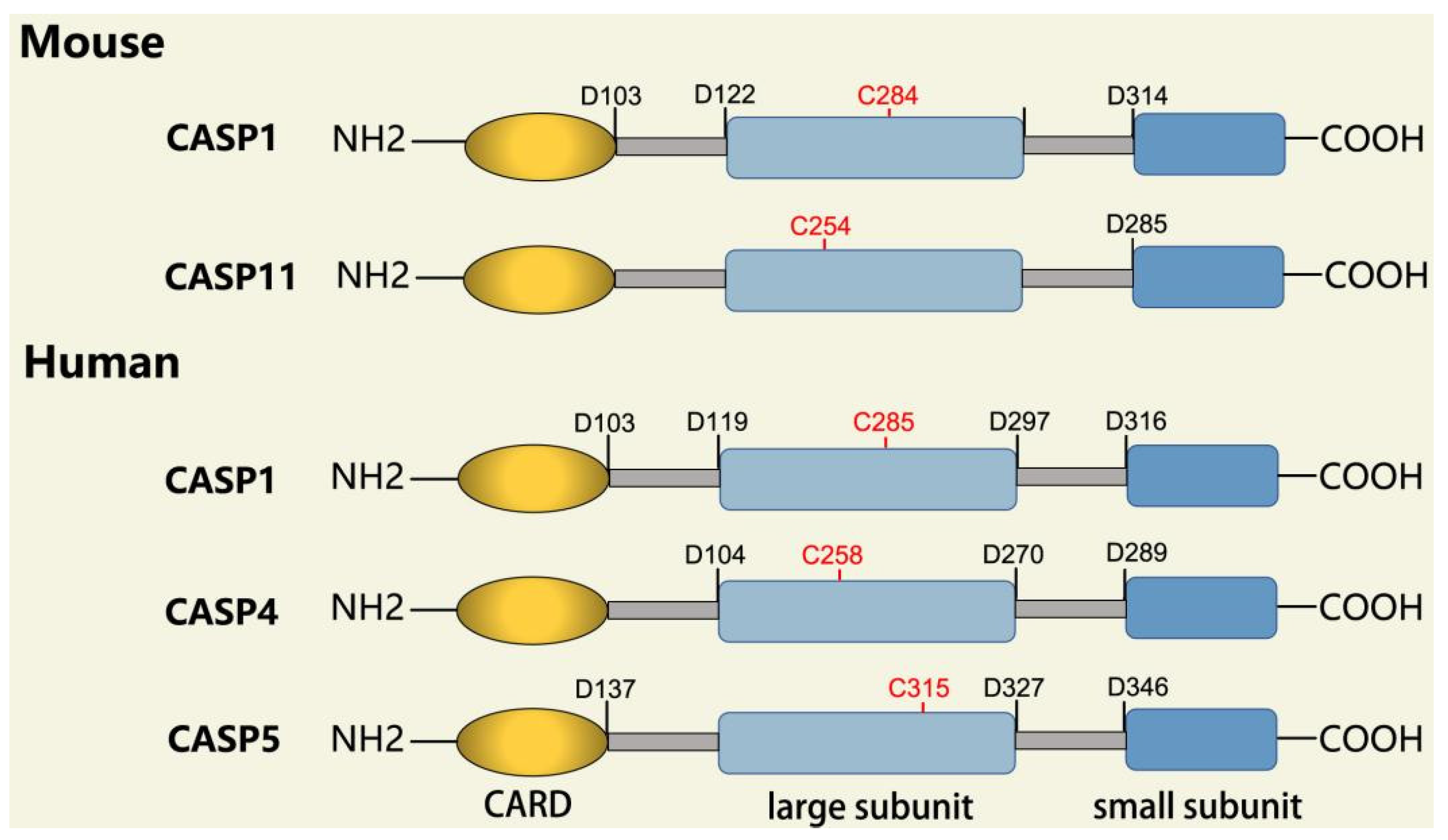
2.2.1. Inflammatory Caspases
2.2.2. Gasdermin Family
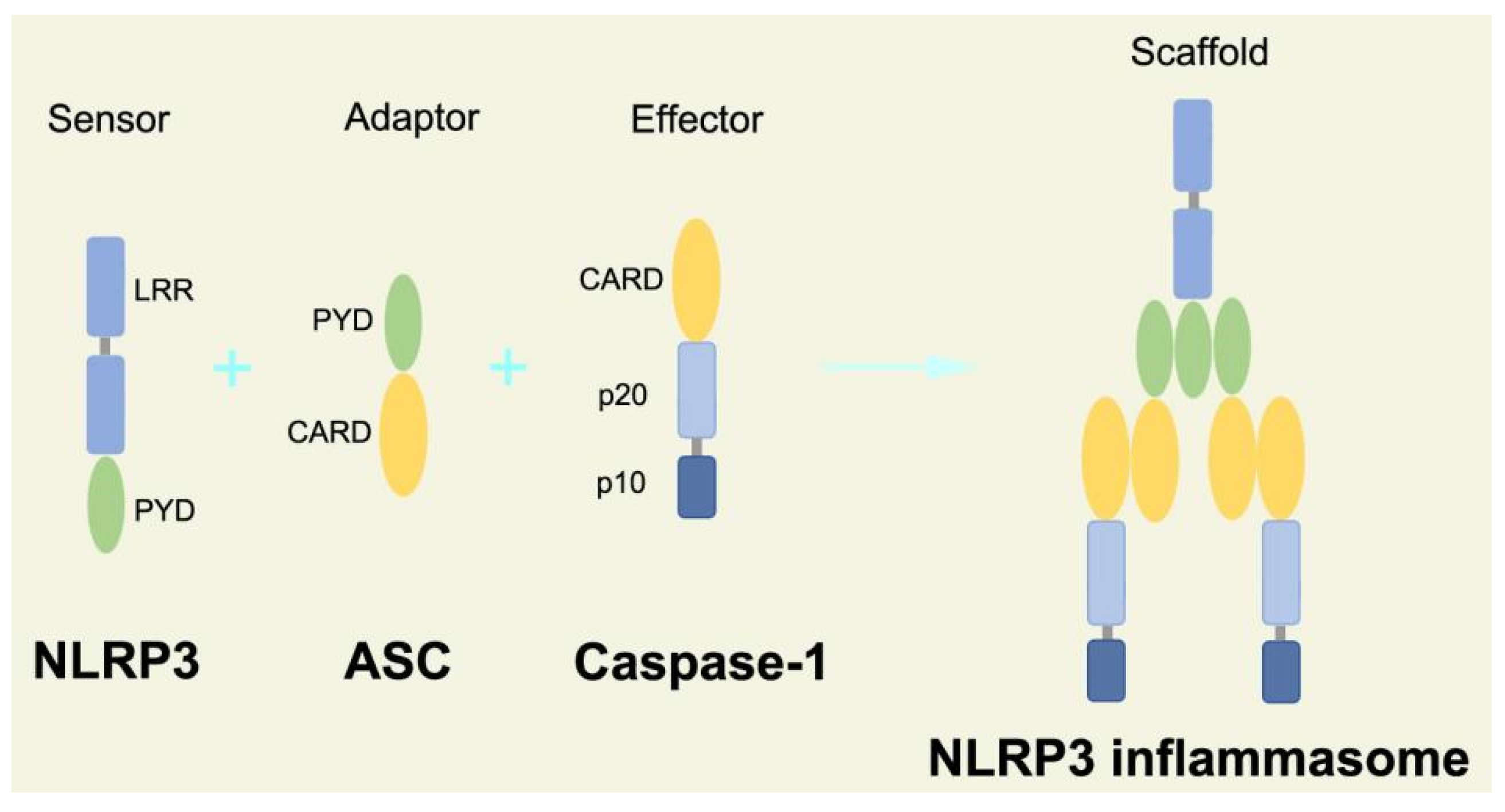
2.2.3. Canonical and Noncanonical Inflammasome
2.3. Mechanism of Pyroptosis
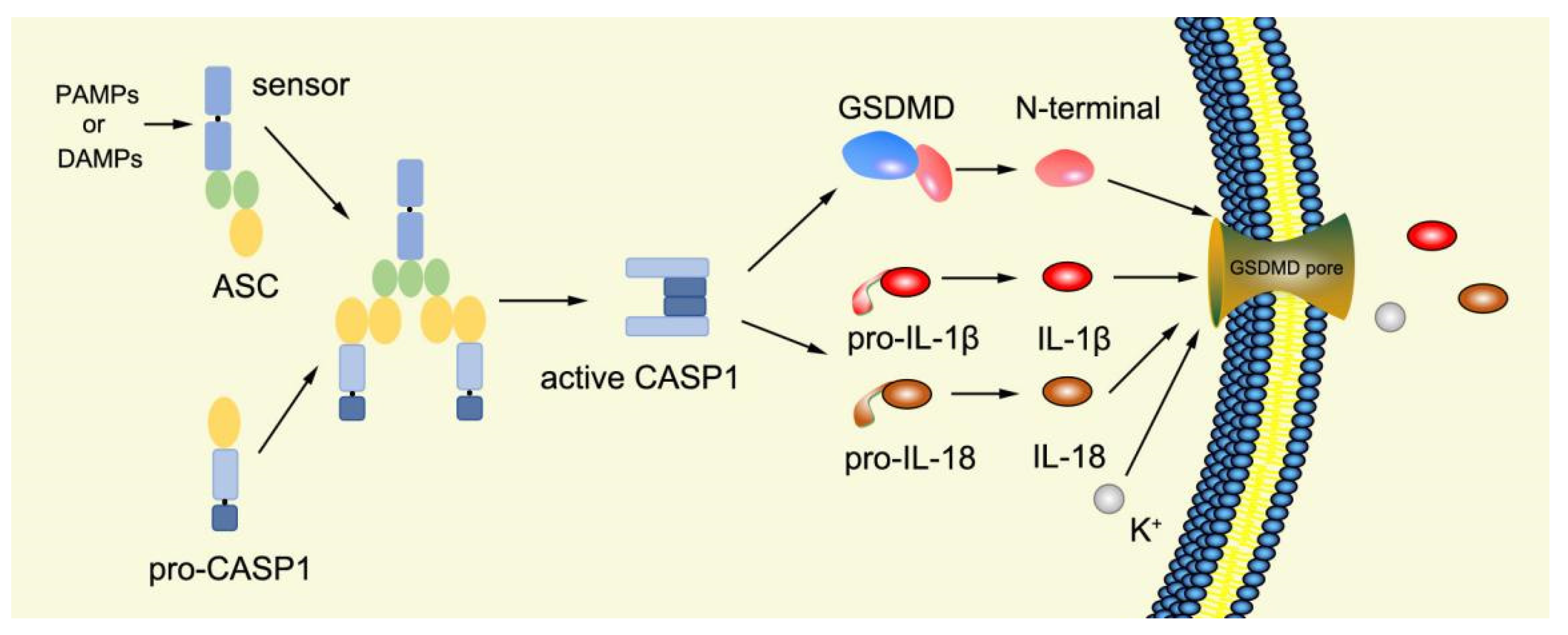
2.3.1. Canonical Pyroptosis Pathway
2.3.2. Noncanonical Pyroptosis Pathway
3. Inhibitors of Pyroptosis
3.1. Chemical Agents as Inhibitors of Pyroptosis
3.2. Natural Product as Inhibitor of Pyroptosis
3.3. Herb Medicine as Inhibitor of Pyroptosis
4. Conclusions and Future Perspectives
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Zychlinsky, A.; Prevost, M.C.; Sansonetti, P.J. Shigella flexneri induces apoptosis in infected macrophages. Nature 1992, 358, 167–169. [Google Scholar] [CrossRef] [PubMed]
- Cookson, B.T.; Brennan, M.A. Pro-inflammatory programmed cell death. Trends Microbiol. 2001, 9, 113–114. [Google Scholar] [CrossRef] [PubMed]
- Broz, P.; Pelegrin, P.; Shao, F. The gasdermins, a protein family executing cell death and inflammation. Nat. Rev. Immunol. 2020, 20, 143–157. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Lu, W.; Zhang, Y.; Zhang, G.; Shi, X.; Hisada, Y.; Grover, S.P.; Zhang, X.; Li, L.; Xiang, B.; et al. Inflammasome Activation Triggers Blood Clotting and Host Death through Pyroptosis. Immunity 2019, 50, 1401–1411.e4. [Google Scholar] [CrossRef] [PubMed]
- Broz, P.; Dixit, V.M. Inflammasomes: Mechanism of assembly, regulation and signalling. Nat. Rev. Immunol. 2016, 16, 407–420. [Google Scholar] [CrossRef]
- Rathinam, V.A.K.; Zhao, Y.; Shao, F. Innate immunity to intracellular LPS. Nat. Immunol. 2019, 20, 527–533. [Google Scholar] [CrossRef]
- Burdette, B.E.; Esparza, A.N.; Zhu, H.; Wang, S. Gasdermin D in pyroptosis. Acta Pharm. Sin. B 2021, 11, 2768–2782. [Google Scholar] [CrossRef]
- Shi, J.; Zhao, Y.; Wang, K.; Shi, X.; Wang, Y.; Huang, H.; Zhuang, Y.; Cai, T.; Wang, F.; Shao, F. Cleavage of GSDMD by inflammatory caspases determines pyroptotic cell death. Nature 2015, 526, 660–665. [Google Scholar] [CrossRef]
- Jorgensen, I.; Rayamajhi, M.; Miao, E.A. Programmed cell death as a defence against infection. Nat. Rev. Immunol. 2017, 17, 151–164. [Google Scholar] [CrossRef]
- Vanden Berghe, T.; Hassannia, B.; Vandenabeele, P. An outline of necrosome triggers. Cell. Mol. Life Sci. 2016, 73, 2137–2152. [Google Scholar] [CrossRef]
- Ge, X.; Li, W.; Huang, S.; Yin, Z.; Xu, X.; Chen, F.; Kong, X.; Wang, H.; Zhang, J.; Lei, P. The pathological role of NLRs and AIM2 inflammasome-mediated pyroptosis in damaged blood-brain barrier after traumatic brain injury. Brain. Res. 2018, 1697, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S. Caspase function in programmed cell death. Cell Death Differ. 2007, 14, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Eckhart, L.; Ballaun, C.; Hermann, M.; VandeBerg, J.L.; Sipos, W.; Uthman, A.; Fischer, H.; Tschachler, E. Identification of novel mammalian caspases reveals an important role of gene loss in shaping the human caspase repertoire. Mol. Biol. Evol. 2008, 25, 831–841. [Google Scholar] [CrossRef]
- Yin, Y.; Pastrana, J.L.; Li, X.; Huang, X.; Mallilankaraman, K.; Choi, E.T.; Madesh, M.; Wang, H.; Yang, X.F. Inflammasomes: Sensors of metabolic stresses for vascular inflammation. Front. Biosci. (Landmark Ed.) 2013, 18, 638–649. [Google Scholar] [CrossRef] [PubMed]
- Crowley, L.C.; Waterhouse, N.J. Detecting Cleaved Caspase-3 in Apoptotic Cells by Flow Cytometry. Cold Spring Harb. Protoc. 2016, 2016, pdb-prot087312. [Google Scholar] [CrossRef]
- Park, H.H. Structural features of caspase-activating complexes. Int. J. Mol. Sci. 2012, 13, 4807–4818. [Google Scholar] [CrossRef]
- Zhaolin, Z.; Guohua, L.; Shiyuan, W.; Zuo, W. Role of pyroptosis in cardiovascular disease. Cell Prolif. 2019, 52, e12563. [Google Scholar] [CrossRef]
- Black, R.A.; Kronheim, S.R.; Sleath, P.R. Activation of interleukin-1 beta by a co-induced protease. FEBS Lett. 1989, 247, 386–390. [Google Scholar] [CrossRef]
- Kostura, M.J.; Tocci, M.J.; Limjuco, G.; Chin, J.; Cameron, P.; Hillman, A.G.; Chartrain, N.A.; Schmidt, J.A. Identification of a monocyte specific pre-interleukin 1 beta convertase activity. Proc. Natl. Acad. Sci. USA 1989, 86, 5227–5231. [Google Scholar] [CrossRef]
- Cerretti, D.P.; Kozlosky, C.J.; Mosley, B.; Nelson, N.; Van Ness, K.; Greenstreet, T.A.; March, C.J.; Kronheim, S.R.; Druck, T.; Cannizzaro, L.A.; et al. Molecular cloning of the interleukin-1 beta converting enzyme. Science 1992, 256, 97–100. [Google Scholar] [CrossRef]
- Thornberry, N.A.; Bull, H.G.; Calaycay, J.R.; Chapman, K.T.; Howard, A.D.; Kostura, M.J.; Miller, D.K.; Molineaux, S.M.; Weidner, J.R.; Aunins, J.; et al. A novel heterodimeric cysteine protease is required for interleukin-1 beta processing in monocytes. Nature 1992, 356, 768–774. [Google Scholar] [CrossRef] [PubMed]
- Ghayur, T.; Banerjee, S.; Hugunin, M.; Butler, D.; Herzog, L.; Carter, A.; Quintal, L.; Sekut, L.; Talanian, R.; Paskind, M.; et al. Caspase-1 processes IFN-gamma-inducing factor and regulates LPS-induced IFN-gamma production. Nature 1997, 386, 619–623. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Kuida, K.; Tsutsui, H.; Ku, G.; Hsiao, K.; Fleming, M.A.; Hayashi, N.; Higashino, K.; Okamura, H.; Nakanishi, K.; et al. Activation of interferon-gamma inducing factor mediated by interleukin-1beta converting enzyme. Science 1997, 275, 206–209. [Google Scholar] [CrossRef] [PubMed]
- Fuentes-Prior, P.; Salvesen, G.S. The protein structures that shape caspase activity, specificity, activation and inhibition. Biochem. J. 2004, 384, 201–232. [Google Scholar] [CrossRef]
- Roschitzki-Voser, H.; Schroeder, T.; Lenherr, E.D.; Frolich, F.; Schweizer, A.; Donepudi, M.; Ganesan, R.; Mittl, P.R.; Baici, A.; Grutter, M.G. Human caspases in vitro: Expression, purification and kinetic characterization. Protein Expr. Purif. 2012, 84, 236–246. [Google Scholar] [CrossRef]
- Denault, J.B.; Salvesen, G.S. Human caspase-7 activity and regulation by its N-terminal peptide. J. Biol. Chem. 2003, 278, 34042–34050. [Google Scholar] [CrossRef]
- Vaidya, S.; Velazquez-Delgado, E.M.; Abbruzzese, G.; Hardy, J.A. Substrate-induced conformational changes occur in all cleaved forms of caspase-6. J. Mol. Biol. 2011, 406, 75–91. [Google Scholar] [CrossRef]
- Shi, Y. Mechanisms of caspase activation and inhibition during apoptosis. Mol. Cell 2002, 9, 459–470. [Google Scholar] [CrossRef]
- Shi, Y. Caspase activation: Revisiting the induced proximity model. Cell 2004, 117, 855–858. [Google Scholar] [CrossRef]
- Kersse, K.; Lamkanfi, M.; Bertrand, M.J.M.; Vanden Berghe, T.; Vandenabeele, P. Interaction patches of procaspase-1 caspase recruitment domains (CARDs) are differently involved in procaspase-1 activation and receptor-interacting protein 2 (RIP2)-dependent nuclear factor kappaB signaling. J. Biol. Chem. 2011, 286, 35874–35882. [Google Scholar] [CrossRef]
- Boucher, D.; Monteleone, M.; Coll, R.C.; Chen, K.W.; Ross, C.M.; Teo, J.L.; Gomez, G.A.; Holley, C.L.; Bierschenk, D.; Stacey, K.J.; et al. Caspase-1 self-cleavage is an intrinsic mechanism to terminate inflammasome activity. J. Exp. Med. 2018, 215, 827–840. [Google Scholar] [CrossRef] [PubMed]
- Ross, C.; Chan, A.H.; Von Pein, J.; Boucher, D.; Schroder, K. Dimerization and auto-processing induce caspase-11 protease activation within the non-canonical inflammasome. Life Sci. Alliance 2018, 1, e201800237. [Google Scholar] [CrossRef] [PubMed]
- Feeney, B.; Clark, A.C. Reassembly of active caspase-3 is facilitated by the propeptide. J. Biol. Chem. 2005, 280, 39772–39785. [Google Scholar] [CrossRef] [PubMed]
- Butt, A.J.; Harvey, N.L.; Parasivam, G.; Kumar, S. Dimerization and autoprocessing of the Nedd2 (caspase-2) precursor requires both the prodomain and the carboxyl-terminal regions. J. Biol. Chem. 1998, 273, 6763–6768. [Google Scholar] [CrossRef]
- Van Criekinge, W.; Beyaert, R.; Van de Craen, M.; Vandenabeele, P.; Schotte, P.; De Valck, D.; Fiers, W. Functional characterization of the prodomain of interleukin-1beta-converting enzyme. J. Biol. Chem. 1996, 271, 27245–27248. [Google Scholar] [CrossRef]
- Stennicke, H.R.; Deveraux, Q.L.; Humke, E.W.; Reed, J.C.; Dixit, V.M.; Salvesen, G.S. Caspase-9 can be activated without proteolytic processing. J. Biol. Chem. 1999, 274, 8359–8362. [Google Scholar] [CrossRef]
- Ponder, K.G.; Boise, L.H. The prodomain of caspase-3 regulates its own removal and caspase activation. Cell Death Discov. 2019, 5, 56. [Google Scholar] [CrossRef]
- Yang, X.; Chang, H.Y.; Baltimore, D. Autoproteolytic activation of pro-caspases by oligomerization. Mol. Cell 1998, 1, 319–325. [Google Scholar] [CrossRef]
- Elliott, J.M.; Rouge, L.; Wiesmann, C.; Scheer, J.M. Crystal structure of procaspase-1 zymogen domain reveals insight into inflammatory caspase autoactivation. J. Biol. Chem. 2009, 284, 6546–6553. [Google Scholar] [CrossRef]
- Wilson, K.P.; Black, J.A.; Thomson, J.A.; Kim, E.E.; Griffith, J.P.; Navia, M.A.; Murcko, M.A.; Chambers, S.P.; Aldape, R.A.; Raybuck, S.A.; et al. Structure and mechanism of interleukin-1 beta converting enzyme. Nature 1994, 370, 270–275. [Google Scholar] [CrossRef]
- Datta, D.; McClendon, C.L.; Jacobson, M.P.; Wells, J.A. Substrate and inhibitor-induced dimerization and cooperativity in caspase-1 but not caspase-3. J. Biol. Chem. 2013, 288, 9971–9981. [Google Scholar] [CrossRef] [PubMed]
- Martinon, F.; Mayor, A.; Tschopp, J. The inflammasomes: Guardians of the body. Annu. Rev. Immunol. 2009, 27, 229–265. [Google Scholar] [CrossRef] [PubMed]
- Riedl, S.J.; Shi, Y. Molecular mechanisms of caspase regulation during apoptosis. Nat. Rev. Mol. Cell Biol. 2004, 5, 897–907. [Google Scholar] [CrossRef] [PubMed]
- Renatus, M.; Stennicke, H.R.; Scott, F.L.; Liddington, R.C.; Salvesen, G.S. Dimer formation drives the activation of the cell death protease caspase 9. Proc. Natl. Acad. Sci. USA 2001, 98, 14250–14255. [Google Scholar] [CrossRef] [PubMed]
- Salvesen, G.S.; Dixit, V.M. Caspase activation: The induced-proximity model. Proc. Natl. Acad. Sci. USA 1999, 96, 10964–10967. [Google Scholar] [CrossRef]
- Boatright, K.M.; Salvesen, G.S. Mechanisms of caspase activation. Curr. Opin. Cell Biol. 2003, 15, 725–731. [Google Scholar] [CrossRef]
- Kumar, S. Mechanisms mediating caspase activation in cell death. Cell Death Differ. 1999, 6, 1060–1066. [Google Scholar] [CrossRef]
- Shi, J.; Zhao, Y.; Wang, Y.; Gao, W.; Ding, J.; Li, P.; Hu, L.; Shao, F. Inflammatory caspases are innate immune receptors for intracellular LPS. Nature 2014, 514, 187–192. [Google Scholar] [CrossRef]
- Hagar, J.A.; Powell, D.A.; Aachoui, Y.; Ernst, R.K.; Miao, E.A. Cytoplasmic LPS activates caspase-11: Implications in TLR4-independent endotoxic shock. Science 2013, 341, 1250–1253. [Google Scholar] [CrossRef]
- Agnew, A.; Nulty, C.; Creagh, E.M. Regulation, Activation and Function of Caspase-11 during Health and Disease. Int. J. Mol. Sci. 2021, 22, 1506. [Google Scholar] [CrossRef]
- Broz, P.; von Moltke, J.; Jones, J.W.; Vance, R.E.; Monack, D.M. Differential requirement for Caspase-1 autoproteolysis in pathogen-induced cell death and cytokine processing. Cell Host Microbe 2010, 8, 471–483. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Xia, S.; Zhang, Z.; Wu, H.; Lieberman, J. Channelling inflammation: Gasdermins in physiology and disease. Nat. Rev. Drug Discov. 2021, 20, 384–405. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.; Fox, D.; Man, S.M. Mechanisms of Gasdermin Family Members in Inflammasome Signaling and Cell Death. J. Mol. Biol. 2018, 430 (Pt B), 3068–3080. [Google Scholar] [CrossRef]
- Man, S.M.; Kanneganti, T.D. Gasdermin D: The long-awaited executioner of pyroptosis. Cell Res. 2015, 25, 1183–1184. [Google Scholar] [CrossRef] [PubMed]
- England, H.; Summersgill, H.R.; Edye, M.E.; Rothwell, N.J.; Brough, D. Release of interleukin-1alpha or interleukin-1beta depends on mechanism of cell death. J. Biol. Chem. 2014, 289, 15942–15950. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Wang, C.; Yang, J.; Zhou, B.; Yang, R.; Ramachandran, R.; Abbott, D.W.; Xiao, T.S. Crystal Structures of the Full-Length Murine and Human Gasdermin D Reveal Mechanisms of Autoinhibition, Lipid Binding, and Oligomerization. Immunity 2019, 51, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Sborgi, L.; Ruhl, S.; Mulvihill, E.; Pipercevic, J.; Heilig, R.; Stahlberg, H.; Farady, C.J.; Muller, D.J.; Broz, P.; Hiller, S. GSDMD membrane pore formation constitutes the mechanism of pyroptotic cell death. EMBO J. 2016, 35, 1766–1778. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, Z.; Ruan, J.; Pan, Y.; Magupalli, V.G.; Wu, H.; Lieberman, J. Inflammasome-activated gasdermin D causes pyroptosis by forming membrane pores. Nature 2016, 535, 153–158. [Google Scholar] [CrossRef]
- Ding, J.; Wang, K.; Liu, W.; She, Y.; Sun, Q.; Shi, J.; Sun, H.; Wang, D.C.; Shao, F. Pore-forming activity and structural autoinhibition of the gasdermin family. Nature 2016, 535, 111–116. [Google Scholar] [CrossRef]
- Ding, J.; Shao, F. Growing a Gasdermin Pore in Membranes of Pyroptotic Cells. EMBO J. 2018, 37, e100067. [Google Scholar] [CrossRef]
- Liu, Z.; Wang, C.; Rathkey, J.K.; Yang, J.; Dubyak, G.R.; Abbott, D.W.; Xiao, T.S. Structures of the Gasdermin D C-Terminal Domains Reveal Mechanisms of Autoinhibition. Structure 2018, 26, 778–784. [Google Scholar] [CrossRef]
- Xia, S. Biological Mechanisms and Therapeutic Relevance of the Gasdermin Family. Mol. Aspects Med. 2020, 76, 100890. [Google Scholar] [CrossRef] [PubMed]
- Mulvihill, E.; Sborgi, L.; Mari, S.A.; Pfreundschuh, M.; Hiller, S.; Muller, D.J. Mechanism of Membrane Pore Formation by Human Gasdermin-D. EMBO J. 2018, 37, e98321. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Shi, J.; Shao, F. Inflammatory Caspases: Activation and Cleavage of Gasdermin-D in Vitro and During Pyroptosis. Methods Mol. Biol. 2018, 1714, 131–148. [Google Scholar] [CrossRef] [PubMed]
- von Moltke, J.; Ayres, J.S.; Kofoed, E.M.; Chavarria-Smith, J.; Vance, R.E. Recognition of bacteria by inflammasomes. Annu. Rev. Immunol. 2013, 31, 73–106. [Google Scholar] [CrossRef] [PubMed]
- Latz, E. The inflammasomes: Mechanisms of activation and function. Curr. Opin. Immunol. 2010, 22, 28–33. [Google Scholar] [CrossRef] [PubMed]
- Lamkanfi, M.; Dixit, V.M. Inflammasomes and their roles in health and disease. Annu. Rev. Cell Dev. Biol. 2012, 28, 137–161. [Google Scholar] [CrossRef]
- Skeldon, A.; Saleh, M. The Inflammasomes: Molecular Effectors of Host Resistance against Bacterial, Viral, Parasitic, and Fungal Infections. Front. Microbiol. 2011, 2, 15. [Google Scholar] [CrossRef]
- Sun, C.; Dong, J.; Kuang, Z.; Yin, M.; Liu, X.; Deng, H. Research progresses on NLRP3 inflammasomes-induced anti-tumor immunity. Acta Pharm. Sin. 2022, 9, 2612–2621. (In Chinese) [Google Scholar] [CrossRef]
- Ting, J.P.; Lovering, R.C.; Alnemri, E.S.; Bertin, J.; Boss, J.M.; Davis, B.K.; Flavell, R.A.; Girardin, S.E.; Godzik, A.; Harton, J.A.; et al. The NLR gene family: A standard nomenclature. Immunity 2008, 28, 285–287. [Google Scholar] [CrossRef]
- Lu, A.; Magupalli, V.G.; Ruan, J.; Yin, Q.; Atianand, M.K.; Vos, M.R.; Schroder, G.F.; Fitzgerald, K.A.; Wu, H.; Egelman, E.H. Unified polymerization mechanism for the assembly of ASC-dependent inflammasomes. Cell 2014, 156, 1193–1206. [Google Scholar] [CrossRef] [PubMed]
- Martinon, F.; Burns, K.; Tschopp, J. The inflammasome: A molecular platform triggering activation of inflammatory caspases and processing of proIL-beta. Mol. Cell 2002, 10, 417–426. [Google Scholar] [CrossRef] [PubMed]
- Agostini, L.; Martinon, F.; Burns, K.; McDermott, M.F.; Hawkins, P.N.; Tschopp, J. NALP3 forms an IL-1beta-processing inflammasome with increased activity in Muckle-Wells autoinflammatory disorder. Immunity 2004, 20, 319–325. [Google Scholar] [CrossRef] [PubMed]
- Bauernfeind, F.G.; Horvath, G.; Stutz, A.; Alnemri, E.S.; MacDonald, K.; Speert, D.; Fernandes-Alnemri, T.; Wu, J.; Monks, B.G.; Fitzgerald, K.A.; et al. Cutting edge: NF-kappaB activating pattern recognition and cytokine receptors license NLRP3 inflammasome activation by regulating NLRP3 expression. J. Immunol. 2009, 183, 787–791. [Google Scholar] [CrossRef]
- Biasizzo, M.; Kopitar-Jerala, N. Interplay between Nlrp3 Inflammasome and Autophagy. Front. Immunol. 2020, 11, 591803. [Google Scholar] [CrossRef]
- Garlanda, C.; Dinarello, C.A.; Mantovani, A. The interleukin-1 family: Back to the future. Immunity 2013, 39, 1003–1018. [Google Scholar] [CrossRef]
- Yang, D.; He, Y.; Munoz-Planillo, R.; Liu, Q.; Nunez, G. Caspase-11 Requires the Pannexin-1 Channel and the Purinergic P2X7 Pore to Mediate Pyroptosis and Endotoxic Shock. Immunity 2015, 43, 923–932. [Google Scholar] [CrossRef]
- He, Y.; Zeng, M.Y.; Yang, D.; Motro, B.; Nunez, G. NEK7 is an essential mediator of NLRP3 activation downstream of potassium efflux. Nature 2016, 530, 354–357. [Google Scholar] [CrossRef]
- He, W.T.; Wan, H.; Hu, L.; Chen, P.; Wang, X.; Huang, Z.; Yang, Z.H.; Zhong, C.Q.; Han, J. Gasdermin D is an executor of pyroptosis and required for interleukin-1beta secretion. Cell Res. 2015, 25, 1285–1298. [Google Scholar] [CrossRef]
- Chen, X.; He, W.T.; Hu, L.; Li, J.; Fang, Y.; Wang, X.; Xu, X.; Wang, Z.; Huang, K.; Han, J. Pyroptosis is driven by non-selective gasdermin-D pore and its morphology is different from MLKL channel-mediated necroptosis. Cell Res. 2016, 26, 1007–1020. [Google Scholar] [CrossRef]
- Christgen, S.; Place, D.E.; Kanneganti, T.D. Toward targeting inflammasomes: Insights into their regulation and activation. Cell Res. 2020, 30, 315–327. [Google Scholar] [CrossRef] [PubMed]
- Case, C.L.; Roy, C.R. Asc modulates the function of NLRC4 in response to infection of macrophages by Legionella pneumophila. mBio 2011, 2, e00117-11. [Google Scholar] [CrossRef] [PubMed]
- Kayagaki, N.; Warming, S.; Lamkanfi, M.; Vande Walle, L.; Louie, S.; Dong, J.; Newton, K.; Qu, Y.; Liu, J.; Heldens, S.; et al. Non-canonical inflammasome activation targets caspase-11. Nature 2011, 479, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Jiang, S.; Zhang, Y.; Li, P.; Wang, K. The Multifaceted Roles of Pyroptotic Cell Death Pathways in Cancer. Cancers 2019, 11, 1313. [Google Scholar] [CrossRef]
- Shi, J.; Gao, W.; Shao, F. Pyroptosis: Gasdermin-Mediated Programmed Necrotic Cell Death. Trends Biochem. Sci. 2017, 42, 245–254. [Google Scholar] [CrossRef]
- He, Y.; Hara, H.; Nunez, G. Mechanism and Regulation of NLRP3 Inflammasome Activation. Trends Biochem. Sci. 2016, 41, 1012–1021. [Google Scholar] [CrossRef]
- Chen, Q.; Shi, P.; Wang, Y.; Zou, D.; Wu, X.; Wang, D.; Hu, Q.; Zou, Y.; Huang, Z.; Ren, J.; et al. GSDMB promotes non-canonical pyroptosis by enhancing caspase-4 activity. J. Mol. Cell Biol. 2019, 11, 496–508. [Google Scholar] [CrossRef]
- Hsu, J.L.; Chou, J.W.; Chen, T.F.; Hsu, J.T.; Su, F.Y.; Lan, J.L.; Wu, P.C.; Hu, C.M.; Lee, E.Y.; Lee, W.H. Glutathione peroxidase 8 negatively regulates caspase-4/11 to protect against colitis. EMBO Mol. Med. 2020, 12, e9386. [Google Scholar] [CrossRef]
- Yang, J.; Zhao, Y.; Shao, F. Non-canonical activation of inflammatory caspases by cytosolic LPS in innate immunity. Curr. Opin. Immunol. 2015, 32, 78–83. [Google Scholar] [CrossRef]
- Kayagaki, N.; Stowe, I.B.; Lee, B.L.; O’Rourke, K.; Anderson, K.; Warming, S.; Cuellar, T.; Haley, B.; Roose-Girma, M.; Phung, Q.T.; et al. Caspase-11 cleaves gasdermin D for non-canonical inflammasome signalling. Nature 2015, 526, 666–671. [Google Scholar] [CrossRef]
- Sarhan, J.; Liu, B.C.; Muendlein, H.I.; Li, P.; Nilson, R.; Tang, A.Y.; Rongvaux, A.; Bunnell, S.C.; Shao, F.; Green, D.R.; et al. Caspase-8 induces cleavage of gasdermin D to elicit pyroptosis during Yersinia infection. Proc. Natl. Acad. Sci. USA 2018, 115, E10888–E10897. [Google Scholar] [CrossRef] [PubMed]
- Ruhl, S.; Broz, P. Caspase-11 activates a canonical NLRP3 inflammasome by promoting K(+) efflux. Eur. J. Immunol. 2015, 45, 2927–2936. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Xu, W.; Zhou, R. NLRP3 inflammasome activation and cell death. Cell. Mol. Immunol. 2021, 18, 2114–2127. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Chen, Z.M.; Wu, X.; Zhang, L.; Cao, Y.; Zhou, P. Distinct Molecular Mechanisms Underlying Potassium Efflux for NLRP3 Inflammasome Activation. Front. Immunol. 2020, 11, 609441. [Google Scholar] [CrossRef]
- Martinon, F.; Tschopp, J. Inflammatory caspases: Linking an intracellular innate immune system to autoinflammatory diseases. Cell 2004, 117, 561–574. [Google Scholar] [CrossRef]
- Lyashchenko, A.K.; Cremers, S. On precision dosing of oral small molecule drugs in oncology. Br. J. Clin. Pharmacol. 2021, 87, 263–270. [Google Scholar] [CrossRef]
- Jiang, H.; He, H.; Chen, Y.; Huang, W.; Cheng, J.; Ye, J.; Wang, A.; Tao, J.; Wang, C.; Liu, Q.; et al. Identification of a selective and direct NLRP3 inhibitor to treat inflammatory disorders. J. Exp. Med. 2017, 214, 3219–3238. [Google Scholar] [CrossRef]
- Zhao, N.; Zhuo, X.; Lu, Y.; Dong, Y.; Ahmed, M.E.; Tucker, D.; Scott, E.L.; Zhang, Q. Intranasal Delivery of a Caspase-1 Inhibitor in the Treatment of Global Cerebral Ischemia. Mol. Neurobiol. 2017, 54, 4936–4952. [Google Scholar] [CrossRef]
- Kumar, A.; Negi, G.; Sharma, S.S. Suppression of NF-kappaB and NF-kappaB regulated oxidative stress and neuroinflammation by BAY 11-7082 (IkappaB phosphorylation inhibitor) in experimental diabetic neuropathy. Biochimie 2012, 94, 1158–1165. [Google Scholar] [CrossRef]
- Gao, R.; Shi, H.; Chang, S.; Gao, Y.; Li, X.; Lv, C.; Yang, H.; Xiang, H.; Yang, J.; Xu, L.; et al. The Selective Nlrp3-Inflammasome Inhibitor Mcc950 Reduces Myocardial Fibrosis and Improves Cardiac Remodeling in a Mouse Model of Myocardial Infarction. Int. Immunopharmacol. 2019, 74, 105575. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, H.; Kouadir, M.; Song, H.; Shi, F. Recent Advances in the Mechanisms of Nlrp3 Inflammasome Activation and Its Inhibitors. Cell Death Dis. 2019, 10, 128. [Google Scholar] [CrossRef] [PubMed]
- Marchetti, C.; Swartzwelter, B.; Gamboni, F.; Neff, C.P.; Richter, K.; Azam, T.; Carta, S.; Tengesdal, I.; Nemkov, T.; D’Alessandro, A.; et al. OLT1177, a beta-sulfonyl nitrile compound, safe in humans, inhibits the NLRP3 inflammasome and reverses the metabolic cost of inflammation. Proc. Natl. Acad. Sci. USA 2018, 115, E1530–E1539. [Google Scholar] [CrossRef] [PubMed]
- Laliberte, R.E.; Perregaux, D.G.; Hoth, L.R.; Rosner, P.J.; Jordan, C.K.; Peese, K.M.; Eggler, J.F.; Dombroski, M.A.; Geoghegan, K.F.; Gabel, C.A. Glutathione s-transferase omega 1-1 is a target of cytokine release inhibitory drugs and may be responsible for their effect on interleukin-1beta posttranslational processing. J. Biol. Chem. 2003, 278, 16567–16578. [Google Scholar] [CrossRef] [PubMed]
- Perregaux, D.G.; McNiff, P.; Laliberte, R.; Hawryluk, N.; Peurano, H.; Stam, E.; Eggler, J.; Griffiths, R.; Dombroski, M.A.; Gabel, C.A. Identification and characterization of a novel class of interleukin-1 post-translational processing inhibitors. J. Pharmacol. Exp. Ther. 2001, 299, 187–197. [Google Scholar] [PubMed]
- van Hout, G.P.; Bosch, L.; Ellenbroek, G.H.; de Haan, J.J.; van Solinge, W.W.; Cooper, M.A.; Arslan, F.; de Jager, S.C.; Robertson, A.A.; Pasterkamp, G.; et al. The selective NLRP3-inflammasome inhibitor MCC950 reduces infarct size and preserves cardiac function in a pig model of myocardial infarction. Eur. Heart J. 2017, 38, 828–836. [Google Scholar] [CrossRef]
- Hill, J.R.; Coll, R.C.; Sue, N.; Reid, J.C.; Dou, J.; Holley, C.L.; Pelingon, R.; Dickinson, J.B.; Biden, T.J.; Schroder, K.; et al. Sulfonylureas as Concomitant Insulin Secretagogues and NLRP3 Inflammasome Inhibitors. ChemMedChem 2017, 12, 1449–1457. [Google Scholar] [CrossRef]
- Lamkanfi, M.; Mueller, J.L.; Vitari, A.C.; Misaghi, S.; Fedorova, A.; Deshayes, K.; Lee, W.P.; Hoffman, H.M.; Dixit, V.M. Glyburide inhibits the Cryopyrin/Nalp3 inflammasome. J. Cell Biol. 2009, 187, 61–70. [Google Scholar] [CrossRef]
- Ye, Y.; Bajaj, M.; Yang, H.C.; Perez-Polo, J.R.; Birnbaum, Y. SGLT-2 Inhibition with Dapagliflozin Reduces the Activation of the Nlrp3/ASC Inflammasome and Attenuates the Development of Diabetic Cardiomyopathy in Mice with Type 2 Diabetes. Further Augmentation of the Effects with Saxagliptin, a DPP4 Inhibitor. Cardiovasc. Drugs Ther. 2017, 31, 119–132. [Google Scholar] [CrossRef]
- Mastrocola, R.; Penna, C.; Tullio, F.; Femmino, S.; Nigro, D.; Chiazza, F.; Serpe, L.; Collotta, D.; Alloatti, G.; Cocco, M.; et al. Pharmacological Inhibition of Nlrp3 Inflammasome Attenuates Myocardial Ischemia/Reperfusion Injury by Activation of Risk and Mitochondrial Pathways. Oxid. Med. Cell Longev. 2016, 2016, 5271251. [Google Scholar] [CrossRef]
- Shi, Y.; Lv, Q.; Zheng, M.; Sun, H.; Shi, F. NLRP3 inflammasome inhibitor INF39 attenuated NLRP3 assembly in macrophages. Int. Immunopharmacol. 2021, 92, 107358. [Google Scholar] [CrossRef]
- Liao, K.C.; Sandall, C.F.; Carlson, D.A.; Ulke-Lemee, A.; Platnich, J.M.; Hughes, P.F.; Muruve, D.A.; Haystead, T.A.J.; MacDonald, J.A. Application of immobilized ATP to the study of NLRP inflammasomes. Arch. Biochem. Biophys. 2019, 670, 104–115. [Google Scholar] [CrossRef] [PubMed]
- Cocco, M.; Pellegrini, C.; Martinez-Banaclocha, H.; Giorgis, M.; Marini, E.; Costale, A.; Miglio, G.; Fornai, M.; Antonioli, L.; Lopez-Castejon, G.; et al. Development of an Acrylate Derivative Targeting the NLRP3 Inflammasome for the Treatment of Inflammatory Bowel Disease. J. Med. Chem. 2017, 60, 3656–3671. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Jiang, H.; Chen, Y.; Wang, X.; Yang, Y.; Tao, J.; Deng, X.; Liang, G.; Zhang, H.; Jiang, W.; et al. Tranilast directly targets NLRP3 to treat inflammasome-driven diseases. EMBO Mol. Med. 2018, 10, e8689. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; He, H.; Lin, B.; Chen, Y.; Deng, X.; Jiang, W.; Zhou, R. RRx-001 ameliorates inflammatory diseases by acting as a potent covalent NLRP3 inhibitor. Cell Mol. Immunol. 2021, 18, 1425–1436. [Google Scholar] [CrossRef]
- Guo, C.; Fulp, J.W.; Jiang, Y.; Li, X.; Chojnacki, J.E.; Wu, J.; Wang, X.Y.; Zhang, S. Development and Characterization of a Hydroxyl-Sulfonamide Analogue, 5-Chloro-N-[2-(4-hydroxysulfamoyl-phenyl)-ethyl]-2-methoxy-benzamide, as a Novel NLRP3 Inflammasome Inhibitor for Potential Treatment of Multiple Sclerosis. ACS Chem. Neurosci. 2017, 8, 2194–2201. [Google Scholar] [CrossRef]
- Juliana, C.; Fernandes-Alnemri, T.; Wu, J.; Datta, P.; Solorzano, L.; Yu, J.W.; Meng, R.; Quong, A.A.; Latz, E.; Scott, C.P.; et al. Anti-inflammatory compounds parthenolide and Bay 11-7082 are direct inhibitors of the inflammasome. J. Biol. Chem. 2010, 285, 9792–9802. [Google Scholar] [CrossRef]
- Shim, D.W.; Shin, W.Y.; Yu, S.H.; Kim, B.H.; Ye, S.K.; Koppula, S.; Won, H.S.; Kang, T.B.; Lee, K.H. BOT-4-one attenuates NLRP3 inflammasome activation: NLRP3 alkylation leading to the regulation of its ATPase activity and ubiquitination. Sci. Rep. 2017, 7, 15020. [Google Scholar] [CrossRef]
- He, Y.; Varadarajan, S.; Munoz-Planillo, R.; Burberry, A.; Nakamura, Y.; Nunez, G. 3,4-methylenedioxy-beta-nitrostyrene inhibits NLRP3 inflammasome activation by blocking assembly of the inflammasome. J. Biol. Chem. 2014, 289, 1142–1150. [Google Scholar] [CrossRef]
- Wannamaker, W.; Davies, R.; Namchuk, M.; Pollard, J.; Ford, P.; Ku, G.; Decker, C.; Charifson, P.; Weber, P.; Germann, U.A.; et al. (S)-1-((S)-2-{[1-(4-amino-3-chloro-phenyl)-methanoyl]-amino}-3,3-dimethyl-butanoy l)-pyrrolidine-2-carboxylic acid ((2R,3S)-2-ethoxy-5-oxo-tetrahydro-furan-3-yl)-amide (VX-765), an orally available selective interleukin (IL)-converting enzyme/caspase-1 inhibitor, exhibits potent anti-inflammatory activities by inhibiting the release of IL-1beta and IL-18. J. Pharmacol. Exp. Ther. 2007, 321, 509–516. [Google Scholar] [CrossRef]
- Li, Y.; Niu, X.; Xu, H.; Li, Q.; Meng, L.; He, M.; Zhang, J.; Zhang, Z.; Zhang, Z. Vx-765 Attenuates Atherosclerosis in Apoe Deficient Mice by Modulating Vsmcs Pyroptosis. Exp. Cell Res. 2020, 389, 111847. [Google Scholar] [CrossRef]
- Mariani, S.M.; Matiba, B.; Armandola, E.A.; Krammer, P.H. Interleukin 1 beta-converting enzyme related proteases/caspases are involved in TRAIL-induced apoptosis of myeloma and leukemia cells. J. Cell Biol. 1997, 137, 221–229. [Google Scholar] [CrossRef]
- Rathkey, J.K.; Zhao, J.; Liu, Z.; Chen, Y.; Yang, J.; Kondolf, H.C.; Benson, B.L.; Chirieleison, S.M.; Huang, A.Y.; Dubyak, G.R.; et al. Chemical disruption of the pyroptotic pore-forming protein gasdermin D inhibits inflammatory cell death and sepsis. Sci. Immunol. 2018, 3, eaat2738. [Google Scholar] [CrossRef] [PubMed]
- Sollberger, G.; Choidas, A.; Burn, G.L.; Habenberger, P.; Di Lucrezia, R.; Kordes, S.; Menninger, S.; Eickhoff, J.; Nussbaumer, P.; Klebl, B.; et al. Gasdermin D plays a vital role in the generation of neutrophil extracellular traps. Sci. Immunol. 2018, 3, eaar6689. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.J.; Liu, X.; Xia, S.; Zhang, Z.; Zhang, Y.; Zhao, J.; Ruan, J.; Luo, X.; Lou, X.; Bai, Y.; et al. FDA-approved disulfiram inhibits pyroptosis by blocking gasdermin D pore formation. Nat. Immunol. 2020, 21, 736–745. [Google Scholar] [CrossRef] [PubMed]
- Humphries, F.; Shmuel-Galia, L.; Ketelut-Carneiro, N.; Li, S.; Wang, B.; Nemmara, V.V.; Wilson, R.; Jiang, Z.; Khalighinejad, F.; Muneeruddin, K.; et al. Succination inactivates gasdermin D and blocks pyroptosis. Science 2020, 369, 1633–1637. [Google Scholar] [CrossRef] [PubMed]
- Ye, A.; Li, W.; Zhou, L.; Ao, L.; Fang, W.; Li, Y. Targeting pyroptosis to regulate ischemic stroke injury: Molecular mechanisms and preclinical evidences. Brain Res. Bull. 2020, 165, 146–160. [Google Scholar] [CrossRef]
- Yan, L.; Li, C.; Xu, L.; Jin, Y.; Bai, W.; Ou, D.; He, X. Inhibitory effects of baicalin on NLRP3 inflammasome activation and pyroptosis and the underlying mechanism. Immunol. J. 2018, 2, 93–100. (In Chinese) [Google Scholar] [CrossRef]
- Hu, Q.; Zhang, T.; Yi, L.; Zhou, X.; Mi, M. Dihydromyricetin inhibits NLRP3 inflammasome-dependent pyroptosis by activating the Nrf2 signaling pathway in vascular endothelial cells. Biofactors 2018, 44, 123–136. [Google Scholar] [CrossRef]
- Martin-Sanchez, F.; Diamond, C.; Zeitler, M.; Gomez, A.I.; Baroja-Mazo, A.; Bagnall, J.; Spiller, D.; White, M.; Daniels, M.J.; Mortellaro, A.; et al. Inflammasome-dependent IL-1beta release depends upon membrane permeabilisation. Cell Death Differ. 2016, 23, 1219–1231. [Google Scholar] [CrossRef]
- Evavold, C.L.; Hafner-Bratkovic, I.; Devant, P.; D’Andrea, J.M.; Ngwa, E.M.; Borsic, E.; Doench, J.G.; LaFleur, M.W.; Sharpe, A.H.; Thiagarajah, J.R.; et al. Control of gasdermin D oligomerization and pyroptosis by the Ragulator-Rag-mTORC1 pathway. Cell 2021, 184, 4495–4511. [Google Scholar] [CrossRef]
- An, X.; Zhang, Y.; Cao, Y.; Chen, J.; Qin, H.; Yang, L. Punicalagin Protects Diabetic Nephropathy by Inhibiting Pyroptosis Based on Txnip/Nlrp3 Pathway. Nutrients 2020, 12, 1516. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Yang, Y.; Lv, Z.; Shu, A.; Du, Q.; Wang, W.; Chen, Y.; Xu, H. Study on the Inhibitive Effect of Catalpol on Diabetic Nephropathy. Life Sci. 2020, 257, 118120. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Chen, Y.; Li, Y.; Huang, M.; Zhao, W. Geniposide Alleviates Diabetic Nephropathy of Mice through Ampk/Sirt1/Nf-Kappab Pathway. Eur. J. Pharmacol. 2020, 886, 173449. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.H.; Tu, Y.; Ni, G.X.; Yan, J.; Yue, L.; Li, Z.L.; Wu, J.J.; Cao, Y.T.; Wan, Z.Y.; Sun, W.; et al. Total Flavones of Abelmoschus manihot Ameliorates Podocyte Pyroptosis and Injury in High Glucose Conditions by Targeting METTL3-Dependent m(6)A Modification-Mediated NLRP3-Inflammasome Activation and PTEN/PI3K/Akt Signaling. Front. Pharmacol. 2021, 12, 667644. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Jiang, H.; Chen, Y.; Ye, J.; Wang, A.; Wang, C.; Liu, Q.; Liang, G.; Deng, X.; Jiang, W.; et al. Oridonin is a covalent NLRP3 inhibitor with strong anti-inflammasome activity. Nat. Commun. 2018, 9, 2550. [Google Scholar] [CrossRef]
- Xu, X.N.; Jiang, Y.; Yan, L.Y.; Yin, S.Y.; Wang, Y.H.; Wang, S.B.; Fang, L.H.; Du, G.H. Aesculin suppresses the NLRP3 inflammasome-mediated pyroptosis via the Akt/GSK3beta/NF-kappaB pathway to mitigate myocardial ischemia/reperfusion injury. Phytomedicine 2021, 92, 153687. [Google Scholar] [CrossRef]
- Tang, K.; Su, W.; Huang, C.; Wu, Y.; Wu, X.; Lu, H. Notoginsenoside R1 suppresses inflammatory response and the pyroptosis of nucleus pulposus cells via inactivating NF-kappaB/NLRP3 pathways. Int. Immunopharmacol. 2021, 101, 107866. [Google Scholar] [CrossRef]
- Wang, M.; Wang, R.; Sun, H.; Sun, G.; Sun, X. Ginsenoside Rb1 ameliorates cardiotoxicity triggered by aconitine via inhibiting calcium overload and pyroptosis. Phytomedicine 2021, 83, 153468. [Google Scholar] [CrossRef]
- Luo, X.; Bao, X.; Weng, X.; Bai, X.; Feng, Y.; Huang, J.; Liu, S.; Jia, H.; Yu, B. The protective effect of quercetin on macrophage pyroptosis via TLR2/Myd88/NF-kappaB and ROS/AMPK pathway. Life Sci. 2022, 291, 120064. [Google Scholar] [CrossRef]
- Salehi, B.; Mishra, A.P.; Nigam, M.; Sener, B.; Kilic, M.; Sharifi-Rad, M.; Fokou, P.V.T.; Martins, N.; Sharifi-Rad, J. Resveratrol: A Double-Edged Sword in Health Benefits. Biomedicines 2018, 6, 91. [Google Scholar] [CrossRef]
- Liu, M.; Lu, J.; Yang, S.; Chen, Y.; Yu, J.; Guan, S. Alliin alleviates LPS-induced pyroptosis via promoting mitophagy in THP-1 macrophages and mice. Food Chem. Toxicol. 2022, 160, 112811. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Zhang, J.; Zhao, Y.; Zhang, Y.; Zhang, X.; Guan, J.; Liu, Y.; Fu, J. Curcumin protects against cognitive impairments in a rat model of chronic cerebral hypoperfusion combined with diabetes mellitus by suppressing neuroinflammation, apoptosis, and pyroptosis. Int. Immunopharmacol. 2021, 93, 107422. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.; Luo, X.; Feng, Y.; Fang, S.; Tian, J.; Yu, B.; Li, J. Luteolin prevents THP-1 macrophage pyroptosis by suppressing ROS production via Nrf2 activation. Chem. Biol. Interact. 2021, 345, 109573. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, Y.; Li, R.; Zhu, L.; Fu, B.; Yan, T. Salidroside ameliorates Parkinson’s disease by inhibiting NLRP3-dependent pyroptosis. Aging (Albany NY) 2020, 12, 9405–9426. [Google Scholar] [CrossRef]
- Ma, D.C.; Zhang, N.N.; Zhang, Y.N.; Chen, H.S. Salvianolic Acids for Injection alleviates cerebral ischemia/reperfusion injury by switching M1/M2 phenotypes and inhibiting NLRP3 inflammasome/pyroptosis axis in microglia in vivo and in vitro. J. Ethnopharmacol. 2021, 270, 113776. [Google Scholar] [CrossRef]
- Kai, J.; Yang, X.; Wang, Z.; Wang, F.; Jia, Y.; Wang, S.; Tan, S.; Chen, A.; Shao, J.; Zhang, F.; et al. Oroxylin a promotes PGC-1alpha/Mfn2 signaling to attenuate hepatocyte pyroptosis via blocking mitochondrial ROS in alcoholic liver disease. Free Radic. Biol. Med. 2020, 153, 89–102. [Google Scholar] [CrossRef]
- Hua, S.; Ma, M.; Fei, X.; Zhang, Y.; Gong, F.; Fang, M. Glycyrrhizin attenuates hepatic ischemia-reperfusion injury by suppressing HMGB1-dependent GSDMD-mediated kupffer cells pyroptosis. Int. Immunopharmacol. 2019, 68, 145–155. [Google Scholar] [CrossRef]
- Xu, L.; Wang, H.; Yu, Q.Q.; Ge, J.R.; Zhang, X.Z.; Mei, D.; Liang, F.Q.; Cai, X.Y.; Zhu, Y.; Shu, J.L.; et al. The monomer derivative of paeoniflorin inhibits macrophage pyroptosis via regulating TLR4/ NLRP3/ GSDMD signaling pathway in adjuvant arthritis rats. Int. Immunopharmacol. 2021, 101, 108169. [Google Scholar] [CrossRef]
- Liang, Q.; Cai, W.; Zhao, Y.; Xu, H.; Tang, H.; Chen, D.; Qian, F.; Sun, L. Lycorine ameliorates bleomycin-induced pulmonary fibrosis via inhibiting NLRP3 inflammasome activation and pyroptosis. Pharmacol. Res. 2020, 158, 104884. [Google Scholar] [CrossRef]
- Zhao, Y.; Li, Z.; Lu, E.; Sheng, Q.; Zhao, Y. Berberine exerts neuroprotective activities against cerebral ischemia/reperfusion injury through up-regulating PPAR-gamma to suppress NF-kappaB-mediated pyroptosis. Brain Res. Bull. 2021, 177, 22–30. [Google Scholar] [CrossRef]
- Lv, X.; Fan, C.; Jiang, Z.; Wang, W.; Qiu, X.; Ji, Q. Isoliquiritigenin alleviates P. gingivalis-LPS/ATP-induced pyroptosis by inhibiting NF-kappaB/ NLRP3/GSDMD signals in human gingival fibroblasts. Int. Immunopharmacol. 2021, 101, 108338. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Yang, J.H.; He, Y.; Zhou, H.F.; Wang, Y.; Ding, Z.S.; Jin, B.; Wan, H.T. Protocatechuic aldehyde prevents ischemic injury by attenuating brain microvascular endothelial cell pyroptosis via lncRNA Xist. Phytomedicine 2022, 94, 153849. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Yan, J.; Yun, M.; Yang, M.; Luo, Y.; Zhang, J.; Guo, M.; Yang, M.; Yuan, W.; Zou, W.; et al. Effect of Sanhuangyilong decoction plus methotrexate on tumor necrosis factor alpha and interferon gamma in serum and synovial fluid in rheumatoid arthritis patients with symptom pattern of damp heat obstruction. J. Tradit. Chin. Med. 2016, 36, 625–633. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Meng, X.; Zhang, X.; Su, X.; Liu, X.; Ren, K.; Ning, C.; Zhang, Q.; Zhang, S. Daphnes Cortex and Its Licorice-Processed Products Suppress Inflammation Via the Tlr4/Nf-Kappab/Nlrp3 Signaling Pathway and Regulation of the Metabolic Profile in the Treatment of Rheumatoid Arthritis. J. Ethnopharmacol. 2022, 283, 114657. [Google Scholar] [CrossRef] [PubMed]
- Jie, F.; Xiao, S.; Qiao, Y.; You, Y.; Feng, Y.; Long, Y.; Li, S.; Wu, Y.; Li, Y.; Du, Q. Kuijieling Decoction Suppresses Nlrp3-Mediated Pyroptosis to Alleviate Inflammation and Experimental Colitis in Vivo and in Vitro. J. Ethnopharmacol. 2021, 264, 113243. [Google Scholar] [CrossRef] [PubMed]
- Chao, L.; Li, Z.; Zhou, J.; Chen, W.; Li, Y.; Lv, W.; Guo, A.; Qu, Q.; Guo, S. Shen-Ling-Bai-Zhu-San Improves Dextran Sodium Sulfate-Induced Colitis by Inhibiting Caspase-1/Caspase-11-Mediated Pyroptosis. Front. Pharmacol. 2020, 11, 814. [Google Scholar] [CrossRef]
- Ye, B.; Lai, L. Yu Shi an Chang Fang Ameliorates Tnbs-Induced Colitis in Mice by Reducing Inflammatory Response and Protecting the Intestinal Mucosal Barrier. Evid. Based Complement. Alternat. Med. 2021, 2021, 8870901. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Drug | Structure | Mechanism of Action | IC50 | References |
|---|---|---|---|---|
| C172 |  | Blocks NLRP3 activation in a CFTR-independent manner | 0.3 μM | [98] |
| CY-09 |  | Binds to NLRP3 to inhibit its assembly and activation, and binds to Walker A site in the NACHT domain to inhibit caspase activity | 6 μM | [98] |
| OLT1177 |  | Inhibits the oligomerization and activation of typical and atypical NLRP3 inflammasomes, and alleviates LPS-induced systemic inflammation | / | [100,102] |
| MCC950 |  | Inhibits the release of IL-18 and IL-1β by inhibiting NLRP3 activation, and blocks ASC oligomerization | 7.5 × 10−3 μM | [98,104] |
| Glyburide |  | Inhibits NLRP3 signaling | 20 µM | [107] |
| sulofenur |  | Inhibits NLRP4 signaling | 0.034 μM | [106] |
| Glimepiride |  | Inhibits NLRP5 signaling | 5.4 × 10−3 μM | [106] |
| Dapagliflozin |  | Inhibits the NLRP3/ASC pathway | 1.6 × 10−3 μM | [108] |
| INF4E |  | Reduces myocardial infarct size and LDH release, and attenuates the formation of NLRP3 inflammasome in a time-dependent manner | / | [109] |
| INF39 |  | Attenuates NLRP3 structural changes detected by BRET and inhibits NLRP3 ATPase activity | 10 μM | [110,111,112] |
| HS-203873 |  | Attenuates NLRP3 activity and signal transduction | 50 μM | [110,111,112] |
| Tranilast |  | Appears to interact with the NLRP3 NACHT domain to disrupt NLRP3 intermolecular interactions and block NLRP3 oligomerization | 25 μM | [113] |
| RRx-001 |  | Covalently interacts with NLRP3 and blocks the NLRP3-NEK7 interaction | 0.117 μM | [114] |
| JC-171 |  | Down-regulates the expression of NLRP3-dependent IL-1β and reduces the level of ASC that is lowered by NLRP3 | 8.45 μM | [115] |
| Bay 11-7082 |  | Blocks the ATPase activity of NLRP3 | 3 μM | [116] |
| Parthenolide |  | Blocks the ATPase activity of NLRP3 | 1.4 μM | [117] |
| BOT-4-one |  | Blocks the ATPase activity of NLRP3 | 1.28 μM | [117] |
| MNS |  | Inhibits NLRP3 activity independently of Syk by directly interacting with NLRP3 to block caspase activity | 3 μM | [118] |
| Drug | Structure | Mechanism of Action | IC50 | References |
|---|---|---|---|---|
| VX-765 |  | VX-765 is the prerequisite drug of VRT-043198, and it can specifically inhibit caspase-1 after in vivo conversion to VRT-043198 | 0.7 μM | [119,120] |
| Boc-D-FMK |  | Inhibits caspase-1 activity | 39 μM | [97] |
| Ac-yvad-cmk |  | Prevents caspase-1 activation | / | [121] |
| Drug | Structure | Mechanism of Action | IC50 | References |
|---|---|---|---|---|
| NSA |  | Binds to GSDMD and inhibits the oligomerization of GSDMD-N | / | [122] |
| LDC7559 |  | Blocks the toxicity of GSDMD-N | / | [123] |
| Disulfiram |  | Inhibits the formation of GSDMD pores | <1 M | [124] |
| Fumarate |  | Acylate cysteine in GSDMD and prevents its interaction with cysteine proteases and subsequent activation | 0.003 μM | [125] |
| Bay11-7082 |  | Covalent modification of the cysteine 191/192 residue of GSDMD, interferes with the formation of GSDMD pore and IL-1β secretion | 10 μM | [126] |
| Drug | Structure | Mechanism of Action | Source | References |
|---|---|---|---|---|
| Baicalin |  | Reduces LDH and IL-1β release, eliminate caspase-1 cleavage and maturation | Scutellariae radix | [127] |
| Dihydromyricetin |  | Reduces LDH and IL-1β release, eliminate caspase-1 cleavage and IL-1β maturation | Ampelopsis grossedentata | [128] |
| Punicalagin |  | Prevents pyroptotic membrane permeability and may interfere with the insertion or oligomerization of GSDMD-N in the plasma membrane | Pomegranate | [129,130,131] |
| Catalpol |  | Inhibits AMPK/SIRT1/NF-κB pathway | Rehmanniae radix | [132] |
| Geniposide |  | Inhibits AMPK/SIRT1/NF-κB pathway | Gardenia jasminoides | [133] |
| Total flavones of Abelmoschus manihot | N/A | Inhibits NLRP3-inflammasome activation and PTEN/PI3K/Akt signaling pathway | Abelmoschus manihot | [134] |
| Oridonin |  | Interacts with NLRP3, covalently modifies NLRP3, and attenuates NLRP3 activation | Rabdosia rubescens | [135] |
| Aesculin |  | Promotes the phosphorylation of Akt/GSK3β, inhibits the phosphorylation of NF-κB, and significantly reduces the expression levels of NLRP3, caspase-1, GSDMD, and IL-1β | Ash bark | [136] |
| Notoginsenoside R1 |  | Inhibits the NF-κB signaling pathway, blocks the activation of NLRP3 inflammasome, prevents the cleavage of caspase-1 and IL-1β, reduces the expression of GSDMDN | Panax notoginseng | [137] |
| Ginsenoside Rb1 |  | Regulates Nrf2 /antioxidant response element signaling pathway and reduces the pyroptosis | Panax notoginseng | [138] |
| Quercetin |  | Inhibits LPS/ATP-induced NLRP3 inflammasome activation and suppresses the expression of GSDMD, NRPS, and IL-1β | Vegetables and fruits | [139] |
| Resveratrol |  | Inhibits the expression of Caspase-1, reduces the cleavage of GSDMD protein and the release of IL-1β and IL-18; suppresses the expression of p62 and the activation of NLRP3 inflammasome | Red grape skins | [140] |
| Alliin |  | Inhibits NLRP3 inflammasome activation and reduces the secretion of IL-1β and IL-18 | Garlic | [141] |
| Curcumin |  | Inhibits the NLRP3 inflammasome-mediated pyroptosis and reduces TLR4/NF-κB pathway | Curcuma longa | [142] |
| Luteolin |  | Inhibits NLRP3 inflammasome activation and reduces the expression of the N-terminal domain of GSDMD and IL-1β by inhibiting Nrf2 and NF-κB signaling pathways | A variety of plants | [143] |
| Salidroside |  | Reduces the levels of IL-1β, IL-18, and GSDMD, and inhibits the NLRP3/caspase-1/GSDMD axis mediated pyroptosis | Rhodiola rosea | [144] |
| Salvianolic acids | N/A | Reduces the cleavage of GSDMD-FL to GSDMD-N and inhibits the NLRP3 inflammasome/pyroptosis activation | Salviae miltiorrhizae | [145] |
| Oroxylin |  | Reduces the activation of NLRP3 inflammasome and caspase-1, and inhibits GSDMD-mediated pyroptosis and the release of IL-1β and IL-18 | Oroxylum indicum | [146] |
| Glycyrrhizin |  | Inhibits caspase-1/GSDMD signaling pathway | Licorice | [147] |
| Paeoniflorin |  | Inhibits the NLRP3/GSDMD signaling pathway | Paeonia lactiflora pall | [148] |
| Berberine |  | Inhibits the NLRP3/GSDMD signaling pathway | Coptis chinensis | [149] |
| Lycorine |  | Inhibits the NLRP3/GSDMD signaling pathway | Lycoris radiata | [150] |
| Isoliquiritigenin |  | Inhibits the NLRP3/GSDMD signaling pathway | Licorice | [151] |
| Protocatechuic Acid |  | Inhibits the NLRP3/GSDMD signaling pathway | Vegetables and fruits | [152] |
| Formulas | Consists of Herbs | Mechanism of Action | References |
|---|---|---|---|
| Baihu Guizhi Tang | Anemarrhena asphodeloides, Gypsum, Licorice, Rice, Cinnamon twig | Inhibits the activation of NLRP3 inflammasome | [153] |
| Daphnes Cortex | Stem and root bark of Daphne giraldii Nitsche | Regulates TLR4/NF-κB/NLRP3 signaling pathway | [154] |
| Kuijieling Tang | Ilicis Rotundae Cortex, Rhizoma Atractylodis, Macrocephalae, Paeoniae Radix Alba, Hirudo, and Radix Glycyrrhizae Preparata | Reduces the expression of NLRP3, caspase-1, GSDMD-N, IL-1β and IL-18 | [155] |
| Shenling Baizhu San | Semen lablab album, Atractylodes macrocephala, poria cocos, Licorice, Radix platycodi, Lotus seed, Ginseng, Fructus amomi, Yam, Coix seed | Inhibits IL-1β, IL-18, and TNF-α; and reduces the mRNA expression levels of NLRP3, ASC, and GSDMD-N | [156] |
| Yushi Anchang Fang | Scutellaria baicalensis, Astragalus, Atractylodes, Licorice, Radix Paeoniae Alba, Cinnamon, Pueraria, Carbonized Catnip, Radix Sanguisorbae, Bletilla Striata, Frankincense, Herba Patriniae, Sargent Gloryvine, and Panax Notoginseng powder | Down-regulates the expressions of NLRP3, ASC, and caspase-1 | [157] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, S.; Feng, M.; Zhang, S. Molecular Characteristics of Cell Pyroptosis and Its Inhibitors: A Review of Activation, Regulation, and Inhibitors. Int. J. Mol. Sci. 2022, 23, 16115. https://doi.org/10.3390/ijms232416115
Wei S, Feng M, Zhang S. Molecular Characteristics of Cell Pyroptosis and Its Inhibitors: A Review of Activation, Regulation, and Inhibitors. International Journal of Molecular Sciences. 2022; 23(24):16115. https://doi.org/10.3390/ijms232416115
Chicago/Turabian StyleWei, Shaoqiang, Min Feng, and Shidong Zhang. 2022. "Molecular Characteristics of Cell Pyroptosis and Its Inhibitors: A Review of Activation, Regulation, and Inhibitors" International Journal of Molecular Sciences 23, no. 24: 16115. https://doi.org/10.3390/ijms232416115
APA StyleWei, S., Feng, M., & Zhang, S. (2022). Molecular Characteristics of Cell Pyroptosis and Its Inhibitors: A Review of Activation, Regulation, and Inhibitors. International Journal of Molecular Sciences, 23(24), 16115. https://doi.org/10.3390/ijms232416115