
iPSC-Derived Macrophages: The Differentiation Protocol Affects Cell Immune Characteristics and Differentiation Trajectories
 , , , , ,
, , , , ,
Abstract
1. Introduction
2. Results
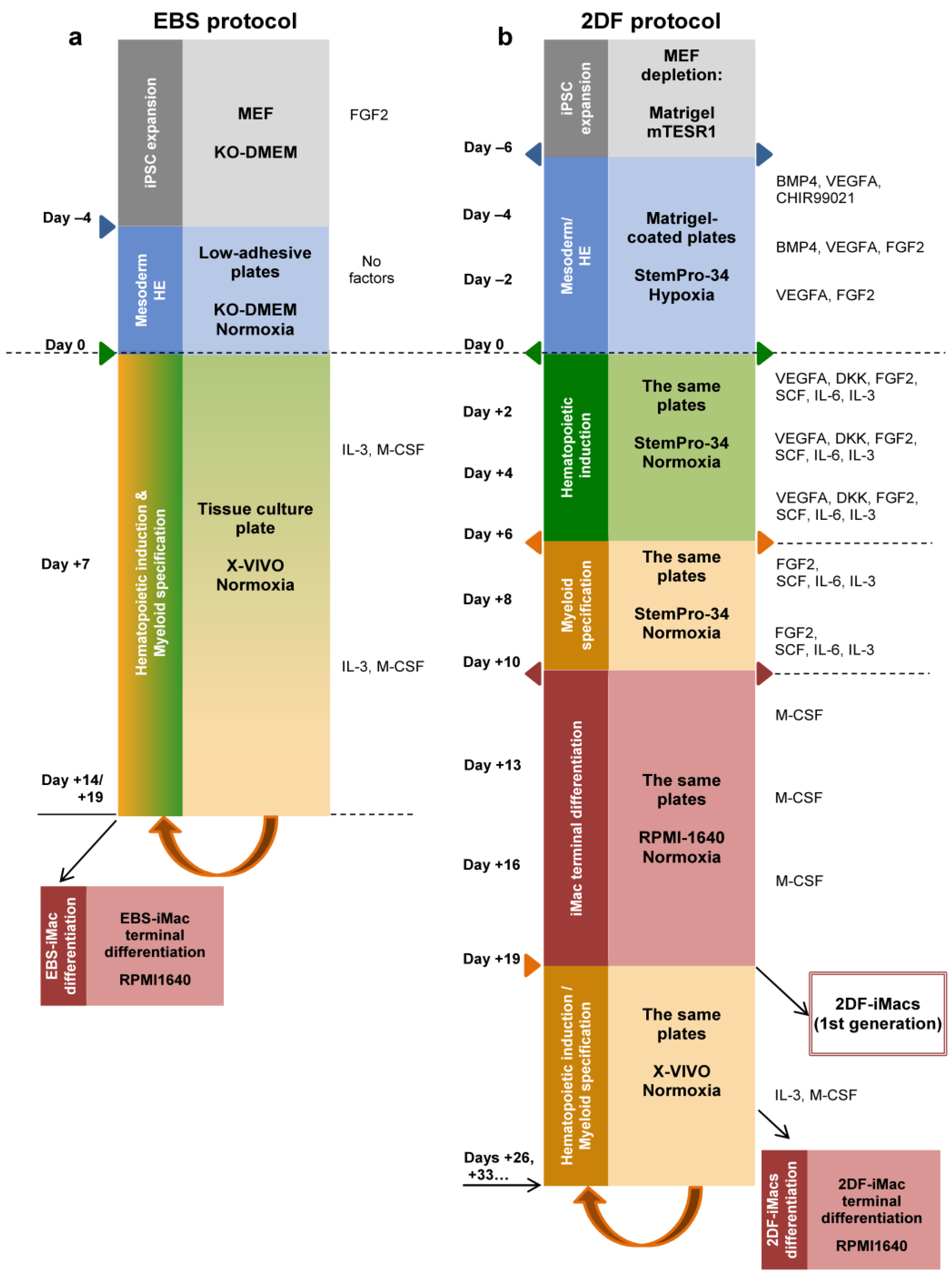
2.1. iMac Differentiation Using EBS and 2DF Protocols: The 2DF Protocol Is More Reliable
2.2. EBS-iMacs and 2DF-iMacs Display Similar Phenotype and Phagocytic Activity
2.3. iMac Secretory Profile Is Characterized by the Co-Production of Pro- and Anti-Inflammatory Factors and Depends on the Differentiation Protocol and Source iPSC Line
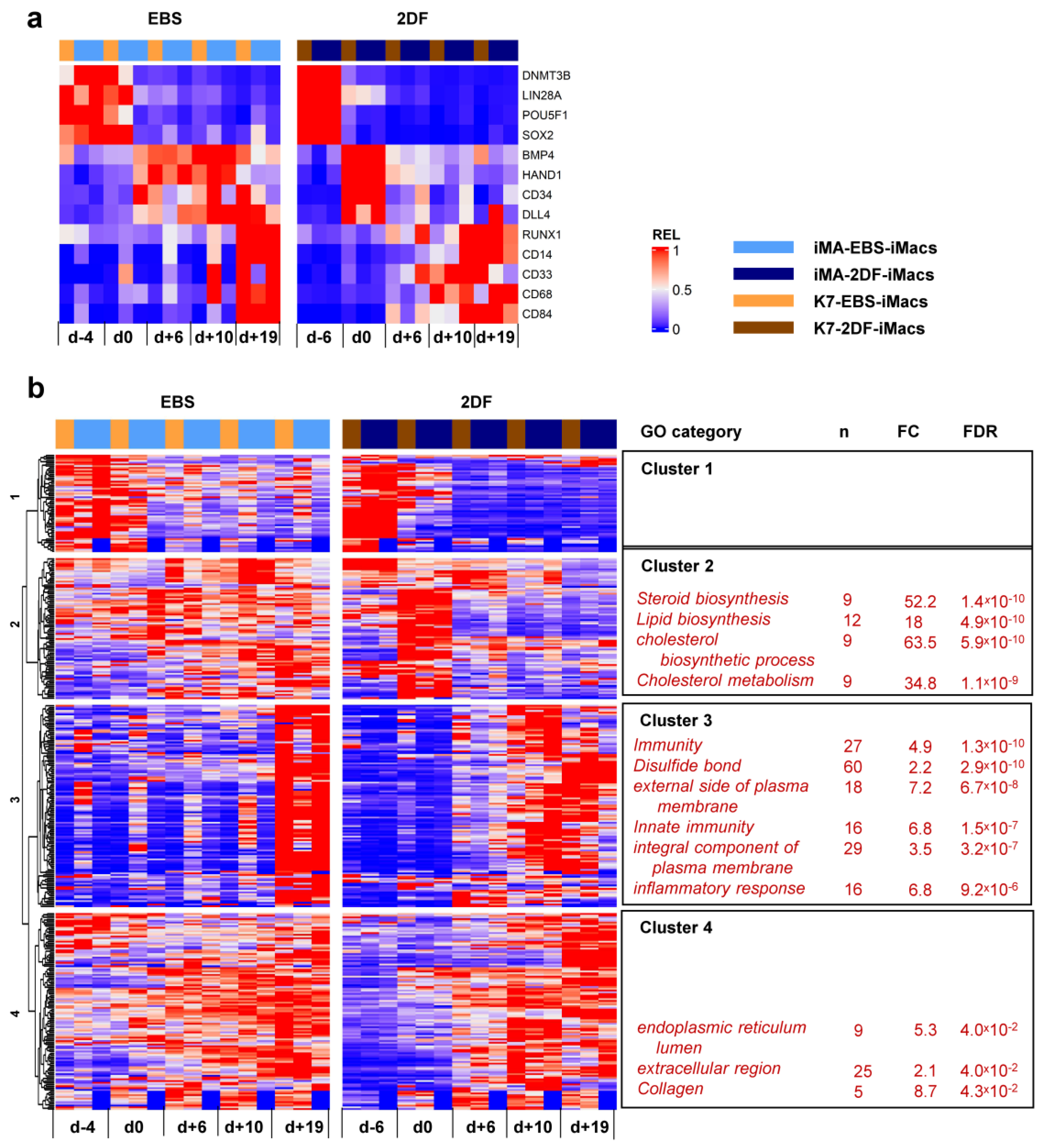
2.4. The Transcriptomic Profiles of EBS- and 2DF-iMacs Differ by the Expression of Genes Involved in Immune Response, Antigen Presentation and Lipid Homeostasis
2.5. Dynamic Transcriptomic Profiles of cells Differentiating Using EBS and 2DF Protocols Differ by the Expression of Genes Involved in the Inflammatory Response and Lipid Homeostasis
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. iPSC Cell Lines
4.3. The Differentiation of iMacs Using the EBS Protocol
4.4. The Differentiation of iMacs Using the 2DF Protocol
4.5. Cell Morphology, Cytospin Preparation and Staining
4.6. Flow Cytometry and Cell Sorting
4.7. iMac Phagocytic Activity
4.8. Multiplex Analysis
4.9. RNA Isolation, Transcriptome Library Preparation and Sequencing
4.10. RNA-Seq Data Analysis and Bioinformatics
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Panicker, L.M.; Miller, D.; Park, T.S.; Pate, B.; Azevedo, J.L.; Awad, O.; Masood, M.A.; Veenstra, T.D.; Goldin, E.; Stubblefield, B.K.; et al. Induced pluripotent stem cell model recapitulates pathologic hallmarks of Gaucher disease. Proc. Natl. Acad. Sci. USA 2012, 109, 18054–18059. [Google Scholar] [CrossRef] [PubMed]
- Brault, J.; Goutagny, E.; Telugu, N.; Shao, K.; Baquié, M.; Satre, V.; Coutton, C.; Grunwald, D.; Brion, J.P.; Barlogis, V.; et al. Optimized generation of functional neutrophils and macrophages from patient-specific induced pluripotent stem cells: Ex vivo models of X(0)-linked, AR22(0)- and AR47(0)- chronic granulomatous diseases. BioResearch 2014, 3, 311–326. [Google Scholar] [CrossRef]
- Zhang, H.; Xue, C.; Shah, R.; Bermingham, K.; Hinkle, C.C.; Li, W.; Rodrigues, A.; Tabita-Martinez, J.; Millar, J.S.; Cuchel, M.; et al. Functional analysis and transcriptomic profiling of iPSC-derived macrophages and their application in modeling Mendelian disease. Circ. Res. 2015, 117, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Klatt, D.; Cheng, E.; Philipp, F.; Selich, A.; Dahlke, J.; Schmidt, R.E.; Schott, J.W.; Büning, H.; Hoffmann, D.; Thrasher, A.J.; et al. Targeted repair of p47-CGD in iPSCs by CRISPR/Cas9: Functional correction without cleavage in the highly homologous pseudogenes. Stem Cell Rep. 2019, 13, 590–598. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Reilly, M.P. Human induced pluripotent stem cell-derived macrophages for unraveling human macrophage biology. Arterioscler. Throm. Vasc. Biol. 2017, 37, 2000–2006. [Google Scholar] [CrossRef]
- Han, H.W.; Seo, H.H.; Jo, H.Y.; Han, H.J.; Falcão, V.C.A.; Delorme, V.; Heo, J.; Shum, D.; Choi, J.H.; Lee, J.M.; et al. Drug discovery platform targeting M. tuberculosis with human embryonic stem cell-derived macrophages. Stem Cell Rep. 2019, 13, 980–991. [Google Scholar] [CrossRef] [PubMed]
- Neehus, A.L.; Lam, J.; Haake, K.; Merkert, S.; Schmidt, N.; Mucci, A.; Ackermann, M.; Schubert, M.; Happle, C.; Kühnel, M.P.; et al. Impaired IFNγ-signaling and mycobacterial clearance in IFNγR1-deficient human iPSC-derived macrophages. Stem Cell Rep. 2018, 10, 7–16. [Google Scholar] [CrossRef]
- O’Keeffe, A.; Hale, C.; Cotton, J.A.; Yardley, V.; Gupta, K.; Ananthanarayanan, A.; Murdan, S.; Croft, S.L. Novel 2D and 3D assays to determine the activity of anti-leishmanial drugs. Microorganisms 2020, 8, 831. [Google Scholar] [CrossRef] [PubMed]
- Iwamoto, Y.; Seki, Y.; Taya, K.; Tanaka, M.; Iriguchi, S.; Miyake, Y.; Nakayama, E.E.; Miura, T.; Shioda, T.; Akari, H.; et al. Generation of macrophages with altered viral sensitivity from genome-edited rhesus macaque iPSCs to model human disease. Mol. Ther. Methods Clin. Dev. 2021, 21, 262–273. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; See, M.; Nim, H.T.; Strumila, K.; Ng, E.S.; Hidalgo, A.; Ramialison, M.; Sutton, P.; Elefanty, A.G.; Sarkar, S.; et al. Human pluripotent stem cell-derived macrophages host Mycobacterium abscessus infection. Stem Cell Rep. 2022, 17, 2156–2166. [Google Scholar] [CrossRef]
- Zhang, H.; Shi, J.; Hachet, M.A.; Xue, C.; Bauer, R.C.; Jiang, H.; Li, W.; Tohyama, J.; Millar, J.; Billheimer, J.; et al. CRISPR/Cas9-mediated gene editing in human iPSC-derived macrophage reveals lysosomal acid lipase function in human macrophages-brief report. Arterioscler. Thromb. Vasc. Biol. 2017, 37, 2156–2160. [Google Scholar] [CrossRef]
- Navarro-Guerrero, E.; Tay, C.; Whalley, J.P.; Cowley, S.A.; Davies, B.; Knight, J.C.; Ebner, D. Genome-wide CRISPR/Cas9-knockout in human induced pluripotent stem cell (iPSC)-derived macrophages. Sci. Rep. 2021, 11, 4245. [Google Scholar] [CrossRef] [PubMed]
- Lyadova, I.; Vasiliev, A. Macrophages derived from pluripotent stem cells: Prospective applications and research gaps. Cell Biosci. 2022, 12, 96. [Google Scholar] [CrossRef] [PubMed]
- Ackermann, M.; Kempf, H.; Hetzel, M.; Hesse, C.; Hashtchin, A.R.; Brinkert, K.; Schott, J.W.; Haake, K.; Kühnel, M.P.; Glage, S.; et al. Bioreactor-based mass production of human iPSC-derived macrophages enables immunotherapies against bacterial airway infections. Nat. Commun. 2018, 9, 5088. [Google Scholar] [CrossRef] [PubMed]
- Hashtchin, A.R.; Fehlhaber, B.; Hetzel, M.; Manstein, F.; Stalp, J.L.; Glage, S.; Abeln, M.; Zweigerdt, R.; Munder, A.; Viemann, D.; et al. Human iPSC-derived macrophages for efficient Staphylococcus aureus clearance in a murine pulmonary infection model. Blood Adv. 2021, 5, 5190–5201. [Google Scholar] [CrossRef]
- Miyashita, A.; Fukushima, S.; Nakahara, S.; Kubo, Y.; Tokuzumi, A.; Yamashita, J.; Aoi, J.; Haruta, M.; Senju, S.; Nishimura, Y.; et al. Immunotherapy against metastatic melanoma with human iPS cell-derived myeloid cell lines producing type I interferons. Cancer Immunol. Res. 2016, 4, 248–258. [Google Scholar] [CrossRef]
- Happle, C.; Lachmann, N.; Ackermann, M.; Mirenska, A.; Göhring, G.; Thomay, K.; Mucci, A.; Hetzel, M.; Glomb, T.; Suzuki, T.; et al. Pulmonary transplantation of human induced pluripotent stem cell-derived macrophages ameliorates pulmonary alveolar proteinosis. Am. J. Respir. Crit. Care Med. 2018, 198, 350–360. [Google Scholar] [CrossRef]
- Mucci, A.; Lopez-Rodriguez, E.; Hetzel, M.; Liu, S.; Suzuki, T.; Happle, C.; Ackermann, M.; Kempf, H.; Hillje, R.; Kunkiel, J.; et al. iPSC-derived macrophages effectively treat pulmonary alveolar proteinosis in Csf2rb-deficient mice. Stem Cell Rep. 2018, 11, 696–710. [Google Scholar] [CrossRef]
- Lee, C.; Kozaki, T.; Ginhoux, F. Studying tissue macrophages in vitro: Are iPSC-derived cells the answer? Nat. Rev. Immunol. 2018, 18, 716–725. [Google Scholar] [CrossRef]
- Lyadova, I.; Gerasimova, T.; Nenasheva, T. Macrophages derived from human induced pluripotent stem cells: The diversity of protocols, future prospects, and outstanding questions. Front. Cell Dev. Biol. 2021, 9, 640703. [Google Scholar] [CrossRef]
- Choi, K.D.; Yu, J.; Smuga-Otto, K.; Salvagiotto, G.; Rehrauer, W.; Vodyanik, M.; Thomson, J.; Slukvin, I. Hematopoietic and endothelial differentiation of human induced pluripotent stem cells. Stem Cells 2009, 27, 559–567. [Google Scholar] [CrossRef] [PubMed]
- van Wilgenburg, B.; Browne, C.; Vowles, J.; Cowley, S.A. Efficient, long term production of monocyte-derived macrophages from human pluripotent stem cells under partly-defined and fully-defined conditions. PLoS ONE 2013, 8, e71098. [Google Scholar] [CrossRef] [PubMed]
- Lachmann, N.; Ackermann, M.; Frenzel, E.; Liebhaber, S.; Brennig, S.; Happle, C.; Hoffmann, D.; Klimenkova, O.; Lüttge, D.; Buchegger, T.; et al. Large-scale hematopoietic differentiation of human induced pluripotent stem cells provides granulocytes or macrophages for cell replacement therapies. Stem Cell Rep. 2015, 4, 282–296. [Google Scholar] [CrossRef] [PubMed]
- Ackermann, M.; Haake, K.; Kempf, H.; Kaschutnig, P.; Weiss, A.C.; Nguyen, A.; Abeln, M.; Merkert, S.; Kühnel, M.P.; Hartmann, D.; et al. A 3D iPSC-differentiation model identifies interleukin-3 as a regulator of early human hematopoietic specification. Haematologica 2021, 106, 1354–1367. [Google Scholar] [CrossRef] [PubMed]
- Buchrieser, J.; James, W.; Moore, M.D. Human induced pluripotent stem cell-derived macrophages share ontogeny with MYB-independent tissue-resident macrophages. Stem Cell Rep. 2017, 8, 334–345. [Google Scholar] [CrossRef] [PubMed]
- Gutbier, S.; Wanke, F.; Dahm, N.; Rümmelin, A.; Zimmermann, S.; Christensen, K.; Köchl, F.; Rautanen, A.; Hatje, K.; Geering, B.; et al. Large-scale production of human iPSC-derived macrophages for drug screening. Int. J. Mol. Sci. 2020, 21, 4808. [Google Scholar] [CrossRef]
- Lopez-Yrigoyen, M.; May, A.; Ventura, T.; Taylor, H.; Fidanza, A.; Cassetta, L.; Pollard, J.W.; Forrester, L.M. Production and characterization of human macrophages from pluripotent stem cells. J. Vis. Exp. 2020, 158, e61038. [Google Scholar] [CrossRef]
- Shi, J.; Xue, C.; Liu, W.; Zhang, H. Differentiation of human-induced pluripotent stem cells to macrophages for disease modeling and functional genomics. Curr. Protoc. Stem Cell Biol. 2019, 48, e74. [Google Scholar] [CrossRef]
- Takata, K.; Kozaki, T.; Lee, C.; Thion, M.S.; Otsuka, M.; Lim, S.; Utami, K.H.; Fidan, K.; Park, D.S.; Malleret, B.; et al. Induced-pluripotent-stem-cell-derived primitive macrophages provide a platform for modeling tissue-resident macrophage differentiation and function. Immunity 2017, 47, 183–198. [Google Scholar] [CrossRef]
- Cao, X.; Yakala, G.K.; van den Hil, F.E.; Cochrane, A.; Mummery, C.L.; Orlova, V.V. Differentiation and functional comparison of monocytes and macrophages from hiPSCs with peripheral blood derivatives. Stem Cell Rep. 2019, 12, 1282–1297. [Google Scholar] [CrossRef]
- Mukherjee, C.; Hale, C.; Mukhopadhyay, S. A simple multistep protocol for differentiating human induced pluripotent stem cells into functional macrophages. Methods Mol. Biol. 2018, 1784, 13–28. [Google Scholar] [CrossRef] [PubMed]
- Nenasheva, T.; Gerasimova, T.; Serdyuk, Y.; Grigor’eva, E.; Kosmiadi, G.; Nikolaev, A.; Dashinimaev, E.; Lyadova, I. Macrophages derived from human induced pluripotent stem cells are low-activated “naïve-like” cells capable of restricting mycobacteria growth. Front. Immunol. 2020, 11, 1016. [Google Scholar] [CrossRef] [PubMed]
- Grigor’eva, E.V.; Malankhanova, T.B.; Surumbayeva, A.; Pavlova, S.V.; Minina, J.M.; Kizilova, E.A.; Suldina, L.A.; Morozova, K.N.; Kiseleva, E.; Sorokoumov, E.D.; et al. Generation of GABAergic striatal neurons by a novel iPSC differentiation protocol enabling scalability and cryopreservation of progenitor cells. Cytotechnology 2020, 72, 649–663. [Google Scholar] [CrossRef] [PubMed]
- Malakhova, A.A.; Grigor’eva, E.V.; Pavlova, S.V.; Malankhanova, T.B.; Valetdinova, K.R.; Vyatkin, Y.V.; Khabarova, E.A.; Rzaev, J.A.; Zakian, S.M.; Medvedev, S.P. Generation of induced pluripotent stem cell lines ICGi021-A and ICGi022-A from peripheral blood mononuclear cells of two healthy individuals from Siberian population. Stem Cell Res. 2020, 48, 101952. [Google Scholar] [CrossRef]
- Jiang, Y.; Cowley, S.A.; Siler, U.; Melguizo, D.; Tilgner, K.; Browne, C.; Dewilton, A.; Przyborski, S.; Saretzki, G.; James, W.S.; et al. Derivation and functional analysis of patient-specific induced pluripotent stem cells as an in vitro model of chronic granulomatous disease. Stem Cells 2012, 30, 599–611. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W. Smyth GK limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]
- Strano, A.; Tuck, E.; Stubbs, V.E.; Livesey, F.J. Variable outcomes in neural differentiation of human PSCs arise from intrinsic differences in developmental signaling pathways. Cell Rep. 2020, 31, 107732. [Google Scholar] [CrossRef]
- Murugesan, G.; Davidson, L.; Jannetti, L.; Crocker, P.R.; Weigle, B. Quantitative proteomics of polarised macrophages derived from induced pluripotent stem cells. Biomedicines 2022, 10, 239. [Google Scholar] [CrossRef]
- Alasoo, K.; Martinez, F.O.; Hale, C.; Gordon, S.; Powrie, F.; Dougan, G.; Mukhopadhyay, S.; Gaffney, D.J. Transcriptional profiling of macrophages derived from monocytes and iPS cells identifies a conserved response to LPS and novel alternative transcription. Sci. Rep. 2015, 5, 12524. [Google Scholar] [CrossRef]
- Joshi, K.; Elso, C.; Motazedian, A.; Labonne, T.; Schiesser, J.V.; Cameron, F.; Mannering, S.I.; Elefanty, A.G.; Stanley, E.G. Induced pluripotent stem cell macrophages present antigen to proinsulin-specific T cell receptors from donor-matched islet-infiltrating T cells in type 1 diabetes. Diabetologia 2019, 62, 2245–2251. [Google Scholar] [CrossRef] [PubMed]
- Libby, P.; Ridker, P.M.; Maseri, A. Inflammation and atherosclerosis. Circulation 2002, 105, 1135–1143. [Google Scholar] [CrossRef] [PubMed]
- Sorci-Thomas, M.G.; Thomas, M.J. Microdomains, inflammation, and atherosclerosis. Circ. Res. 2016, 118, 679–691. [Google Scholar] [CrossRef] [PubMed]
- Wolf, D.; Ley, K. Immunity and inflammation in atherosclerosis. Circ. Res. 2019, 124, 315–327. [Google Scholar] [CrossRef]
- Spann, N.J.; Garmire, L.X.; McDonald, J.G.; Myers, D.S.; Milne, S.B.; Shibata, N.; Reichart, D.; Fox, J.N.; Shaked, I.; Heudobler, D.; et al. Regulated accumulation of desmosterol integrates macrophage lipid metabolism and inflammatory responses. Cell 2012, 151, 138–152. [Google Scholar] [CrossRef]
- Araldi, E.; Fernández-Fuertes, M.; Canfrán-Duque, A.; Tang, W.; Cline, G.W.; Madrigal-Matute, J.; Pober, J.S.; Lasunción, M.A.; Wu, D.; Fernández-Hernando, C.; et al. Lanosterol modulates TLR4-mediated innate immune responses in macrophages. Cell Rep. 2017, 19, 2743–2755. [Google Scholar] [CrossRef]
- Lappalainen, J.; Yeung, N.; Nguyen, S.D.; Jauhiainen, M.; Kovanen, P.T.; Lee-Rueckert, M. Cholesterol loading suppresses the atheroinflammatory gene polarization of human macrophages induced by colony stimulating factors. Sci. Rep. 2021, 11, 4923. [Google Scholar] [CrossRef]
- Bagnati, M.; Moreno-Moral, A.; Ko, J.H.; Nicod, J.; Harmston, N.; Imprialou, M.; Game, L.; Gil, J.; Petretto, E.; Behmoaras, J. Systems genetics identifies a macrophage cholesterol network associated with physiological wound healing. JCI Insight 2019, 4, e125736. [Google Scholar] [CrossRef]
- Ensan, S.; Li, A.; Besla, R.; Degousee, N.; Cosme, J.; Roufaiel, M.; Shikatani, E.A.; El-Maklizi, M.; Williams, J.W.; Robins, L.; et al. Robbins CS Self-renewing resident arterial macrophages arise from embryonic CX3CR1(+) precursors and circulating monocytes immediately after birth. Nat. Immunol. 2016, 17, 159–168. [Google Scholar] [CrossRef]
- Swirski, F.K.; Robbins, C.S.; Nahrendorf, M. Development and function of arterial and cardiac macrophages. Trends immunol. 2016, 37, 32–40. [Google Scholar] [CrossRef]
- Honold, L.; Nahrendorf, M. Resident and monocyte-derived macrophages in cardiovascular disease. Circ. Res. 2018, 122, 113–127. [Google Scholar] [CrossRef] [PubMed]
- Lee-Rueckert, M.; Lappalainen, J.; Kovanen, P.T.; Escola-Gil, J.C. Lipid-laden macrophages and inflammation in atherosclerosis and cancer: An Integrative View. Front. Cardiovasc. Med. 2022, 9, 777822. [Google Scholar] [CrossRef] [PubMed]
- Weinberger, T.; Esfandyari, D.; Messerer, D.; Percin, G.; Schleifer, C.; Thaler, R.; Liu, L.; Stremmel, C.; Schneider, V.; Vagnozzi, R.J.; et al. Ontogeny of arterial macrophages defines their functions in homeostasis and inflammation. Nat. Commun. 2020, 11, 4549. [Google Scholar] [CrossRef] [PubMed]
- Vanhee, S.; De Mulder, K.; Van Caeneghem, Y.; Verstichel, G.; Van Roy, N.; Menten, B.; Velghe, I.; Philippé, J.; De Bleser, D.; Lambrecht, B.N.; et al. In vitro human embryonic stem cell hematopoiesis mimics MYB-independent yolk sac hematopoiesis. Haematologica 2015, 100, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Konttinen, H.; Cabral-da-Silva, M.; Ohtonen, S.; Wojciechowski, S.; Shakirzyanova, A.; Caligola, S.; Giugno, R.; Ishchenko, Y.; Hernández, D.; Fazaludeen, M.F.; et al. PSEN1ΔE9, APPswe, and APOE4 confer disparate phenotypes in human iPSC-derived microglia. Stem Cell Rep. 2019, 13, 669–683. [Google Scholar] [CrossRef]
- Monkley, S.; Krishnaswamy, J.K.; Göransson, M.; Clausen, M.; Meuller, J.; Thörn, K.; Hicks, R.; Delaney, S.; Stjernborg, L. Optimised generation of iPSC-derived macrophages and dendritic cells that are functionally and transcriptionally similar to their primary counterparts. PLoS ONE 2020, 15, e0243807. [Google Scholar] [CrossRef]
- Jo, H.Y.; Seo, H.H.; Gil, D.; Par, Y.; Han, H.J.; Han, H.W.; Thimmulappa, R.K.; Kim, S.C.; Kim, J.H. Single-Cell RNA sequencing of human pluripotent stem cell-derived macrophages for quality control of the cell therapy product. Front. Genet. 2022, 12, 658862. [Google Scholar] [CrossRef]
- Vaughan-Jackson, A.; Stodolak, S.; Ebrahimi, K.H.; Browne, C.; Reardon, P.K.; Pires, E.; Gilbert-Jaramillo, J.; Cowley, S.A.; James, W.S. Differentiation of human induced pluripotent stem cells to authentic macrophages using a defined, serum-free, open-source medium. Stem Cell Rep. 2021, 16, 1735–1748. [Google Scholar] [CrossRef]
- Shen, J.; Xu, Y.; Zhang, S.; Lyu, S.; Huo, Y.; Zhu, Y.; Tang, K.; Mou, J.; Li, X.; Hoyle, D.L.; et al. Single-cell transcriptome of early hematopoiesis guides arterial endothelial-enhanced functional T cell generation from human PSCs. Sci. Adv. 2021, 7, eabi9787. [Google Scholar] [CrossRef]
- Korotkevich, G.; Sukhov, V.; Budin, N.; Shpak, B.; Artyomov, M.N.; Sergushichev, A. Fast gene set enrichment analysis. bioRxiv 2021. [Google Scholar] [CrossRef]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef]
- Liberzon, A.; Birger, C.; Thorvaldsdóttir, H.; Ghandi, M.; Mesirov, J.P.; Tamayo, P. The Molecular Signatures Database (MSigDB) hallmark gene set collection. Cell Syst. 2015, 1, 417–425. [Google Scholar] [CrossRef] [PubMed]
- Dolgalev, I. MSigDB Gene Sets for Multiple Organisms in a Tidy Data Format. 2022. Available online: https://igordot.github.io/msigdbr/ (accessed on 30 March 2022).
- Gu, Z.; Eils, R.; Schlesner, M. Complex heatmaps reveal patterns and correlations in multidimensional genomic data. Bioinformatics 2016, 32, 2847–2849. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Lin, Y.; Sun, Z.; Yuan, X.; Chen, L.; Shen, B. Knowledge-guided bioinformatics model for identifying autism spectrum disorder diagnostic microRNA biomarkers. Sci. Rep. 2016, 6, 39663. [Google Scholar] [CrossRef] [PubMed]
- Apweiler, R.; Bairoch, A.; Wu, C.H.; Barker, W.C.; Boeckmann, B.; Ferro, S.; Gasteiger, E.; Huang, H.; Lopez, R.; Magrane, M.; et al. UniProt: The universal protein knowledgebase. Nucleic Acids Res. 2004, 32, D115–D119. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Hochberg, Y.; Benjamini, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Statist. Soc. B 1995, 57, 289–300. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Comparison | Pattern (DEG Number) | Biological Process/ Pathway Category | Source | DEG Number | FC | FDR | Identity of DEGs |
|---|---|---|---|---|---|---|---|
| iMA- EBS-iMacs versus iMA- 2DF-iMacs | Up-regulated in EBS-iMacs (49) * | cellular response to interleukin-1 | GOTERM_BP_DIRECT | 5 | 10.2 | 2.3 × 10−2 | CCL2, CCL8, MYC, CHI3L1 and ICAM1 |
| cellular response to tumor necrosis factor | GOTERM_BP_DIRECT | 6 | 15.7 | 2.3 × 10−2 | CCL2, CCL8, GATA3, CHI3L1, ICAM1 and POSTN | ||
| inflammatory response | GOTERM_BP_DIRECT | 8 | 7.2 | 2.3 × 10−2 | BCL6, CCL2, CCL8, NLRP2, CHI3L1, FASN, IL1RN and PTGER2 | ||
| inflammatory response | UP_KW_BIOLOGICAL_ PROCESS | 5 | 9.6 | 4.6 × 10−2 | BCL6, CCL2, CCL8, NLRP2 and CHI3 | ||
| Up-regulated in 2DF-iMacs (118) * | ZN_FING:C2H2-type 10 | UP_SEQ_FEATURE | 15 | 6.7 | 4.1 × 10−6 | ZNF117, ZNF253, ZNF287, ZNF329, ZNF331, ZNF347, ZNF 404, ZNF415, ZNF471, ZNF486, ZNF568, ZNF595, ZNF626, ZNF677 and ZNF93 | |
| Herpes simplex virus 1 infection | KEGG_PATHWAY | 13 | 3.6 | 2.0 × 10−2 | HLA-F, HLA-DRB1, ZNF253, ZNF331, ZNF347, ZNF404, ZNF415, ZNF471, ZNF486, ZNF568, ZNF595, ZNF677 and ZNF93 | ||
| RNA polymerase II transcription factor activity and sequence-specific DNA binding | GOTERM_MF_DIRECT | 23 | 3.3 | 1.6 × 10−4 | ELF3, SOX4, TBX15, ATOH8, PAX8, TSHZ3, ZSCAN18, ZNF117, ZNF253, ZNF287, ZNF329, ZNF331, ZNF347, ZNF404, ZNF415, ZNF471, ZNF486, ZNF568, ZNF595, ZNF626, ZNF677, ZNF844 and ZNF93 | ||
| K7- EBS-iMacs versus K7- 2DF-iMacs | Up-regulated in K7-iMacs (126) * | endocytic vesicle membrane | GOTERM_CC_DIRECT | 8 | 15.5 | 1.4 × 10−4 | CD74, CD9, WNT5A, CSF3R, HBEGF, MSR1, HLA-DPA1 and HLA-DQB1 |
| MHC class II protein complex | GOTERM_CC_DIRECT | 4 | 30.4 | 6.3 × 10−3 | CD74, HLA-DMB, HLA-DPA1 and HLA-DQB1 | ||
| antigen processing and presentation of exogenous peptide antigen via MHC class II | GOTERM_BP_DIRECT | 5 | 26.9 | 3.2 × 10−2 | CD74, FCGR2B, HLA-DMB, HLA-DPA1 and HLA-DQB1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Klepikova, A.; Nenasheva, T.; Sheveleva, O.; Protasova, E.; Antonov, D.; Gainullina, A.; Chikina, E.; Sakovnich, O.; Gerasimova, T.; Nikitina, I.; et al. iPSC-Derived Macrophages: The Differentiation Protocol Affects Cell Immune Characteristics and Differentiation Trajectories. Int. J. Mol. Sci. 2022, 23, 16087. https://doi.org/10.3390/ijms232416087
Klepikova A, Nenasheva T, Sheveleva O, Protasova E, Antonov D, Gainullina A, Chikina E, Sakovnich O, Gerasimova T, Nikitina I, et al. iPSC-Derived Macrophages: The Differentiation Protocol Affects Cell Immune Characteristics and Differentiation Trajectories. International Journal of Molecular Sciences. 2022; 23(24):16087. https://doi.org/10.3390/ijms232416087
Chicago/Turabian StyleKlepikova, Anna, Tatiana Nenasheva, Olga Sheveleva, Elena Protasova, Daniil Antonov, Anastasiia Gainullina, Evgeniia Chikina, Olga Sakovnich, Tatiana Gerasimova, Irina Nikitina, and et al. 2022. "iPSC-Derived Macrophages: The Differentiation Protocol Affects Cell Immune Characteristics and Differentiation Trajectories" International Journal of Molecular Sciences 23, no. 24: 16087. https://doi.org/10.3390/ijms232416087
APA StyleKlepikova, A., Nenasheva, T., Sheveleva, O., Protasova, E., Antonov, D., Gainullina, A., Chikina, E., Sakovnich, O., Gerasimova, T., Nikitina, I., Shevalie, D., & Lyadova, I. (2022). iPSC-Derived Macrophages: The Differentiation Protocol Affects Cell Immune Characteristics and Differentiation Trajectories. International Journal of Molecular Sciences, 23(24), 16087. https://doi.org/10.3390/ijms232416087