

AKBA Promotes Axonal Regeneration via RhoA/Rictor to Repair Damaged Sciatic Nerve
 ,
,
Abstract
1. Introduction
2. Results
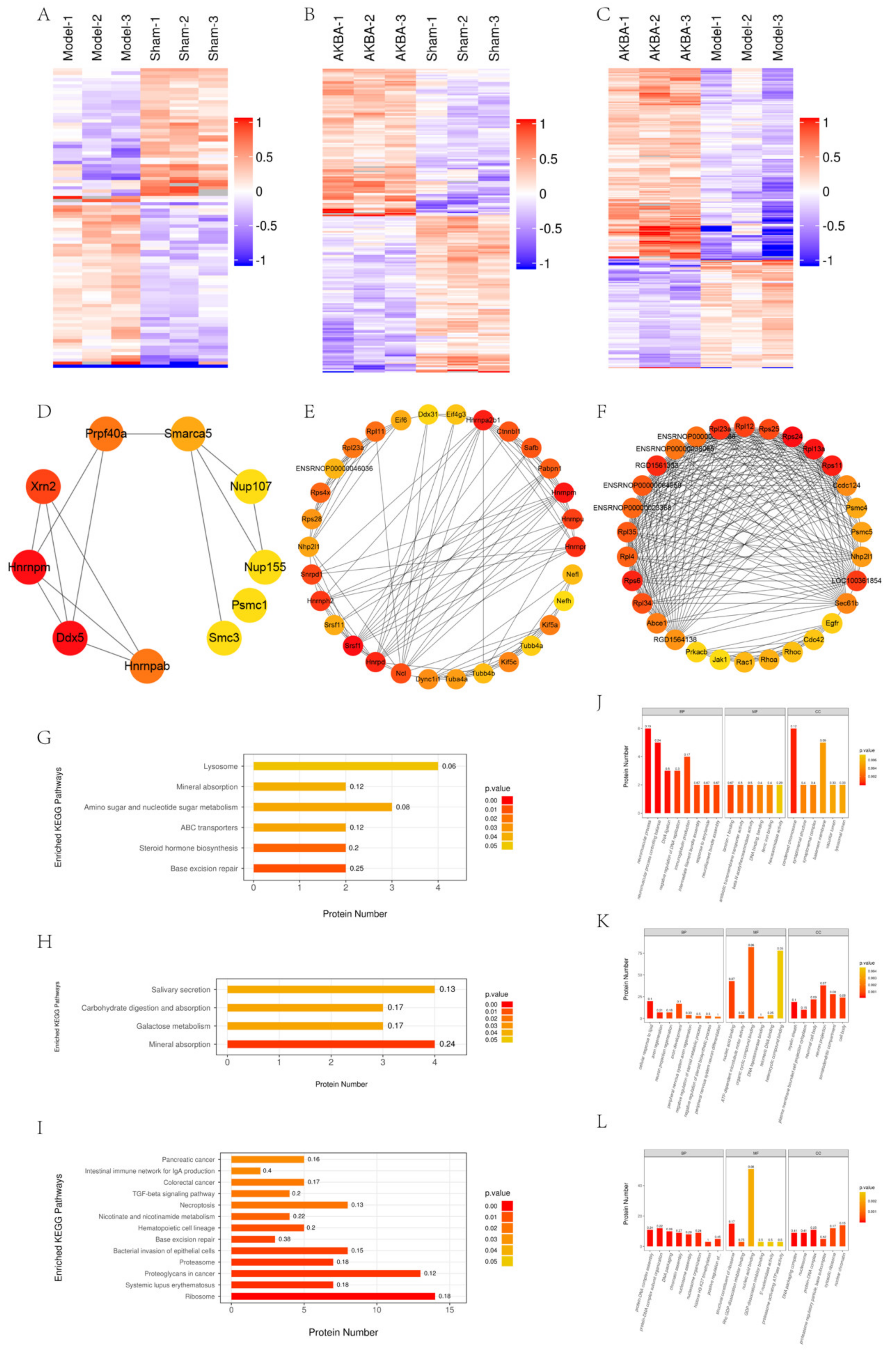
2.1. Bioinformatic Analysis of The Proteomics of Rat Sciatic Nerve Samples
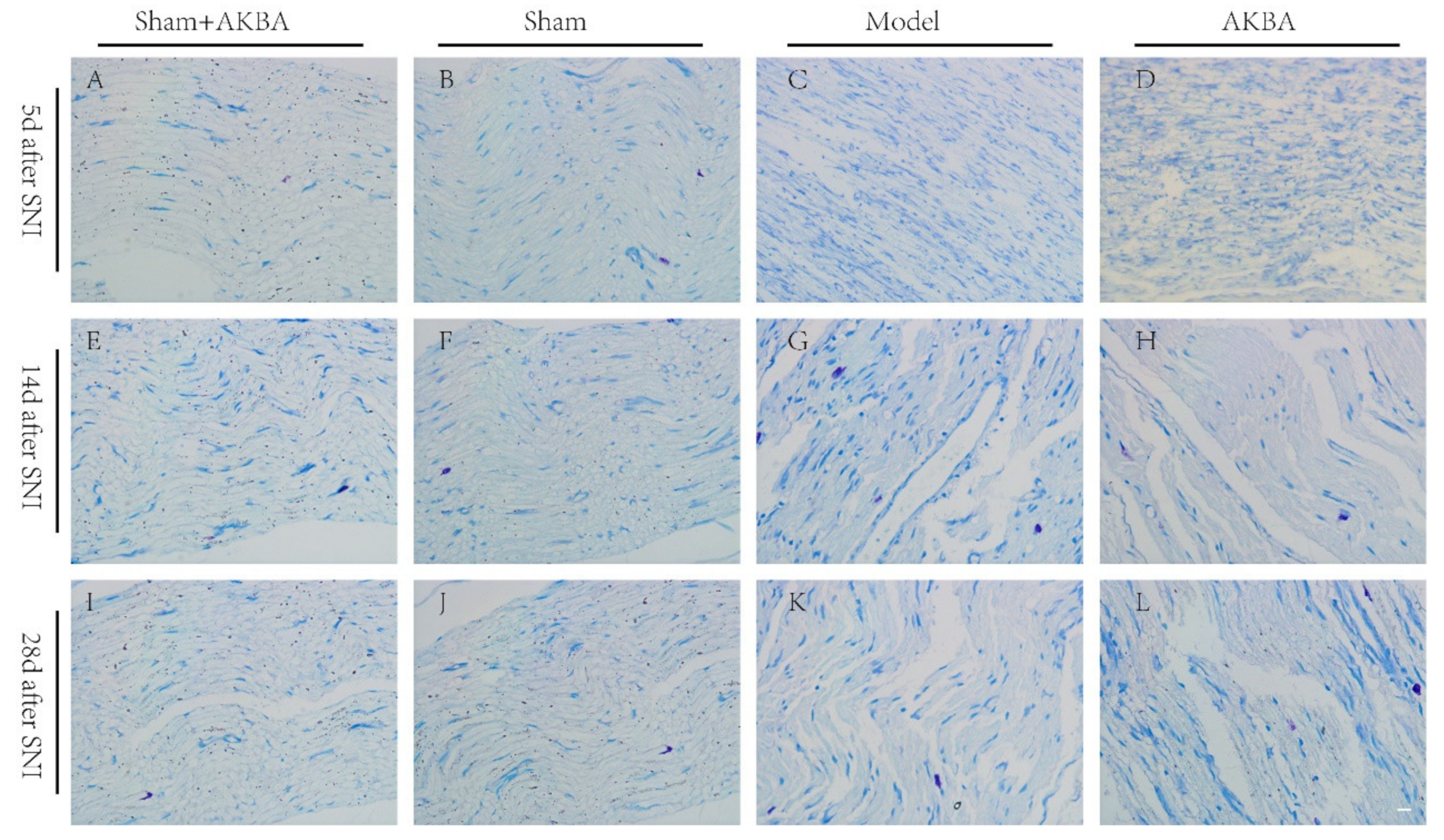
2.2. Toluidine Blue Staining of The Injured Sciatic Nerve in Rats
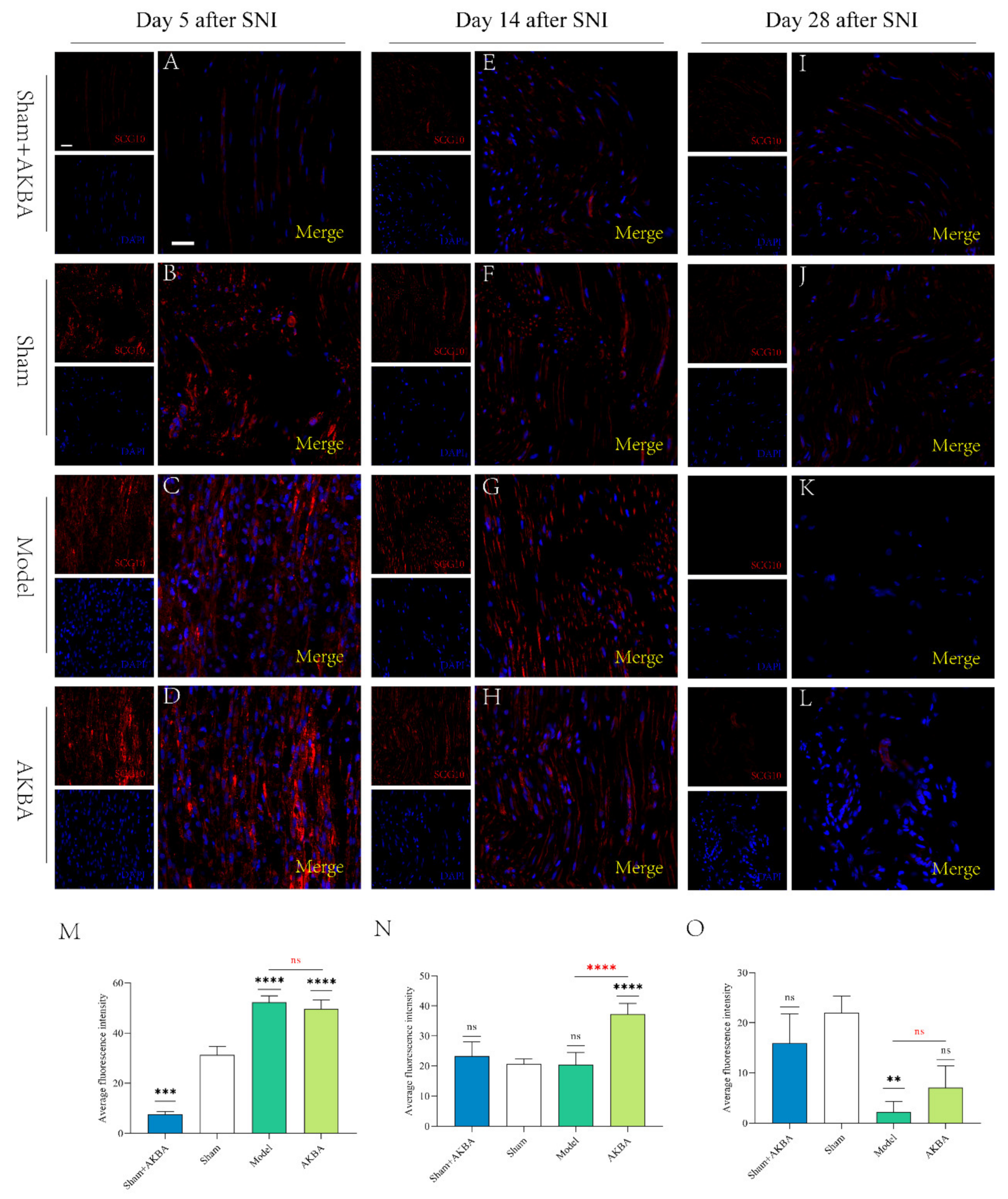
2.3. Immunofluorescence Detection of Nascent Axons by SCG10
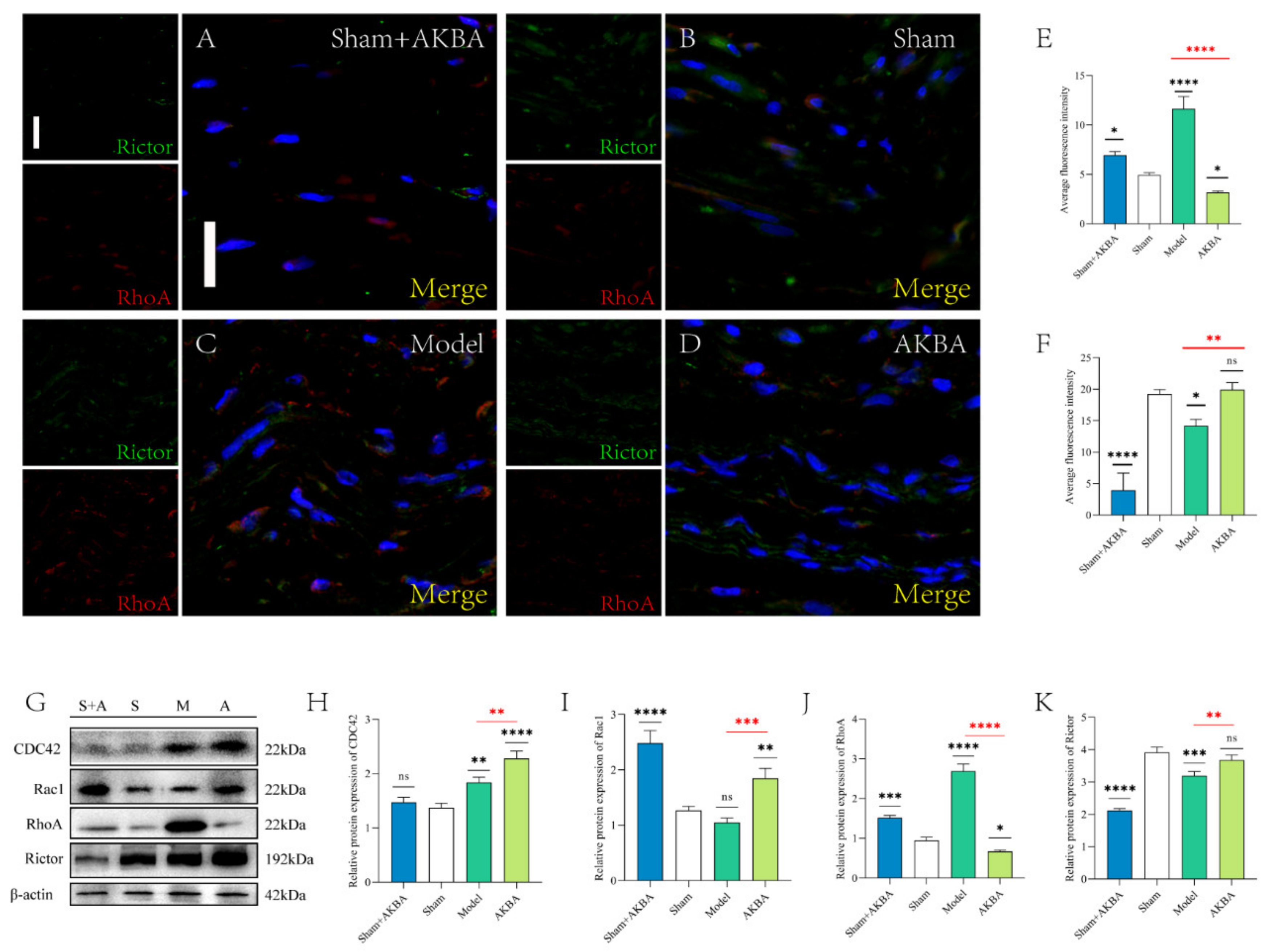
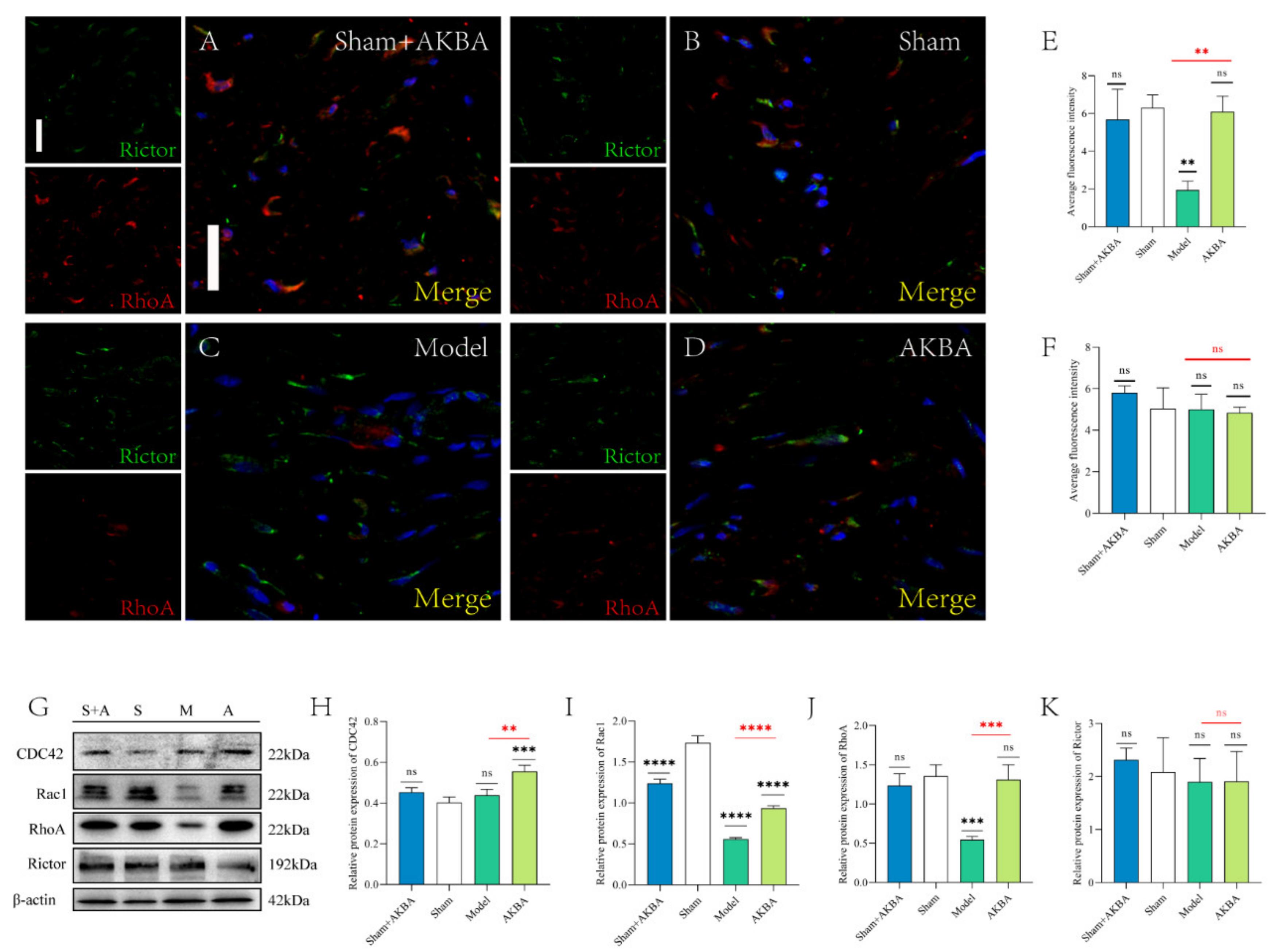
2.4. Protein Expression of The Injured Sciatic Nerve after 5 d of AKBA Intervention
2.5. Protein Expression of The Injured Sciatic Nerve 14 d after AKBA Intervention
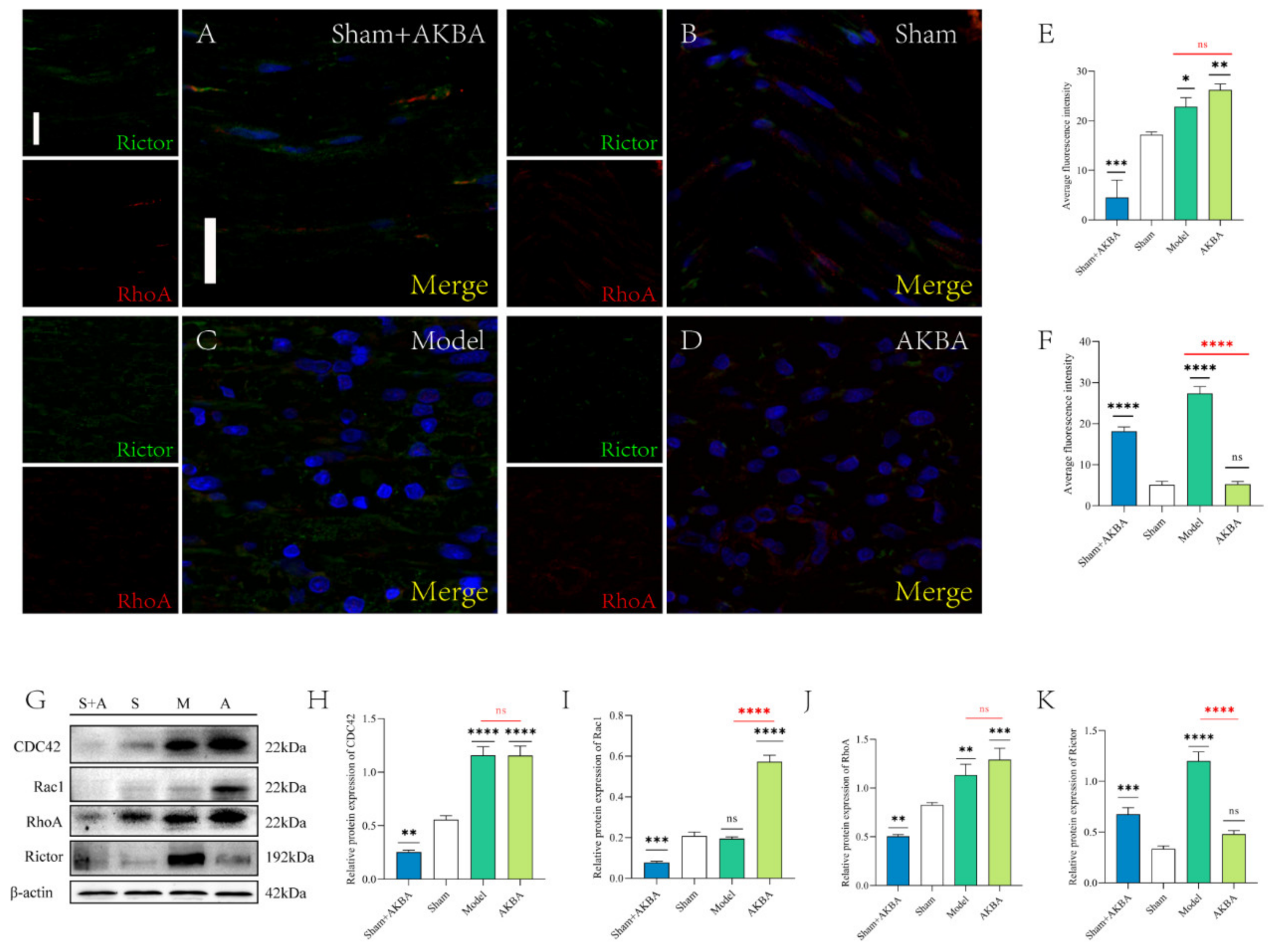
2.6. Protein Expression of The Injured Sciatic Nerve after 28 d of AKBA Intervention
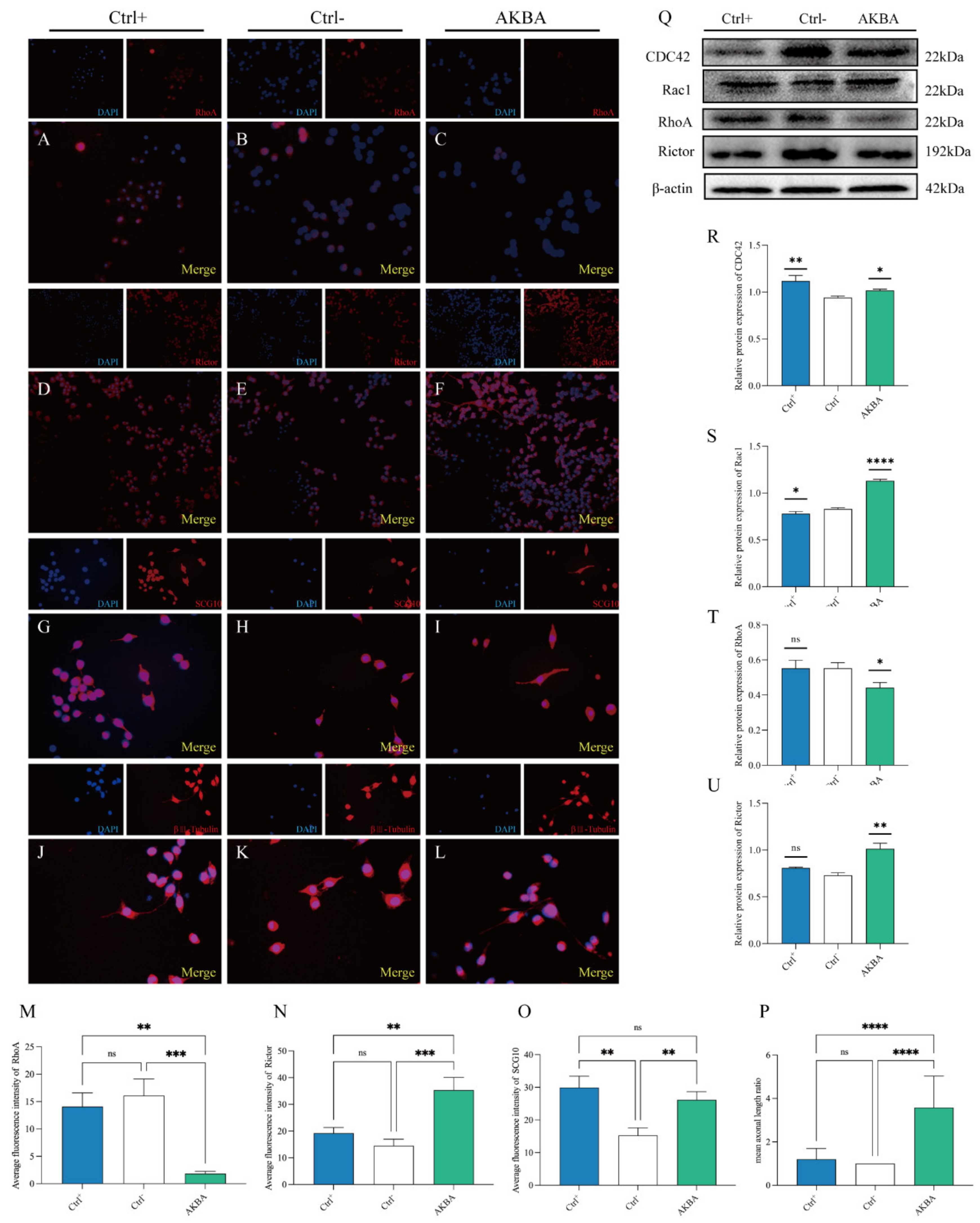
2.7. Changes in Protein Expression after AKBA Action on PC12 Cells
3. Discussion
3.1. Effect of AKBA Intervention on the Bioinformatics of Late SNI
3.2. Effect of AKBA Intervention on Regeneration of Injured Sciatic Nerve
3.3. Effect of AKBA Intervention on Axon-Related Protein Expression in Injured Sciatic Nerve
4. Methods and Materials
4.1. Establishment of a Model of Sciatic Nerve Injury in Rats
4.2. Experimental Grouping and Drug Administration in Rats
4.3. Proteomic Analysis of Rat Sciatic Nerve Samples
4.4. Western Blot Assay of Rat Sciatic Nerve Samples
4.5. Toluidine Blue Staining of Rat Sciatic Nerve Samples
4.6. Immunofluorescence Detection of Rat Sciatic Nerve Samples
4.7. Cytotoxicity Assay of PC12 Cells
4.8. Effect of AKBA on PC12 Cells by Immunofluorescence Detection and Immunoblotting
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ding, Y.; Qiao, Y.; Wang, M.; Zhang, H.; Li, L.; Zhang, Y.; Ge, J.; Song, Y.; Li, Y.; Wen, A. Enhanced neuroprotection of acetyl-11-keto-β-boswellic acid (AKBA)-loaded O-carboxymethyl chitosan nanoparticles through antioxidant and anti-inflammatory pathways. Mol. Neurobiol. 2016, 53, 3842–3853. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Al-Harrasi, A.; Rehman, N.U.; Sarwar, R.; Ahmad, T.; Ghaffar, R.; Khan, H.; Al-Amri, I.; Csuk, R.; Al-Rawahi, A. Loading AKBA on surface of silver nanoparticles to improve their sedative-hypnotic and anti-inflammatory efficacies. Nanomedicine 2019, 14, 2783–2798. [Google Scholar] [CrossRef] [PubMed]
- Minj, E.; Upadhayay, S.; Mehan, S. Nrf2/HO-1 Signaling Activator Acetyl-11-keto-beta Boswellic Acid (AKBA)-Mediated Neuroprotection in Methyl Mercury-Induced Experimental Model of ALS. Neurochem. Res. 2021, 46, 2867–2884. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Chen, M.; Wang, M.; Wang, M.; Zhang, T.; Park, J.; Zhu, Y.; Guo, C.; Jia, Y.; Li, Y.; et al. Neuroprotection by acetyl-11-keto-β-boswellic acid, in ischemic brain injury involves the Nrf2/HO-1 defense pathway. Sci. Rep. 2014, 4, 7002. [Google Scholar] [CrossRef] [PubMed]
- Forouzanfar, F.; Hosseinzadeh, H.; Ebrahimzadeh Bideskan, A.; Sadeghnia, H.R. Aqueous and ethanolic extracts of Boswellia serrata protect against focal cerebral ischemia and reperfusion injury in rats. Phytother. Res. 2016, 30, 1954–1967. [Google Scholar] [CrossRef]
- Jiang, X.W.; Zhang, B.Q.; Qiao, L.; Liu, L.; Wang, X.W.; Yu, W.H. Acetyl-11-keto-beta-boswellic acid extracted from Boswellia serrata promotes Schwann cell proliferation and sciatic nerve function recovery. Neural Regen. Res. 2018, 13, 484–491. [Google Scholar] [CrossRef]
- Jiang, X.; Wang, Y.; Zhang, B.; Fei, X.; Gou, X.; Jia, Y.; Yu, W. Acetyl-11-keto-beta-boswellic acid regulates the repair of rat sciatic nerve injury by promoting the proliferation of Schwann cells. Life Sci. 2019, 254, 116887. [Google Scholar] [CrossRef]
- Matsukawa, T.; Morita, K.; Omizu, S.; Kato, S.; Koriyama, Y. Mechanisms of RhoA inactivation and CDC42 and Rac1 activation during zebrafish optic nerve regeneration. Neurochem. Int. 2018, 112, 71–80. [Google Scholar] [CrossRef]
- Hu, J.; Selzer, M.E. RhoA as a target to promote neuronal survival and axon regeneration. Neural Regen. Res. 2017, 12, 525. [Google Scholar]
- Fournier, A.E.; Takizawa, B.T.; Strittmatter, S.M. Rho kinase inhibition enhances axonal regeneration in the injured CNS. J. Neurosci. 2003, 23, 1416–1423. [Google Scholar] [CrossRef]
- Quarta, S.; Camprubí-Robles, M.; Schweigreiter, R.; Matusica, D.; Haberberger, R.V.; Proia, R.L.; Bandtlow, C.E.; Ferrer-Montiel, A.; Kress, M. Sphingosine-1-phosphate and the S1P3 receptor initiate neuronal retraction via RhoA/ROCK associated with CRMP2 phosphorylation. Front. Mol. Neurosci. 2017, 10, 317. [Google Scholar] [CrossRef]
- Jiu, Y.; Peränen, J.; Schaible, N.; Cheng, F.; Eriksson, J.E.; Krishnan, R.; Lappalainen, P. Vimentin intermediate filaments control actin stress fiber assembly through GEF-H1 and RhoA. J. Cell Sci. 2017, 130, 892–902. [Google Scholar] [CrossRef]
- Patel, S.; McKeon, D.; Sao, K.; Yang, C.; Naranjo, N.M.; Svitkina, T.M.; Petrie, R.J. Myosin II and Arp2/3 cross-talk governs intracellular hydraulic pressure and lamellipodia formation. Mol. Biol. Cell 2021, 32, 579–589. [Google Scholar] [CrossRef]
- Marsh, E.; Gonzalez, D.G.; Lathrop, E.A.; Boucher, J.; Greco, V. Positional stability and membrane occupancy define skin fibroblast homeostasis in vivo. Cell 2018, 175, 1620–1633.e1613. [Google Scholar] [CrossRef]
- Adams, J.C.; Schwartz, M.A. Stimulation of fascin spikes by thrombospondin-1 is mediated by the GTPases Rac and Cdc42. J. Cell Biol. 2000, 150, 807–822. [Google Scholar] [CrossRef]
- Aspenström, P. The intrinsic gdp/gtp exchange activities of Cdc42 and rac1 are critical determinants for their specific effects on mobilization of the actin filament system. Cells 2019, 8, 759. [Google Scholar] [CrossRef]
- García-Ortiz, A.; Serrador, J.M. ERM proteins at the crossroad of leukocyte polarization, migration and intercellular adhesion. Int. J. Mol. Sci. 2020, 21, 1502. [Google Scholar] [CrossRef]
- Pardo-Pastor, C.; Rubio-Moscardo, F.; Vogel-González, M.; Serra, S.A.; Afthinos, A.; Mrkonjic, S.; Destaing, O.; Abenza, J.F.; Fernández-Fernández, J.M.; Trepat, X.; et al. Piezo2 channel regulates RhoA and actin cytoskeleton to promote cell mechanobiological responses. Proc. Natl. Acad. Sci. USA 2018, 115, 1925–1930. [Google Scholar] [CrossRef]
- Chen, H.-C.; Fong, T.-H.; Hsu, P.-W.; Chiu, W.-T. Multifaceted effects of rapamycin on functional recovery after spinal cord injury in rats through autophagy promotion, anti-inflammation, and neuroprotection. J. Surg. Res. 2013, 179, e203–e210. [Google Scholar] [CrossRef]
- Rajnicek, A.M.; Foubister, L.E.; McCaig, C.D. Prioritising guidance cues: Directional migration induced by substratum contours and electrical gradients is controlled by a rho/cdc42 switch. Dev. Biol. 2007, 312, 448–460. [Google Scholar] [CrossRef]
- Sekiguchi, A.; Kanno, H.; Ozawa, H.; Yamaya, S.; Itoi, E. Rapamycin promotes autophagy and reduces neural tissue damage and locomotor impairment after spinal cord injury in mice. J. Neurotrauma 2012, 29, 946–956. [Google Scholar] [CrossRef] [PubMed]
- Thomanetz, V.; Angliker, N.; Cloëtta, D.; Lustenberger, R.M.; Schweighauser, M.; Oliveri, F.; Suzuki, N.; Rüegg, M.A. Ablation of the mTORC2 component rictor in brain or Purkinje cells affects size and neuron morphology. J. Cell Biol. 2013, 201, 293–308. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Lu, N.; Ding, Y.; Wang, Y.; Chan, L.T.; Wang, X.; Gao, X.; Jiang, S.; Liu, K. Rapamycin-resistant mTOR activity is required for sensory axon regeneration induced by a conditioning lesion. eNeuro 2016, 3, ENEURO.0358-16. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Zhou, P.; Liu, X.; Li, J.; Wan, Y.; Liu, S.; Wei, F. Overexpression of Rictor in the injured spinal cord promotes functional recovery in a rat model of spinal cord injury. FASEB J. 2020, 34, 6984–6998. [Google Scholar] [CrossRef] [PubMed]
- Grier, M.D.; West, K.L.; Kelm, N.D.; Fu, C.; Does, M.D.; Parker, B.; McBrier, E.; Lagrange, A.H.; Ess, K.C.; Carson, R.P. Loss of mTORC2 signaling in oligodendrocyte precursor cells delays myelination. PLoS ONE 2017, 12, e0188417. [Google Scholar] [CrossRef]
- Kiss, A.; Fischer, I.; Kleele, T.; Misgeld, T.; Propst, F. Neuronal growth cone size-dependent and-independent parameters of microtubule polymerization. Front. Cell. Neurosci. 2018, 12, 195. [Google Scholar] [CrossRef]
- Bernabo, P.; Tebaldi, T.; Groen, E.J.N.; Lane, F.M.; Perenthaler, E.; Mattedi, F.; Newbery, H.J.; Zhou, H.; Zuccotti, P.; Potrich, V.; et al. In vivo translatome profiling in spinal muscular atrophy reveals a role for SMN protein in ribosome biology. Cell Rep. 2017, 21, 953–965. [Google Scholar] [CrossRef]
- Terenzio, M.; Koley, S.; Samra, N.; Rishal, I.; Zhao, Q.; Sahoo, P.K.; Urisman, A.; Marvaldi, L.; Oses-Prieto, J.A.; Forester, C.; et al. Locally translated mTOR controls axonal local translation in nerve injury. Science 2018, 359, 1416–1421. [Google Scholar] [CrossRef]
- Spillane, M.; Ketschek, A.; Merianda, T.T.; Twiss, J.L.; Gallo, G. Mitochondria coordinate sites of axon branching through localized intra-axonal protein synthesis. Cell Rep. 2013, 5, 1564–1575. [Google Scholar] [CrossRef]
- Schachtrup, C.; Ryu, J.K.; Helmrick, M.J.; Vagena, E.; Galanakis, D.K.; Degen, J.L.; Margolis, R.U.; Akassoglou, K. Fibrinogen triggers astrocyte scar formation by promoting the availability of active TGF-β after vascular damage. J. Neurosci. 2010, 30, 5843–5854. [Google Scholar] [CrossRef]
- Dupraz, S.; Hilton, B.J.; Husch, A.; Santos, T.E.; Coles, C.H.; Stern, S.; Brakebusch, C.; Bradke, F. RhoA Controls Axon Extension Independent of Specification in the Developing Brain. Curr. Biol. 2019, 29, 3874–3886.e3879. [Google Scholar] [CrossRef]
- Wen, J.; Tan, D.; Li, L.; Wang, X.; Pan, M.; Guo, J. RhoA regulates Schwann cell differentiation through JNK pathway. Exp. Neurol. 2018, 308, 26–34. [Google Scholar] [CrossRef]
- Zhang, G.; Hu, J.; Rodemer, W.; Li, S.; Selzer, M.E. RhoA activation in axotomy-induced neuronal death. Exp. Neurol. 2018, 306, 76–91. [Google Scholar] [CrossRef]
- Chen, W.; Chen, W.; Chen, S.; Uosef, A.; Ghobrial, R.M.; Kloc, M. Fingolimod (FTY720) prevents chronic rejection of rodent cardiac allografts through inhibition of the RhoA pathway. Transpl. Immunol. 2021, 65, 101347. [Google Scholar] [CrossRef]
- Zhang, L.; You, J.; Sidhu, J.; Tejpal, N.; Ganachari, M.; Skelton, T.S.; Kloc, M.; Li, X.C.; Ghobrial, R.M. Abrogation of chronic rejection in rat model system involves modulation of the mTORC1 and mTORC2 pathways. Transplantation 2013, 96, 782–790. [Google Scholar] [CrossRef]
- Chen, G.; Li, X.; Zhu, H.; Wu, H.; He, D.; Shi, L.; Wei, F.; Liu, X.; Chen, N.; Liu, S. Transplanting neurofibromatosis-1 gene knockout neural stem cells improve functional recovery in rats with spinal cord injury by enhancing the mTORC2 pathway. Exp. Mol. Med. 2022, 54, 1766–1777. [Google Scholar] [CrossRef]
- Jin, Z.; Strittmatter, S.M. Rac1 mediates collapsin-1-induced growth cone collapse. J. Neurosci. 1997, 17, 6256–6263. [Google Scholar] [CrossRef]
- Liu, B.; Xin, W.; Tan, J.R.; Zhu, R.P.; Li, T.; Wang, D.; Kan, S.S.; Xiong, D.K.; Li, H.H.; Zhang, M.M.; et al. Myelin sheath structure and regeneration in peripheral nerve injury repair. Proc. Natl. Acad. Sci. USA 2019, 116, 22347–22352. [Google Scholar] [CrossRef]
- Luís, A.L.; Rodrigues, J.M.; Geuna, S.; Amado, S.; Simões, M.J.; Fregnan, F.; Ferreira, A.J.; Veloso, A.P.; Armada-da-Silva, P.A.; Varejão, A.S.; et al. Neural cell transplantation effects on sciatic nerve regeneration after a standardized crush injury in the rat. Microsurgery 2008, 28, 458–470. [Google Scholar] [CrossRef]
- Wang, Y.; Xiong, Z.L.; Ma, X.L.; Zhou, C.; Huo, M.H.; Jiang, X.W.; Yu, W.H. Acetyl-11-keto-beta-boswellic acid promotes sciatic nerve repair after injury: Molecular mechanism. Neural Regen. Res. 2022, 17, 2778–2784. [Google Scholar] [CrossRef]
- Chi, Q.; Hu, X.; Liu, Z.; Han, Y.; Tao, D.; Xu, S.; Li, S. H2S exposure induces cell death in the broiler thymus via the ROS-initiated JNK/MST1/FOXO1 pathway. Ecotoxicol. Environ. Saf. 2021, 222, 112488. [Google Scholar] [CrossRef] [PubMed]
- Catenaccio, A.; Court, F.A. Teased Fiber Preparation of Myelinated Nerve Fibers from Peripheral Nerves for Vital Dye Staining and Immunofluorescence Analysis. Methods Mol. Biol. 2018, 1739, 329–337. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Application | Cat. No | Brands |
|---|---|---|---|
| RhoA | WB, IF | BM4479 | Boster, China |
| CDC42 | WB | bs-3555R | Bioss, China |
| Rac1 | WB | A7720 | Abclonal, China |
| ApoE | WB | WL03172 | Wanlei, China |
| Rictor | WB | WL04494 | Wanlei, China |
| Rictor | IF | MB0063 | ABmart, China |
| SCG10 | IF | A04729–2 | Boster, China |
| βIII-Tubulin | IF | A17913 | Abclonal, China |
| P16 ARC | WB | AF1069 | Beyotime, China |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Xiong, Z.; Zhou, C.; Zhang, Q.; Liu, S.; Dong, S.; Jiang, X.; Yu, W. AKBA Promotes Axonal Regeneration via RhoA/Rictor to Repair Damaged Sciatic Nerve. Int. J. Mol. Sci. 2022, 23, 15903. https://doi.org/10.3390/ijms232415903
Wang Y, Xiong Z, Zhou C, Zhang Q, Liu S, Dong S, Jiang X, Yu W. AKBA Promotes Axonal Regeneration via RhoA/Rictor to Repair Damaged Sciatic Nerve. International Journal of Molecular Sciences. 2022; 23(24):15903. https://doi.org/10.3390/ijms232415903
Chicago/Turabian StyleWang, Yao, Zongliang Xiong, Chong Zhou, Qiyuan Zhang, Shuang Liu, Sainan Dong, Xiaowen Jiang, and Wenhui Yu. 2022. "AKBA Promotes Axonal Regeneration via RhoA/Rictor to Repair Damaged Sciatic Nerve" International Journal of Molecular Sciences 23, no. 24: 15903. https://doi.org/10.3390/ijms232415903
APA StyleWang, Y., Xiong, Z., Zhou, C., Zhang, Q., Liu, S., Dong, S., Jiang, X., & Yu, W. (2022). AKBA Promotes Axonal Regeneration via RhoA/Rictor to Repair Damaged Sciatic Nerve. International Journal of Molecular Sciences, 23(24), 15903. https://doi.org/10.3390/ijms232415903