
Rat Hepatocytes Mitigate Cadmium Toxicity by Forming Annular Gap Junctions and Degrading Them via Endosome–Lysosome Pathway

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Cd-Induced Microtubule Damage Promotes Cx43 Internalization and Degradation Accompanied by Increased Phosphorylation at Ser373 Site
2.2. Cd-Induced Damage to Microtubules Promotes Cx43 Internalization
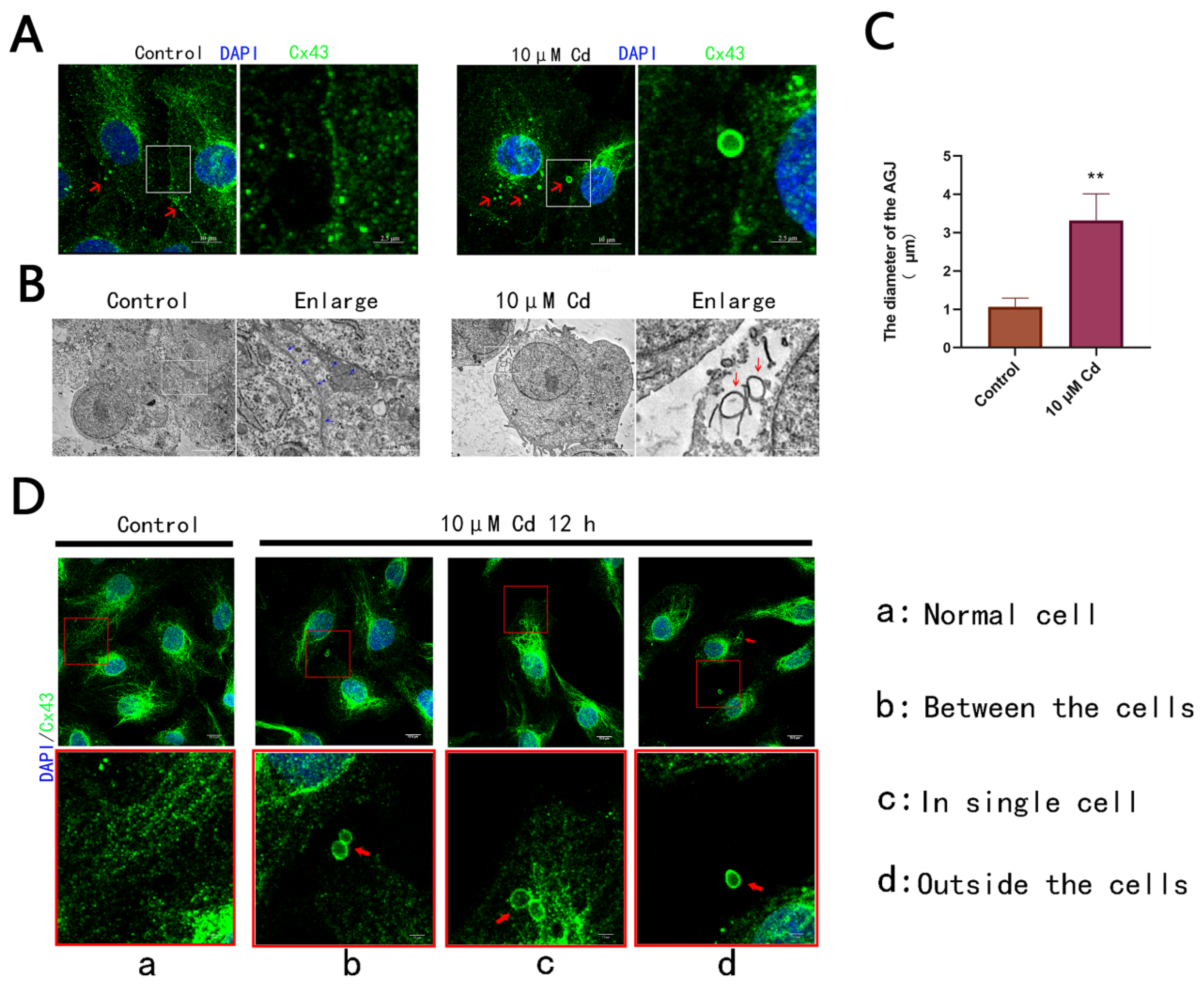
2.3. Cd Exposure Induces the Formation of AGJs in Rat Hepatocytes
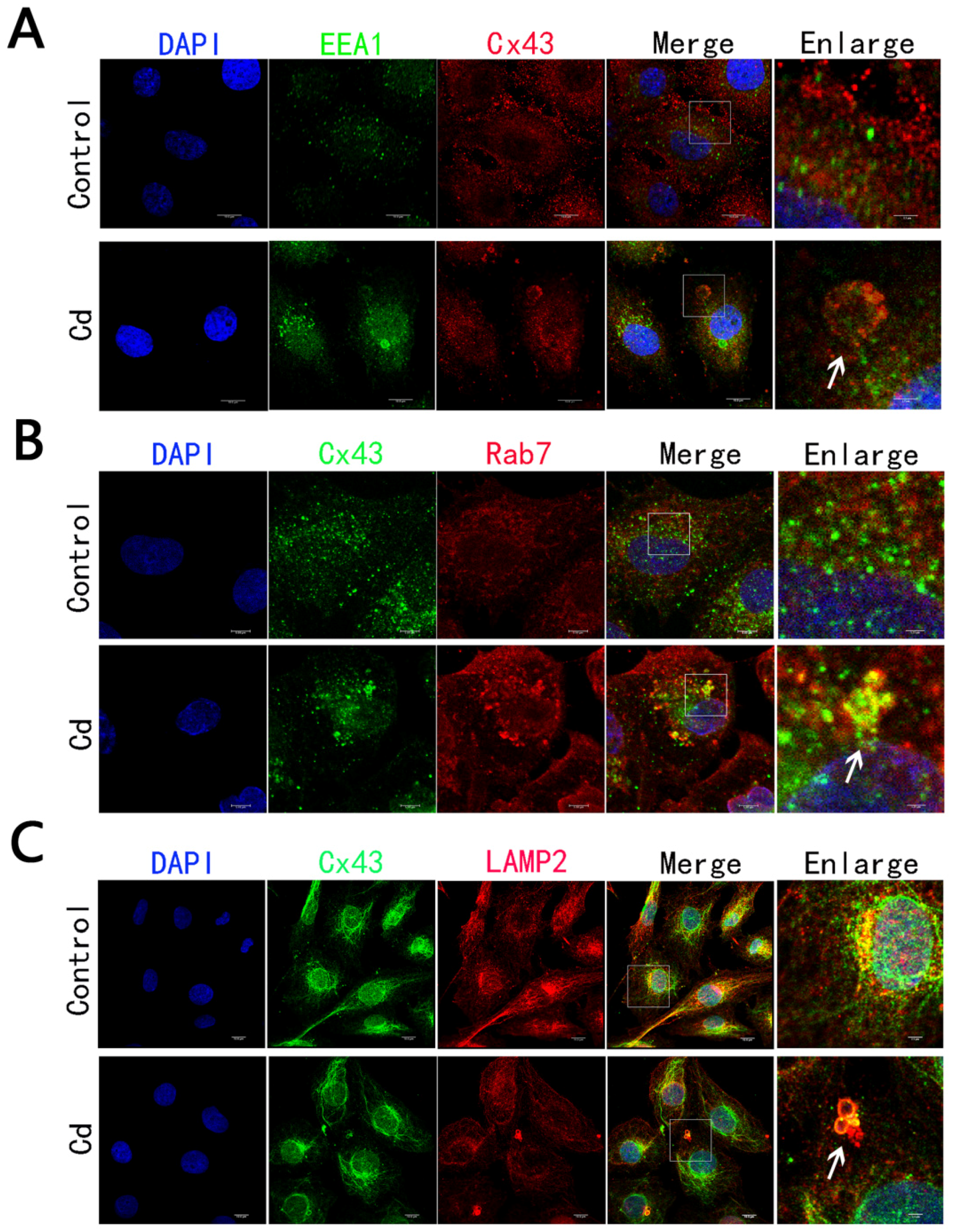
2.4. Cd Treatment Caused the Degradation of Internalized AGJ through Endosome-Lysosome Pathway
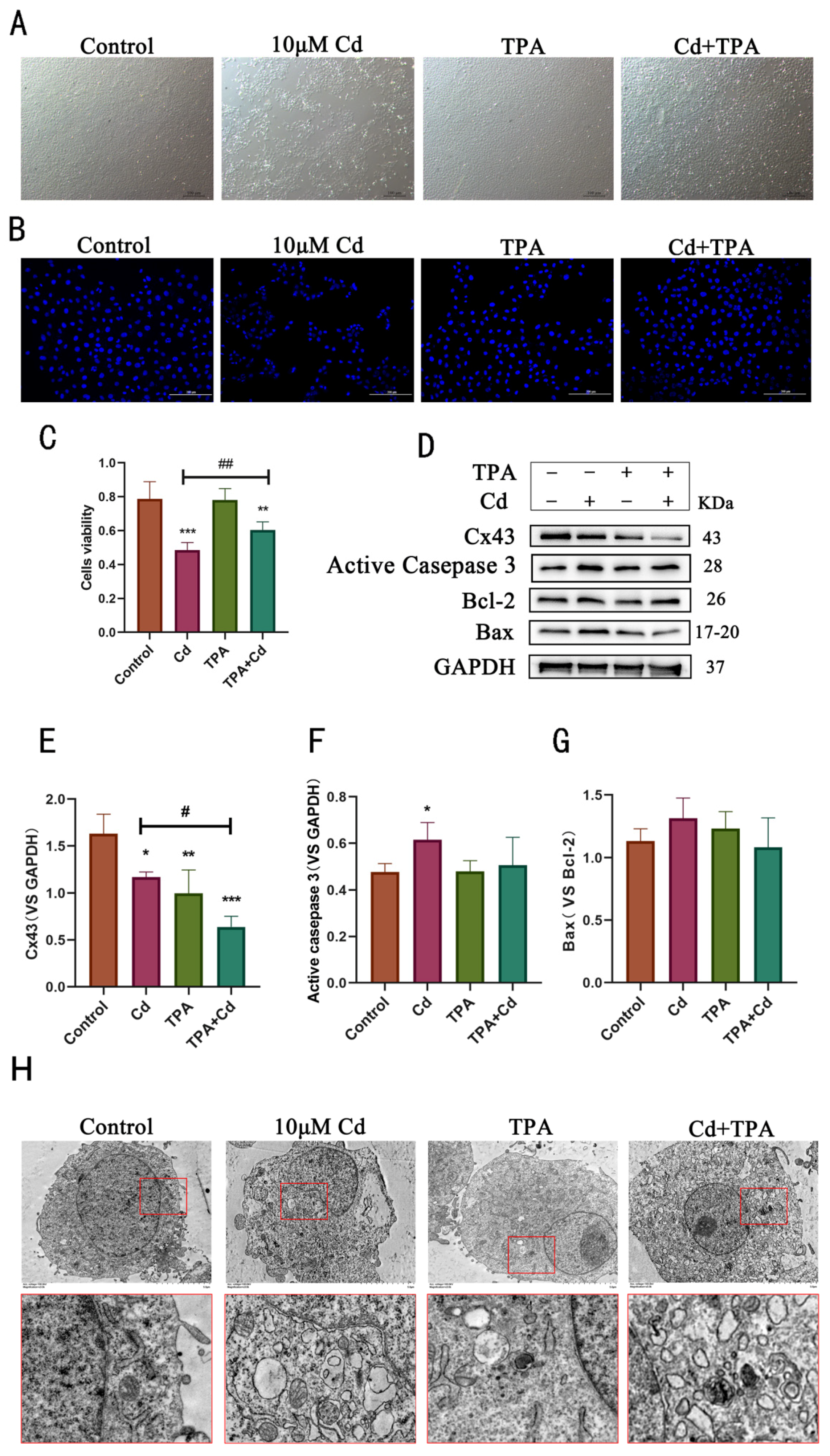
2.5. Internalization of Cx43 Protects Rat Hepatocytes from Cd-Induced Apoptosis and Alleviates Cell Damage
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture and Morphological Observation
4.3. Immunofluorescence Assay
- Different concentrations of Cd (0, 5, 10, and 20 µM) were added to 24-well plates to co-stain Cx43 and microfilaments.
- Different concentrations of Cd (0, 5, 10, and 20 µM) were added to 24-well plates to co-stain Cx43 and α-tubulin.
- The control, Cd, nocodazole, and nocodazole + Cd were added to 24-well plates. Cells were then stained for Cx43.
- Three experimental groups were set up in 24-well plates. The control and Cd were added to two wells in each group, respectively. Cells of the three groups were then stained with antibodies from different species against Cx43 and EEA1, Cx43 and Rab7a, Cx43, and LAMP2.
- The control, Cd, TPA (0.5 µM in DMSO), and Cd + TPA were added to 24-well plates. The cells were then stained with Hochest 33258.
4.4. Transmission Electron Microscopy
4.5. Western Blot Assay
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Turner, A. Cadmium pigments in consumer products and their health risks. Sci. Total Environ. 2019, 657, 1409–1418. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Ma, Z.; van der Kuijp, T.J.; Yuan, Z.; Huang, L. A review of soil heavy metal pollution from mines in China: Pollution and health risk assessment. Sci. Total Environ. 2014, 468–469, 843–853. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.J.; Ding, K.B.; Zhang, P.; Qiu, H.; Cloquet, C.; Wen, H.J.; Morel, J.L.; Qiu, R.L.; Tang, Y.T. Cadmium stable isotope variation in a mountain area impacted by acid mine drainage. Sci. Total Environ. 2019, 646, 696–703. [Google Scholar] [CrossRef] [PubMed]
- Huff, J.; Lunn, R.M.; Waalkes, M.P.; Tomatis, L.; Infante, P.F. Cadmium-induced cancers in animals and in humans. Int. J. Occup. Environ. Health 2007, 13, 202–212. [Google Scholar] [CrossRef]
- Luo, G.; Han, Z.; Xiong, J.; He, Y.; Liao, J.; Wu, P. Heavy metal pollution and ecological risk assessment of tailings in the Qinglong Dachang antimony mine, China. Environ. Sci. Pollut. Res. Int. 2021, 28, 33491–33504. [Google Scholar] [CrossRef]
- Outa, J.O.; Kowenje, C.O.; Plessl, C.; Jirsa, F. Distribution of arsenic, silver, cadmium, lead and other trace elements in water, sediment and macrophytes in the Kenyan part of Lake Victoria: Spatial, temporal and bioindicative aspects. Environ. Sci. Pollut. Res. Int. 2020, 27, 1485–1498. [Google Scholar] [CrossRef]
- Sharma, T.; Litoria, P.K.; Bajwa, B.S.; Kaur, I. Appraisal of groundwater quality and associated risks in Mansa district (Punjab, India). Environ. Monit. Assess. 2021, 193, 159. [Google Scholar] [CrossRef]
- Kim, J.J.; Kim, Y.S.; Kumar, V. Heavy metal toxicity: An update of chelating therapeutic strategies. J. Trace Elem. Med. Biol. 2019, 54, 226–231. [Google Scholar] [CrossRef]
- Ahamed, M.; Akhtar, M.J.; Khan, M.A.M.; Alhadlaq, H.A. Reduced graphene oxide mitigates cadmium-induced cytotoxicity and oxidative stress in HepG2 cells. Food Chem. Toxicol. 2020, 143, 111515. [Google Scholar] [CrossRef]
- Yuan, J.; Huang, X.; Gu, J.; Yuan, Y.; Liu, Z.; Zou, H.; Bian, J. Honokiol reduces cadmium-induced oxidative injury and endosomal/lysosomal vacuolation via protecting mitochondrial function in quail (Coturnix japonica) liver tissues. Sci. Total Environ. 2022, 857, 159626. [Google Scholar] [CrossRef]
- Sughis, M.; Penders, J.; Haufroid, V.; Nemery, B.; Nawrot, T.S. Bone resorption and environmental exposure to cadmium in children: A cross--sectional study. Environ. Health 2011, 10, 104. [Google Scholar] [CrossRef] [PubMed]
- Severs, N.J.; Rothery, S.; Dupont, E.; Coppen, S.R.; Yeh, H.I.; Ko, Y.S.; Matsushita, T.; Kaba, R.; Halliday, D. Immunocytochemical analysis of connexin expression in the healthy and diseased cardiovascular system. Microsc. Res. Tech. 2001, 52, 301–322. [Google Scholar] [CrossRef] [PubMed]
- Kelly, J.J.; Simek, J.; Laird, D.W. Mechanisms linking connexin mutations to human diseases. Cell Tissue Res. 2015, 360, 701–721. [Google Scholar] [CrossRef] [PubMed]
- Mugisho, O.O.; Green, C.R. The NLRP3 inflammasome in age-related eye disease: Evidence-based connexin hemichannel therapeutics. Exp. Eye Res. 2022, 215, 108911. [Google Scholar] [CrossRef]
- Willebrords, J.; Crespo Yanguas, S.; Maes, M.; Decrock, E.; Wang, N.; Leybaert, L.; da Silva, T.C.; Veloso Alves Pereira, I.; Jaeschke, H.; Cogliati, B.; et al. Structure, Regulation and Function of Gap Junctions in Liver. Cell Commun. Adhes. 2015, 22, 29–37. [Google Scholar] [CrossRef]
- Falk, M.M.; Baker, S.M.; Gumpert, A.M.; Segretain, D.; Buckheit, R.W. Gap Junction Turnover Is Achieved by the Internalization of Small Endocytic Double-Membrane Vesicles. Mol. Biol. Cell 2009, 20, 3342–3352. [Google Scholar] [CrossRef]
- Nickel, B.M.; DeFranco, B.H.; Gay, V.L.; Murray, S.A. Clathrin and Cx43 gap junction plaque endoexocytosis. Biochem. Biophys. Res. Commun. 2008, 374, 679–682. [Google Scholar] [CrossRef]
- Leithe, E.; Mesnil, M.; Aasen, T. The connexin 43 C-terminus: A tail of many tales. Biochim. Biophys. Acta 2018, 1860, 48–64. [Google Scholar] [CrossRef]
- Thévenin, A.F.; Kowal, T.J.; Fong, J.T.; Kells, R.M.; Fisher, C.G.; Falk, M.M. Proteins and mechanisms regulating gap-junction assembly, internalization, and degradation. Physiology 2013, 28, 93–116. [Google Scholar] [CrossRef]
- Solan, J.L.; Hingorani, S.R.; Lampe, P.D. Cx43 phosphorylation sites regulate pancreatic cancer metastasis. Oncogene 2021, 40, 1909–1920. [Google Scholar] [CrossRef]
- Tonkin, R.S.; Bowles, C.; Perera, C.J.; Keating, B.A.; Makker, P.G.S.; Duffy, S.S.; Lees, J.G.; Tran, C.; Don, A.S.; Fath, T.; et al. Attenuation of mechanical pain hypersensitivity by treatment with Peptide5, a connexin-43 mimetic peptide, involves inhibition of NLRP3 inflammasome in nerve-injured mice. Exp. Neurol. 2018, 300, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Boengler, K.; Rohrbach, S.; Weissmann, N.; Schulz, R. Importance of Cx43 for Right Ventricular Function. Int. J. Mol. Sci. 2021, 22, 987. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Zhang, Z.; Peng, X.; Wu, Y.; Zhu, Y.; Wang, L.; Zhou, H.; Li, T.; Liu, L. The protective effect of pericytes on vascular permeability after hemorrhagic shock and their relationship with Cx43. Front. Physiol. 2022, 13, 948541. [Google Scholar] [CrossRef] [PubMed]
- Amino, M.; Yoshioka, K.; Kamada, T.; Furusawa, Y. The Potential Application of Heavy Ion Beams in the Treatment of Arrhythmia: The Role of Radiation-Induced Modulation of Connexin43 and the Sympathetic Nervous System. Int. J. Part. Ther. 2018, 5, 140–150. [Google Scholar] [CrossRef]
- Falk, M.M.; Kells, R.M.; Berthoud, V.M. Degradation of connexins and gap junctions. FEBS Lett. 2014, 588, 1221–1229. [Google Scholar] [CrossRef]
- Dunn, C.A.; Su, V.; Lau, A.F.; Lampe, P.D. Activation of Akt, not connexin 43 protein ubiquitination, regulates gap junction stability. J. Biol. Chem. 2012, 287, 2600–2607. [Google Scholar] [CrossRef]
- Solan, J.L.; Lampe, P.D. Specific Cx43 phosphorylation events regulate gap junction turnover in vivo. FEBS Lett. 2014, 588, 1423–1429. [Google Scholar] [CrossRef]
- Girão, H.; Catarino, S.; Pereira, P. Eps15 interacts with ubiquitinated Cx43 and mediates its internalization. Exp. Cell Res. 2009, 315, 3587–3597. [Google Scholar] [CrossRef]
- Jovic, M.; Sharma, M.; Rahajeng, J.; Caplan, S.J. The early endosome: A busy sorting station for proteins at the crossroads. Histol. Histopathol. 2010, 25, 99–112. [Google Scholar]
- Naava, N.; Steve, C. The enigmatic endosome—Sorting the ins and outs of endocytic trafficking. J. Cell Sci. 2018, 131, jcs216499. [Google Scholar]
- Platta, H.W.; Stenmark, H. Endocytosis and signaling. Curr. Opin. Cell Biol. 2011, 23, 393–403. [Google Scholar] [CrossRef] [PubMed]
- Langemeyer, L.; Fröhlich, F.; Ungermann, C. Rab GTPase Function in Endosome and Lysosome Biogenesis. Trends Cell Biol. 2018, 28, 957–970. [Google Scholar] [CrossRef] [PubMed]
- Scott, C.C.; Vacca, F.; Gruenberg, J. Endosome maturation, transport and functions. Semin. Cell Dev. Biol. 2014, 31, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Xiang, X.; Qiu, R.; Yao, X.; Arst, H.N.; Peñalva, M.A.; Zhang, J.J.C.; Sciences, M.L. Cytoplasmic dynein and early endosome transport. Cell. Mol. Life Sci. 2015, 72, 3267–3280. [Google Scholar] [CrossRef]
- Skjeldal, F.M.; Strunze, S.; Bergeland, T.; Walseng, E.; Gregers, T.F.; Bakke, O. The fusion of early endosomes induces molecular-motor-driven tubule formation and fission. J. Cell Sci. 2012, 125, 1910–1919. [Google Scholar] [CrossRef]
- Leithe, E.; Kjenseth, A.; Sirnes, S.; Stenmark, H.; Brech, A.; Rivedal, E. Ubiquitylation of the gap junction protein connexin-43 signals its trafficking from early endosomes to lysosomes in a process mediated by Hrs and Tsg101. J. Cell Sci. 2009, 122, 3883–3893. [Google Scholar] [CrossRef]
- Piehl, M.; Lehmann, C.; Gumpert, A.; Denizot, J.P.; Segretain, D.; Falk, M.M. Internalization of Large Double-Membrane Intercellular Vesicles by a Clathrin-dependent Endocytic Process. Mol. Biol. Cell 2007, 18, 337–347. [Google Scholar] [CrossRef]
- Xia, Y.; Gong, K.Z.; Xu, M.; Zhang, Y.Y.; Guo, J.H.; Song, Y.; Zhang, P. Regulation of gap-junction protein connexin 43 by beta-adrenergic receptor stimulation in rat cardiomyocytes. Acta Pharmacol. Sin. 2009, 30, 928–934. [Google Scholar] [CrossRef]
- Mally, A.; Decker, M.; Bekteshi, M.; Dekant, W. Ochratoxin A alters cell adhesion and gap junction intercellular communication in MDCK cells. Toxicology 2006, 223, 15–25. [Google Scholar] [CrossRef]
- Baker, S.M.; Kim, N.; Gumpert, A.M.; Segretain, D.; Falk, M.M. Acute internalization of gap junctions in vascular endothelial cells in response to inflammatory mediator-induced G-protein coupled receptor activation. FEBS Lett. 2008, 582, 4039–4046. [Google Scholar] [CrossRef]
- Liao, C.K.; Jeng, C.J.; Wang, H.S.; Wang, S.H.; Wu, J.C. Lipopolysaccharide induces degradation of connexin43 in rat astrocytes via the ubiquitin-proteasome proteolytic pathway. PLoS ONE 2013, 8, e79350. [Google Scholar] [CrossRef]
- Hervé, J.; Derangeon, M.; Sarrouilhe, D.; Giepmans, B.; Bourmeyster, N. Gap junctional channels are parts of multiprotein complexes. Biochim. Biophys. Acta 2012, 1818, 1844–1865. [Google Scholar] [CrossRef] [PubMed]
- Cinzia, A.; Cynthia, R.; Gaelle, S.; Gabriel, C.; Angela, C.; Grintsevich, E.E.; Sosinsky, G.E.; Sorgen, P.L.; Michael, K. Connexin43 Forms Supramolecular Complexes through Non-Overlapping Binding Sites for Drebrin, Tubulin, and ZO-1. PLoS ONE 2016, 11, e0157073. [Google Scholar]
- Templeton, D.M.; Liu, Y. Effects of cadmium on the actin cytoskeleton in renal mesangial cells. Can. J. Physiol. Pharmacol. 2013, 91, 1–7. [Google Scholar] [CrossRef]
- Wang, L.; Yan, M.; Wu, S.; Mao, B.; Wong, C.K.C.; Ge, R.; Sun, F.; Cheng, C.Y. Microtubule Cytoskeleton and Spermatogenesis-Lesson From Studies of Toxicant Models. Toxicol. Sci. 2020, 177, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Schalper, K.A.; Carvajal-Hausdorf, D.; Oyarzo, M.P. Possible role of hemichannels in cancer. Front. Physiol. 2014, 5, 237. [Google Scholar] [CrossRef]
- Gemel, J.; Kilkus, J.; Dawson, G.; Beyer, E.C. Connecting Exosomes and Connexins. Cancers 2019, 11, 476. [Google Scholar] [CrossRef]
- Shimaoka, M.; Kawamoto, E.; Gaowa, A.; Okamoto, T.; Park, E.J. Connexins and Integrins in Exosomes. Cancers 2019, 11, 106. [Google Scholar] [CrossRef] [PubMed]
- Zou, H.; Wang, T.; Yuan, J.; Sun, J.; Yuan, Y.; Gu, J.; Liu, X.; Bian, J.; Liu, Z. Cadmium-induced cytotoxicity in mouse liver cells is associated with the disruption of autophagic flux via inhibiting the fusion of autophagosomes and lysosomes. Toxicol. Lett. 2020, 321, 32–43. [Google Scholar] [CrossRef]
- Saidi Brikci-Nigassa, A.; Clement, M.J.; Ha-Duong, T.; Adjadj, E.; Ziani, L.; Pastre, D.; Curmi, P.A.; Savarin, P. Phosphorylation controls the interaction of the connexin43 C-terminal domain with tubulin and microtubules. Biochemistry 2012, 51, 4331–4342. [Google Scholar] [CrossRef] [PubMed]
- Frankel, E.B.; Audhya, A. ESCRT-dependent cargo sorting at multivesicular endosomes. Semin. Cell Dev. Biol. 2018, 74, 4–10. [Google Scholar] [CrossRef] [PubMed]
- Zou, H.; Yuan, J.; Zhang, Y.; Wang, T.; Chen, Y.; Yuan, Y.; Bian, J.; Liu, Z. Gap Junction Intercellular Communication Negatively Regulates Cadmium-Induced Autophagy and Inhibition of Autophagic Flux in Buffalo Rat Liver 3A Cells. Front. Pharmacol. 2020, 11, 596046. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Zhao, Y.; Bai, Y.; Gu, J.; Yuan, Y.; Liu, X.; Liu, Z.; Zou, H.; Bian, J. Cadmium induces endosomal/lysosomal enlargement and blocks autophagy flux in rat hepatocytes by damaging microtubules. Ecotoxicol. Environ. Saf. 2021, 228, 112993. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, J.; Huang, X.; Zhao, Y.; Gu, J.; Yuan, Y.; Liu, Z.; Zou, H.; Bian, J. Rat Hepatocytes Mitigate Cadmium Toxicity by Forming Annular Gap Junctions and Degrading Them via Endosome–Lysosome Pathway. Int. J. Mol. Sci. 2022, 23, 15607. https://doi.org/10.3390/ijms232415607
Yuan J, Huang X, Zhao Y, Gu J, Yuan Y, Liu Z, Zou H, Bian J. Rat Hepatocytes Mitigate Cadmium Toxicity by Forming Annular Gap Junctions and Degrading Them via Endosome–Lysosome Pathway. International Journal of Molecular Sciences. 2022; 23(24):15607. https://doi.org/10.3390/ijms232415607
Chicago/Turabian StyleYuan, Junzhao, Xiaoqian Huang, Yumeng Zhao, Jianhong Gu, Yan Yuan, Zongping Liu, Hui Zou, and Jianchun Bian. 2022. "Rat Hepatocytes Mitigate Cadmium Toxicity by Forming Annular Gap Junctions and Degrading Them via Endosome–Lysosome Pathway" International Journal of Molecular Sciences 23, no. 24: 15607. https://doi.org/10.3390/ijms232415607
APA StyleYuan, J., Huang, X., Zhao, Y., Gu, J., Yuan, Y., Liu, Z., Zou, H., & Bian, J. (2022). Rat Hepatocytes Mitigate Cadmium Toxicity by Forming Annular Gap Junctions and Degrading Them via Endosome–Lysosome Pathway. International Journal of Molecular Sciences, 23(24), 15607. https://doi.org/10.3390/ijms232415607