
SP600125 Enhances Temperature-Controlled Repeated Thermal Stimulation-Induced Neurite Outgrowth in PC12-P1F1 Cells

 ,
,  , , ,
, , ,  ,
,  ,
,  and
and
Abstract
1. Introduction
2. Results
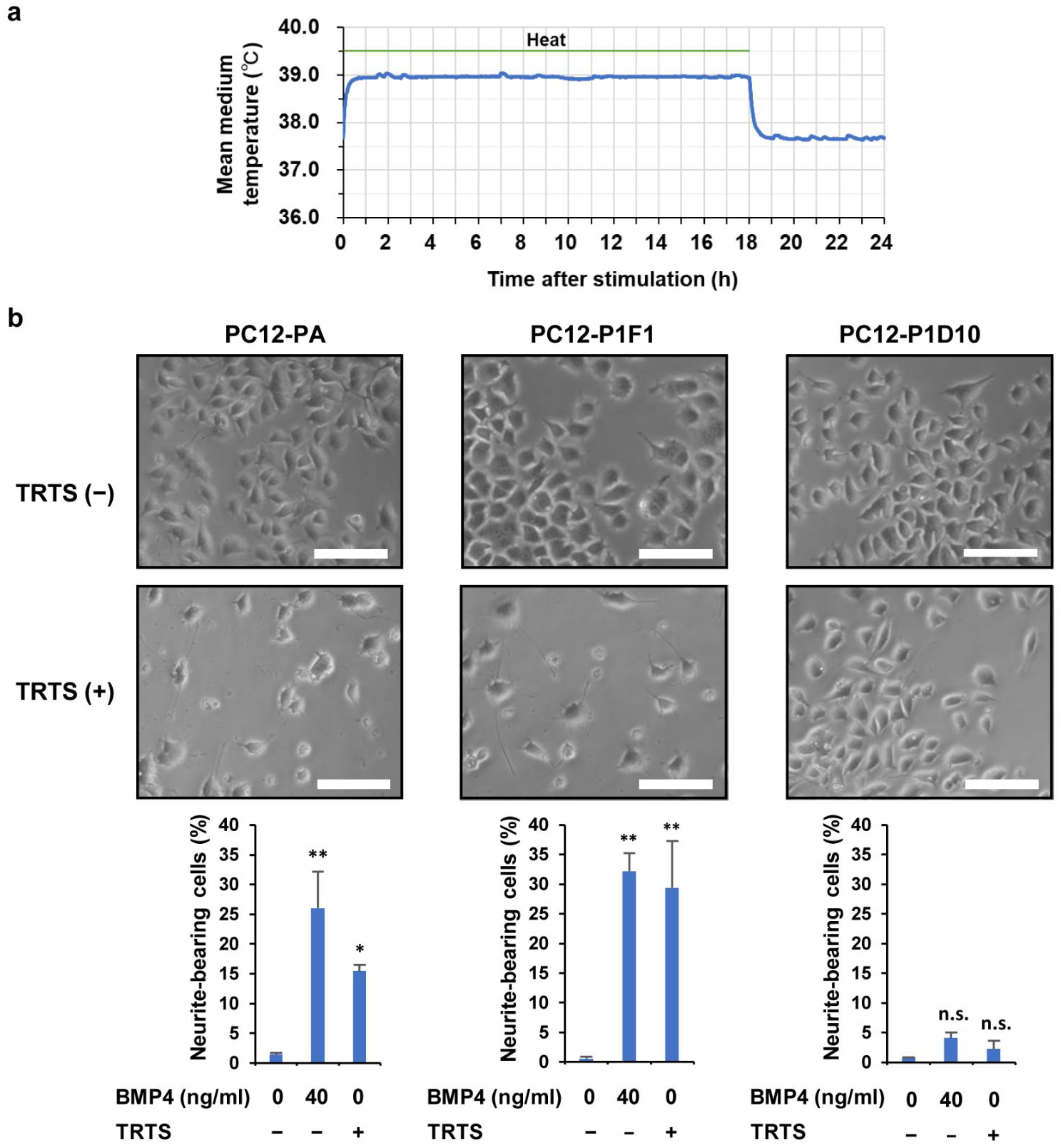
2.1. Regulation of TRTS
2.2. Promotion of TRTS-Mediated Neuritogenesis by SP600125 Treatment
2.3. Time Course and Morphological Characteristics of TRTS Plus SP60015-Induced Neuritogenesis
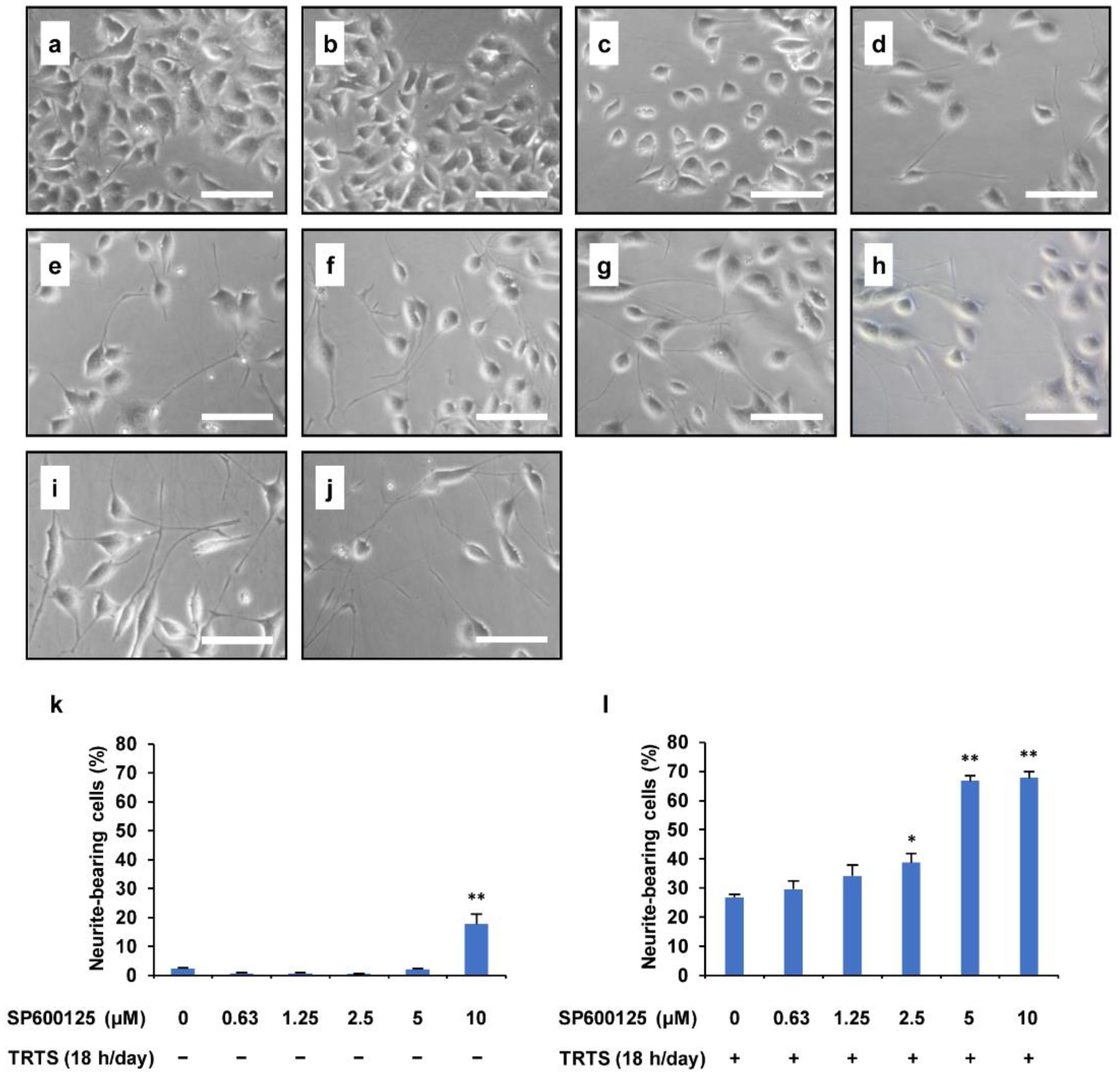
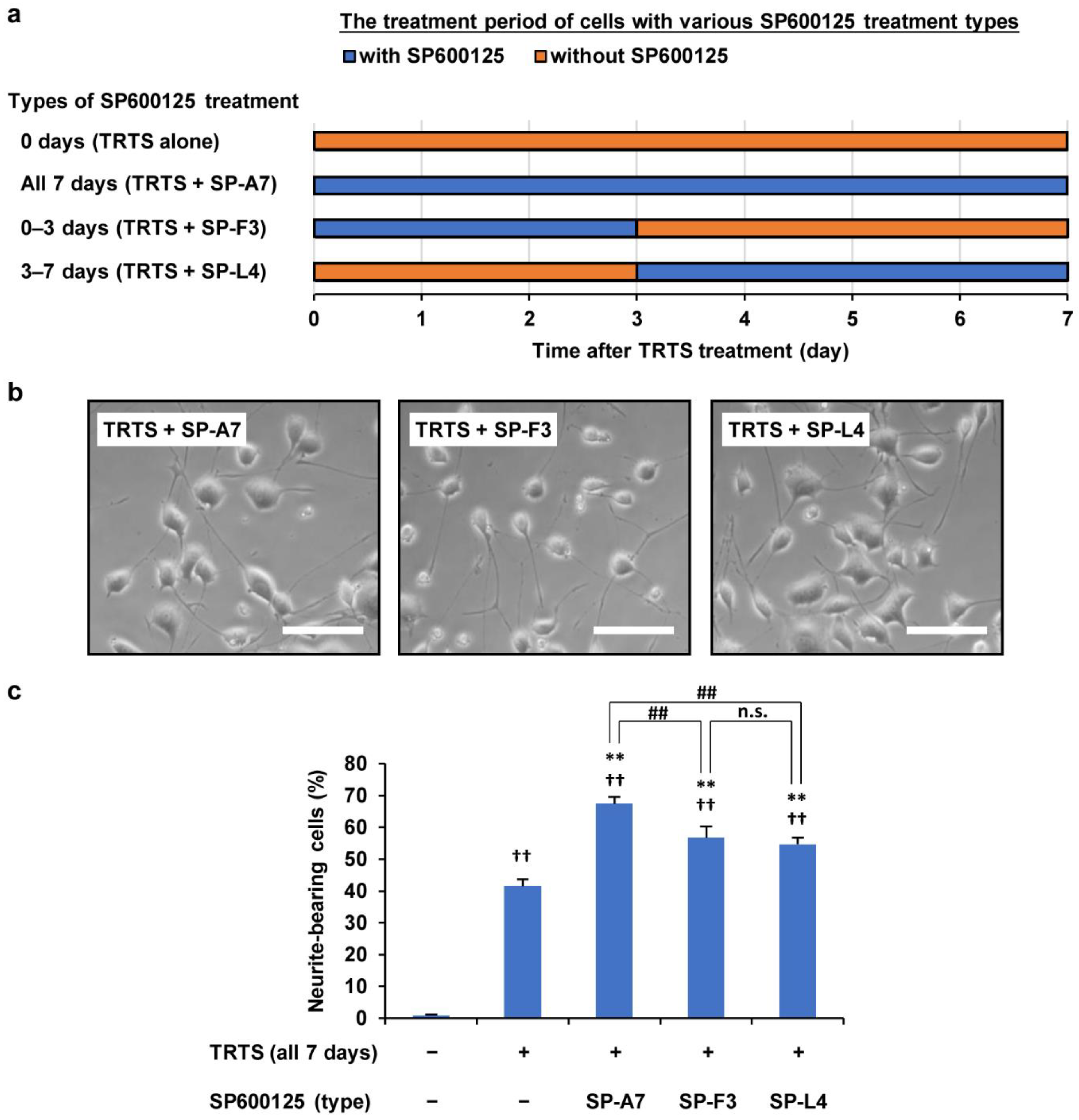
2.4. Effects of Dose and Treatment Time of SP600125 on TRTS-Induced Neuritogenesis
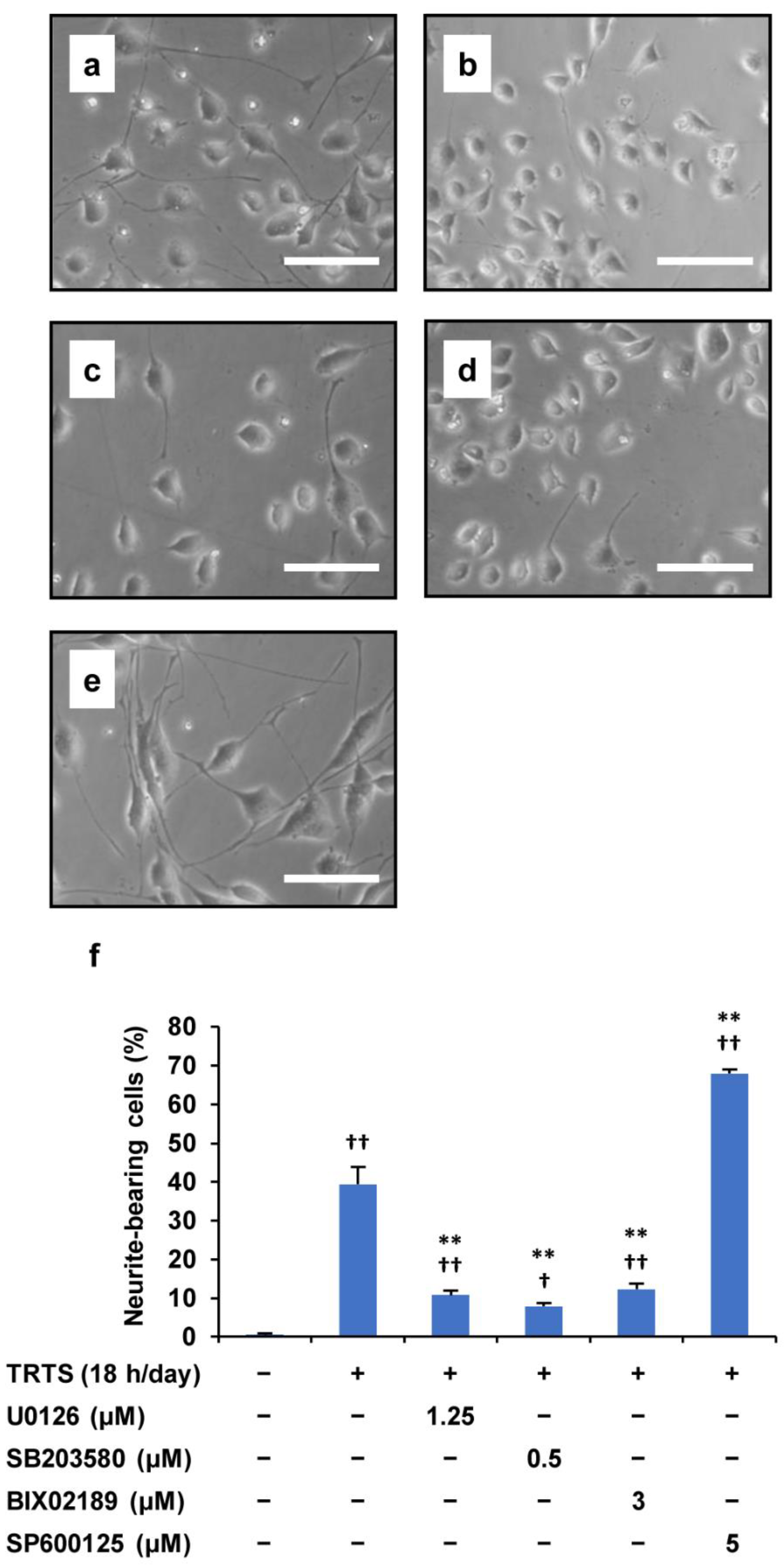
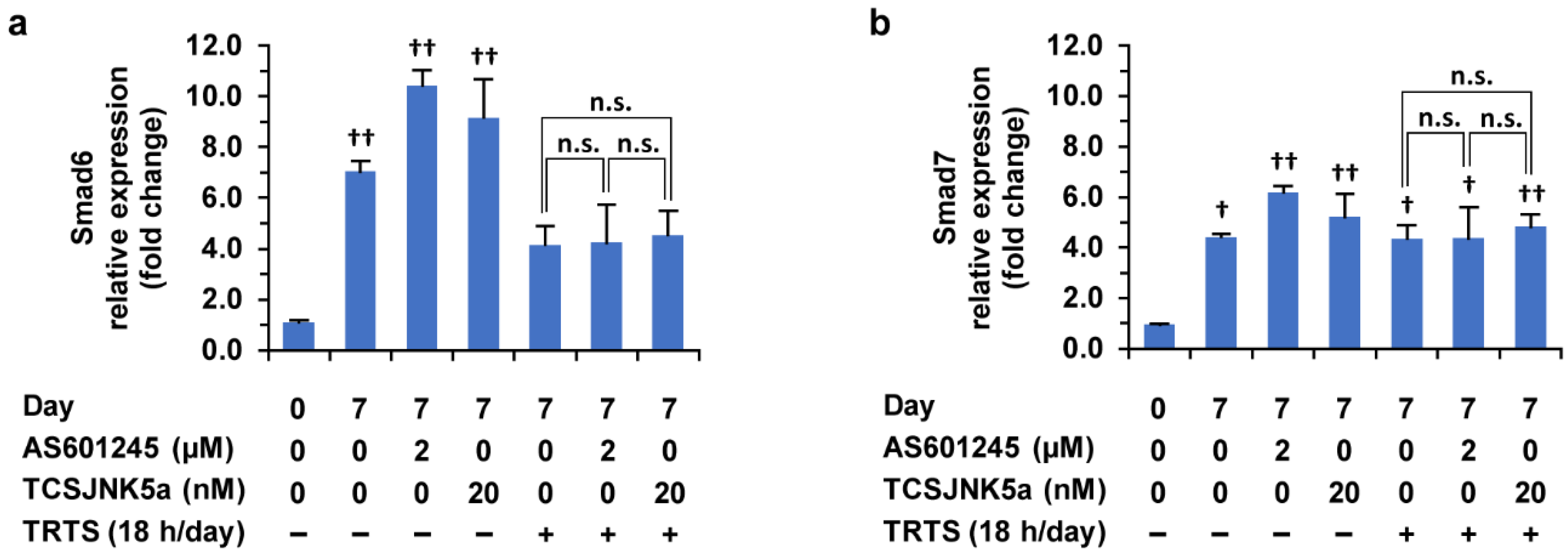
2.5. Relationship between the SP600125-Mediated Promoting Effect and the Target of SP600125
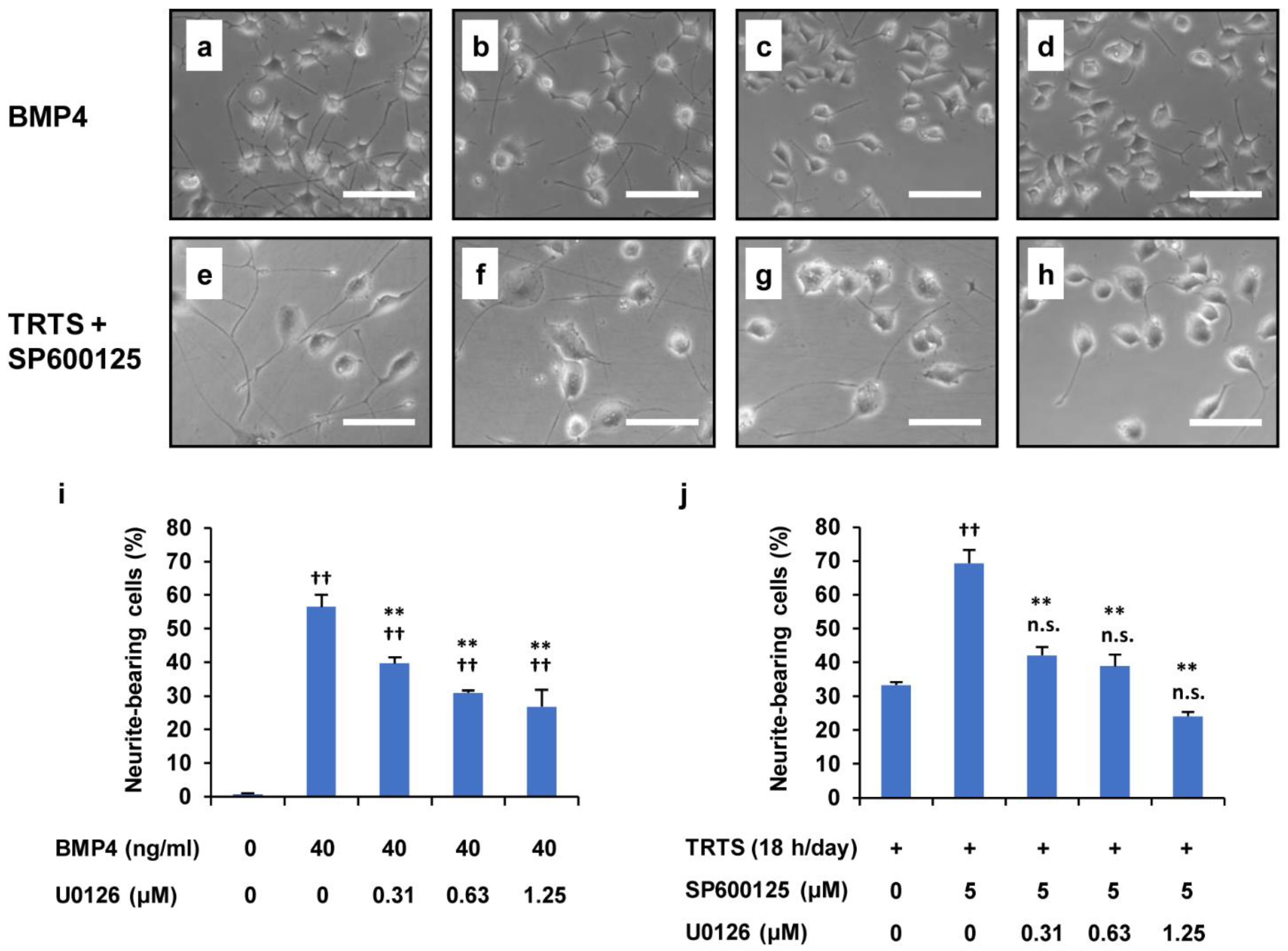
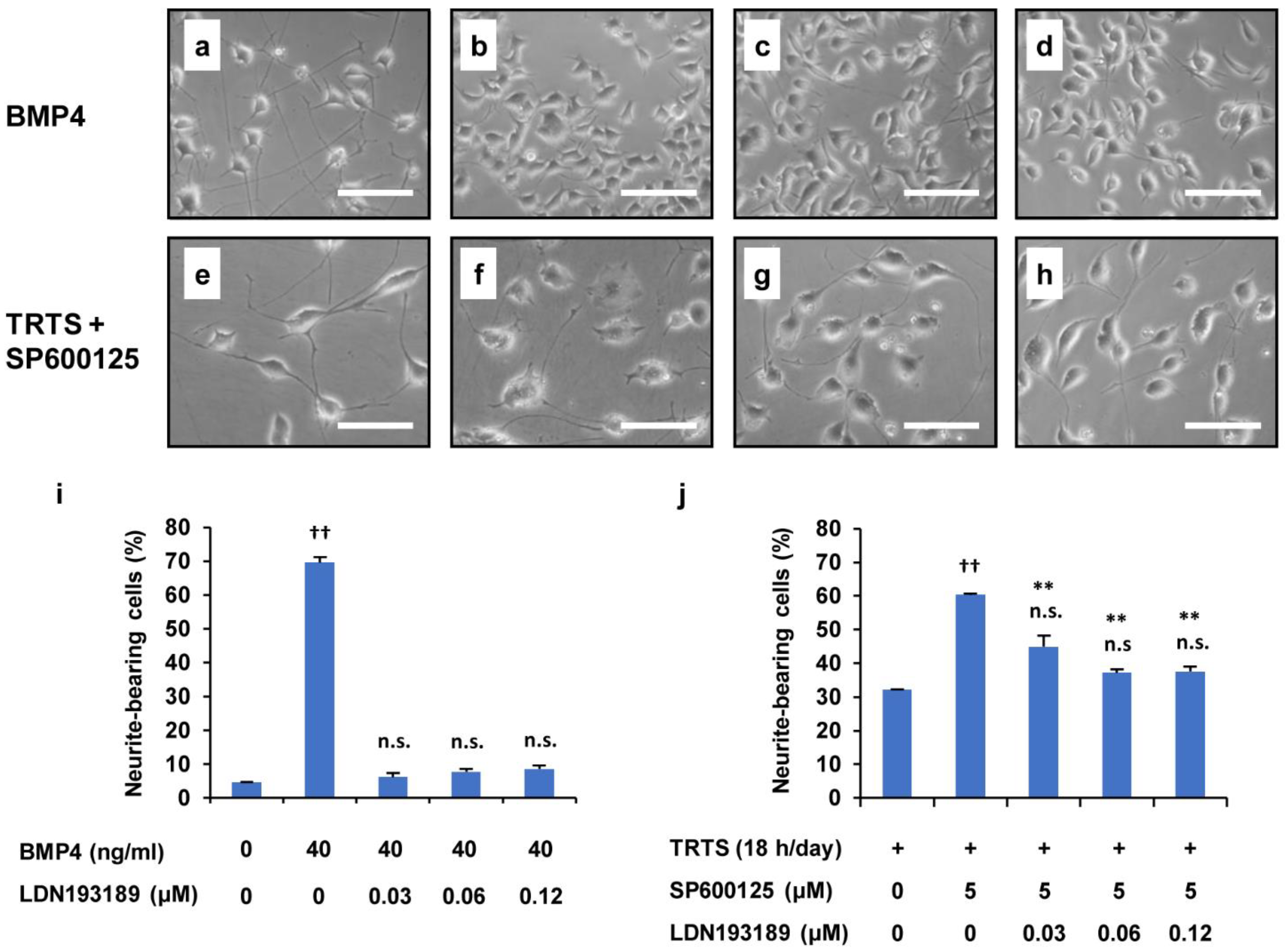
2.6. Suppression of the SP600125-Mediated Promotion of TRTS-Induced Neuritogenesis by U0126 and LDN193189
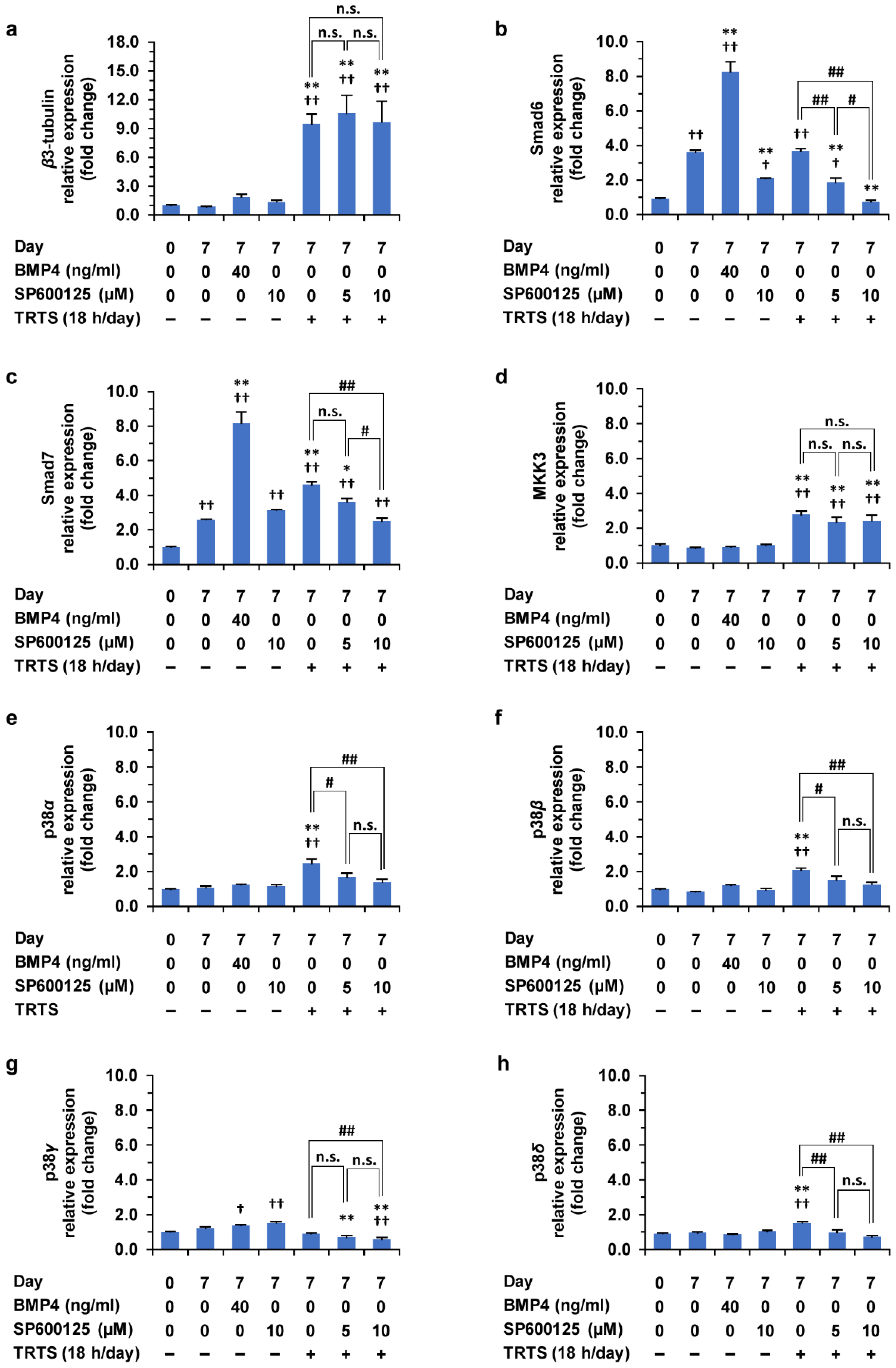
2.7. Effects of TRTS in the Presence or Absence of SP600125 on Gene Expressions
3. Discussion
4. Materials and Methods
4.1. Cells and Reagents
4.2. Cell Culture and the Induction of Neurite Outgrowth
4.3. Thermal Evaluation of the Medium
4.4. Live Cell Imaging with Fluorescein Diacetate Staining
4.5. Neurite Length Measurement of Live Cells
4.6. Quantitative Real-Time Polymerase Chain Reaction
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Teng, K.K.; Georgieff, I.S.; Nunez, J.; Shelanski, M.L.; Greene, L.A. Characterization of a PC12 cell sub-clone (PC12-C41) with enhanced neurite outgrowth with capacity: Implications for a modulatory role of high molecular weight tau in neuritogenesis. J. Cell Sci. 1993, 106, 611–626. [Google Scholar] [CrossRef] [PubMed]
- Gokoffski, K.K.; Peng, M.; Alas, B.; Lam, P. Neuro-protection and neuro-regeneration of the optic nerve: Recent advances and future directions. Curr. Opin. Neurol. 2020, 33, 93–105. [Google Scholar] [CrossRef] [PubMed]
- Higgins, S.; Lee, J.S.; Ha, L.; Lim, J.Y. Inducing neurite outgrowth by mechanical cell stretch. Biores. Open Access 2013, 2, 212–216. [Google Scholar] [CrossRef] [PubMed]
- Kudo, T.; Kanetaka, H.; Mochizuki, K.; Tominami, K. Induction of neurite outgrowth in PC12 cells treated with temperature-controlled repeated thermal stimulation. PLoS ONE 2015, 10, e0124024. [Google Scholar] [CrossRef] [PubMed]
- Wiatrak, B.; Kubis-Kubiak, A.; Piwowar, A.; Barg, E. PC12 cell line: Cell types, coating of culture vessels, differentiation and other culture conditions. Cells 2020, 9, 958. [Google Scholar] [CrossRef]
- Radio, N.M.; Mundy, W.R. Developmental neurotoxicity testing in vitro: Models for assessing chemical effects on neurite outgrowth. Neurotoxicology 2008, 29, 361–376. [Google Scholar] [CrossRef] [PubMed]
- Greene, L.A.; Tischler, A.S. Establishment of a noradrenergic clonal line of rat adrenal pheochromocytoma cells which respond to nerve growth factor. Proc. Natl. Acad. Sci. USA 1976, 73, 2424–2428. [Google Scholar] [CrossRef] [PubMed]
- Gill, J.S.; Schenone, A.E.; Podratz, J.L.; Windebank, A.J. Autocrine regulation of neurite outgrowth from PC12 cells by nerve growth factor. Mol. Brain Res. 1998, 57, 123–131. [Google Scholar] [CrossRef]
- Mielke, K.; Herdegen, T. JNK and p38 stresskinases--degenerative effectors of signal-transduction-cascades in the nervous system. Prog. Neurobiol. 2000, 61, 45–60. [Google Scholar] [CrossRef]
- Ambrosino, C.; Mace, G.; Galban, S.; Fritsch, C.; Vintersten, K.; Black, E.; Gorospe, M.; Nebreda, A.R. Negative feedback regulation of MKK6 mRNA stability by p38alpha mitogen-activated protein kinase. Mol. Cell. Biol. 2003, 23, 370–381. [Google Scholar] [CrossRef]
- Ana, C.; Angel, R.N. Mechanisms and functions of p38 MAPK signalling. Biochem. J. 2010, 429, 403–417. [Google Scholar]
- Raingeaud, J.; Whitmarsh, A.J.; Barrett, T.; Dérijard, B.; Davis, R.J. MKK3- and MKK6-regulated gene expression is mediated by the p38 mitogen-activated protein kinase signal transduction pathway. Mol. Cell. Biol. 1996, 16, 1247–1255. [Google Scholar] [CrossRef] [PubMed]
- Burton, J.C.; Grimsey, N.J. Ubiquitination as a Key Regulator of Endosomal Signaling by GPCRs. Front. Cell Dev. Biol. 2019, 7, 43. [Google Scholar] [CrossRef] [PubMed]
- Johnson, G.L.; Lapadat, R. Mitogen-activated protein kinase pathways mediated by ERK, JNK, and p38 protein kinases. Science 2002, 298, 1911–1912. [Google Scholar] [CrossRef]
- Cargnello, M.; Roux, P.P. Activation and function of the MAPKs and their substrates, the MAPK-activated protein kinases. Microbiol. Mol. Biol. Rev. 2011, 75, 50–83. [Google Scholar] [CrossRef]
- Vaudry, D.; Stork, P.J.S.; Lazarovici, P.; Eiden, L.E. Signaling pathways for PC12 cell differentiation: Making the right connections. Science 2002, 296, 1648–1649. [Google Scholar] [CrossRef]
- Obara, Y.; Yamauchi, A.; Takehara, S.; Nemoto, W.; Takahashi, M.; Stork, P.J.S.; Nakahata, N. ERK5 activity is required for nerve growth factor-induced neurite outgrowth and stabilization of tyrosine hydroxylase in PC12 cells. J. Biol. Chem. 2009, 284, 564–573. [Google Scholar] [CrossRef]
- Kashino, Y.; Obara, Y.; Okamoto, Y.; Saneyoshi, T.; Hayashi, Y.; Ishii, K. ERK5 phosphorylates Kv4.2 and inhibits inactivation of the A-type current in PC12 cells. Int. J. Mol. Sci. 2018, 19, 2008. [Google Scholar] [CrossRef]
- Iwasaki, S.; Iguchi, M.; Watanabe, K.; Hoshino, R.; Tsujimoto, M.; Kohno, M. Specific Activation of the p38 mitogen-activated protein kinase signaling pathway and induction of neurite outgrowth in PC12 cells by bone morphogenetic protein-2. J. Biol. Chem. 1999, 274, 26503–26510. [Google Scholar] [CrossRef]
- Liu, J.; Lin, A. Role of JNK activation in apoptosis: A double-edged sword. Cell Res. 2005, 15, 36–42. [Google Scholar] [CrossRef]
- Xiao, J.; Zhou, Q.; Liu, Y. Variant PC12 cell line that spontaneously differentiates and extends neuritic processes. J. Neurosci. Res. 2002, 69, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Pradhan, A.; Liu, Y. Functional role of JNK in neuritogenesis of PC12-N1 cells. Neurosci. Lett. 2006, 392, 231–234. [Google Scholar] [CrossRef] [PubMed]
- Ebendal, T.; Bengtsson, H.; Söderström, S. Bone morphogenetic proteins and their receptors: Potential functions in the brain. J. Neurosci. Res. 1998, 51, 139–146. [Google Scholar] [CrossRef]
- Iwasaki, S.; Hattori, A.; Sato, M.; Tsujimoto, M.; Kohno, M. Characterization of the bone morphogenetic protein-2 as a neurotrophic factor. Induction of neuronal differentiation of PC12 cells in the absence of mitogen-activated protein kinase activation. J. Biol. Chem. 1996, 271, 17360–17365. [Google Scholar] [CrossRef] [PubMed]
- Althini, S.; Usoskin, D.; Kylberg, A.; Kaplan, P.L.; Ebendal, T. Blocked MAP kinase activity selectively enhances neurotrophic growth responses. Mol. Cell. Neurosci. 2004, 25, 345–354. [Google Scholar] [CrossRef] [PubMed]
- Lönn, P.; Zaia, K.; Israelsson, C.; Althini, S.; Usoskin, D.; Kylberg, A.; Ebendal, T. BMP enhances transcriptional responses to NGF during PC12 cell differentiation. Neurochem. Res. 2005, 30, 753–765. [Google Scholar] [CrossRef]
- Kimura, N.; Matsuo, R.; Shibuya, H.; Nakashima, K.; Taga, T. BMP2-induced apoptosis is mediated by activation of the TAK1-p38 kinase pathway that is negatively regulated by Smad6. J. Biol. Chem. 2000, 275, 17647–17652. [Google Scholar] [CrossRef]
- Kudo, T.; Kanetaka, H.; Mizuno, K.; Ryu, Y.; Miyamoto, Y.; Nunome, S.; Zhang, Y.; Kano, M.; Shimizu, Y.; Hayashi, H. Dor somorphin stimulates neurite outgrowth in PC12 cells via activation of a protein kinase A-dependent MEK-ERK1⁄2 signaling pathway. Genes Cells 2011, 16, 1121–1132. [Google Scholar] [CrossRef]
- Yanagisawa, M.; Nakashima, K.; Takeda, K.; Ochiai, W.; Takizawa, T.; Ueno, M.; Takizawa, M.; Shibuya, H.; Taga, T. Inhibition of BMP2-induced, TAK1 kinase-mediated neurite outgrowth by Smad6 and Smad7. Genes Cells 2001, 6, 1091–1099. [Google Scholar] [CrossRef]
- Hirata, Y.; Takahashi, M.; Morishita, T.; Noguchi, T.; Matsuzawa, A. Post-Translational Modifications of the TAK1-TAB Complex. Int. J. Mol. Sci. 2017, 18, 205. [Google Scholar] [CrossRef]
- Kudo, T.; Kanetaka, H.; Watanabe, A.; Okumoto, A.; Asano, M.; Zhang, Y.; Zhao, F.; Kano, M.; Shimizu, Y.; Tamura, S.; et al. Investigating bone morphogenetic protein (BMP) signaling in a newly established human cell line expressing BMP receptor type II. Tohoku. J. Exp. Med. 2010, 222, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Long, Q.; Wu, B.; Yang, Y.; Wang, S.; Shen, Y.; Bao, Q.; Xu, F. Nerve guidance conduit promoted peripheral nerve regeneration in rats. Artif. Organs 2021, 45, 616–624. [Google Scholar] [CrossRef] [PubMed]
- Kano, Y.; Nohno, T.; Takahashi, R.; Hasegawa, T.; Hiragami, F.; Kawamura, K.; Motoda, H.; Sugiyama, T. CAMP and calcium ionophore induce outgrowth of neuronal processes in PC12 mutant cells in which nerve growth factor-induced outgrowth of neuronal processes is impaired. Neurosci. Lett. 2001, 303, 21–24. [Google Scholar] [CrossRef] [PubMed]
- Murai, H.; Hiragami, F.; Kawamura, K.; Motoda, H.; Koike, Y.; Inoue, S.; Kumagishi, K.; Ohtsuka, A.; Kano, Y. Differential response of heat-shock-induced p38 MAPK and JNK activity in PC12 mutant and PC12 parental cells for differentiation and apoptosis. Acta Med. Okayama 2010, 64, 55–62. [Google Scholar] [CrossRef]
- Kudo, T.; Tominami, K.; Izumi, S.; Hayashi, Y.; Noguchi, T.; Matsuzawa, A.; Hong, G.; Nakai, J. Characterization of PC12 cell subclones with different sensitivities to programmed thermal stimulation. Int. J. Mol. Sci. 2020, 21, 8356. [Google Scholar] [CrossRef]
- Bennett, B.L.; Sasaki, D.T.; Murray, B.W.; O’Leary, E.C.; Sakata, S.T.; Xu, W.; Leisten, J.C.; Motiwala, A.; Pierce, S.; Satoh, Y.; et al. SP600125, an anthrapyrazolone inhibitor of jun N-terminal kinase. Proc. Natl. Acad. Sci. USA 2001, 98, 13681–13686. [Google Scholar] [CrossRef]
- Vaishnav, D.; Jambal, P.; Reusch, J.E.B.; Pugazhenthi, S. SP600125, an inhibitor of c-Jun N-terminal kinase, activates CREB by a p38 MAPK-mediated pathway. Biochem. Biophys. Res. Commun. 2003, 307, 855–860. [Google Scholar] [CrossRef]
- Bonni, A.; Ginty, D.D.; Dudek, H.; Greenberg, M.E. Serine 133-phosphorylated CREB induces transcription via a cooperative mechanism that may confer specificity to neurotrophin signals. Mol. Cell. Neurosci. 1995, 6, 168–183. [Google Scholar] [CrossRef]
- Angell, R.M.; Atkinson, F.L.; Brown, M.J.; Chuang, T.T.; Christopher, J.A.; Cichy-Knight, M.; Dunn, A.K.; Hightower, K.E.; Malkakorpi, S.; Musgrave, J.R.; et al. N-(3-Cyano-4,5,6,7-Tetrahydro-1-Benzothien-2-Yl)amides as potent, selective, inhibitors of JNK2 and JNK3. Bioorganic Med. Chem. Lett. 2007, 17, 1296–1301. [Google Scholar] [CrossRef]
- Nunome, S.; Kanetaka, H.; Kudo, T.; Endoh, K.; Hosoda, H.; Igarashi, K. In vitro evaluation of biocompatibility of Ti-Mo-Sn-Zr superelastic alloy. J. Biomater. Appl. 2015, 30, 119–130. [Google Scholar] [CrossRef]
- Hayashi, Y.; Otsuka, K.; Ebina, M.; Igarashi, K.; Takehara, A.; Matsumoto, M.; Kanai, A.; Igarashi, K.; Soga, T.; Matsui, Y. Distinct requirements for energy metabolism in mouse primordial germ cells and their reprogramming to embryonic germ cells. Proc. Natl. Acad. Sci. USA 2017, 114, 8289–8294. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genbank Accession Number | Gene | Forward Primer (5′-3′) | Reverse Primer (5′-3′) |
|---|---|---|---|
| NM_139254.2 | β3-tubulin | TCCACCTTCATCGGCAACA | CGGTGAACTCCATCTCATCCA |
| NM_001109002.3 | Smad6 | CACTGCTCCGGGTGAATTCTC | AGTATGCCACGCTGCACCA |
| NM_030858.2 | Smad7 | AGCAAGAGTCAGCACTGCCAAG | TGACAACTGAAATGCTGATCCAAAG |
| NM_001100674.1 | MKK3 | GTCTGGAGCCTTGGCATCAC | CCTGCTTCAGCTGCTGGAAC |
| NM_031020.3 | p38α | ATGCAGTCCAGCTCCACGTC | TCCTAACACAGCATGGCCACA |
| NM_001109532.2 | p38β | GGCAAAGATATCCTCGGAGCA | TGGTCACTGTCTAGCACCAGCA |
| NM_021746.2 | p38γ | TGGCTGTGAACGAGGACTGTG | GATGACCTCTGGTGCCCGATA |
| NM_019231.3 | p38δ | GAAGGTCCAGTATTTGGTGTACCAG | CTTCATTCACGGCCAGGTTG |
| NM_031144.3 | β-actin | GGAGATTACTGCCCTGGCTCCTA | GACTCATCGTACTCCTGCTTGCTG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, Y.-R.; Kudo, T.-a.; Tominami, K.; Izumi, S.; Tanaka, T.; Hayashi, Y.; Noguchi, T.; Matsuzawa, A.; Nakai, J.; Hong, G.; et al. SP600125 Enhances Temperature-Controlled Repeated Thermal Stimulation-Induced Neurite Outgrowth in PC12-P1F1 Cells. Int. J. Mol. Sci. 2022, 23, 15602. https://doi.org/10.3390/ijms232415602
Luo Y-R, Kudo T-a, Tominami K, Izumi S, Tanaka T, Hayashi Y, Noguchi T, Matsuzawa A, Nakai J, Hong G, et al. SP600125 Enhances Temperature-Controlled Repeated Thermal Stimulation-Induced Neurite Outgrowth in PC12-P1F1 Cells. International Journal of Molecular Sciences. 2022; 23(24):15602. https://doi.org/10.3390/ijms232415602
Chicago/Turabian StyleLuo, You-Ran, Tada-aki Kudo, Kanako Tominami, Satoshi Izumi, Takakuni Tanaka, Yohei Hayashi, Takuya Noguchi, Atsushi Matsuzawa, Junichi Nakai, Guang Hong, and et al. 2022. "SP600125 Enhances Temperature-Controlled Repeated Thermal Stimulation-Induced Neurite Outgrowth in PC12-P1F1 Cells" International Journal of Molecular Sciences 23, no. 24: 15602. https://doi.org/10.3390/ijms232415602
APA StyleLuo, Y.-R., Kudo, T.-a., Tominami, K., Izumi, S., Tanaka, T., Hayashi, Y., Noguchi, T., Matsuzawa, A., Nakai, J., Hong, G., & Wang, H. (2022). SP600125 Enhances Temperature-Controlled Repeated Thermal Stimulation-Induced Neurite Outgrowth in PC12-P1F1 Cells. International Journal of Molecular Sciences, 23(24), 15602. https://doi.org/10.3390/ijms232415602