
Influence of the Photodegradation of Azathioprine on DNA and Cells
 , , ,
, , , 
Abstract
1. Introduction
2. Results and Discussion
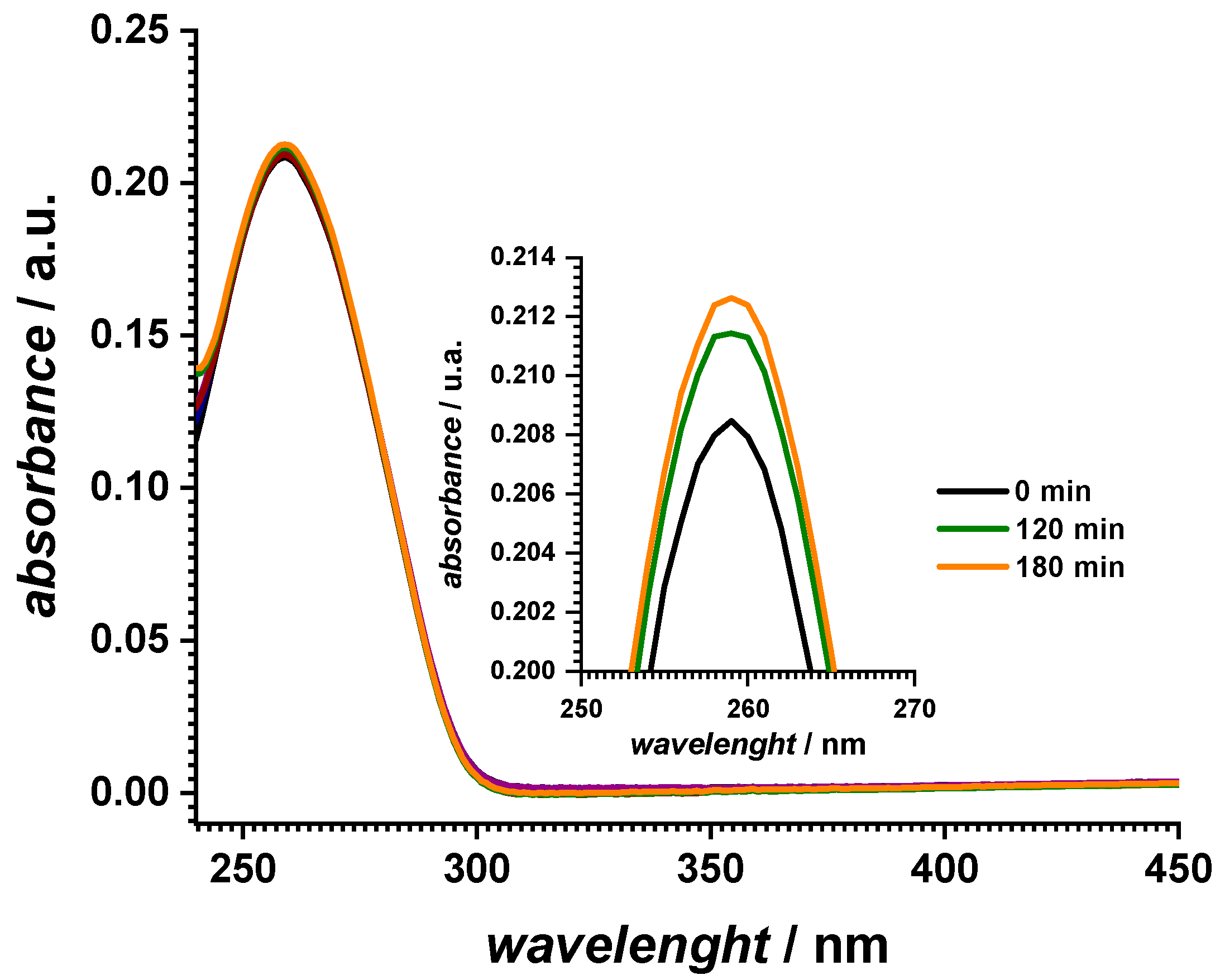
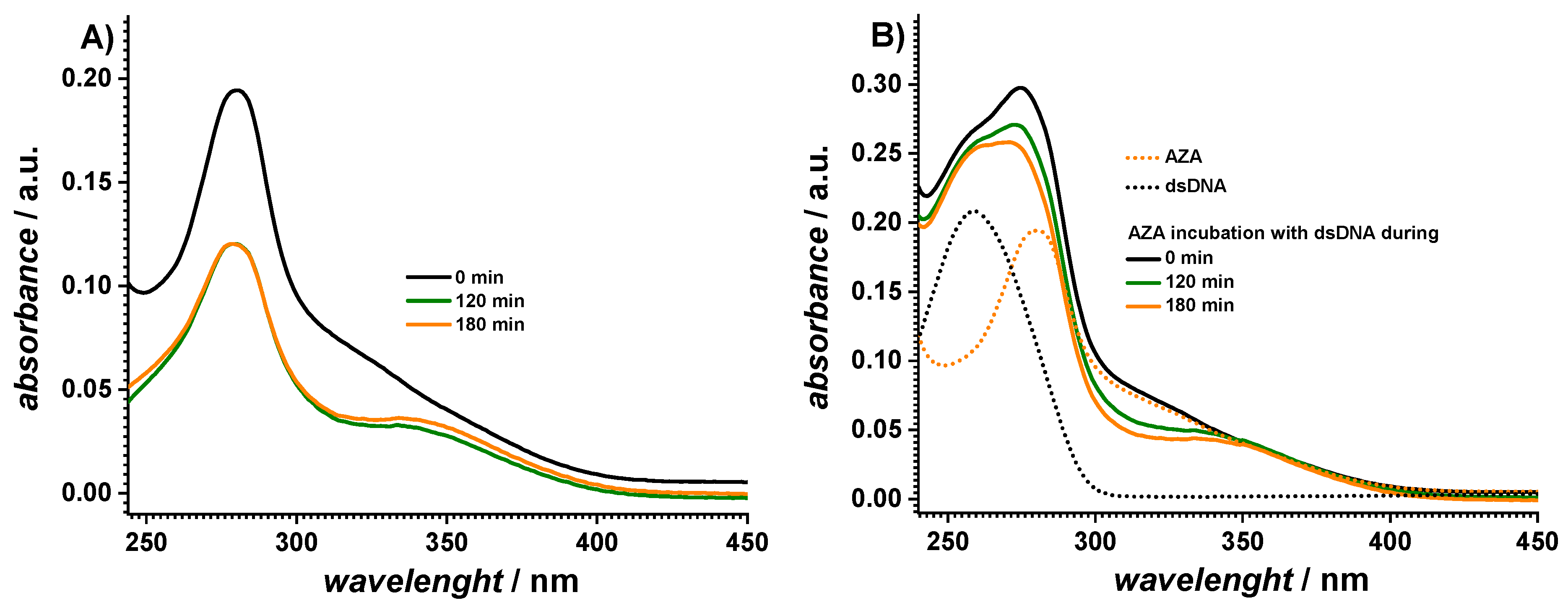
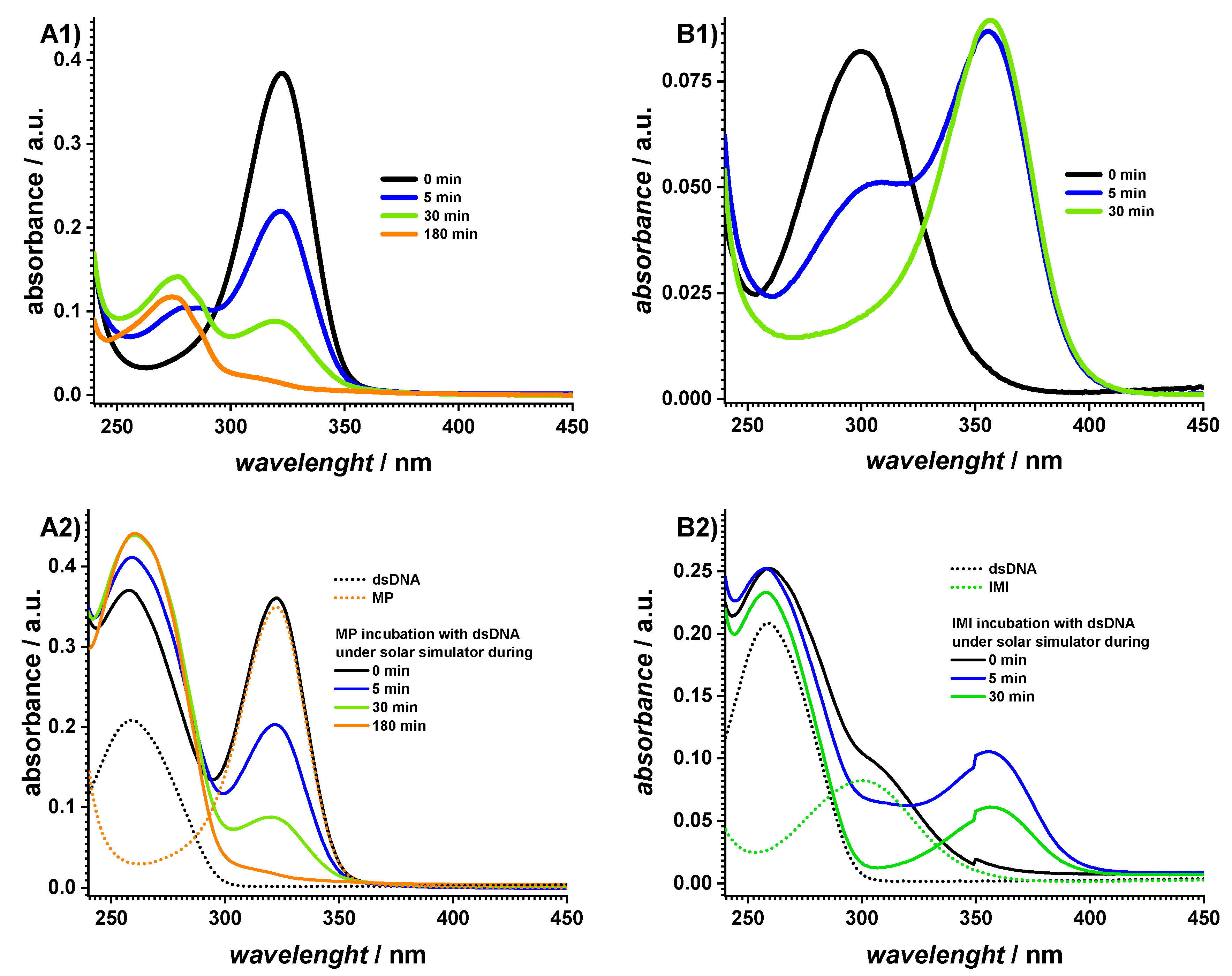
2.1. Spectrophotometric Characterization
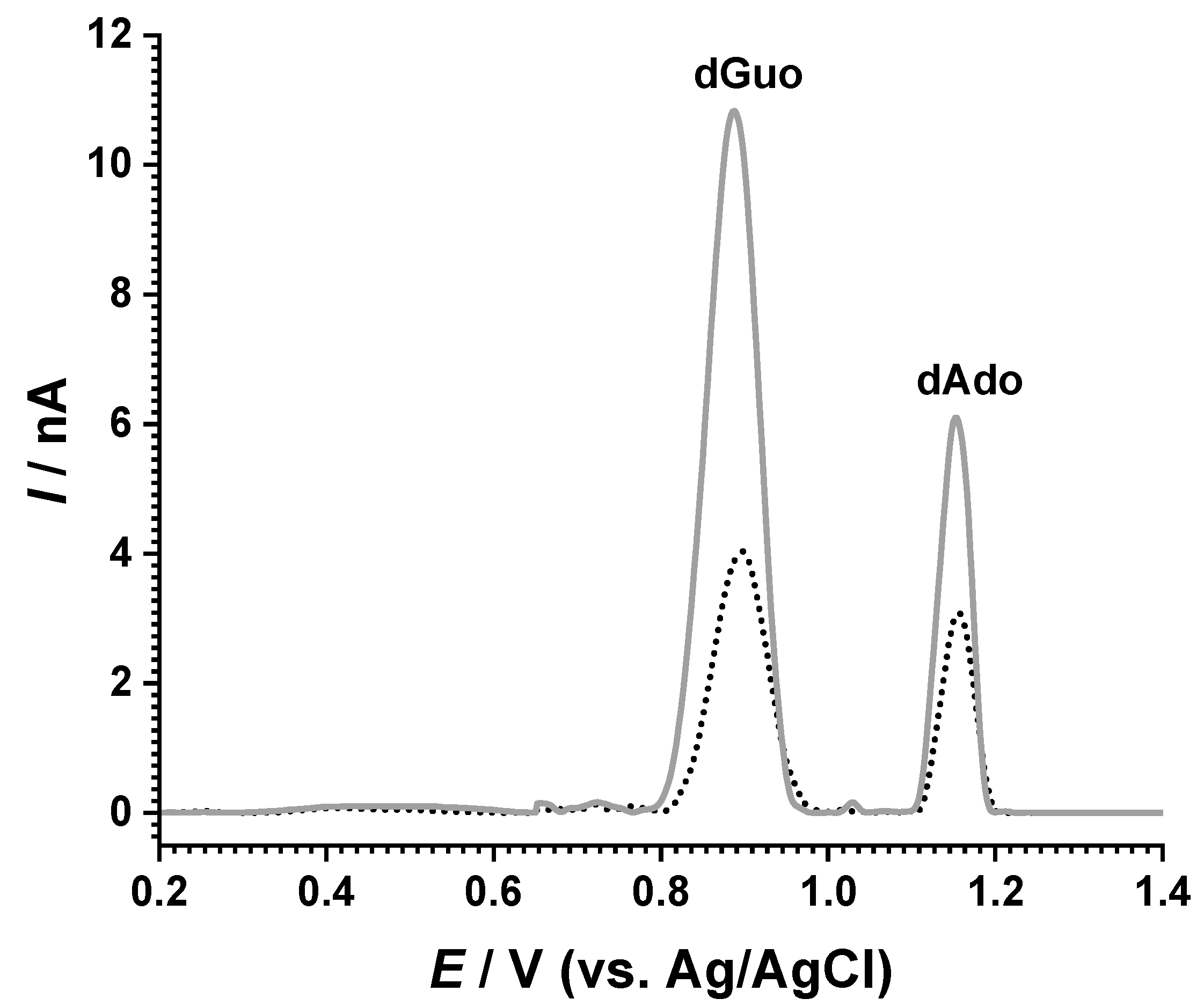
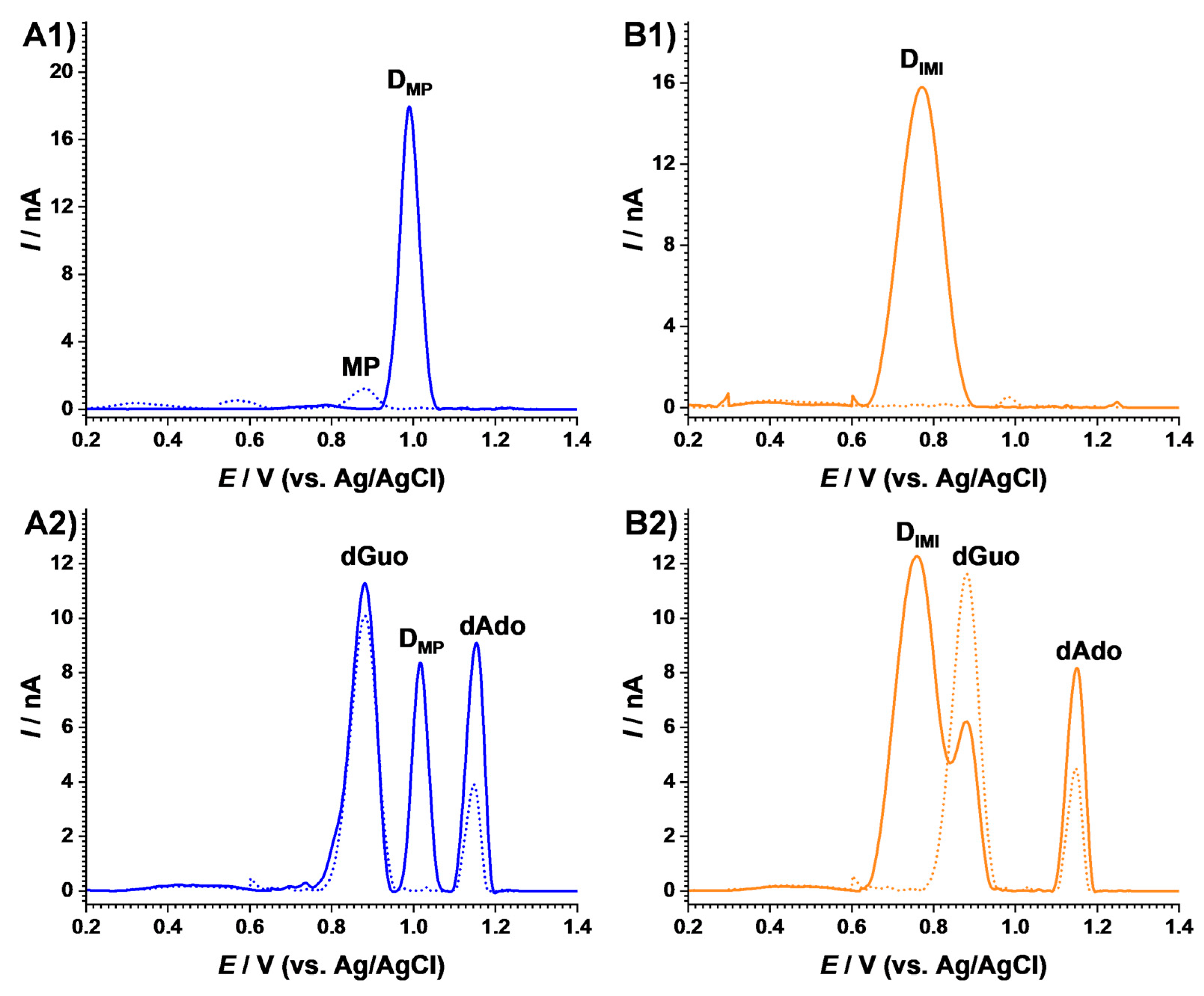
2.2. Electrochemical Characterization
2.3. Cell Viability and Proliferation
3. Materials and Methods
3.1. Materials
3.2. Instrumentation
3.2.1. Solar Simulator Irradiation
3.2.2. UV-Vis Spectrophotometry
3.2.3. Voltammetric Parameters and Electrochemical Cells
3.2.4. Fluorescence Microscopy
3.3. Procedures
3.3.1. Photodegradation
3.3.2. Viability Assay
3.3.3. Fluorescence Microscopy
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hemmens, V.J.; Moore, D.E. Photochemical Sensitization by Azathioprine and Its Metabolites—Ii. Azathioprine and Nitroimidazole Metabolites. Photochem. Photobiol. 1986, 43, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Gupta, R.L. A Critical Study on Degradation of Azathioprine in Aqueous Solutions. Int. J. Pharm. 1988, 42, 263–266. [Google Scholar] [CrossRef]
- Cuffari, C. A Physician’s Guide to Azathioprine Metabolite Testing. Gastroenterol. Hepatol. 2006, 2, 58. [Google Scholar]
- Maltzman, J.S.; Koretzky, G.A. Azathioprine: Old Drug, New Actions. J. Clin. Investig. 2003, 111, 1122–1124. [Google Scholar] [CrossRef] [PubMed]
- Weersma, R.K.; Peters, F.T.M.; Oostenbrug, L.E.; Van Den Berg, A.P.; Van Haastert, M.; Ploeg, R.J.; Posthumus, M.D.; Homan Van Der Heide, J.J.; Jansen, P.L.M.; Van Dullemen, H.M. Increased Incidence of Azathioprine-Induced Pancreatitis in Crohn’s Disease Compared with Other Diseases. Aliment. Pharmacol. Ther. 2004, 20, 843–850. [Google Scholar] [CrossRef] [PubMed]
- Brooke, B.N.; Hoffmann, D.C.; Swarbrick, E.T. Azathioprine for Crohn’s Disease. Lancet 1969, 2, 612–614. [Google Scholar] [CrossRef]
- Mishra, K.; Pramanik, S.; Sandal, R.; Jandial, A.; Sahu, K.K.; Singh, K.; Khera, S.; Meshram, A.; Khurana, H.; Somasundaram, V.; et al. Safety and Efficacy of Azathioprine in Immune Thrombocytopenia. Am. J. Blood Res. 2021, 11, 217–226. [Google Scholar]
- Broekman, M.M.T.J. Optimization of Treatment with Conventional Thiopurines in Inflammatory Bowel Disease. Ph.D. Thesis, Radboud University Medical Centre, Nijmegen, The Netherlands, 2021. [Google Scholar]
- Claeys, E.; Vermeire, K. Immunosuppressive Drugs in Organ Transplantation to Prevent Allograft Rejection: Mode of Action and Side Effects. J. Immunol. Sci. 2019, 3, 14–21. [Google Scholar] [CrossRef]
- Herman-Edelstein, M.; Rozen-Zvi, B.; Zingerman, B.; Lichtenberg, S.; Malachi, T.; Gafter, U.; Ori, Y. Effect of Immunosuppressive Drugs on DNA Repair in Human Peripheral Blood Mononuclear Cells. Biomed. Pharmacother. 2012, 66, 111–115. [Google Scholar] [CrossRef]
- Hoffmann, M.; Rychlewski, J.; Chrzanowska, M.; Hermann, T. Mechanism of Activation of an Immunosuppressive Drug: Azathioprine. Quantum Chemical Study on the Reaction of Azathioprine with Cysteine. J. Am. Chem. Soc. 2001, 123, 6404–6409. [Google Scholar] [CrossRef]
- Rehman, S.U.; Yaseen, Z.; Husain, M.A.; Sarwar, T.; Ishqi, H.M.; Tabish, M. Interaction of 6 Mercaptopurine with Calf Thymus DNA—Deciphering the Binding Mode and Photoinduced DNA Damage. PLoS ONE 2014, 9, e93913. [Google Scholar] [CrossRef] [PubMed]
- Lavezzo, M.M.; Maria, F.; Souto, S.; Hirata, C.E. Early Azathioprine in Acute Vogt-Koyanagi-Harada Disease: A Prospective 24-Month Follow-up Study with Multimodal Imaging and Electroretinogram. Available online: https://www.researchsquare.com/article/rs-1251714/v1 (accessed on 4 October 2022).
- Cangemi, M.; Montico, B.; Faè, D.A.; Steffan, A.; Dolcetti, R. Dissecting the Multiplicity of Immune Effects of Immunosuppressive Drugs to Better Predict the Risk of de Novo Malignancies in Solid Organ Transplant Patients. Front. Oncol. 2019, 9, 160. [Google Scholar] [CrossRef] [PubMed]
- Attard, N.R.; Karran, P. UVA Photosensitization of Thiopurines and Skin Cancer in Organ Transplant Recipients. Photochem. Photobiol. Sci. 2012, 11, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Gueranger, Q.; Li, F.; Peacock, M.; Larnicol-Fery, A.; Brem, R.; Macpherson, P.; Egly, J.M.; Karran, P. Protein Oxidation and DNA Repair Inhibition by 6-Thioguanine and UVA Radiation. J. Investig. Dermatol. 2014, 134, 1408–1417. [Google Scholar] [CrossRef]
- Perrett, C.M.; Walker, S.L.; O’Donovan, P.; Warwick, J.; Harwood, C.A.; Karran, P.; McGregor, J.M. Azathioprine Treatment Photosensitizes Human Skin to Ultraviolet A Radiation. Br. J. Dermatol. 2008, 159, 198–204. [Google Scholar] [CrossRef] [PubMed]
- Ulrich, C.; Stockfleth, E. Azathioprine, UV Light, and Skin Cancer in Organ Transplant Patients Do We Have an Answer? Med. Monatsschr. Pharm. 2000, 23, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Jalali, F.; Rasaee, G. Electrochemical, Spectroscopic, and Theoretical Studies on the Interaction between Azathioprine and DNA. Int. J. Biol. Macromol. 2015, 81, 427–434. [Google Scholar] [CrossRef]
- Hlavata, L.; Benikova, K.; Vyskocil, V.; Labuda, J. Evaluation of Damage to DNA Induced by UV-C Radiation and Chemical Agents Using Electrochemical Biosensor Based on Low Molecular Weight DNA and Screen-Printed Carbon Electrode. Electrochim. Acta 2012, 71, 134–139. [Google Scholar] [CrossRef]
- Oliveira-Brett, A.M.; Paquim, A.M.C.; Diculescu, V.C.; Piedade, J.A.P. Electrochemistry of Nanoscale DNA Surface Films on Carbon. Med. Eng. Phys. 2006, 28, 963–970. [Google Scholar] [CrossRef]
- Karran, P.; Attard, N. Thiopurines in Current Medical Practice: Molecular Mechanisms and Contributions to Therapy-Related Cancer. Nat. Rev. Cancer 2008, 8, 24–36. [Google Scholar] [CrossRef]
- Smaranda, I.; Nila, A.; Manta, C.M.; Samohvalov, D.; Gherca, D.; Baibarac, M. The Influence of UV Light on the Azathioprine Photodegradation: New Evidences by Photoluminescence. Results Phys. 2019, 14, 102443. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, J.; Xiao, Y.; Chang, V.W.C.; Lim, T.T. Kinetic and Mechanistic Investigation of Azathioprine Degradation in Water by UV, UV/H2O2 and UV/Persulfate. Chem. Eng. J. 2016, 302, 526–534. [Google Scholar] [CrossRef]
- Bunea, M.C.; Diculescu, V.C.; Enculescu, M.; Iovu, H.; Enache, T.A. Redox Mechanism of Azathioprine and Its Interaction with DNA. Int. J. Mol. Sci. 2021, 22, 6805. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | AZA | MP | IMI | dsDNA | AZA + dsDNA | MP + dsDNA | IMI + dsDNA | |
|---|---|---|---|---|---|---|---|---|
| A260 | 0 min | 0.19 | 0.03 | 0.03 | 0.21 | 0.26 | 0.37 | 0.25 |
| 180 min | 0.12 | 0.09 | 0.02 | 0.21 | 0.26 | 0.44 | 0.23 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bunea, M.-C.; Diculescu, V.-C.; Enculescu, M.; Oprea, D.; Enache, T.A. Influence of the Photodegradation of Azathioprine on DNA and Cells. Int. J. Mol. Sci. 2022, 23, 14438. https://doi.org/10.3390/ijms232214438
Bunea M-C, Diculescu V-C, Enculescu M, Oprea D, Enache TA. Influence of the Photodegradation of Azathioprine on DNA and Cells. International Journal of Molecular Sciences. 2022; 23(22):14438. https://doi.org/10.3390/ijms232214438
Chicago/Turabian StyleBunea, Mihaela-Cristina, Victor-Constantin Diculescu, Monica Enculescu, Daniela Oprea, and Teodor Adrian Enache. 2022. "Influence of the Photodegradation of Azathioprine on DNA and Cells" International Journal of Molecular Sciences 23, no. 22: 14438. https://doi.org/10.3390/ijms232214438
APA StyleBunea, M.-C., Diculescu, V.-C., Enculescu, M., Oprea, D., & Enache, T. A. (2022). Influence of the Photodegradation of Azathioprine on DNA and Cells. International Journal of Molecular Sciences, 23(22), 14438. https://doi.org/10.3390/ijms232214438