
Effector-Dependent and -Independent Molecular Mechanisms of Soybean–Microbe Interaction

 ,
,  and
and 
Abstract
1. Introduction
2. Interaction between Nematode Effectors and Soybean R Genes

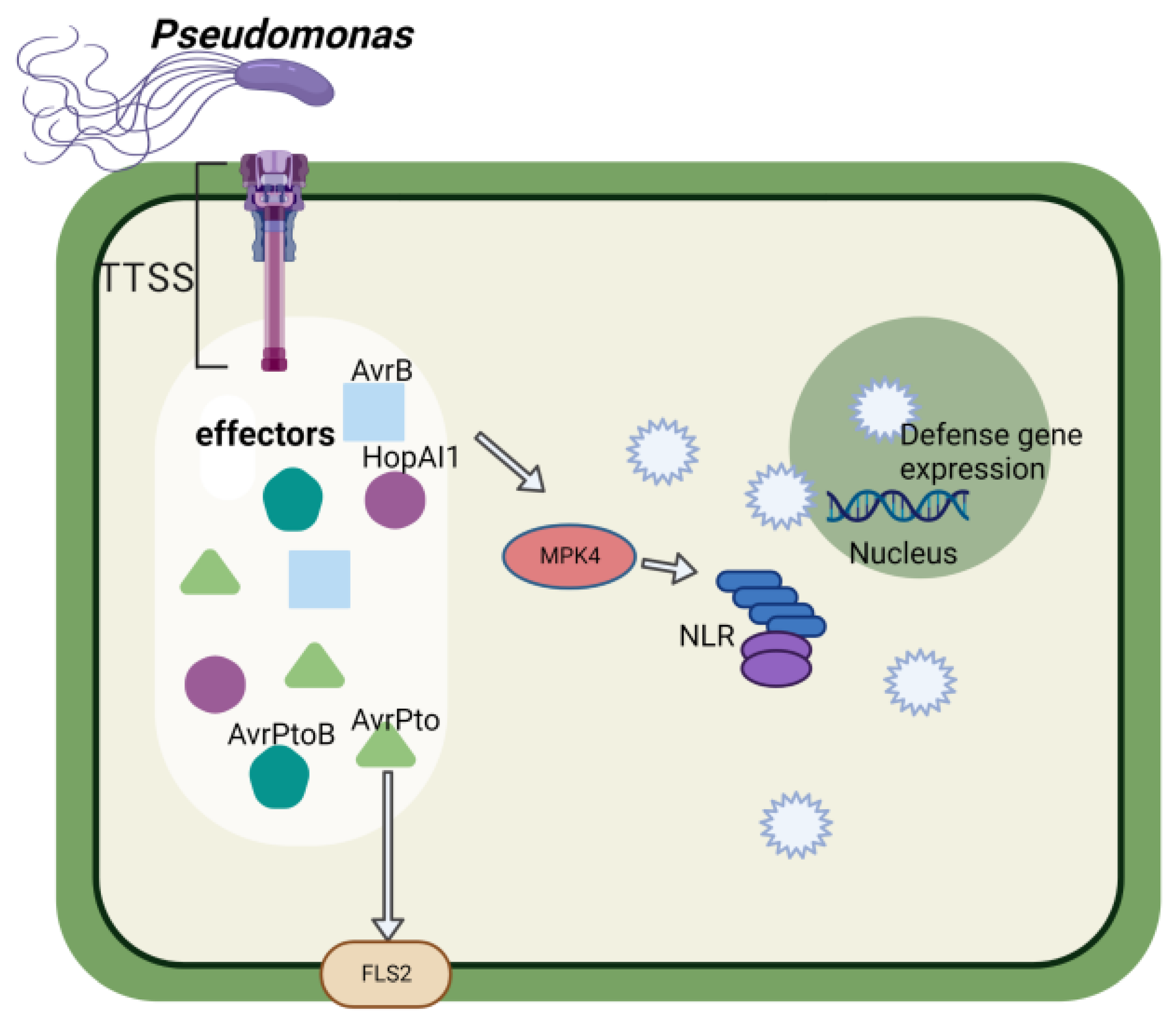
3. Function of Pseudomonas Effectors in Soybean–Pseudomonas Interaction
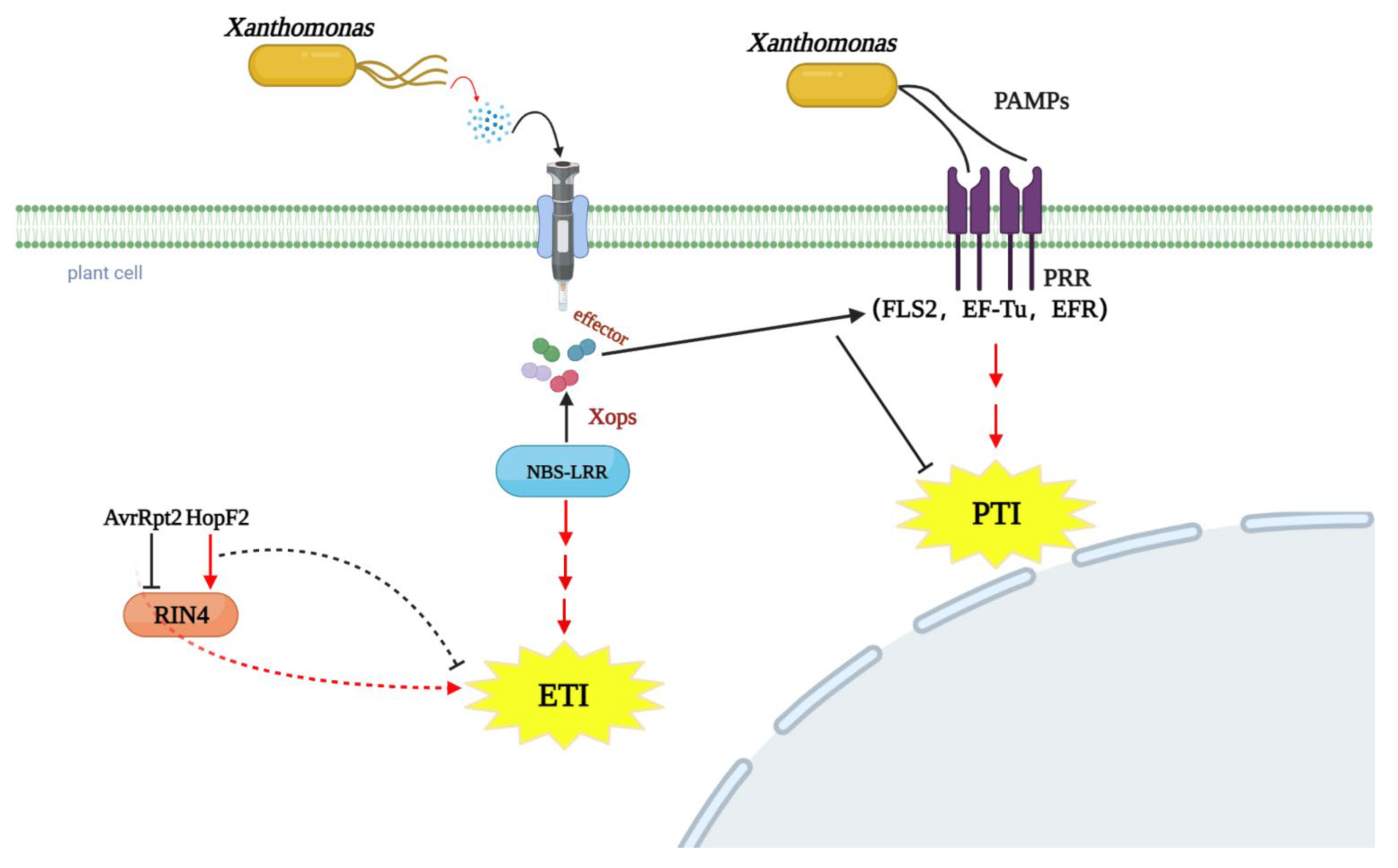
4. The Infection Molecular Signaling of Xanthomonas on Legume
5. Conservation of Pseudomonas and Xanthomonas Effector Targets
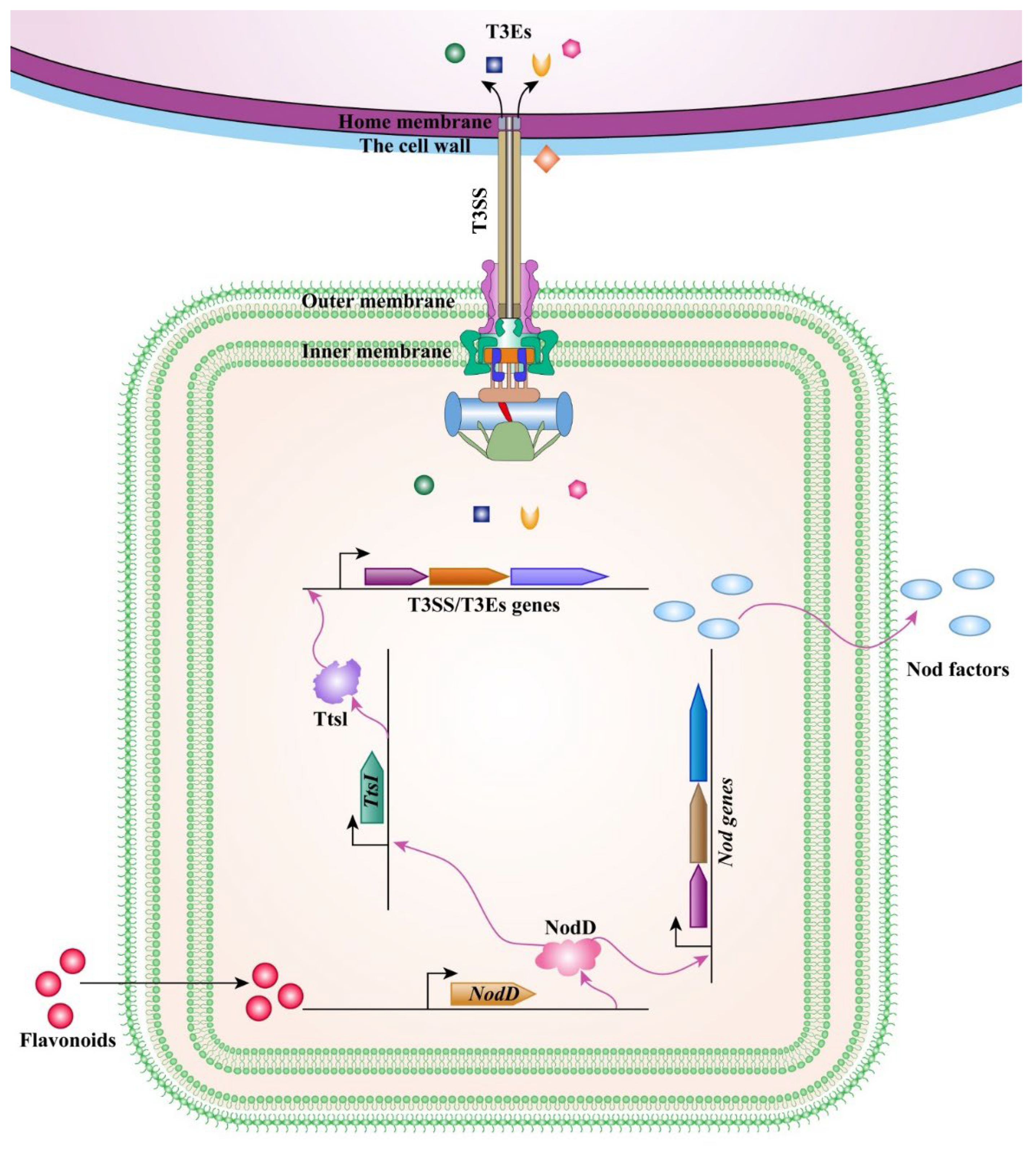
6. Rhizobial Type III Effector Underlying Symbiosis Establishment
7. Conclusions and Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Liu, S.; Zhang, M.; Feng, F.; Tian, Z. Toward a “Green Revolution” for Soybean. Mol. Plant 2020, 13, 688–697. [Google Scholar] [CrossRef] [PubMed]
- Whitham, S.A.; Qi, M.; Innes, R.W.; Ma, W.; Lopes-Caitar, V.; Hewezi, T. Molecular Soybean-Pathogen Interactions. Annu. Rev. Phytopathol. 2016, 54, 443–468. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Liu, W.; Nandety, R.S.; Crook, A.; Mysore, K.S.; Pislariu, C.I.; Frugoli, J.; Dickstein, R.; Udvardi, M.K. Celebrating 20 Years of Genetic Discoveries in Legume Nodulation and Symbiotic Nitrogen Fixation. Plant Cell 2020, 32, 15–41. [Google Scholar] [CrossRef] [PubMed]
- Mitchum, M.G.; Hussey, R.S.; Baum, T.J.; Wang, X.; Elling, A.A.; Wubben, M.; Davis, E.L. Nematode effector proteins: An emerging paradigm of parasitism. New Phytol. 2013, 199, 879–894. [Google Scholar] [CrossRef]
- Okazaki, S.; Kaneko, T.; Sato, S.; Saeki, K. Hijacking of leguminous nodulation signaling by the rhizobial type III secretion system. Proc. Natl. Acad. Sci. USA 2013, 110, 17131–17136. [Google Scholar] [CrossRef]
- Wagner, S.; Grin, I.; Malmsheimer, S.; Singh, N.; Torres-Vargas, C.E.; Westerhausen, S. Bacterial type III secretion systems: A complex device for the delivery of bacterial effector proteins into eukaryotic host cells. FEMS Microbiol. Lett. 2018, 365, fny201. [Google Scholar] [CrossRef]
- Bullones-Bolaños, A.; Bernal-Bayard, J.; Ramos-Morales, F. The NEL Family of Bacterial E3 Ubiquitin Ligases. Int. J. Mol. Sci. 2022, 23, 7725. [Google Scholar] [CrossRef]
- Miwa, H.; Okazaki, S. How effectors promote beneficial interactions. Curr. Opin. Plant Biol. 2017, 38, 148–154. [Google Scholar] [CrossRef]
- Costa, S.R.; Ng, J.L.P.; Mathesius, U. Interaction of Symbiotic Rhizobia and Parasitic Root-Knot Nematodes in Legume Roots: From Molecular Regulation to Field Application. Mol. Plant Microbe Interact. 2021, 34, 470–490. [Google Scholar] [CrossRef]
- Hu, J.; Worrall, L.J.; Strynadka, N.C. Towards capture of dynamic assembly and action of the T3SS at near atomic resolution—ScienceDirect. Curr. Opin. Struct. Biol. 2020, 61, 71–78. [Google Scholar] [CrossRef]
- Buttner, D.; He, S.Y. Type III Protein Secretion in Plant Pathogenic Bacteria. Plant Physiol. 2009, 150, 1656–1664. [Google Scholar] [CrossRef] [PubMed]
- Sugawara, M.; Takahashi, S.; Umehara, Y.; Iwano, H.; Tsurumaru, H.; Odake, H.; Suzuki, Y.; Kondo, H.; Konno, Y.; Yamakawa, T.; et al. Variation in bradyrhizobial NopP effector determines symbiotic incompatibility with Rj2-soybeans via effector-triggered immunity. Nat. Commun. 2018, 9, 3139. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Guerrero, I.; Acosta-Jurado, S.; Medina, C.; Ollero, F.J.; Alias-Villegas, C.; Vinardell, J.M.; Pérez-Montaño, F.; López-Baena, F.J. The Sinorhizobium fredii HH103 type III secretion system effector NopC blocks nodulation with Lotus japonicus Gifu. J. Exp. Bot. 2020, 71, 6043–6056. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Montaño, F.; Jiménez-Guerrero, I.; Acosta-Jurado, S.; Navarro-Gómez, P.; Ollero, F.J.; Ruiz-Sainz, J.E.; López-Baena, F.J.; Vinardell, J.M. A transcriptomic analysis of the effect of genistein on Sinorhizobium fredii HH103 reveals novel rhizobial genes putatively involved in symbiosis. Sci. Rep. 2016, 6, 31592. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Kandoth, P.K.; Warren, S.D.; Yeckel, G.; Heinz, R.; Alden, J.; Yang, C.; Jamai, A.; El-Mellouki, T.; Juvale, P.S.; et al. A soybean cyst nematode resistance gene points to a new mechanism of plant resistance to pathogens. Nature 2012, 492, 256–260. [Google Scholar] [CrossRef] [PubMed]
- Niblack, T.L. Soybean Cyst Nematode Management Reconsidered. Plant Dis. 2005, 89, 1020–1026. [Google Scholar] [CrossRef]
- Qi, M.; Link, T.I.; Müller, M.; Hirschburger, D.; Pudake, R.N.; Pedley, K.F.; Braun, E.; Voegele, R.T.; Baum, T.J.; Whitham, S.A. A Small Cysteine-Rich Protein from the Asian Soybean Rust Fungus, Phakopsora pachyrhizi, Suppresses Plant Immunity. PLoS Pathog. 2016, 12, e1005827. [Google Scholar] [CrossRef]
- Yuan, M.; Ngou, B.P.M.; Ding, P.; Xin, X.F. PTI-ETI crosstalk: An integrative view of plant immunity. Curr. Opin. Plant Biol. 2021, 62, 102030. [Google Scholar] [CrossRef]
- Gassmann, W.; Bhattacharjee, S. Effector-triggered immunity signaling: From gene-for-gene pathways to protein-protein interaction networks. Mol. Plant Microbe Interact. 2012, 25, 862–868. [Google Scholar] [CrossRef]
- Waheed, A.; Haxim, Y.; Islam, W.; Kahar, G.; Liu, X.; Zhang, D. Role of pathogen’s effectors in understanding host-pathogen interaction. Biochim. Biophys. Acta Mol. Cell Res. 2022, 1869, 119347. [Google Scholar] [CrossRef]
- Dodds, P.N.; Rathjen, J.P. Plant immunity: Towards an integrated view of plant-pathogen interactions. Nat. Rev. Genet. 2010, 11, 539–548. [Google Scholar] [CrossRef] [PubMed]
- Block, A.; Li, G.; Fu, Z.Q.; Alfano, J.R. Phytopathogen type III effector weaponry and their plant targets. Curr. Opin. Plant Biol. 2008, 11, 396–403. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Qin, R.; Li, C.; Liu, C.; Jiang, Y.; Yu, J.; Chang, D.; Roberts, P.A.; Chen, Q.; Wang, C. Transgressive resistance to Heterodera glycines in chromosome segment substitution lines derived from susceptible soybean parents. Plant Genome 2021, 14, e20091. [Google Scholar] [CrossRef] [PubMed]
- Wrather, J.A.; Stienstra, W.C.; Koenning, S.R. Soybean disease loss estimates for the United States from 1996 to 1998. Can. J. Plant Pathol. 2001, 23, 122–131. [Google Scholar] [CrossRef]
- Hua, C.; Li, C.; Hu, Y.; Mao, Y.; You, J.; Wang, M.; Chen, J.; Tian, Z.; Wang, C. Identification of HG Types of Soybean Cyst Nematode and Resistance Screening on Soybean Genotypes in Northeast China. J. Nematol. 2018, 50, 41–50. [Google Scholar] [CrossRef]
- Bent, A.F. Exploring Soybean Resistance to Soybean Cyst Nematode. Annu. Rev. Phytopathol. 2022, 60, 379–409. [Google Scholar] [CrossRef]
- Arjoune, Y.; Sugunaraj, N.; Peri, S.; Nair, S.V.; Skurdal, A.; Ranganathan, P.; Johnson, B. Soybean cyst nematode detection and management: A review. Plant Methods 2022, 18, 110. [Google Scholar] [CrossRef]
- Kofsky, J.; Zhang, H.; Song, B.H. Novel resistance strategies to soybean cyst nematode (SCN) in wild soybean. Sci. Rep. 2021, 11, 7967. [Google Scholar] [CrossRef]
- Bissonnette, K.M.; Marett, C.C.; Mullaney, M.P.; Gebhart, G.D.; Kyveryga, P.M.; Mueller, T.A.; Tylka, G.L. Effects of ILeVO Seed Treatment on Heterodera glycines Reproduction and Soybean Yield in Small-Plot and Strip-Trial Experiments in Iowa. Plant Dis. 2020, 104, 2914–2920. [Google Scholar] [CrossRef]
- Clifton, E.H.; Tylka, G.L.; Gassmann, A.J.; Hodgson, E.W. Interactions of effects of host plant resistance and seed treatments on soybean aphid (Aphis glycines Matsumura) and soybean cyst nematode (Heterodera glycines Ichinohe). Pest Manag. Sci. 2018, 74, 992–1000. [Google Scholar] [CrossRef]
- Wang, J.; Niblack, T.L.; Tremain, J.A.; Wiebold, W.J.; Tylka, G.L.; Marett, C.C.; Noel, G.R.; Myers, O.; Schmidt, M.E. Soybean Cyst Nematode Reduces Soybean Yield Without Causing Obvious Aboveground Symptoms. Plant Dis. 2003, 87, 623–628. [Google Scholar] [CrossRef] [PubMed]
- Usovsky, M.; Robbins, R.T.; Fultz Wilkes, J.; Crippen, D.; Shankar, V.; Vuong, T.D.; Agudelo, P.; Nguyen, H.T. Classification Methods and Identification of Reniform Nematode Resistance in Known Soybean Cyst Nematode-Resistant Soybean Genotypes. Plant Dis. 2022, 106, 382–389. [Google Scholar] [CrossRef] [PubMed]
- Davis, E.L.; Hussey, R.S.; Baum, T.J. Getting to the roots of parasitism by nematodes. Trends Parasitol. 2004, 20, 134–141. [Google Scholar] [CrossRef] [PubMed]
- Hewezi, T.; Baum, T.J. Manipulation of plant cells by cyst and root-knot nematode effectors. Mol. Plant Microbe Interact. 2013, 26, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Vieira, P.; Gleason, C. Plant-parasitic nematode effectors—Insights into their diversity and new tools for their identification. Curr. Opin. Plant Biol. 2019, 50, 37–43. [Google Scholar] [CrossRef]
- Ohtsu, M.; Sato, Y.; Kurihara, D.; Suzaki, T.; Kawaguchi, M.; Maruyama, D.; Higashiyama, T. Spatiotemporal deep imaging of syncytium induced by the soybean cyst nematode Heterodera glycines. Protoplasma 2017, 254, 2107–2115. [Google Scholar] [CrossRef]
- Carpentier, J.; Grenier, E.; Esquibet, M.; Hamel, L.P.; Moffett, P.; Manzanares-Dauleux, M.J.; Kerlan, M.C. Evolution and variability of Solanum RanGAP2, a cofactor in the incompatible interaction between the resistance protein GPA2 and the Globodera pallida effector Gp-RBP-1. BMC Evol. Biol. 2013, 13, 87. [Google Scholar] [CrossRef]
- Torres, J.; Wilbers, R.; Gawronski, P.; Boshoven, J.C.; Smant, G. Dual Cf-2-mediated disease resistance in tomato requires a common virulence target of a fungus and a nematode. In Proceedings of the Immunomodulation by Plant-Associated Organisms, Fallen Leaf Lake, CA, USA, 16–19 September 2012. [Google Scholar]
- Bournaud, C.; Gillet, F.-X.; Murad, A.M.; Bresso, E.; Albuquerque, E.V.S.; Grossi-de-Sá, M.F. Meloidogyne incognita PASSE-MURAILLE (MiPM) Gene Encodes a Cell-Penetrating Protein That Interacts With the CSN5 Subunit of the COP9 Signalosome. Front. Plant Sci. 2018, 9, 904. [Google Scholar] [CrossRef]
- Sadia, B.; Domier, L.L.; Biruk, G.; Naoufal, L.; Khalid, M.; Lambert, K.N.; Philippe, C.S. A SNARE-Like Protein and Biotin Are Implicated in Soybean Cyst Nematode Virulence. PLoS ONE 2015, 10, e0145601. [Google Scholar]
- Klink, V.P.; Hosseini, P.; Matsye, P.; Alkharouf, N.W.; Matthews, B.F. A gene expression analysis of syncytia laser microdissected from the roots of the Glycine max (soybean) genotype PI 548402 (Peking) undergoing a resistant reaction after infection by Heterodera glycines (soybean cyst nematode). Plant Mol. Biol. 2009, 71, 525–567. [Google Scholar] [CrossRef]
- Klink, V.P.; Hosseini, P.; Matsye, P.D.; Alkharouf, N.W.; Matthews, B.F. Differences in gene expression amplitude overlie a conserved transcriptomic program occurring between the rapid and potent localized resistant reaction at the syncytium of the Glycine max genotype Peking (PI 548402) as compared to the prolonged and potent resistant reaction of PI 88788. Plant Mol. Biol. 2011, 75, 141–165. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Chen, Q.; Liu, S.; Wang, J.; Peng, D.; Kong, L. Combining targeted metabolite analyses and transcriptomics to reveal the specific chemical composition and associated genes in the incompatible soybean variety PI437654 infected with soybean cyst nematode HG1.2.3.5.7. BMC Plant Biol. 2021, 21, 217. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Kandoth, P.K.; Lakhssassi, N.; Kang, J.; Colantonio, V.; Heinz, R.; Yeckel, G.; Zhou, Z.; Bekal, S.; Dapprich, J.; et al. The soybean GmSNAP18 gene underlies two types of resistance to soybean cyst nematode. Nat. Commun. 2017, 8, 14822. [Google Scholar] [CrossRef] [PubMed]
- Brucker, E.; Carlson, S.; Wright, E.; Niblack, T.; Diers, B. Rhg1 alleles from soybean PI 437654 and PI 88788 respond differentially to isolates of Heterodera glycines in the greenhouse. Appl. Genet. 2005, 111, 44–49. [Google Scholar] [CrossRef]
- Klink, V.P.; Hosseini, P.; Matsye, P.D.; Alkharouf, N.W.; Matthews, B.F. Syncytium gene expression in Glycine max([PI 88788]) roots undergoing a resistant reaction to the parasitic nematode Heterodera glycines. Plant Physiol. Biochem. 2010, 48, 176–193. [Google Scholar] [CrossRef]
- Yuan, C.P.; Wang, Y.J.; Zhao, H.K.; Zhang, L.; Wang, Y.M.; Liu, X.D.; Zhong, X.F.; Dong, Y.S. Genetic diversity of rhg1 and Rhg4 loci in wild soybeans resistant to soybean cyst nematode race 3. Genet. Mol. Res. 2016, 15, gmr.15027386. [Google Scholar] [CrossRef]
- Wu, X.Y.; Zhou, G.C.; Chen, Y.X.; Wu, P.; Liu, L.W.; Ma, F.F.; Wu, M.; Liu, C.C.; Zeng, Y.J.; Chu, A.E.; et al. Soybean Cyst Nematode Resistance Emerged via Artificial Selection of Duplicated Serine Hydroxymethyltransferase Genes. Front. Plant Sci. 2016, 7, 998. [Google Scholar] [CrossRef]
- Appaji Rao, N.; Ambili, M.; Jala, V.R.; Subramanya, H.S.; Savithri, H.S. Structure-function relationship in serine hydroxymethyltransferase. Biochim. Biophys. Acta 2003, 1647, 24–29. [Google Scholar] [CrossRef]
- Hanson, A.D.; Gage, D.A.; Shachar-Hill, Y. Plant one-carbon metabolism and its engineering. Trends Plant Sci. 2000, 5, 206–213. [Google Scholar] [CrossRef]
- Schirch, L. Serine hydroxymethyltransferase. Adv. Enzym. Relat. Areas Mol. Biol. 1982, 53, 83–112. [Google Scholar] [CrossRef]
- Nissan, N.; Mimee, B.; Cober, E.R.; Golshani, A.; Smith, M.; Samanfar, B. A Broad Review of Soybean Research on the Ongoing Race to Overcome Soybean Cyst Nematode. Biology 2022, 11, 211. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Q.; Putker, V.; Goverse, A. Molecular and Cellular Mechanisms Involved in Host-Specific Resistance to Cyst Nematodes in Crops. Front. Plant. Sci. 2021, 12, 641582. [Google Scholar] [CrossRef] [PubMed]
- Lakhssassi, N.; Piya, S.; Bekal, S.; Liu, S.; Zhou, Z.; Bergounioux, C.; Miao, L.; Meksem, J.; Lakhssassi, A.; Jones, K.; et al. A pathogenesis-related protein GmPR08-Bet VI promotes a molecular interaction between the GmSHMT08 and GmSNAP18 in resistance to Heterodera glycines. Plant Biotechnol. J. 2020, 18, 1810–1829. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.S.; Vuong, T.D.; Qiu, D.; Robbins, R.T.; Grover Shannon, J.; Li, Z.; Nguyen, H.T. Advancements in breeding, genetics, and genomics for resistance to three nematode species in soybean. Appl. Genet. 2016, 129, 2295–2311. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Liu, B.; Shi, X.; Reif, J.C.; Guan, R.; Li, Y.-h.; Qiu, L.-j. Deep genotyping of the gene GmSNAP facilitates pyramiding resistance to cyst nematode in soybean. Crop J. 2019, 7, 677–684. [Google Scholar] [CrossRef]
- Suzuki, C.; Taguchi-Shiobara, F.; Ikeda, C.; Iwahashi, M.; Matsui, T.; Yamashita, Y.; Ogura, R. Mapping soybean rhg2 locus, which confers resistance to soybean cyst nematode race 1 in combination with rhg1 and Rhg4 derived from PI 84751. Breed Sci. 2020, 70, 474–480. [Google Scholar] [CrossRef]
- Butler, K.J.; Chen, S.; Smith, J.M.; Wang, X.; Bent, A.F. Soybean Resistance Locus Rhg1 Confers Resistance to Multiple Cyst Nematodes in Diverse Plant Species. Phytopathology 2019, 109, 2107–2115. [Google Scholar] [CrossRef]
- Dong, J.; Hudson, M.E. WI12(Rhg1) interacts with DELLAs and mediates soybean cyst nematode resistance through hormone pathways. Plant Biotechnol. J. 2022, 20, 283–296. [Google Scholar] [CrossRef]
- Yang, D.L.; Yao, J.; Mei, C.S.; Tong, X.H.; Zeng, L.J.; Li, Q.; Xiao, L.T.; Sun, T.P.; Li, J.; Deng, X.W.; et al. Plant hormone jasmonate prioritizes defense over growth by interfering with gibberellin signaling cascade. Proc. Natl. Acad. Sci. USA 2012, 109, E1192–E1200. [Google Scholar] [CrossRef]
- Verma, V.; Ravindran, P.; Kumar, P.P. Plant hormone-mediated regulation of stress responses. BMC Plant Biol. 2016, 16, 86. [Google Scholar] [CrossRef]
- Davière, J.M.; Achard, P. A Pivotal Role of DELLAs in Regulating Multiple Hormone Signals. Mol. Plant 2016, 9, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Ntoukakis, V.; Balmuth, A.L.; Mucyn, T.S.; Gutierrez, J.R.; Jones, A.M.E.; Rathjen, J.P.; Dangl, J.L. The Tomato Prf Complex Is a Molecular Trap for Bacterial Effectors Based on Pto Transphosphorylation. PLoS Pathog. 2013, 9, e1003123. [Google Scholar] [CrossRef] [PubMed]
- Cunnac, S.; Lindeberg, M.; Collmer, A. Pseudomonas syringae type III secretion system effectors: Repertoires in search of functions. Curr. Opin. Microbiol. 2009, 12, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Taguchi, F.; Takeuchi, K.; Katoh, E.; Murata, K.; Suzuki, T.; Marutani, M.; Kawasaki, T.; Eguchi, M.; Katoh, S.; Kaku, H.; et al. Identification of glycosylation genes and glycosylated amino acids of flagellin in Pseudomonas syringae pv. tabaci. Cell. Microbiol. 2006, 8, 923–938. [Google Scholar] [CrossRef]
- Keen, N.T. Gene-for-gene complementarity in plant-pathogen interactions. Annu. Rev. Genet. 1990, 24, 447–463. [Google Scholar] [CrossRef]
- Dangl, J.L.; Jones, J.D.G. Plant pathogens and integrated defence responses to infection. Nature 2001, 411, 826–833. [Google Scholar] [CrossRef]
- Lindeberg, M.; Cartinhour, S.; Myers, C.; Schechter, L.; Schneider, D.; Collmer, A. Closing the Circle on the Discovery of Genes Encoding Hrp Regulon Members and Type III Secretion System Effectors in the Genomes of Three Model Pseudomonas syringae Strains. Mol. Plant-Microbe Interact. 2006, 19, 1151–1158. [Google Scholar] [CrossRef]
- Nomura, K.; Melotto, M.; He, S.-Y. Suppression of host defense in compatible plant-Pseudomonas syringae interactions. Curr. Opin. Plant Biol. 2005, 8, 361–368. [Google Scholar] [CrossRef]
- Abramovitch, R.; Anderson, J.; Martin, G. Bacterial elicitation and evasion of plant innate immunity. Nat. Rev. Mol. Cell Biol. 2006, 7, 601–611. [Google Scholar] [CrossRef]
- Vance, R.E.; Isberg, R.R.; Portnoy, D.A. Patterns of pathogenesis: Discrimination of pathogenic and nonpathogenic microbes by the innate immune system. Cell Host Microbe 2009, 6, 10–21. [Google Scholar] [CrossRef]
- Macho, A.P.; Zipfel, C. Targeting of plant pattern recognition receptor-triggered immunity by bacterial type-III secretion system effectors. Curr. Opin. Microbiol. 2015, 23, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Asai, S.; Shirasu, K. Plant cells under siege: Plant immune system versus pathogen effectors. Curr. Opin. Plant Biol. 2015, 28, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Zipfel, C.; Robatzek, S.; Navarro, L.; Oakeley, E.; Jones, J.; Felix, G.; Boller, T. Bacterial disease resistance in Arabidopsis through flagellin perception. Nature 2004, 428, 764–767. [Google Scholar] [CrossRef] [PubMed]
- Buscaill, P.; Chandrasekar, B.; Sanguankiattichai, N.; Kourelis, J.; Kaschani, F.; Thomas, E.L.; Morimoto, K.; Kaiser, M.; Preston, G.M.; Ichinose, Y.; et al. Glycosidase and glycan polymorphism control hydrolytic release of immunogenic flagellin peptides. Science 2019, 364, eaav0748. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Dunning, F.M.; Pfund, C.; Weingarten, R.; Bent, A.F. Within-species flagellin polymorphism in Xanthomonas campestris pv campestris and its impact on elicitation of Arabidopsis FLAGELLIN SENSING2-dependent defenses. Plant Cell 2006, 18, 764–779. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Sun, Z.; Wang, H.; Liu, L.; Lu, F.; Yang, J.; Zhang, M.; Zhang, S.; Guo, Z.; Bent, A.F.; et al. Rice OsFLS2-Mediated Perception of Bacterial Flagellins Is Evaded by Xanthomonas oryzae pvs. oryzae and oryzicola. Mol. Plant 2015, 8, 1024–1037. [Google Scholar] [CrossRef]
- Shang, Y.; Li, X.; Cui, H.; He, P.; Thilmony, R.; Chintamanani, S.; Zwiesler-Vollick, J.; Gopalan, S.; Tang, X.; Zhou, J.-M. RAR1, a central player in plant immunity, is targeted by Pseudomonas syringae effector AvrB. Proc. Natl. Acad. Sci. USA 2006, 103, 19200–19205. [Google Scholar] [CrossRef]
- Wang, L.; Eggenberger, L.; Hill, J.; Bogdanove, A. Pseudomonas syringae Effector avrB Confers Soybean Cultivar-Specific Avirulence on Soybean mosaic virus Adapted for Transgene Expression but Effector avrPto Does Not. Mol. Plant-Microbe Interact. 2006, 19, 304–312. [Google Scholar] [CrossRef]
- Ong, L.E.; Innes, R.W. AvrB mutants lose both virulence and avirulence activities on soybean and Arabidopsis. Mol. Microbiol. 2006, 60, 951–962. [Google Scholar] [CrossRef]
- Ashfield, T.; Redditt, T.; Russell, A.; Kessens, R.; Rodibaugh, N.; Galloway, L.; Kang, Q.; Podicheti, R.; Innes, R. Evolutionary Relationship of Disease Resistance Genes in Soybean and Arabidopsis Specific for the Pseudomonas syringae Effectors AvrB and AvrRpm1. Plant Physiol. 2014, 166, 235–251. [Google Scholar] [CrossRef]
- Russell, A.; Ashfield, T.; Innes, R. Pseudomonas syringae Effector AvrPphB Suppresses AvrB-Induced Activation of RPM1 but Not AvrRpm1-Induced Activation. Mol. Plant-Microbe Interact. 2015, 28, 727–735. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Cheng, F.; Sun, Y.; Ma, H.; Yang, X.-Q. Structure-Function Relationship of a Novel PR-5 Protein with Antimicrobial Activity from Soy Hulls. J. Agric. Food Chem. 2016, 64, 948–959. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.-M.; Chai, J. Plant pathogenic bacterial type III effectors subdue host responses. Curr. Opin. Microbiol. 2008, 11, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Liu, L.; Bent, A.F. Type III secretion-dependent host defence elicitation and type III secretion-independent growth within leaves by Xanthomonas campestris pv. campestris. Mol. Plant Pathol. 2011, 12, 731–745. [Google Scholar] [CrossRef] [PubMed]
- Büttner, D.; Bonas, U. Regulation and secretion of Xanthomonas virulence factors. FEMS Microbiol. Rev. 2010, 34, 107–133. [Google Scholar] [CrossRef] [PubMed]
- Segonzac, C.; Zipfel, C. Activation of plant pattern-recognition receptors by bacteria. Curr. Opin. Microbiol. 2011, 14, 54–61. [Google Scholar] [CrossRef]
- Boller, T.; Felix, G. A Renaissance of Elicitors: Perception of Microbe-Associated Molecular Patterns and Danger Signals by Pattern-Recognition Receptors. Annu. Rev. Plant Biol. 2009, 60, 379–406. [Google Scholar] [CrossRef]
- De Bruyne, L.; Höfte, M.; De Vleesschauwer, D. Connecting Growth and Defense: The Emerging Roles of Brassinosteroids and Gibberellins in Plant Innate Immunity. Mol. Plant 2014, 7, 943–959. [Google Scholar] [CrossRef]
- Ryan, R.P.; Vorh?Lter, F.J.R.; Potnis, N.; Jones, J.B.; Van Sluys, M.A.; Bogdanove, A.J.; Dow, J.M. Pathogenomics of Xanthomonas: Understanding bacterium-plant interactions. Nat. Rev. Microbiol. 2011, 9, 344–355. [Google Scholar] [CrossRef]
- Swings, J.; Civetta, L. Xanthomonas; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Sharma, A. Xanthomonas. In Encyclopedia of Food Microbiology; Academic Press: Cambridge, MA, USA, 1999. [Google Scholar]
- Starr, M.P.; Stolp, H.; Trüper, H.G.; Balows, A.; Schlegel, H.G. The Genus Xanthomonas. In The Prokaryotes: A Handbook on Habitats. Isolation and Identification of Bacteria; Springer: Berlin, Germany, 1981; pp. 742–763. [Google Scholar]
- Leyns, F.; De Cleene, M.; Swings, J.-G.; De Ley, J. The host range of the genusXanthomonas. Bot. Rev. 1984, 50, 308–356. [Google Scholar] [CrossRef]
- Zipfel, C.; Kunze, G.; Chinchilla, D.; Caniard, A.; Jones, J.D.G.; Boller, T.; Felix, G. Perception of the bacterial PAMP EF-Tu by the receptor EFR restricts Agrobacterium-mediated transformation. Cell 2006, 125, 749–760. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.M.; Mansfield, J.; Grant, M. Considerations on post-translational modification and protein targeting in the arabidopsis defense proteome. Plant Signal. Behav. 2007, 2, 153–154. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Tsuda, K.; Katagiri, F. Comparing signaling mechanisms engaged in pattern-triggered and effector-triggered immunity. Curr. Opin. Plant Biol. 2010, 13, 459–465. [Google Scholar] [CrossRef] [PubMed]
- Gerlach, R.G.; Hensel, M. Protein secretion systems and adhesins: The molecular armory of Gram-negative pathogens. Int. J. Med. Microbiol. 2007, 297, 401–415. [Google Scholar] [CrossRef]
- Büttner, D.; Noël, L.; Thieme, F.; Bonas, U. Genomic approaches in Xanthomonas campestris pv. vesicatoria allow fishing for virulence genes. J. Biotechnol. 2003, 106, 203–214. [Google Scholar] [CrossRef]
- Tampakaki, A.P.; Fadouloglou, V.E.; Gazi, A.D.; Panopoulos, N.J.; Kokkinidis, M. Conserved features of type III secretion. Cell. Microbiol. 2010, 6, 805–816. [Google Scholar] [CrossRef]
- Newberry, E.A.; Bhandari, R.; Minsavage, G.V.; Timilsina, S.; Potnis, N. Independent Evolution with the Gene Flux Originating from Multiple Xanthomonas Species Explains Genomic Heterogeneity in Xanthomonas perforans. Appl. Environ. Microbiol. 2019, 85, e00885-19. [Google Scholar] [CrossRef]
- Constantin, E.C.; Haegeman, A.; Van Vaerenbergh, J.; Baeyen, S.; Van Malderghem, C.; Maes, M.; Cottyn, B. Pathogenicity and virulence gene content of Xanthomonas strains infecting Araceae, formerly known as Xanthomonas axonopodis pv. dieffenbachiae. Plant Pathol. 2017, 66, 1539–1554. [Google Scholar] [CrossRef]
- Ichida, H.; Maeda, K.; Ichise, H.; Matsuyama, T.; Abe, T.; Yoneyama, K.; Koba, T. In silico restriction landmark genome scanning analysis of Xanthomonas oryzae pathovar oryzae MAFF 311018. Biochem. Biophys. Res. Commun. 2007, 363, 852–856. [Google Scholar] [CrossRef]
- da Silva, A.C.; Ferro, J.A.; Reinach, F.D.; Farah, C.S.; Furlan, L.R.; Quaggio, R.B.; Monteiro-Vitorello, C.B.; Van Sluys, M.A.; Almeida, N.A.; Alves, L.M.; et al. Comparison of the genomes of two Xanthomonas pathogens with differing host specificities. Nature 2002, 417, 459–463. [Google Scholar] [CrossRef]
- White, F.F.; Potnis, N.; Jones, J.B.; Koebnik, R. The type III effectors of Xanthomonas. Mol. Plant Pathol. 2010, 10, 749–766. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.G.; Taylor, K.W.; Hotson, A.; Keegan, M.; Schmelz, E.A.; Mudgett, M.B. XopD SUMO Protease Affects Host Transcription, Promotes Pathogen Growth, and Delays Symptom Development in Xanthomonas-Infected Tomato Leaves. Plant Cell 2008, 20, 1915–1929. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.G.; Stork, W.; Mudgett, M.B. Xanthomonas type III effector XopD desumoylates tomato transcription factor SlERF4 to suppress ethylene responses and promote pathogen growth. Cell Host Microbe 2013, 13, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-G.; Li, X.; Roden, J.A.; Taylor, K.W.; Aakre, C.D.; Su, B.; Lalonde, S.; Kirik, A.; Chen, Y.; Baranage, G. Xanthomonas T3S Effector XopN Suppresses PAMP-Triggered Immunity and Interacts with a Tomato Atypical Receptor-Like Kinase and TFT1. Plant Cell Online 2009, 21, 1305–1323. [Google Scholar] [CrossRef] [PubMed]
- Jiang, B.L.; He, Y.Q.; Cen, W.J.; Wei, H.Y.; Jiang, G.F.; Jiang, W.; Hang, X.H.; Feng, J.X.; Lu, G.T.; Tang, D.J. The type III secretion effector XopXccN of Xanthomonas campestris pv. campestris is required for full virulence. Res. Microbiol. 2008, 159, 216–220. [Google Scholar] [CrossRef] [PubMed]
- Deb, S.; Ghosh, P.; Patel, H.K.; Sonti, R.V. Interaction of the Xanthomonas effectors XopQ and XopX results in induction of rice immune responses. Plant J. 2020, 104, 332–350. [Google Scholar] [CrossRef]
- Ishikawa, K.; Yamaguchi, K.; Sakamoto, K.; Yoshimura, S.; Inoue, K.; Tsuge, S.; Kojima, C.; Kawasaki, T. Bacterial effector modulation of host E3 ligase activity suppresses PAMP-triggered immunity in rice. Nat. Commun. 2014, 5, 5430. [Google Scholar] [CrossRef]
- Swords, K.; Dahlbeck, D.; Kearney, B.; Roy, M.; Staskawicz, B.J. Spontaneous and induced mutations in a single open reading frame alter both virulence and avirulence in Xanthomonas campestris pv. vesicatoria avrBs2. J. Bacteriol. 1996, 178, 4661–4669. [Google Scholar] [CrossRef]
- Gürlebeck, D.; Thieme, F.; Bonas, U. Type III effector proteins from the plant pathogen Xanthomonas and their role in the interaction with the host plant. J. Plant Physiol. 2006, 163, 233–255. [Google Scholar] [CrossRef]
- Li, S.; Wang, Y.; Wang, S.; Fang, A.; Sun, W. The Type III Effector AvrBs2 in Xanthomonas oryzae pv. oryzicola Suppresses Rice Immunity and Promotes Disease Development Running title: AvrBs2 in Xoc suppresses rice immunity. Mol. Plant-Microbe Interact. 2015, 28, 869–880. [Google Scholar] [CrossRef]
- Nissan, G.; Manulis-Sasson, S.; Weinthal, D.; Mor, H.; Sessa, G.; Barash, I. The type III effectors HsvG and HsvB of gall-forming Pantoea agglomerans determine host specificity and function as transcriptional activators. Mol. Microbiol. 2006, 61, 1118–1131. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; White, F.F. Diverse Members of the AvrBs3/PthA Family of Type III Effectors Are Major Virulence Determinants in Bacterial Blight Disease of Rice. Mol. Plant-Microbe Interact. 2004, 17, 1192–1200. [Google Scholar] [CrossRef] [PubMed]
- Swarup, S. A pathogenicity locus from Xanthomonas citri enable strains from several pathovars ofelicit can-ker-like lesion on ctirus. Phytopathology 1991, 81, 802–809. [Google Scholar] [CrossRef]
- Yang, B.; Sugio, A.; White, F. Os8N3 is a host disease-susceptibility gene for bacterial blight of rice. Proc. Natl. Acad. Sci. USA 2006, 103, 10503–10508. [Google Scholar] [CrossRef] [PubMed]
- Kladsuwan, L.; Athinuwat, D.; Bogdanove, A.J.; Prathuangwong, S. AvrBs3-like genes and TAL effectors specific to race structure in Xanthomonas axonopodis pv. glycines. Thai J. Agric. Sci. 2017, 50, 121–145. [Google Scholar]
- Athinuwat, D.; Prathuangwong, S.; Cursino, L.; Burr, T. Xanthomonas axonopodis pv. glycines Soybean Cultivar Virulence Specificity Is Determined by avrBs3 Homolog avrXg1. Phytopathology 2009, 99, 996–1004. [Google Scholar] [CrossRef]
- Vauterin, L.; Hoste, B.; Kersters, K.; Swings, J. Reclassification of Xanthomonas. Int. J. Syst. Bacteriol. 1995, 45, 472–489. [Google Scholar] [CrossRef]
- Narvel, J.M.; Jakkula, L.R.; Phillips, D.V.; Wang, T.; Leem, S.-H.; Boerma, H.R. Molecular Mapping of Rxp Conditioning Reaction to Bacterial Pustule in Soybean. J. Hered. 2001, 92, 267–270. [Google Scholar] [CrossRef]
- Guo, W.; Gao, J.; Chen, Q.; Ma, B.; Fang, Y.; Liu, X.; Chen, G.; Liu, J. Crp-like protein (Clp) plays both positive and negative roles in regulating the pathogenicity of bacterial pustule pathogen Xanthomonas axonopodis pv. glycines. Phytopathology 2019, 109, 1171–1183. [Google Scholar] [CrossRef]
- Prathuangwong, S.; Amnuaykit, K. Studies on tolerance and rate-reducing bacterial pustule of soybean cultivars/lines. Witthayasan Kasetsart Sakha Witthayasat 1987, 21, 408–426. [Google Scholar]
- Chatnaparat, T.; Prathuangwong, S.; Lindow, S.E. Global Pattern of Gene Expression of Xanthomonas axonopodis pv. glycines Within Soybean Leaves. Mol. Plant Microbe Interact. 2016, 29, 508–522. [Google Scholar] [CrossRef] [PubMed]
- Athinuwat, D.; Brooks, S.; Burr, T.J.; Prathuangwong, S. Flagella and Pili of Xanthomonas axonopodis pv. glycines are associated with motility, biofilm formation and virulence on soybean. J. Phytopathol. 2018, 166, 590–600. [Google Scholar] [CrossRef]
- Carpenter, S.; Lawan, K.; Sang-Wook, H.; Sutruedee, P.; Bogdanove, A.J. Complete Genome Sequences of Xanthomonas axonopodis pv. glycines Isolates from the United States and Thailand Reveal Conserved Transcription Activator-Like Effectors. Genome Biol. Evol. 2019, 11, 1380–1384. [Google Scholar] [CrossRef] [PubMed]
- Gómez, J.; Vital, C.E.; Oliveira, M.; Ramos, H.; Lightfoot, D.A. Broad range flavonoid profiling by LC/MS of soybean genotypes contrasting for resistance to Anticarsia gemmatalis (Lepidoptera: Noctuidae). PLoS ONE 2018, 13, e0205010. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.J.; Pang, Y.B.; Wang, X.Y.; Jiang, Y.H.; Herrera-Balandrano, D.D.; Jin, Y.; Wang, S.Y.; Laborda, P. Exogenous genistein enhances soybean resistance to Xanthomonas axonopodis pv. glycines. Pest Manag. Sci. 2022, 78, 3664–3675. [Google Scholar] [CrossRef]
- Athinuwat, D.; Brooks, S. The OmpA Gene of Xanthomonas axonopodis pv. glycines is Involved in Pathogenesis of Pustule Disease on Soybean. Curr. Microbiol. 2019, 76, 879–887. [Google Scholar] [CrossRef]
- Bonas, U.; Stall, R.E.; Staskawicz, B. Genetic and structural characterization of the avirulence gene avrBs3 from Xanthomonas campestris pv. vesicatoria. Mol. Gen. Genet. 1989, 218, 127–136. [Google Scholar] [CrossRef]
- Shiotani, H.; Ozaki, K.; Tsuyumu, S. Pathogenic Interactions between Xanthomonas axonopodis pv. citri and Cultivars of Pummelo (Citrus grandis). Phytopathology 2000, 90, 1383–1389. [Google Scholar] [CrossRef]
- Yang, Y. Watersoaking function(s) of xcmh1005 are redundantly encoded by members of the xanthomonas avr/pth gene family. Mol. Plant-Microbe Interact. 1996, 9, 105–113. [Google Scholar] [CrossRef]
- Kanamori, H.; Tsuyumu, S. Comparison of nucleotide sequences of canker-forming and non-canker-forming homologues in pv. Ann. Phytopathol. Soc. Jpn. 1998, 64, 462–470. [Google Scholar] [CrossRef]
- Kim, J.G.; Choi, S.; Oh, J.; Moon, J.S.; Hwang, I. Comparative analysis of three indigenous plasmids from Xanthomonas axonopodis pv. glycines. Plasmid 2006, 56, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Staskawicz, B.J.; Dahlbeck, D.; Keen, N.T. Cloned avirulence gene of Pseudomonas syringae pv. glycinea determines race-specific incompatibility on Glycine max (L.) Merr. Proc. Natl. Acad. Sci. USA 1984, 81, 6024–6028. [Google Scholar] [CrossRef] [PubMed]
- Grant, S.R.; Fisher, E.J.; Chang, J.H.; Mole, B.M.; Dangl, J.L. Subterfuge and manipulation: Type III effector proteins of phytopathogenic bacteria. Annu. Rev. Microbiol. 2006, 60, 425–449. [Google Scholar] [CrossRef] [PubMed]
- Chinchilla, D.; Zipfel, C.; Robatzek, S.; Kemmerling, B.; Nürnberger, T.; Jones, J.D.; Felix, G.; Boller, T. A flagellin-induced complex of the receptor FLS2 and BAK1 initiates plant defence. Nature 2007, 448, 497–500. [Google Scholar] [CrossRef] [PubMed]
- Lu, D.; Wu, S.; Gao, X.; Zhang, Y.; Shan, L.; He, P. A receptor-like cytoplasmic kinase, BIK1, associates with a flagellin receptor complex to initiate plant innate immunity. Proc. Natl. Acad. Sci. USA 2010, 107, 496–501. [Google Scholar] [CrossRef]
- Xin, X.-F.; He, S.Y. Pseudomonas syringae pv. tomato DC3000: A model pathogen for probing disease susceptibility and hormone signaling in plants. Annu. Rev. Phytopathol. 2013, 51, 473–498. [Google Scholar] [CrossRef]
- Sonnewald, S.; Priller, J.P.; Schuster, J.; Glickmann, E.; Hajirezaei, M.-R.; Siebig, S.; Mudgett, M.B.; Sonnewald, U. Regulation of cell wall-bound invertase in pepper leaves by Xanthomonas campestris pv. vesicatoria type three effectors. PLoS ONE 2012, 7, e51763. [Google Scholar] [CrossRef]
- Rodríguez-Herva, J.J.; González-Melendi, P.; Cuartas-Lanza, R.; Antúnez-Lamas, M.; Río-Alvarez, I.; Li, Z.; López-Torrejón, G.; Díaz, I.; Del Pozo, J.C.; Chakravarthy, S. A bacterial cysteine protease effector protein interferes with photosynthesis to suppress plant innate immune responses. Cell. Microbiol. 2012, 14, 669–681. [Google Scholar] [CrossRef]
- Feng, F.; Zhou, J.-M. Plant–bacterial pathogen interactions mediated by type III effectors. Curr. Opin. Plant Biol. 2012, 15, 469–476. [Google Scholar] [CrossRef]
- Zhang, J.; Shao, F.; Li, Y.; Cui, H.; Chen, L.; Li, H.; Zou, Y.; Long, C.; Lan, L.; Chai, J. A Pseudomonas syringae effector inactivates MAPKs to suppress PAMP-induced immunity in plants. Cell Host Microbe 2007, 1, 175–185. [Google Scholar] [CrossRef]
- Teper, D.; Salomon, D.; Sunitha, S.; Kim, J.G.; Mudgett, M.B.; Sessa, G. Xanthomonas euvesicatoria type III effector X op Q interacts with tomato and pepper 14–3–3 isoforms to suppress effector-triggered immunity. Plant J. 2014, 77, 297–309. [Google Scholar] [CrossRef]
- López-Baena, F.J.; Ruiz-Sainz, J.E.; Rodríguez-Carvajal, M.A.; Vinardell, J.M. Bacterial Molecular Signals in the Sinorhizobium fredii-Soybean Symbiosis. Int. J. Mol. Sci. 2016, 17, 755. [Google Scholar] [CrossRef] [PubMed]
- Roy, S. Goldilocks Principle: MtNFH1 Ensures Optimal Nod Factor Activity. Plant Cell 2018, 30, 267–268. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Halane, M.K.; Gassmann, W.; Stacey, G. The Role of Plant Innate Immunity in the Legume-Rhizobium Symbiosis. Annu. Rev. Plant Biol. 2017, 68, 535–561. [Google Scholar] [CrossRef]
- Teulet, A.; Camuel, A.; Perret, X.; Giraud, E. The Versatile Roles of Type III Secretion Systems in Rhizobia-Legume Symbioses. Annu. Rev. Microbiol. 2022, 76, 45–65. [Google Scholar] [CrossRef]
- Deng, W.; Marshall, N.C.; Rowland, J.L.; McCoy, J.M.; Worrall, L.J.; Santos, A.S.; Strynadka, N.C.J.; Finlay, B.B. Assembly, structure, function and regulation of type III secretion systems. Nat. Rev. Microbiol. 2017, 15, 323–337. [Google Scholar] [CrossRef] [PubMed]
- Kambara, K.; Ardissone, S.; Kobayashi, H.; Saad, M.M.; Schumpp, O.; Broughton, W.J.; Deakin, W.J. Rhizobia utilize pathogen-like effector proteins during symbiosis. Mol. Microbiol. 2009, 71, 92–106. [Google Scholar] [CrossRef] [PubMed]
- Büttner, D. Protein export according to schedule: Architecture, assembly, and regulation of type III secretion systems from plant- and animal-pathogenic bacteria. Microbiol. Mol. Biol. Rev. 2012, 76, 262–310. [Google Scholar] [CrossRef] [PubMed]
- Abby, S.S.; Rocha, E.P. The non-flagellar type III secretion system evolved from the bacterial flagellum and diversified into host-cell adapted systems. PLoS Genet. 2012, 8, e1002983. [Google Scholar] [CrossRef]
- Bergeron, J.R.C.; Fernández, L.; Wasney, G.A.; Vuckovic, M.; Reffuveille, F.; Hancock, R.E.W.; Strynadka, N.C.J. The Structure of a Type 3 Secretion System (T3SS) Ruler Protein Suggests a Molecular Mechanism for Needle Length Sensing. J. Biol. Chem. 2016, 291, 1676–1691. [Google Scholar] [CrossRef]
- Deakin, W.J.; Marie, C.; Saad, M.M.; Krishnan, H.B.; Broughton, W.J. NopA is associated with cell surface appendages produced by the type III secretion system of Rhizobium sp. strain NGR234. Mol. Plant Microbe Interact. 2005, 18, 499–507. [Google Scholar] [CrossRef] [PubMed]
- Lorio, J.C.; Kim, W.S.; Krishnan, H.B. NopB, a soybean cultivar-specificity protein from Sinorhizobium fredii USDA257, is a type III secreted protein. Mol. Plant Microbe Interact. 2004, 17, 1259–1268. [Google Scholar] [CrossRef] [PubMed]
- Teulet, A.; Gully, D.; Rouy, Z.; Camuel, A.; Koebnik, R.; Giraud, E.; Lassalle, F. Phylogenetic distribution and evolutionary dynamics of nod and T3SS genes in the genus Bradyrhizobium. Microb. Genomes 2020, 6, mgen000407. [Google Scholar] [CrossRef] [PubMed]
- Reeve, W.; van Berkum, P.; Ardley, J.; Tian, R.; Gollagher, M.; Marinova, D.; Elia, P.; Reddy, T.B.K.; Pillay, M.; Varghese, N.; et al. High-quality permanent draft genome sequence of the Bradyrhizobium elkanii type strain USDA 76(T), isolated from Glycine max (L.) Merr. Stand Genom. Sci. 2017, 12, 26. [Google Scholar] [CrossRef] [PubMed]
- Quilbé, J.; Montiel, J.; Arrighi, J.F.; Stougaard, J. Molecular Mechanisms of Intercellular Rhizobial Infection: Novel Findings of an Ancient Process. Front. Plant Sci. 2022, 13, 922982. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Guerrero, I.; Pérez-Montaño, F.; Medina, C.; Ollero, F.J.; López-Baena, F.J. NopC Is a Rhizobium-Specific Type 3 Secretion System Effector Secreted by Sinorhizobium (Ensifer) fredii HH103. PLoS ONE 2015, 10, e0142866. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Ma, C.; Ma, S.; Zheng, H.; Tian, H.; Wang, X.; Wang, Y.; Jiang, H.; Wang, J.; Zhang, Z.; et al. Genetic variation in GmCRP contributes to nodulation in soybean (Glycine max Merr.). Crop J. 2022; in press. [Google Scholar] [CrossRef]
- Rodrigues, J.A.; López-Baena, F.J.; Ollero, F.J.; Vinardell, J.M.; Espuny Mdel, R.; Bellogín, R.A.; Ruiz-Sainz, J.E.; Thomas, J.R.; Sumpton, D.; Ault, J.; et al. NopM and NopD are rhizobial nodulation outer proteins: Identification using LC-MALDI and LC-ESI with a monolithic capillary column. J. Proteome Res. 2007, 6, 1029–1037. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Q.W.; Bai, J.; Cai, J.; Huang, Q.Y.; Wang, Y.; Liang, Y.; Zhong, Z.; Wagner, C.; Xie, Z.P.; Staehelin, C. NopD of Bradyrhizobium sp. XS1150 Possesses SUMO Protease Activity. Front. Microbiol. 2020, 11, 386. [Google Scholar] [CrossRef]
- Canonne, J.; Marino, D.; Jauneau, A.; Pouzet, C.; Brière, C.; Roby, D.; Rivas, S. The Xanthomonas type III effector XopD targets the Arabidopsis transcription factor MYB30 to suppress plant defense. Plant Cell 2011, 23, 3498–3511. [Google Scholar] [CrossRef]
- Wang, J.; Wang, J.; Ma, C.; Zhou, Z.; Yang, D.; Zheng, J.; Wang, Q.; Li, H.; Zhou, H.; Sun, Z.; et al. QTL Mapping and Data Mining to Identify Genes Associated With the Sinorhizobium fredii HH103 T3SS Effector NopD in Soybean. Front. Plant Sci. 2020, 11, 453. [Google Scholar] [CrossRef]
- Skorpil, P.; Saad, M.M.; Boukli, N.M.; Kobayashi, H.; Ares-Orpel, F.; Broughton, W.J.; Deakin, W.J. NopP, a phosphorylated effector of Rhizobium sp. strain NGR234, is a major determinant of nodulation of the tropical legumes Flemingia congesta and Tephrosia vogelii. Mol. Microbiol 2005, 57, 1304–1317. [Google Scholar] [CrossRef]
- Jiménez-Guerrero, I.; Pérez-Montaño, F.; Medina, C.; Ollero, F.J.; López-Baena, F.J. The Sinorhizobium (Ensifer) fredii HH103 Nodulation Outer Protein NopI Is a Determinant for Efficient Nodulation of Soybean and Cowpea Plants. Appl. Environ. Microbiol. 2017, 83, e02770-16. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, J.; Liu, C.; Ma, C.; Li, C.; Zhang, Y.; Qi, Z.; Zhu, R.; Shi, Y.; Zou, J.; et al. Identification of Soybean Genes Whose Expression is Affected by the Ensifer fredii HH103 Effector Protein NopP. Int. J. Mol. Sci. 2018, 19, 3438. [Google Scholar] [CrossRef] [PubMed]
- Tsukui, T.; Eda, S.; Kaneko, T.; Sato, S.; Okazaki, S.; Kakizaki-Chiba, K.; Itakura, M.; Mitsui, H.; Yamashita, A.; Terasawa, K.; et al. The type III Secretion System of Bradyrhizobium japonicum USDA122 mediates symbiotic incompatibility with Rj2 soybean plants. Appl. Environ. Microbiol. 2013, 79, 1048–1051. [Google Scholar] [CrossRef]
- Zhang, B.; Wang, M.; Sun, Y.; Zhao, P.; Liu, C.; Qing, K.; Hu, X.; Zhong, Z.; Cheng, J.; Wang, H.; et al. Glycine max NNL1 restricts symbiotic compatibility with widely distributed bradyrhizobia via root hair infection. Nat. Plants 2021, 7, 73–86. [Google Scholar] [CrossRef] [PubMed]
- Bartsev, A.V.; Deakin, W.J.; Boukli, N.M.; McAlvin, C.B.; Stacey, G.; Malnoë, P.; Broughton, W.J.; Staehelin, C. NopL, an effector protein of Rhizobium sp. NGR234, thwarts activation of plant defense reactions. Plant Physiol. 2004, 134, 871–879. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Chen, X.J.; Lu, H.B.; Xie, Z.P.; Staehelin, C. Functional analysis of the type 3 effector nodulation outer protein L (NopL) from Rhizobium sp. NGR234: Symbiotic effects, phosphorylation, and interference with mitogen-activated protein kinase signaling. J. Biol. Chem. 2011, 286, 32178–32187. [Google Scholar] [CrossRef]
- Ge, Y.Y.; Xiang, Q.W.; Wagner, C.; Zhang, D.; Xie, Z.P.; Staehelin, C. The type 3 effector NopL of Sinorhizobium sp. strain NGR234 is a mitogen-activated protein kinase substrate. J. Exp. Bot. 2016, 67, 2483–2494. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, X.; Chen, L.; Fu, Y.; Li, C.; Qi, Z.; Zou, J.; Zhu, R.; Li, S.; Wei, W.; et al. Mining for genes encoding proteins associated with NopL of Sinorhizobium fredii HH103 using quantitative trait loci in soybean (Glycine max Merr.) recombinant inbred lines. Plant Soil 2018, 431, 245–255. [Google Scholar] [CrossRef]
- Xin, D.W.; Liao, S.; Xie, Z.P.; Hann, D.R.; Steinle, L.; Boller, T.; Staehelin, C. Functional analysis of NopM, a novel E3 ubiquitin ligase (NEL) domain effector of Rhizobium sp. strain NGR234. PLoS Pathog. 2012, 8, e1002707. [Google Scholar] [CrossRef]
- Xu, C.C.; Zhang, D.; Hann, D.R.; Xie, Z.P.; Staehelin, C. Biochemical properties and in planta effects of NopM, a rhizobial E3 ubiquitin ligase. J. Biol. Chem. 2018, 293, 15304–15315. [Google Scholar] [CrossRef] [PubMed]
- Nakano, M.; Oda, K.; Mukaihara, T. Ralstonia solanacearum novel E3 ubiquitin ligase (NEL) effectors RipAW and RipAR suppress pattern-triggered immunity in plants. Microbiology 2017, 163, 992–1002. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Wadood, S.F.; Chen, M.; Wang, Y.; Xie, Z.P.; Staehelin, C. Effector-triggered inhibition of nodulation: A rhizobial effector protease targets soybean kinase GmPBS1-1. Plant Physiol. 2022, 189, 2382–2395. [Google Scholar] [CrossRef]
- Qi, D.; Dubiella, U.; Kim, S.H.; Sloss, D.I.; Dowen, R.H.; Dixon, J.E.; Innes, R.W. Recognition of the protein kinase AVRPPHB SUSCEPTIBLE1 by the disease resistance protein RESISTANCE TO PSEUDOMONAS SYRINGAE5 is dependent on s-acylation and an exposed loop in AVRPPHB SUSCEPTIBLE1. Plant Physiol. 2014, 164, 340–351. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Wang, Z.; Shabab, M.; Oeljeklaus, J.; Verhelst, S.H.; Kaschani, F.; Kaiser, M.; Bogyo, M.; van der Hoorn, R.A. A substrate-inspired probe monitors translocation, activation, and subcellular targeting of bacterial type III effector protease AvrPphB. Chem. Biol. 2013, 20, 168–176. [Google Scholar] [CrossRef]
- Luo, Y.; Liu, D.; Jiao, S.; Liu, S.; Wang, X.; Shen, X.; Wei, G. Identification of Robinia pseudoacacia target proteins responsive to Mesorhizobium amphore CCNWGS0123 effector protein NopT. J. Exp. Bot. 2020, 71, 7347–7363. [Google Scholar] [CrossRef]
- Jiménez-Guerrero, I.; Pérez-Montaño, F.; Zdyb, A.; Beutler, M.; Werner, G.; Göttfert, M.; Ollero, F.J.; Vinardell, J.M.; López-Baena, F.J. GunA of Sinorhizobium (Ensifer) fredii HH103 is a T3SS-secreted cellulase that differentially affects symbiosis with cowpea and soybean. Plant Soil 2019, 435, 15–26. [Google Scholar] [CrossRef]
- Wang, J.; Ma, C.; Ma, S.; Zheng, H.; Feng, H.; Wang, Y.; Wang, J.; Liu, C.; Xin, D.; Chen, Q.; et al. GmARP is Related to the Type III Effector NopAA to Promote Nodulation in Soybean (Glycine max). Front. Genet. 2022, 13, 889795. [Google Scholar] [CrossRef]
- Ma, Z.; Song, T.; Zhu, L.; Ye, W.; Wang, Y.; Shao, Y.; Dong, S.; Zhang, Z.; Dou, D.; Zheng, X.; et al. A Phytophthora sojae Glycoside Hydrolase 12 Protein Is a Major Virulence Factor during Soybean Infection and Is Recognized as a PAMP. Plant Cell 2015, 27, 2057–2072. [Google Scholar] [CrossRef]
- Ma, Z.; Zhu, L.; Song, T.; Wang, Y.; Zhang, Q.; Xia, Y.; Qiu, M.; Lin, Y.; Li, H.; Kong, L.; et al. A paralogous decoy protects Phytophthora sojae apoplastic effector PsXEG1 from a host inhibitor. Science 2017, 355, 710–714. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Composition | Function | Target in Host Cells | Effect of Mutation on Nodule |
|---|---|---|---|---|
| Structural Components of T3SS | ||||
| 1 | RhcN | ATPase | - | Nod− |
| 2 | NolV | Stator | - | - |
| 3 | RhcQ | Cytoplasmic ring | - | - |
| 4 | RhcO | Stalk | - | - |
| 5 | RhcV | Export gate | - | - |
| 6 | RhcU | Autoprotease | - | - |
| 7 | RhcR | Inner membrane component | - | - |
| 8 | RhcS | Inner membrane component | - | - |
| 9 | RhcT | Inner membrane component | - | - |
| 10 | RhcJ | Inner membrane ring | - | - |
| 11 | RhcD | Inner membrane ring | - | - |
| 12 | RhcC1 | Secretin, outer membrane ring | - | - |
| 13 | NolU | Inner rod | - | Nod− |
| 14 | RhcC1 | Secretin, outer membrane ring | - | Nod− |
| 15 | NopA | Needle | - | Nod− |
| 16 | NopB | Needle | - | Nod− |
| 17 | NopX | Translocation pore | - | Nod− and Delayed nodulation |
| 18 | NopE | Translocation pore | - | - |
| 19 | NopH | Translocation pore | - | - |
| Secretable effector of T3Es | ||||
| 20 | NopC | Suppress immunity | Unknown | Nod− |
| 21 | NopD | DeSUMOylation | Rj4 | Nod− |
| 22 | NopL | Substrate for plant kinase | SIPK | Nod− |
| 23 | NopJ | Putative acetyltransferase | Unknown | Nod− or Nod+ |
| 24 | NopM | E3-ubiquitin ligase | Unknown | Nod− |
| 25 | NopP | Substrate for plant kinase | NNL1 | Nod− |
| 26 | NopT | Cysteine proteases | PBS1 | Nod− or Nod+ |
| 27 | NopZ | Putative effector | Unknown | - |
| 28 | InnB | Unknown | Unknown | Nod− |
| 29 | NopAA | Cellulase | Unknown | Nod− |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Ni, H.; Chen, L.; Zou, J.; Liu, C.; Chen, Q.; Ratet, P.; Xin, D. Effector-Dependent and -Independent Molecular Mechanisms of Soybean–Microbe Interaction. Int. J. Mol. Sci. 2022, 23, 14184. https://doi.org/10.3390/ijms232214184
Wang J, Ni H, Chen L, Zou J, Liu C, Chen Q, Ratet P, Xin D. Effector-Dependent and -Independent Molecular Mechanisms of Soybean–Microbe Interaction. International Journal of Molecular Sciences. 2022; 23(22):14184. https://doi.org/10.3390/ijms232214184
Chicago/Turabian StyleWang, Jinhui, Hejia Ni, Lin Chen, Jianan Zou, Chunyan Liu, Qingshan Chen, Pascal Ratet, and Dawei Xin. 2022. "Effector-Dependent and -Independent Molecular Mechanisms of Soybean–Microbe Interaction" International Journal of Molecular Sciences 23, no. 22: 14184. https://doi.org/10.3390/ijms232214184
APA StyleWang, J., Ni, H., Chen, L., Zou, J., Liu, C., Chen, Q., Ratet, P., & Xin, D. (2022). Effector-Dependent and -Independent Molecular Mechanisms of Soybean–Microbe Interaction. International Journal of Molecular Sciences, 23(22), 14184. https://doi.org/10.3390/ijms232214184