
The Hypothalamic–Pituitary–Thyroid Axis Equivalent in Normal and Cancerous Oral Tissues: A Scoping Review



Abstract
1. Introduction
2. Materials and Methods
2.1. Protocol and Search Strategy
2.2. Eligibility Criteria
2.3. Study Selection and Date of Collection
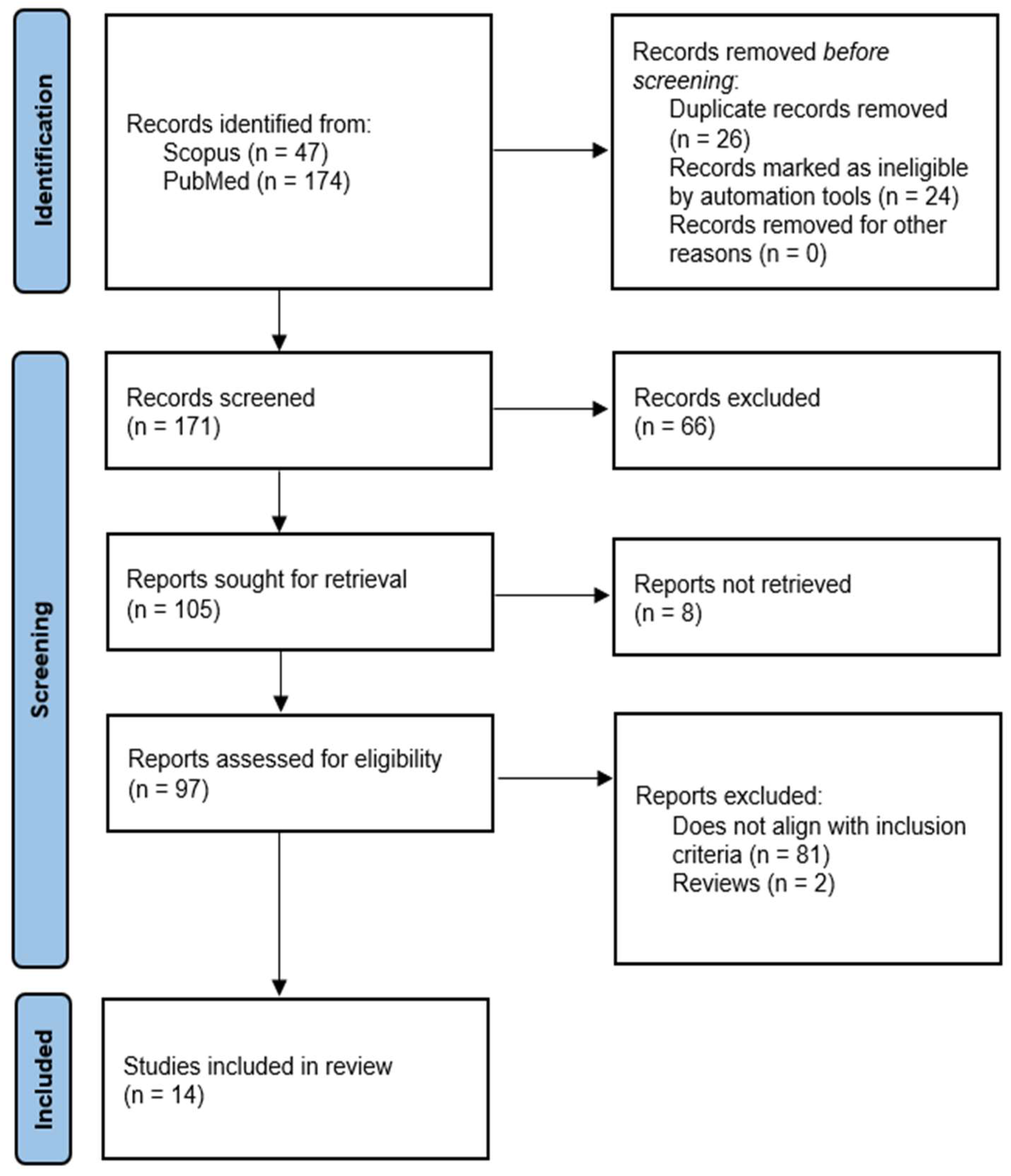
3. Results
3.1. Overview of Search Results
3.2. Summary of Findings
3.2.1. Loss of Heterozygosity (LOH) in THRB Gene Loci
3.2.2. Methylated CpG Site of Thyrotropin-Releasing Hormone Gene Sequence
3.2.3. THRA in Tongue Squamous Cell Carcinoma (TSCC) Progression
3.2.4. TSH and Antithyroid Antibody Expression in OLP Patients
3.2.5. Effects of HPT Axis Components on Oral Cell Lines
3.2.6. TSH Levels and Association with Disease Outcome in Head and Neck Squamous Cell Carcinoma (HNSCC)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zoeller, R.T.; Tan, S.W.; Tyl, R.W. General background on the hypothalamic-pituitary-thyroid (HPT) axis. Crit. Rev. Toxicol. 2007, 37, 11–53. [Google Scholar] [CrossRef] [PubMed]
- Mariotti, S.; Beck-Peccoz, P. Physiology of the hypothalamic-pituitary-thyroid axis. In Endotext [Internet]; MDText.com, Inc.: South Dartmouth, MA, USA, 2021. [Google Scholar]
- Slominski, A.T.; Zmijewski, M.A.; Skobowiat, C.; Zbytek, B.; Slominski, R.M.; Steketee, J.D. Sensing the environment: Regulation of local and global homeostasis by the skin’s neuroendocrine system. Adv. Anat. Embryol. Cell. Biol. 2012, 212, 1–115. [Google Scholar]
- Slominski, A.; Pisarchik, A.; Wortsman, J.; Kohn, L.; Ain, K.B.; Venkataraman, G.M.; Chung, J.H.; Giuliani, C.; Thornton, M.; Slugocki, G.; et al. Expression of Hypothalamic–Pituitary–Thyroid Axis Related Genes in the Human Skin. J. Investig. Dermatol. 2002, 119, 1449–1455. [Google Scholar] [CrossRef] [PubMed]
- Cianfarani, F.; Baldini, E.; Cavalli, A.; Marchioni, E.; Lembo, L.; Teson, M.; Persechino, S.; Zambruno, G.; Ulisse, S.; Odorisio, T.; et al. TSH receptor and thyroid-specific gene expression in human skin. J. Investig. Dermatol. 2010, 130, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Gáspár, E.; Hardenbicker, C.; Bodó, E.; Wenzel, B.; Ramot, Y.; Funk, W.; Kromminga, A.; Paus, R. Thyrotropin releasing hormone (TRH): A new player in human hair-growth control. FASEB J. 2010, 24, 393–403. [Google Scholar] [CrossRef]
- Bodó, E.; Kany, B.; Gáspár, E.; Knüver, J.; Kromminga, A.; Ramot, Y.; Bíro, T.; Tiede, S.; Van Beek, N.; Poeggeler, B.; et al. Thyroid-stimulating hormone, a novel, locally produced modulator of human epidermal functions, is regulated by thyrotropin-releasing hormone and thyroid hormones. Endocrinology 2010, 151, 1633–1642. [Google Scholar] [CrossRef]
- Contreras-Jurado, C.; García-Serrano, L.; Gómez-Ferrería, M.; Costa, C.; Paramio, J.M.; Aranda, A. The thyroid hormone receptors as modulators of skin proliferation and inflammation. J. Biol. Chem. 2011, 286, 24079–24088. [Google Scholar] [CrossRef]
- Van Beek, N.; Bodo, E.; Kromminga, A.; Gáspár, E.; Meyer, K.; Zmijewski, M.A.; Slominski, A.; Wenzel, B.E.; Paus, R. Thyroid hormones directly alter human hair follicle functions: Anagen prolongation and stimulation of both hair matrix keratinocyte proliferation and hair pigmentation. J. Clin. Endocrinol. Metab. 2008, 93, 4381–4388. [Google Scholar] [CrossRef]
- Krashin, E.; Piekiełko-Witkowska, A.; Ellis, M.; Ashur-Fabian, O. Thyroid hormones and cancer: A comprehensive review of preclinical and clinical studies. Front. Endocrinol. 2019, 10, 59. [Google Scholar] [CrossRef]
- Moeller, L.C.; Führer, D. Thyroid hormone, thyroid hormone receptors, and cancer: A clinical perspective. Endocr. Relat. Cancer 2013, 20, R19–R29. [Google Scholar] [CrossRef]
- Martínez-Iglesias, O.; Garcia-Silva, S.; Tenbaum, S.P.; Regadera, J.; Larcher, F.; Paramio, J.M.; Vennström, B.; Aranda, A. Thyroid hormone receptor β1 acts as a potent suppressor of tumor invasiveness and metastasis. Cancer Res. 2009, 69, 501–509. [Google Scholar] [CrossRef] [PubMed]
- Turabelidze, A.; Guo, S.; Chung, A.Y.; Chen, L.; Dai, Y.; Marucha, P.T.; DiPietro, L.A. Intrinsic differences between oral and skin keratinocytes. PLoS ONE 2014, 9, e101480. [Google Scholar] [CrossRef] [PubMed]
- Szpaderska, A.M.; Walsh, C.G.; Steinberg, M.J.; DiPietro, L.A. Distinct patterns of angiogenesis in oral and skin wounds. J. Dent. Res. 2005, 84, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Arbieva, Z.H.; Guo, S.; Marucha, P.T.; Mustoe, T.A.; DiPietro, L.A. Positional differences in the wound transcriptome of skin and oral mucosa. BMC Genom. 2010, 11, 471. [Google Scholar] [CrossRef] [PubMed]
- Cirillo, N. The Local Neuropeptide System of Keratinocytes. Biomedicines 2021, 9, 1854. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. Syst. Rev. 2021, 10, 89. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, Y.; Chin, Y.; Li, Z.; Shih, Y.; Yang, Y.; Ou, C.A.C.; Su, P.-Y.; Wang, S.-H.; Wu, Y.-H.; et al. Thyroid hormone-induced expression of inflammatory cytokines interfere with resveratrol-induced anti-proliferation of oral cancer cells. Food Chem. Toxicol. 2019, 132, 110693. [Google Scholar] [CrossRef]
- El-Naggar, A.; Lee, M.; Wang, G.; Luna, M.; Goepfert, H.; Batsakis, J. Polymerase chain reaction-based restriction fragment length polymorphism analysis of the short arm of chromosome 3 in primary head and neck squamous carcinoma. Cancer 1993, 72, 881–886. [Google Scholar] [CrossRef]
- Formanek, M.; Millesi, W.; Willheim, M.; Scheiner, O.; Kornfehl, J. Optimized growth medium for primary culture of human oral keratinocytes. Int. J. Oral Maxillofac. Surg. 1996, 25, 157–160. [Google Scholar] [CrossRef]
- Ho, Y.; Wu, C.; Chin, Y.; Li, Z.; Pan, Y.; Huang, T.; Su, P.-Y.; Lee, S.-Y.; Crawford, D.R.; Su, K.-W.; et al. NDAT suppresses pro-inflammatory gene expression to enhance resveratrol-induced anti-proliferation in oral cancer cells. Food Chem. Toxicol. 2020, 136, 111092. [Google Scholar] [CrossRef]
- Jank, B.; Haas, M.; Schnoell, J.; Schlederer, M.; Heiduschka, G.; Kenner, L.; Kadletz-Wanke, L. µ-Crystallin Is Associated with Disease Outcome in Head and Neck Squamous Cell Carcinoma. J. Pers. Med. 2021, 11, 1330. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Chin, Y.; Shih, Y.; Chen, Y.; Chung, Y.; Lin, C.; Hsiung, C.-N.; Whang-Peng, J.; Lee, S.-Y.; Lin, H.-Y.; et al. Resveratrol antagonizes thyroid hormone-induced expression of checkpoint and proliferative genes in oral cancer cells. J. Dent. Sci. 2019, 14, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Chin, Y.; Ho, Y.; Chou, S.; Sh Yang, Y.; Nana, A.; Su, K.-W.; Lim, Y.-T.; Wang, K.; Lee, S.-Y.; et al. Nano-diamino-tetrac (NDAT) inhibits PD-L1 expression which is essential for proliferation in oral cancer cells. Food Chem. Toxicol. 2018, 120, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Miyashita, H.; Mori, S.; Fukumoto, Y.; Sato, A.; Fukumoto, M.; Kawamura, H. Loss of heterozygosity of the PTH/PTHrP type 1 receptor in oral squamous cell carcinoma. Mol. Med. Rep. 2008, 1, 821–825. [Google Scholar] [CrossRef]
- Partridge, M.; Emilion, G.; Pateromichelakis, S.; A’Hern, R.; Lee, G.; Phillips, E.; Langdon, J. The prognostic significance of allelic imbalance at key chromosomal loci in oral cancer. Br. J. Cancer 1999, 79, 1821–1827. [Google Scholar] [CrossRef]
- Puttipanyalears, C.; Arayataweegool, A.; Chalertpet, K.; Rattanachayoto, P.; Mahattanasakul, P.; Tangjaturonsasme, N.; Kerekhanjanarong, V.; Mutirangura, A.; Kitkumthorn, N. TRH site-specific methylation in oral and oropharyngeal squamous cell carcinoma. BMC Cancer 2018, 18, 786. [Google Scholar] [CrossRef]
- Robledo-Sierra, J.; Landin-Wilhelmsen, K.; Filipsson Nyström, H.; Eggertsen, R.; Larsson, L.; Dafar, A.; Warfvinge, G.; Mattsson, U.; Jontell, M. A mechanistic linkage between oral lichen planus and autoimmune thyroid disease. Oral Dis. 2018, 24, 1001–1011. [Google Scholar] [CrossRef]
- Rowley, H.; Jones, A.; Spandidos, D.; Field, J. Definition of a Tumor Suppressor Gene Locus on the Short Arm of Chromosome 3 in Squamous Cell Carcinoma of the Head and Neck by Means of Microsatellite Markers. Arch. Otolaryngol.-Head Neck Surg. 1996, 122, 497–501. [Google Scholar] [CrossRef]
- Vehviläinen, M.; Salem, A.; Asghar, M.; Salo, T.; Siponen, M. No detection of TSH or TSHR in oral lichen planus lesions in patients with or without hypothyroidism. Acta Odontol. Scand. 2020, 78, 337–344. [Google Scholar] [CrossRef]
- Zhang, B.; Min, S.; Guo, Q.; Huang, Y.; Guo, Y.; Liang, X.; Wu, L.; Yu, G.; Wang, X. 7SK Acts as an Anti-tumor Factor in Tongue Squamous Cell Carcinoma. Front. Genet. 2021, 12, 642969. [Google Scholar] [CrossRef]
- Nelson, M.; Hercbergs, A.; Rybicki, L.; Strome, M. Association between development of hypothyroidism and improved survival in patients with head and neck cancer. Arch. Otolaryngol.-Head Neck Surg. 2006, 132, 1041–1046. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Reference | Type of Study | Population/Specimen | Intervention | Control | Outcome | Biomarkers | Assay Methodology |
|---|---|---|---|---|---|---|---|
| Chen et al. (2019) [18] | in vitro | Human (OEC-M1, SCC-25) | 10−7 M T4, resveratrol (40 μM or 10 μM) for 96 h or 24 h | Untreated cells | Proliferation and gene expression changes | IL-1b, CCND1, PD-L1 (CD274), COX-2, BAD, TGF-b1, STAT3, PI3K, ERK1/2 | Cell proliferation assay, qPCR, confocal microscopy, WB analysis |
| el-Naggar et al. (1993) [19] | clinical | Human (HNSCC tissue specimens) | NA | Normal mucosa from same patients | LOH | Restriction enzymes at the D3F1552, D3S32, THRB loci | PCR, flow cytometry, southern blotting |
| Formanek et al. (1996) [20] | in vitro | Human (primary cultures of oral cells) | Hydrocortisone, insulin, EGF, T3, bovine pituitary extract, transferrin, cholera toxin, adenine, KGM | DMEM (additive free control) | Proliferation | 3H-labeled thymidine | Proliferation assays using 3H-labeled thymidine incorporation. Flow cytometry |
| Ho et al. (2020) [21] | in vitro | Human (OEC-M1, SCC-25) | 10−7 M T4, 40 μM resveratrol, 10−7 NDAT, STAT3 inhibitor (S31-201, 40 μM) | Unstimulated cells | Gene expression changes, cell viability | IL-1B, TNF-a, BAD, CCND1, PD-L1, COX-2. | Cell viability test (alamar Blue Assay kit), qPCR, confocal microscopy |
| Jank et al. (2021) [22] | clinical | Human HNSCC patients | NA | NR | Clinical out-come, TSH level, CRYM | µ-Crystallin. Thyrotropin | Tissue Microarray/IHC |
| Lin et al. (2019) [23] | in vitro | Human (OEC-M1, SCC-25) | 10−7 M T4, 40 μM resveratrol and their combination for 24 h | Unstimulated cells | Gene expression, nuclear PD-L1 | BAD, CCND1, PD-L1, BTLA | qPCR, confocal microscopic analysis |
| Lin et al. (2018) [24] | in vitro | Human (OEC-M1, SCC-25) | 10−8 M to 10−6 M T4, 10−8 M to 10−6 M NDAT | Unstimulated cells | Expression/localization of PD-L1 | PD-L1, pPI3K | qRT-PCR, Western blotting |
| Miyashita et al. (2008) [25] | clinical | Human OSCC tissue | NA | Adjacent non-neoplastic tissue | LOH and MSI | PTHR1, THRB, p53, APC, BRCA1, BRCA2, DCC, FHIT | H&E staining, laser captured microdissection, PCR, electrophoresis |
| Patridge et al. (1999) [26] | clinical | Human OSCC, blood | NA | Normal samples | LOH, AI, FAL | Microsatellite markers (3p, 8p21-23, 9p13-21 9q22, 13q14.2) p53, Rb, DCC | Toluidine blue staining, microdissection, PCR-RFLP, gel electrophoresis |
| Puttipanyalears et al. (2018) [27] | clinical | Human oral SCC (tissue, swab, rinse) | NA | Healthy human oral tissue | Methylation of CpG | cg01009664 TRH (CpG methylated site) | Microdissection, pyrosequencing, RT-PCR |
| Robledo-Sierra et al. (2018) [28] | clinical | Human blood samples from OLP patients | NA | Healthy subjects | Antithyroid antibodies and OLP (association) | Circulating TgAb, TPOAb, FT3, FT4 Abs; thyroid proteins (Tg, TPO, TSHR) | IHC and qPCR analyses |
| Rowley (1996) [29] | clinical | Human tumors of the head and neck | NA | Normal tissue samples from same patients | LOH, microsatellite instability | Microsatellite markers D3S1304D3S656, D3S1252D3S1293, THRB, and D3S1266. | PCR |
| Vehviläinen et al. (2020) [30] | clinical | Human oral mucosal tissue (OLP) | NA | Healthy controls without oral lesions | Measurement of TSH and TSHR expression | TSH and TSHR | qRT-PCR, ddPCR, IHC, H&E staining |
| Zhang et al. (2021) [31] | in vitro | Human (SCC15) | Transfection of cells by ShRNAs targeting 7SK | PLKO negative control | Expression of 7SK, cell proliferation and migration, apoptosis | NR | RT-qPCR, RNA sequencing, cell migration assay, cell proliferation assay, cytometry |
| in vivo | BALB/c nude mice | Subcutaneous injection of 7SK-/- SCC15 cells at the armpits | Subcutaneous injection of control cells | Identification of enriched genes | NR | H&E staining, ChIRP-Seq analysis |
| Author (Year) | Human Carcinoma Sample | Chromosomal Location | LOH/Informative Sample (%) |
|---|---|---|---|
| el-Naggar et al. (1993) [19] | HNSCC | 3p24 | 3/9 (33%) |
| Miyashita et al. (2008) [25] | OSCC | 3p24.1-22 | 2/7 (28.6%) |
| Patridge et al. (1999) [26] | OSCC | 3p24-26 | 10/36 (28%) |
| Rowley et al. (1996) [29] | Head and neck tumours | 3p24 | 2/11 (18%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, L.; Xu, S.; Yang, B.; Yang, J.; Yee, C.; Cirillo, N. The Hypothalamic–Pituitary–Thyroid Axis Equivalent in Normal and Cancerous Oral Tissues: A Scoping Review. Int. J. Mol. Sci. 2022, 23, 14096. https://doi.org/10.3390/ijms232214096
Wu L, Xu S, Yang B, Yang J, Yee C, Cirillo N. The Hypothalamic–Pituitary–Thyroid Axis Equivalent in Normal and Cancerous Oral Tissues: A Scoping Review. International Journal of Molecular Sciences. 2022; 23(22):14096. https://doi.org/10.3390/ijms232214096
Chicago/Turabian StyleWu, Lisa, Stephen Xu, Brian Yang, Jenny Yang, Claire Yee, and Nicola Cirillo. 2022. "The Hypothalamic–Pituitary–Thyroid Axis Equivalent in Normal and Cancerous Oral Tissues: A Scoping Review" International Journal of Molecular Sciences 23, no. 22: 14096. https://doi.org/10.3390/ijms232214096
APA StyleWu, L., Xu, S., Yang, B., Yang, J., Yee, C., & Cirillo, N. (2022). The Hypothalamic–Pituitary–Thyroid Axis Equivalent in Normal and Cancerous Oral Tissues: A Scoping Review. International Journal of Molecular Sciences, 23(22), 14096. https://doi.org/10.3390/ijms232214096