
Immune Microenvironment and Immunotherapeutic Management in Virus-Associated Digestive System Tumors

 ,
,  ,
, 
 ,
,  and
and 
Abstract
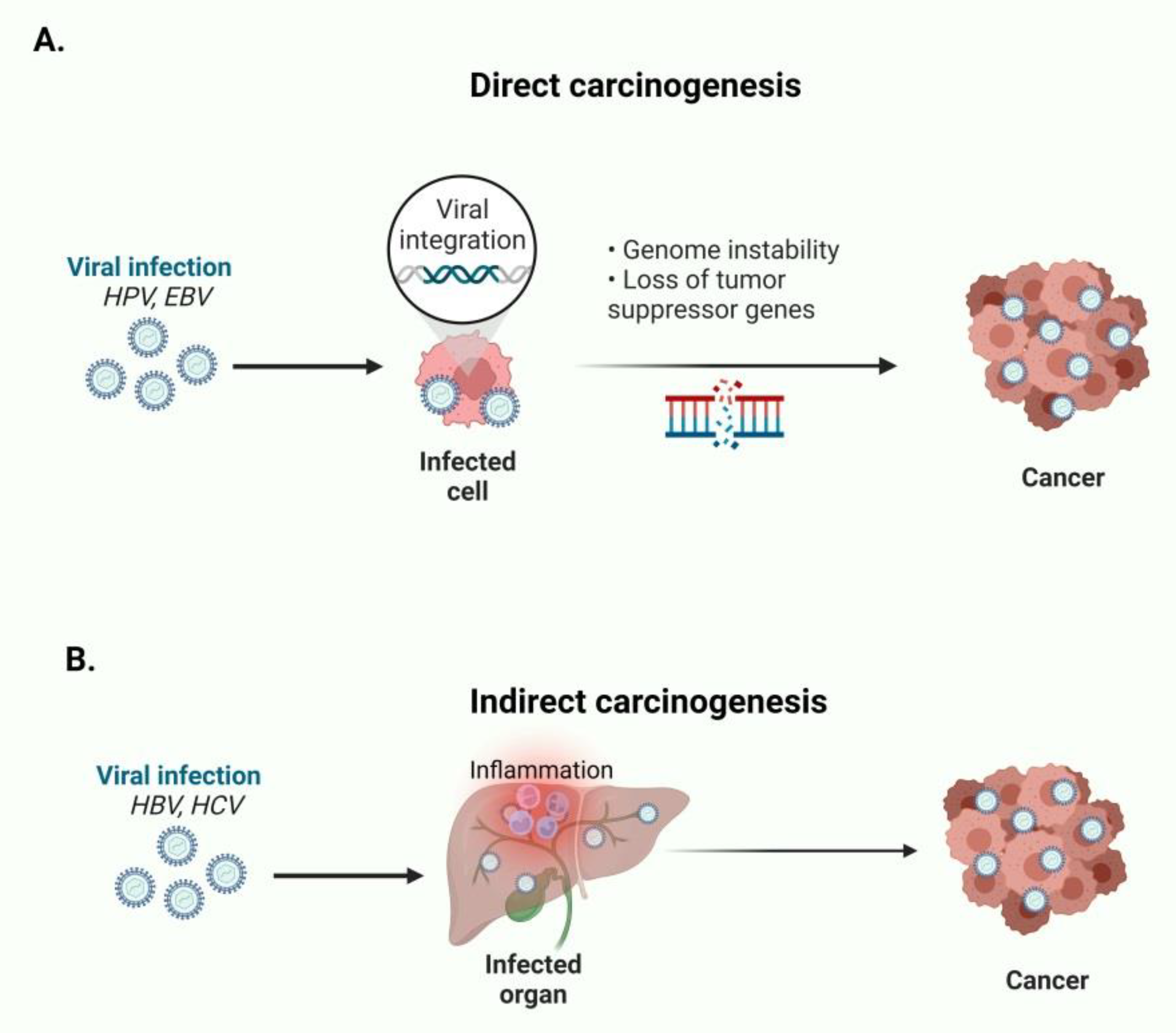
1. Introduction
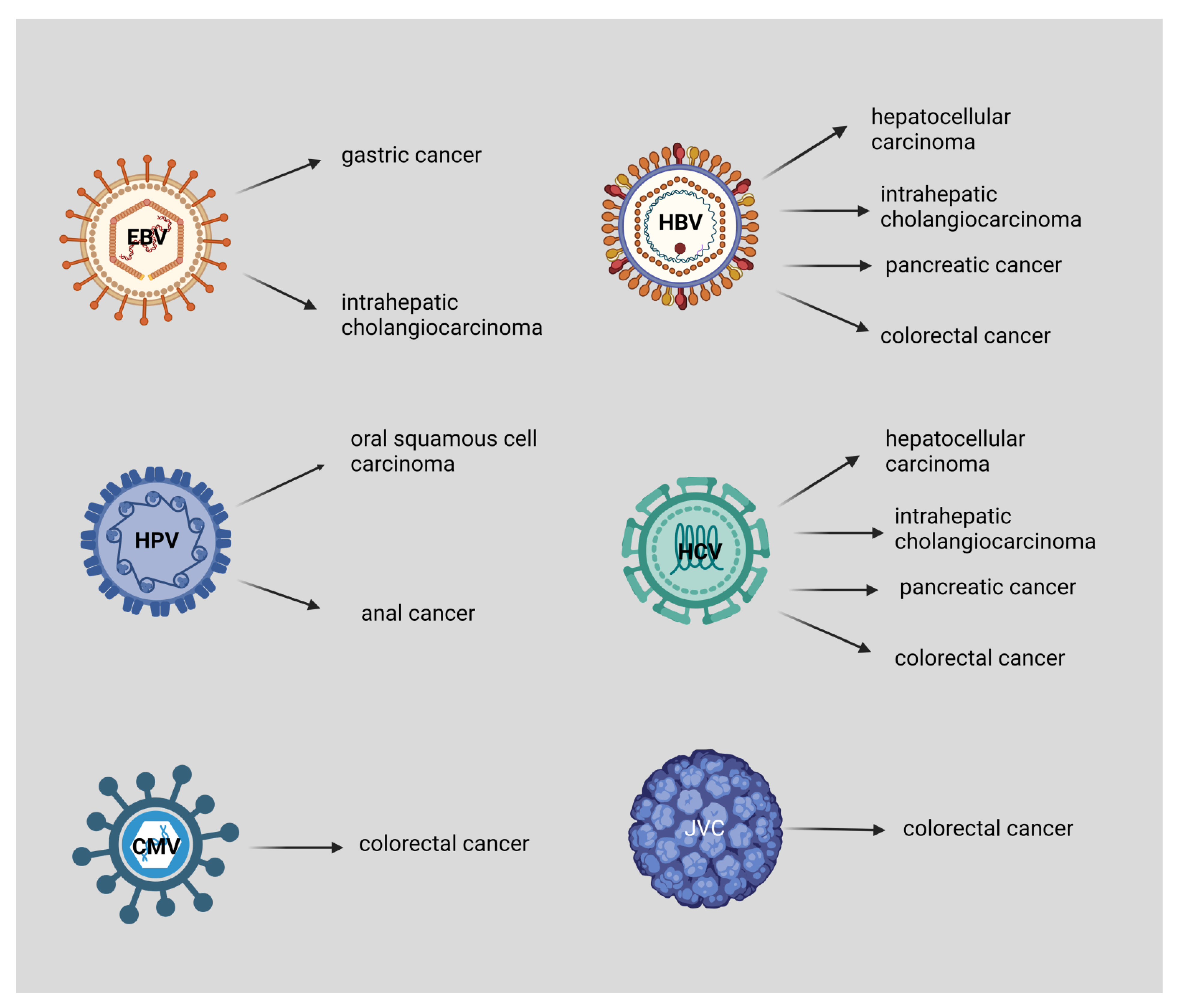
2. Hepatitis B Virus (HBV)
2.1. HBV-Associated Hepatocellular Carcinoma (HCC)
2.2. HBV-Associated Cholangiocarcinoma (CCA)
2.3. HΒV-Associated Colorectal Cancer (CRC)
2.4. HBV-Associated Pancreatic Adenocarcinoma (PAC)
3. Hepatitis C Virus (HCV)
3.1. HCV-Related HCC
3.2. HCV-Related Cholangiocarcinoma (CCA)
3.3. HCV-Related Pancreatic and CRC Cancers
4. John Cunningham Virus (JCV) and Human Cytomegalovirus (CMV)-Associated CRC
5. Epstein-Barr Virus (EBV)
5.1. EBV-Associated Gastric Cancer (GC)
5.2. EBV-Associated Intrahepatic Cholangiocarcinoma (iCCA)
6. Human Papilloma Virus (HPV)
6.1. HPV-Associated Oral Squamous Cell Carcinomas (OSCC)
6.2. HPV-Associated Anal Squamous Cell Carcinomas (ASCC)
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Grochowska, M.; Perlejewski, K.; Laskus, T.; Radkowski, M. The Role of Gut Microbiota in Gastrointestinal Tract Cancers. Arch. Immunol. Ther. Exp. 2022, 70, 7. [Google Scholar] [CrossRef] [PubMed]
- de Martel, C.; Ferlay, J.; Franceschi, S.; Vignat, J.; Bray, F.; Forman, D.; Plummer, M. Global Burden of Cancers Attributable to Infections in 2008: A Review and Synthetic Analysis. Lancet Oncol. 2012, 13, 607–615. [Google Scholar] [CrossRef]
- Morales-Sánchez, A.; Fuentes-Pananá, E.M. Human Viruses and Cancer. Viruses 2014, 6, 4047–4079. [Google Scholar] [CrossRef]
- Zárate, S.; Taboada, B.; Yocupicio-Monroy, M.; Arias, C.F. Human Virome. Arch. Med. Res. 2017, 48, 701–716. [Google Scholar] [CrossRef]
- Marônek, M.; Link, R.; Monteleone, G.; Gardlík, R.; Stolfi, C. Viruses in Cancers of the Digestive System: Active Contributors or Idle Bystanders? Int. J. Mol. Sci. 2020, 21, 8133. [Google Scholar] [CrossRef]
- Rai, K.R.; Shrestha, P.; Yang, B.; Chen, Y.; Liu, S.; Maarouf, M.; Chen, J.L. Acute Infection of Viral Pathogens and Their Innate Immune Escape. Front. Microbiol. 2021, 12, 1358. [Google Scholar] [CrossRef]
- Greten, F.R.; Grivennikov, S.I. Inflammation and Cancer: Triggers, Mechanisms, and Consequences. Immunity 2019, 51, 27–41. [Google Scholar] [CrossRef]
- Zhang, R.; Shen, C.; Zhao, L.; Wang, J.; McCrae, M.; Chen, X.; Lu, F. Dysregulation of Host Cellular Genes Targeted by Human Papillomavirus (HPV) Integration Contributes to HPV-Related Cervical Carcinogenesis. Int. J. Cancer 2016, 138, 1163–1174. [Google Scholar] [CrossRef]
- McLaughlin-Drubin, M.E.; Munger, K. Viruses Associated with Human Cancer. Biochim. Biophys. Acta 2008, 1782, 127–150. [Google Scholar] [CrossRef]
- Varn, F.S.; Schaafsma, E.; Wang, Y.; Cheng, C. Genomic Characterization of Six Virus-Associated Cancers Identifies Changes in the Tumor Immune Microenvironment and Altered Genetic Programs. Cancer Res. 2018, 78, 6413. [Google Scholar] [CrossRef] [PubMed]
- Marongiu, L.; Allgayer, H. Viruses in Colorectal Cancer. Mol. Oncol. 2022, 16, 1423–1450. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.J.; You, S.L.; Hsu, W.L.; Yang, H.I.; Lee, M.H.; Chen, H.C.; Chen, Y.Y.; Liu, J.; Hu, H.H.; Lin, Y.J.; et al. Epidemiology of Virus Infection and Human Cancer. Recent Results Cancer Res. 2021, 217, 13–45. [Google Scholar] [CrossRef] [PubMed]
- Bokas, A.; Papakotoulas, P.; Sarantis, P.; Papadimitropoulou, A.; Papavassiliou, A.G.; Karamouzis, M.v. Mechanisms of the Antitumor Activity of Low Molecular Weight Heparins in Pancreatic Adenocarcinomas. Cancers 2020, 12, 432. [Google Scholar] [CrossRef] [PubMed]
- Hao, X.; Sun, G.; Zhang, Y.; Kong, X.; Rong, D.; Song, J.; Tang, W.; Wang, X. Targeting Immune Cells in the Tumor Microenvironment of HCC: New Opportunities and Challenges. Front. Cell Dev. Biol. 2021, 9, 3149. [Google Scholar] [CrossRef] [PubMed]
- Koustas, E.; Trifylli, E.M.; Sarantis, P.; Papadopoulos, N.; Karapedi, E.; Aloizos, G.; Damaskos, C.; Garmpis, N.; Garmpi, A.; Papavassiliou, K.A.; et al. Immunotherapy as a Therapeutic Strategy for Gastrointestinal Cancer-Current Treatment Options and Future Perspectives. Int. J. Mol. Sci. 2022, 23, 6664. [Google Scholar] [CrossRef] [PubMed]
- Kaur, S.P.; Talat, A.; Karimi-Sari, H.; Grees, A.; Chen, H.W.; Lau, D.T.Y.; Catana, A.M. Hepatocellular Carcinoma in Hepatitis B Virus-Infected Patients and the Role of Hepatitis B Surface Antigen (HBsAg). J. Clin. Med. 2022, 11, 1126. [Google Scholar] [CrossRef]
- Song, C.; Lv, J.; Liu, Y.; Chen, J.G.; Ge, Z.; Zhu, J.; Dai, J.; Du, L.B.; Yu, C.; Guo, Y.; et al. Associations Between Hepatitis B Virus Infection and Risk of All Cancer Types. JAMA Netw. Open 2019, 2, e195718. [Google Scholar] [CrossRef]
- Liang, T.J. Hepatitis B: The Virus and Disease. Hepatology 2009, 49, S13–S21. [Google Scholar] [CrossRef]
- Kamiza, A.B.; Su, F.H.; Wang, W.C.; Sung, F.C.; Chang, S.N.; Yeh, C.C. Chronic Hepatitis Infection Is Associated with Extrahepatic Cancer Development: A Nationwide Population-Based Study in Taiwan. BMC Cancer 2016, 16, 861. [Google Scholar] [CrossRef]
- Ivanov, A.V.; Valuev-Elliston, V.T.; Tyurina, D.A.; Ivanova, O.N.; Kochetkov, S.N.; Bartosch, B.; Isaguliants, M.G. Oxidative Stress, a Trigger of Hepatitis C and B Virus-Induced Liver Carcinogenesis. Oncotarget 2017, 8, 3895–3932. [Google Scholar] [CrossRef] [PubMed]
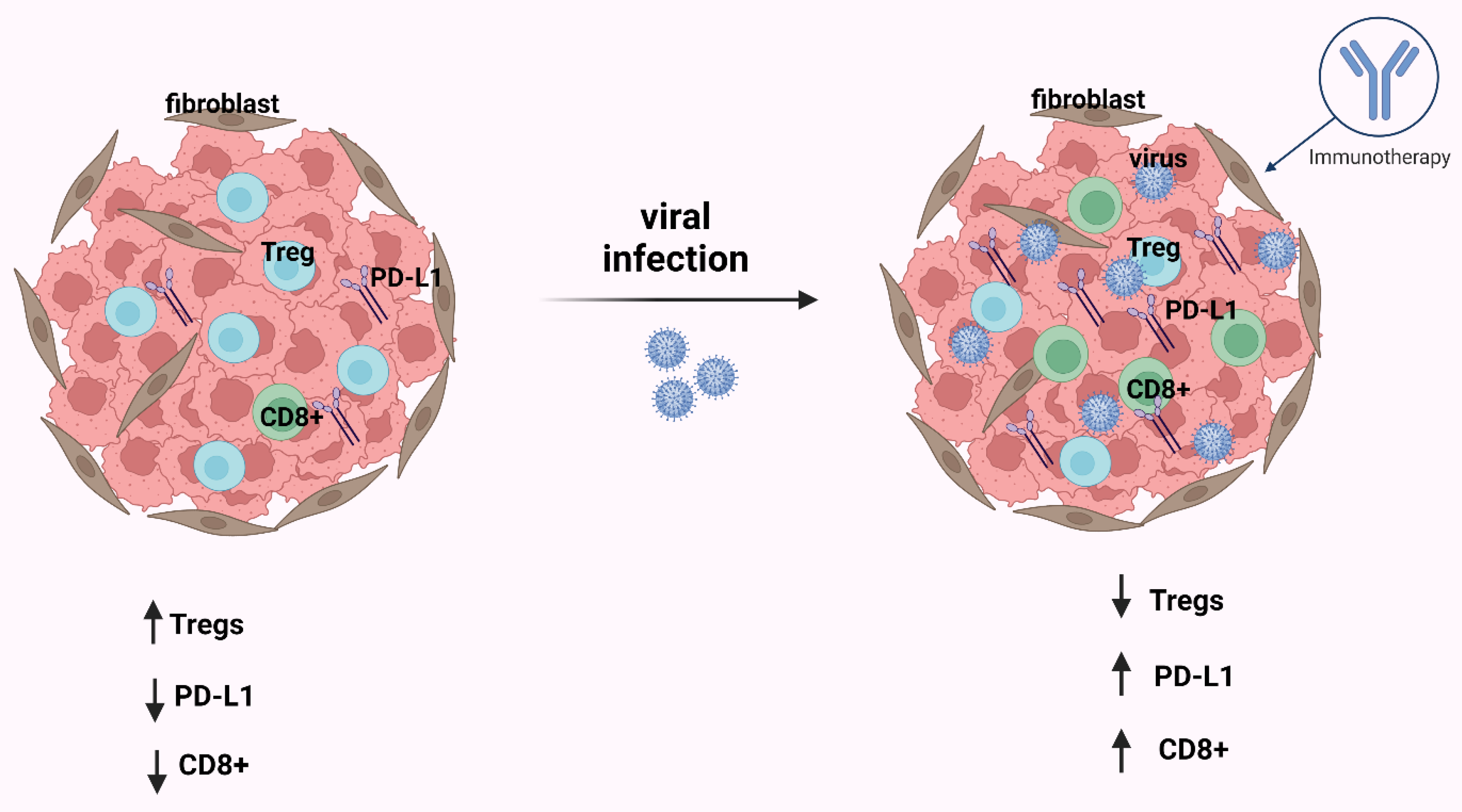
- Labani-Motlagh, A.; Ashja-Mahdavi, M.; Loskog, A. The Tumor Microenvironment: A Milieu Hindering and Obstructing Antitumor Immune Responses. Front. Immunol. 2020, 11, 940. [Google Scholar] [CrossRef] [PubMed]
- Teng, C.F.; Li, T.C.; Wang, T.; Liao, D.C.; Wen, Y.H.; Wu, T.H.; Wang, J.; Wu, H.C.; Shyu, W.C.; Su, I.J.; et al. Increased Infiltration of Regulatory T Cells in Hepatocellular Carcinoma of Patients with Hepatitis B Virus Pre-S2 Mutant. Sci. Rep. 2021, 11, 1136. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Sun, H.; Zhang, C.; Tian, Z. NK Cell Receptor Imbalance and NK Cell Dysfunction in HBV Infection and Hepatocellular Carcinoma. Cell Mol. Immunol. 2015, 12, 292–302. [Google Scholar] [CrossRef]
- Taghiabadi, M.; Hosseini, S.Y.; Gorzin, A.A.; Taghavi, S.A.; Monavari, S.H.R.; Sarvari, J. Comparison of Pre-S1/S2 Variations of Hepatitis B Virus between Asymptomatic Carriers and Cirrhotic/Hepatocellular Carcinoma-Affected Individuals. Clin. Exp. Hepatol. 2019, 5, 161–168. [Google Scholar] [CrossRef]
- Lim, C.J.; Lee, Y.H.; Pan, L.; Lai, L.; Chua, C.; Wasser, M.; Lim, T.K.H.; Yeong, J.; Toh, H.C.; Lee, S.Y.; et al. Multidimensional Analyses Reveal Distinct Immune Microenvironment in Hepatitis B Virus-Related Hepatocellular Carcinoma. Gut 2019, 68, 916–927. [Google Scholar] [CrossRef]
- de Battista, D.; Zamboni, F.; Gerstein, H.; Sato, S.; Markowitz, T.E.; Lack, J.; Engle, R.E.; Farci, P. Molecular Signature and Immune Landscape of HCV-Associated Hepatocellular Carcinoma (HCC): Differences and Similarities with HBV-HCC. J. Hepatocell. Carcinoma 2021, 8, 1399–1413. [Google Scholar] [CrossRef]
- Torresi, J.; Tran, B.M.; Christiansen, D.; Earnest-Silveira, L.; Schwab, R.H.M.; Vincan, E. HBV-Related Hepatocarcinogenesis: The Role of Signalling Pathways and Innovative Ex Vivo Research Models. BMC Cancer 2019, 19, 707. [Google Scholar] [CrossRef]
- Sangro, B.; Gomez-Martin, C.; de La Mata, M.; Iñarrairaegui, M.; Garralda, E.; Barrera, P.; Riezu-Boj, J.I.; Larrea, E.; Alfaro, C.; Sarobe, P.; et al. A Clinical Trial of CTLA-4 Blockade with Tremelimumab in Patients with Hepatocellular Carcinoma and Chronic Hepatitis C. J. Hepatol. 2013, 59, 81–88. [Google Scholar] [CrossRef]
- Duffy, A.G.; Ulahannan, S.V.; Makorova-Rusher, O.; Rahma, O.; Wedemeyer, H.; Pratt, D.; Davis, J.L.; Hughes, M.S.; Heller, T.; ElGindi, M.; et al. Tremelimumab in Combination with Ablation in Patients with Advanced Hepatocellular Carcinoma. J. Hepatol. 2017, 66, 545–551. [Google Scholar] [CrossRef]
- Kudo, M.; Finn, R.S.; Edeline, J.; Cattan, S.; Ogasawara, S.; Palmer, D.H.; Verslype, C.; Zagonel, V.; Fartoux, L.; Vogel, A.; et al. Updated Efficacy and Safety of KEYNOTE-224: A Phase II Study of Pembrolizumab in Patients with Advanced Hepatocellular Carcinoma Previously Treated with Sorafenib. Eur. J. Cancer 2022, 167, 1–12. [Google Scholar] [CrossRef]
- Ren, Z.; Xu, J.; Bai, Y.; Xu, A.; Cang, S.; Du, C.; Li, Q.; Lu, Y.; Chen, Y.; Guo, Y.; et al. Sintilimab plus a Bevacizumab Biosimilar (IBI305) versus Sorafenib in Unresectable Hepatocellular Carcinoma (ORIENT-32): A Randomised, Open-Label, Phase 2-3 Study. Lancet Oncol. 2021, 22, 977–990. [Google Scholar] [CrossRef]
- CIK Treatment for HCC Patient Underwent Radical Resection—Full Text View—ClinicalTrials.Gov. Available online: https://clinicaltrials.gov/ct2/show/NCT01749865 (accessed on 24 October 2022).
- Xu, L.; Wang, J.; Kim, Y.; Shuang, Z.Y.; Zhang, Y.J.; Lao, X.M.; Li, Y.Q.; Chen, M.S.; Pawlik, T.M.; Xia, J.C.; et al. A Randomized Controlled Trial on Patients with or without Adjuvant Autologous Cytokine-Induced Killer Cells after Curative Resection for Hepatocellular Carcinoma. Oncoimmunology 2016, 5, e1083671. [Google Scholar] [CrossRef] [PubMed]
- Trifylli, E.M.; Koustas, E.; Papadopoulos, N.; Sarantis, P.; Aloizos, G.; Damaskos, C.; Garmpis, N.; Garmpi, A.; Karamouzis, M.V. An Insight into the Novel Immunotherapy and Targeted Therapeutic Strategies for Hepatocellular Carcinoma and Cholangiocarcinoma. Life 2022, 12, 665. [Google Scholar] [CrossRef] [PubMed]
- Llovet, J.M.; Castet, F.; Heikenwalder, M.; Maini, M.K.; Mazzaferro, V.; Pinato, D.J.; Pikarsky, E.; Zhu, A.X.; Finn, R.S. Immunotherapies for Hepatocellular Carcinoma. Nat. Rev. Clin. Oncol. 2022, 19, 151–172. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Tian, Y.; Li, Y.; Zhang, W.; Cai, W.; Liu, Y.; Ren, Y.; Liang, Z.; Zhou, P.; Zhang, Y.; et al. In Vivo Therapeutic Effects of Affinity-Improved-TCR Engineered T-Cells on HBV-Related Hepatocellular Carcinoma. J. Immunother. Cancer 2020, 8, e001748. [Google Scholar] [CrossRef]
- Yoo, S.Y.; Badrinath, N.; Woo, H.Y.; Heo, J. Oncolytic Virus-Based Immunotherapies for Hepatocellular Carcinoma. Mediat. Inflamm. 2017, 2017, 5198798. [Google Scholar] [CrossRef]
- Heo, J.; Reid, T.; Ruo, L.; Breitbach, C.J.; Rose, S.; Bloomston, M.; Cho, M.; Lim, H.Y.; Chung, H.C.; Kim, C.W.; et al. Randomized Dose-Finding Clinical Trial of Oncolytic Immunotherapeutic Vaccinia JX-594 in Liver Cancer. Nat. Med. 2013, 19, 329–336. [Google Scholar] [CrossRef]
- Park, B.H.; Hwang, T.; Liu, T.C.; Sze, D.Y.; Kim, J.S.; Kwon, H.C.; Oh, S.Y.; Han, S.Y.; Yoon, J.H.; Hong, S.H.; et al. Use of a Targeted Oncolytic Poxvirus, JX-594, in Patients with Refractory Primary or Metastatic Liver Cancer: A Phase I Trial. Lancet Oncol. 2008, 9, 533–542. [Google Scholar] [CrossRef]
- Breitbach, C.J.; Burke, J.; Jonker, D.; Stephenson, J.; Haas, A.R.; Chow, L.Q.M.; Nieva, J.; Hwang, T.H.; Moon, A.; Patt, R.; et al. Intravenous Delivery of a Multi-Mechanistic Cancer-Targeted Oncolytic Poxvirus in Humans. Nature 2011, 477, 99–102. [Google Scholar] [CrossRef]
- Fragkou, N.; Sideras, L.; Panas, P.; Sinakos, E.; Emmanouilides, C. Update on the Association of Hepatitis B with Intrahepatic Cholangiocarcinoma: Is There New Evidence? World J. Gastroenterol. 2021, 27, 4252–4275. [Google Scholar] [CrossRef] [PubMed]
- Sarantis, P.; Tzanetatou, E.D.; Ioakeimidou, E.; Vallilas, C.; Androutsakos, T.; Damaskos, C.; Garmpis, N.; Garmpi, A.; Papavassiliou, A.G.; Karamouzis, M.V. Cholangiocarcinoma: The Role of Genetic and Epigenetic Factors; Current and Prospective Treatment with Checkpoint Inhibitors and Immunotherapy. Am. J. Transl. Res. 2021, 13, 13246. [Google Scholar] [PubMed]
- Thompson, A.I.; Conroy, K.P.; Henderson, N.C. Hepatic Stellate Cells: Central Modulators of Hepatic Carcinogenesis. BMC Gastroenterol. 2015, 15, 63. [Google Scholar] [CrossRef] [PubMed]
- Fabris, L.; Sato, K.; Alpini, G.; Strazzabosco, M. The Tumor Microenvironment in Cholangiocarcinoma Progression. Hepatology 2021, 73 (Suppl. 1), 75–85. [Google Scholar] [CrossRef]
- Koustas, E.; Sarantis, P.; Papavassiliou, A.G.; Karamouzis, M.v. The Resistance Mechanisms of Checkpoint Inhibitors in Solid Tumors. Biomolecules 2020, 10, 666. [Google Scholar] [CrossRef]
- An, J.; Kim, D.; Oh, B.; Oh, Y.J.; Song, J.; Park, N.; Kim, H.I.; Kang, H.J.; Oh, J.H.; Kim, W.; et al. Comprehensive Characterization of Viral Integrations and Genomic Aberrations in HBV-Infected Intrahepatic Cholangiocarcinomas. Hepatology 2022, 75, 997–1011. [Google Scholar] [CrossRef]
- Zhou, M.; Wang, C.; Lu, S.; Xu, Y.; Li, Z.; Jiang, H.; Ma, Y. Tumor-Associated Macrophages in Cholangiocarcinoma: Complex Interplay and Potential Therapeutic Target. EBioMedicine 2021, 67, 103375. [Google Scholar] [CrossRef]
- Deng, M.; Li, S.H.; Fu, X.; Yan, X.P.; Chen, J.; Qiu, Y.D.; Guo, R.P. Relationship between PD-L1 Expression, CD8+ T-Cell Infiltration and Prognosis in Intrahepatic Cholangiocarcinoma Patients. Cancer Cell Int. 2021, 21, 371. [Google Scholar] [CrossRef]
- Lin, J.; Dai, Y.; Sang, C.; Song, G.; Xiang, B.; Zhang, M.; Dong, L.; Xia, X.; Ma, J.; Shen, X.; et al. Multimodule Characterization of Immune Subgroups in Intrahepatic Cholangiocarcinoma Reveals Distinct Therapeutic Vulnerabilities. J. Immunother. Cancer 2022, 10, e004892. [Google Scholar] [CrossRef]
- Zhou, H.B.; Hu, J.Y.; Hu, H.P. Hepatitis B Virus Infection and Intrahepatic Cholangiocarcinoma. World J. Gastroenterol. 2014, 20, 5721–5729. [Google Scholar] [CrossRef]
- Song, G.; Shi, Y.; Zhang, M.; Goswami, S.; Afridi, S.; Meng, L.; Ma, J.; Chen, Y.; Lin, Y.; Zhang, J.; et al. Global Immune Characterization of HBV/HCV-Related Hepatocellular Carcinoma Identifies Macrophage and T-Cell Subsets Associated with Disease Progression. Cell Discov. 2020, 6, 2085. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.S.; Kim, N.H.; Park, J.H.; Park, D.I.; Sohn, C. il Correlation between Hepatitis B Virus Infection and Colorectal Neoplasia. J. Clin. Med. 2019, 8, 2085. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Li, W.; Zhang, Y.; Siyin, S.T.; Zhang, Q.; Song, M.; Zhang, K.; Liu, S.; Shi, H. Associations between Hepatitis B Virus Infection and Risk of Colorectal Cancer: A Population-Based Prospective Study. BMC Cancer 2021, 21, 1119. [Google Scholar] [CrossRef]
- Nam, S.; Choi, Y.J.; Kim, D.W.; Park, E.C.; Kang, J.G. Risk Factors for Colorectal Cancer in Korea: A Population-Based Retrospective Cohort Study. Ann. Coloproctol. 2019, 35, 347. [Google Scholar] [CrossRef]
- Zhou, J.; Guo, X.; Huang, P.; Tan, S.; Lin, R.; Zhan, H.; Wu, X.; Li, T.; Huang, M.; Huang, M. HBV Infection Status Indicates Different Risks of Synchronous and Metachronous Liver Metastasis in Colorectal Cancer: A Retrospective Study of 3132 Patients with a 5-Year Follow-Up. Cancer Manag. Res. 2022, 14, 1581–1594. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.K.; Chen, P.; Chen, D.W.; Lin, Z.S.; Ye, S.B.; Lan, P. Comparative Safety, Efficacy and Survival Outcome of Anti-PD-1 Immunotherapy in Colorectal Cancer Patients With vs Without Hepatitis B Virus Infection: A Multicenter Cohort Study. Clin. Transl. Gastroenterol. 2022, 13, e00475. [Google Scholar] [CrossRef] [PubMed]
- Sarantis, P.; Bokas, A.; Papadimitropoulou, A.; Koustas, E.; Theocharis, S.; Papakotoulas, P.; Schizas, D.; Papalampros, A.; Felekouras, E.; Papavassiliou, A.G.; et al. Combinatorial Treatment of Tinzaparin and Chemotherapy Can Induce a Significant Antitumor Effect in Pancreatic Cancer. Int. J. Mol. Sci. 2021, 22, 7053. [Google Scholar] [CrossRef] [PubMed]
- Maisonneuve, P.; Lowenfels, A.B. Risk Factors for Pancreatic Cancer: A Summary Review of Meta-Analytical Studies. Int. J. Epidemiol. 2015, 44, 186–198. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, Z.H.; Jiang, F. Hepatitis B Virus Infection Increases the Risk of Pancreatic Cancer: A Meta-Analysis. Scand J. Gastroenterol. 2021, 56, 252–258. [Google Scholar] [CrossRef]
- Olakowski, M.; Bułdak, Ł. Modifiable and Non-Modifiable Risk Factors for the Development of Non-Hereditary Pancreatic Cancer. Medicina 2022, 58, 978. [Google Scholar] [CrossRef]
- Sherman, M. Pancreatic Cancer in Chronic Hepatitis B. Liver Int. 2010, 30, 339–341. [Google Scholar] [CrossRef]
- Dumitrascu, T.; Pineau, P. Is Hepatitis B Virus a Player in Pancreatic Cancer? Chirurgia 2018, 113, 344–352. [Google Scholar] [CrossRef] [PubMed]
- Shi, G.; Suzuki, T. Molecular Basis of Encapsidation of Hepatitis C Virus Genome. Front. Microbiol. 2018, 9, 396. [Google Scholar] [CrossRef] [PubMed]
- Li, H.C.; Lo, S.Y. Hepatitis C Virus: Virology, Diagnosis and Treatment. World J. Hepatol. 2015, 7, 1377–1389. [Google Scholar] [CrossRef] [PubMed]
- Flores, J.E.; Thompson, A.J.; Ryan, M.; Howell, J. The Global Impact of Hepatitis B Vaccination on Hepatocellular Carcinoma. Vaccines 2022, 10, 793. [Google Scholar] [CrossRef]
- Modi, A.A.; Liang, T.J. Hepatitis C: A Clinical Review. Oral. Dis. 2008, 14, 10–14. [Google Scholar] [CrossRef]
- Saud, L.R.C.; Chagas, A.L.; Maccali, C.; Pinto, P.V.A.; Horvat, N.; Alencar, R.S.S.M.; Tani, C.M.; Abdala, E.; Carrilho, F.J. Hepatocellular Carcinoma in Patients Co-Infected with Hepatitis B or C and HIV: More Aggressive Tumor Behavior? Eur. J. Gastroenterol. Hepatol. 2021, 33, 583. [Google Scholar] [CrossRef]
- Oh, J.K.; Shin, H.R.; Lim, M.K.; Cho, H.; Kim, D.I.; Jee, Y.; Yun, H.; Yoo, K.Y. Multiplicative Synergistic Risk of Hepatocellular Carcinoma Development among Hepatitis B and C Co-Infected Subjects in HBV Endemic Area: A Community-Based Cohort Study. BMC Cancer 2012, 12, 452. [Google Scholar] [CrossRef]
- Hasson, H.; Merli, M.; Messina, E.; Bhoori, S.; Salpietro, S.; Morsica, G.; Regalia, E.; Bagaglio, S.; Lazzarin, A.; Uberti-Foppa, C.; et al. Occurrence of Hepatocellular Carcinoma in HIV/HCV Co-Infected Patients Treated with Direct-Acting Antivirals. J. Hepatol. 2017, 67, 415–417. [Google Scholar] [CrossRef]
- Zhao, P.; Malik, S.; Xing, S. Epigenetic Mechanisms Involved in HCV-Induced Hepatocellular Carcinoma (HCC). Front. Oncol. 2021, 11, 677926. [Google Scholar] [CrossRef]
- Dhanasekaran, R.; Kwo, P.Y. Hepatitis C and Hepatocellular Cancer: To Treat or Not to Treat. Clin. Liver Dis. 2021, 17, 169–173. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.S.; Lin, C.L.; Lu, M.J.; Yeh, C.T.; Liang, K.H.; Sun, C.C.; Shyu, Y.C.; Chien, R.N. Diabetes, Hepatocellular Carcinoma, and Mortality in Hepatitis C-Infected Patients: A Population-Based Cohort Study. J. Gastroenterol. Hepatol. 2017, 32, 1355–1362. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Xu, H.; Gao, Y.; Pan, M.; Wang, L.; Gao, P. Diabetes Mellitus Increases the Risk of Hepatocellular Carcinoma in Treatment-Naïve Chronic Hepatitis C Patients in China. Medicine 2017, 96, e6508. [Google Scholar] [CrossRef] [PubMed]
- Irshad, M.; Gupta, P.; Irshad, K. Molecular Basis of Hepatocellular Carcinoma Induced by Hepatitis C Virus Infection. World J. Hepatol. 2017, 9, 1305. [Google Scholar] [CrossRef]
- Wang, H.; Wang, P.; Xu, M.; Song, X.; Wu, H.; Evert, M.; Calvisi, D.F.; Zeng, Y.; Chen, X. Distinct Functions of Transforming Growth Factor-β Signaling in c-MYC Driven Hepatocellular Carcinoma Initiation and Progression. Cell Death Dis. 2021, 12, 200. [Google Scholar] [CrossRef]
- Jin, X.; Zhang, S.; Wang, N.; Guan, L.; Shao, C.; Lin, Y.; Liu, J.; Li, Y. High Expression of TGF-Β1 Contributes to Hepatocellular Carcinoma Prognosis via Regulating Tumor Immunity. Front. Oncol. 2022, 12, 861601. [Google Scholar] [CrossRef]
- Banerjee, A.; Ray, R.B.; Ray, R. Oncogenic Potential of Hepatitis C Virus Proteins. Viruses 2010, 2, 2108–2133. [Google Scholar] [CrossRef]
- Virzì, A.; Suarez, A.A.R.; Baumert, T.F.; Lupberger, J. Oncogenic Signaling Induced by HCV Infection. Viruses 2018, 10, 538. [Google Scholar] [CrossRef]
- Mahmoudvand, S.; Shokri, S.; Taherkhani, R.; Farshadpour, F. Hepatitis C Virus Core Protein Modulates Several Signaling Pathways Involved in Hepatocellular Carcinoma. World J. Gastroenterol. 2019, 25, 42–58. [Google Scholar] [CrossRef]
- Xie, Y.; Sun, R.; Gao, L.; Guan, J.; Wang, J.; Bell, A.; Zhu, J.; Zhang, M.; Xu, M.; Lu, P.; et al. Chronic Activation of LXRα Sensitizes Mice to Hepatocellular Carcinoma. Hepatol. Commun. 2022, 6, 1123–1139. [Google Scholar] [CrossRef]
- Giraud, J.; Chalopin, D.; Blanc, J.F.; Saleh, M. Hepatocellular Carcinoma Immune Landscape and the Potential of Immunotherapies. Front. Immunol. 2021, 12, 655697. [Google Scholar] [CrossRef] [PubMed]
- Goto, K.; Andres Roca Suarez, A.; Wrensch, F.; Baumert, T.F.; Lupberger, J. Hepatitis C Virus and Hepatocellular Carcinoma: When the Host Loses Its Grip. Int. J. Mol. Sci. 2020, 21, 3057. [Google Scholar] [CrossRef] [PubMed]
- Dash, S.; Aydin, Y.; Widmer, K.E.; Nayak, L. Hepatocellular Carcinoma Mechanisms Associated with Chronic HCV Infection and the Impact of Direct-Acting Antiviral Treatment. J. Hepatocell. Carcinoma 2020, 7, 45–76. [Google Scholar] [CrossRef] [PubMed]
- Oe, N.; Takeda, H.; Eso, Y.; Takai, A.; Marusawa, H. Clinical and Molecular Basis of Hepatocellular Carcinoma after Hepatitis C Virus Eradication. Pathogens 2022, 11, 430. [Google Scholar] [CrossRef]
- Luxenburger, H.; Neumann-Haefelin, C.; Thimme, R.; Boettler, T. HCV-Specific T Cell Responses During and After Chronic HCV Infection. Viruses 2018, 10, 645. [Google Scholar] [CrossRef]
- Park, S.-J.; Hahn, Y.S. Hepatocytes Infected with Hepatitis C Virus Change Immunological Features in the Liver Microenvironment. Clin. Mol. Hepatol. 2022. [Google Scholar] [CrossRef]
- Ravi, R.; Noonan, K.A.; Pham, V.; Bedi, R.; Zhavoronkov, A.; Ozerov, I.V.; Makarev, E.; Artemov, A.v.; Wysocki, P.T.; Mehra, R.; et al. Bifunctional Immune Checkpoint-Targeted Antibody-Ligand Traps That Simultaneously Disable TGFβ Enhance the Efficacy of Cancer Immunotherapy. Nat. Commun. 2018, 9, 741. [Google Scholar] [CrossRef]
- Ren, Z.; Qin, S.; Meng, Z.; Chen, Z.; Chai, X.; Xiong, J.; Bai, Y.; Yang, L.; Zhu, H.; Fang, W.; et al. A Phase 2 Study of Camrelizumab for Advanced Hepatocellular Carcinoma: Two-Year Outcomes and Continued Treatment beyond First RECIST-Defined Progression. Liver Cancer 2021, 10, 500–509. [Google Scholar] [CrossRef]
- Qin, S.; Finn, R.S.; Kudo, M.; Meyer, T.; Vogel, A.; Ducreux, M.; Macarulla, T.M.; Tomasello, G.; Boisserie, F.; Hou, J.; et al. RATIONALE 301 Study: Tislelizumab versus Sorafenib as First-Line Treatment for Unresectable Hepatocellular Carcinoma. Future Oncol. 2019, 15, 1811–1822. [Google Scholar] [CrossRef]
- Desai, J.; Deva, S.; Lee, J.S.; Lin, C.C.; Yen, C.J.; Chao, Y.; Keam, B.; Jameson, M.; Hou, M.M.; Kang, Y.K.; et al. Phase IA/IB Study of Single-Agent Tislelizumab, an Investigational Anti-PD-1 Antibody, in Solid Tumors. J. Immunother Cancer 2020, 8, e000453. [Google Scholar] [CrossRef]
- Tada, F.; Abe, M.; Hirooka, M.; Ikeda, Y.; Hiasa, Y.; Lee, Y.; Jung, N.C.; Lee, W.B.; Lee, H.S.; Bae, Y.S.; et al. Phase I/II Study of Immunotherapy Using Tumor Antigen-Pulsed Dendritic Cells in Patients with Hepatocellular Carcinoma. Int. J. Oncol. 2012, 41, 1601–1609. [Google Scholar] [CrossRef] [PubMed]
- Repáraz, D.; Aparicio, B.; Llopiz, D.; Hervás-stubbs, S.; Sarobe, P. Therapeutic Vaccines against Hepatocellular Carcinoma in the Immune Checkpoint Inhibitor Era: Time for Neoantigens? Int. J. Mol. Sci. 2022, 23, 2022. [Google Scholar] [CrossRef] [PubMed]
- Tsuchiya, N.; Yoshikawa, T.; Fujinami, N.; Saito, K.; Mizuno, S.; Sawada, Y.; Endo, I.; Nakatsura, T. Immunological Efficacy of Glypican-3 Peptide Vaccine in Patients with Advanced Hepatocellular Carcinoma. Oncoimmunology 2017, 6, e1346764. [Google Scholar] [CrossRef] [PubMed]
- Banales, J.M.; Marin, J.J.G.; Lamarca, A.; Rodrigues, P.M.; Khan, S.A.; Roberts, L.R.; Cardinale, V.; Carpino, G.; Andersen, J.B.; Braconi, C.; et al. Cholangiocarcinoma 2020: The next Horizon in Mechanisms and Management. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 557–588. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Hu, B.; Zhou, Z.Q.; Guan, J.; Zhang, Z.Y.; Zhou, G.W. Hepatitis C Virus Infection and the Risk of Intrahepatic Cholangiocarcinoma and Extrahepatic Cholangiocarcinoma: Evidence from a Systematic Review and Meta-Analysis of 16 Case-Control Studies. World J. Surg. Oncol. 2015, 13, 161. [Google Scholar] [CrossRef]
- Navas, M.C.; Glaser, S.; Dhruv, H.; Celinski, S.; Alpini, G.; Meng, F. Hepatitis C Virus Infection and Cholangiocarcinoma: An Insight into Epidemiologic Evidences and Hypothetical Mechanisms of Oncogenesis. Am. J. Pathol. 2019, 189, 1122–1132. [Google Scholar] [CrossRef]
- Seo, J.W.; Kwan, B.S.; Cheon, Y.K.; Lee, T.Y.; Shim, C.S.; Kwon, S.Y.; Choe, W.H.; Yoo, B.C.; Yoon, J.M.; Lee, J.H. Prognostic Impact of Hepatitis B or C on Intrahepatic Cholangiocarcinoma. Korean J. Intern. Med. 2020, 35, 566–573. [Google Scholar] [CrossRef]
- Li, T.; Li, D.; Cheng, L.; Wu, H.; Gao, Z.; Liu, Z.; Jiang, W.; Gao, Y.H.; Tian, F.; Zhao, L.; et al. Epithelial-Mesenchymal Transition Induced by Hepatitis C Virus Core Protein in Cholangiocarcinoma. Ann. Surg. Oncol. 2010, 17, 1937–1944. [Google Scholar] [CrossRef]
- Su, F.H.; Bai, C.H.; Le, T.N.; Muo, C.H.; Chang, S.N.; Te, A.; Sung, F.C.; Yeh, C.C. Patients With Chronic Hepatitis C Virus Infection Are at an Increased Risk of Colorectal Cancer: A Nationwide Population-Based Case-Control Study in Taiwan. Front. Oncol. 2021, 10, 561420. [Google Scholar] [CrossRef]
- Darvishian, M.; Butt, Z.A.; Wong, S.; Yoshida, E.M.; Khinda, J.; Otterstatter, M.; Yu, A.; Binka, M.; Rossi, C.; McKee, G.; et al. Elevated Risk of Colorectal, Liver, and Pancreatic Cancers among HCV, HBV and/or HIV (Co)Infected Individuals in a Population Based Cohort in Canada. Ther. Adv. Med. Oncol. 2021, 13, 1758835921992987. [Google Scholar] [CrossRef]
- Delbue, S.; Comar, M.; Ferrante, P. Review on the Role of the Human Polyomavirus JC in the Development of Tumors. Infect. Agent Cancer 2017, 12, 10. [Google Scholar] [CrossRef] [PubMed]
- Goel, A.; Li, M.S.; Nagasaka, T.; Shin, S.K.; Fuerst, F.; Ricciardiello, L.; Wasserman, L.; Boland, C.R. Association of JC Virus T-Antigen Expression With the Methylator Phenotype in Sporadic Colorectal Cancers. Gastroenterology 2006, 130, 1950–1961. [Google Scholar] [CrossRef] [PubMed]
- Navand, A.H.; Teimoori, A.; Makvandi, M.; Nisi, N.; Seyedian, S.S.; Ranjbari, N.; Angali, K.A.; Keyani, H.; Tabasi, M.; Pourjabari, K. Study on JV Virus in Patients with Colon Cancer Type Adenocarcinoma. Asian Pac J. Cancer Prev. 2019, 20, 1147–1151. [Google Scholar] [CrossRef] [PubMed]
- Butcher, L.D.; Garcia, M.; Arnold, M.; Ueno, H.; Goel, A.; Boland, C.R. Immune Response to JC Virus T Antigen in Patients with and without Colorectal Neoplasia. Gut Microbes 2014, 5, 468–475. [Google Scholar] [CrossRef] [PubMed]
- Kimla, L.J. John Cunningham Virus Infection and the Risk of Colorectal Cancer: A Meta-Analysis of Case-Control Studies. J. Clin. Oncol. 2018, 36, e15650. [Google Scholar] [CrossRef]
- Holtappels, R.; Freitag, K.; Renzaho, A.; Becker, S.; Lemmermann, N.A.W.; Reddehase, M.J. Revisiting CD8 T-Cell “Memory Inflation”: New Insights with Implications for Cytomegaloviruses as Vaccine Vectors. Vaccines 2020, 8, 402. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.H.; Meng, Q.; Rao, M.; Liu, Z.; Paraschoudi, G.; Dodoo, E.; Maeurer, M. The Impact of Inflationary Cytomegalovirus-Specific Memory T Cells on Anti-Tumour Immune Responses in Patients with Cancer. Immunology 2018, 155, 294–308. [Google Scholar] [CrossRef] [PubMed]
- Schlick, K.; Grundbichler, M.; Auberger, J.; Kern, J.M.; Hell, M.; Hohla, F.; Hopfinger, G.; Greil, R. Cytomegalovirus Reactivation and Its Clinical Impact in Patients with Solid Tumors. Infect. Agent Cancer 2015, 10, 45. [Google Scholar] [CrossRef]
- van den Berg, S.P.H.; Pardieck, I.N.; Lanfermeijer, J.; Sauce, D.; Klenerman, P.; van Baarle, D.; Arens, R. The Hallmarks of CMV-Specific CD8 T-Cell Differentiation. Med. Microbiol. Immunol. 2019, 208, 365–373. [Google Scholar] [CrossRef]
- Gordon, C.L.; Miron, M.; Thome, J.J.C.; Matsuoka, N.; Weiner, J.; Rak, M.A.; Igarashi, S.; Granot, T.; Lerner, H.; Goodrum, F.; et al. Tissue Reservoirs of Antiviral T Cell Immunity in Persistent Human CMV Infection. J. Exp. Med. 2017, 214, 651–667. [Google Scholar] [CrossRef]
- Lee, M.S.; Selitsky, S.R.; Parker, J.S. Prognostic Impact of Cytomegalovirus (CMV) Detection in Colorectal Cancer (CRC). J. Clin. Oncol. 2017, 35, e15120. [Google Scholar] [CrossRef]
- Chen, H.P.; Chan, Y.J. The Oncomodulatory Role of Human Cytomegalovirus in Colorectal Cancer: Implications for Clinical Trials. Front. Oncol 2014, 4, 314. [Google Scholar] [CrossRef] [PubMed]
- Longnecker, R.; Neipel, F. Introduction to the Human γ-Herpesviruses. In Human Herpesviruses: Biology, Therapy, and Immunoprophylaxis; Cambridge University Press: Cambridge, UK, 2007; pp. 341–359. [Google Scholar] [CrossRef]
- Han, S.; Tay, J.K.; Loh, C.J.L.; Chu, A.J.M.; Yeong, J.P.S.; Lim, C.M.; Toh, H.C. Epstein–Barr Virus Epithelial Cancers—A Comprehensive Understanding to Drive Novel Therapies. Front. Immunol. 2021, 12, 5102. [Google Scholar] [CrossRef] [PubMed]
- Teng, M.W.L.; Ngiow, S.F.; Ribas, A.; Smyth, M.J. Classifying Cancers Based on T-Cell Infiltration and PD-L1. Cancer Res. 2015, 75, 2139–2145. [Google Scholar] [CrossRef]
- Iizasa, H.; Nanbo, A.; Nishikawa, J.; Jinushi, M.; Yoshiyama, H. Epstein-Barr Virus (EBV)-Associated Gastric Carcinoma. Viruses 2012, 4, 3420–3439. [Google Scholar] [CrossRef] [PubMed]
- Camargo, M.C.; Koriyama, C.; Matsuo, K.; Kim, W.H.; Herrera-Goepfert, R.; Liao, L.M.; Yu, J.; Carrasquilla, G.; Sung, J.J.Y.; Alvarado-Cabrero, I.; et al. Case-Case Comparison of Smoking and Alcohol Risk Associations with Epstein-Barr Virus-Positive Gastric Cancer. Int. J. Cancer 2014, 134, 948–953. [Google Scholar] [CrossRef]
- Chen, P.C.H.; Pan, C.C.; Hsu, W.H.; Ka, H.J.; Yang, A.H. Epstein-Barr Virus-Associated Lymphoepithelioma-like Carcinoma of the Esophagus. Hum. Pathol. 2003, 34, 407–410. [Google Scholar] [CrossRef]
- Nishikawa, J.; Yoshiyama, H.; Iizasa, H.; Kanehiro, Y.; Nakamura, M.; Nishimura, J.; Saito, M.; Okamoto, T.; Sakai, K.; Suehiro, Y.; et al. Epstein-Barr Virus in Gastric Carcinoma. Cancers 2014, 6, 2259. [Google Scholar] [CrossRef]
- Yanai, H.; Nishikawa, J.; Mizugaki, Y.; Shimizu, N.; Takada, K.; Matsusaki, K.; Toda, T.; Matsumoto, Y.; Tada, M.; Okita, K. Endoscopic and Pathologic Features of Epstein-Barr Virus-Associated Gastric Carcinoma. Gastrointest. Endosc. 1997, 45, 236–242. [Google Scholar] [CrossRef]
- Zhao, K.; Zhang, Y.; Xia, S.; Feng, L.; Zhou, W.; Zhang, M.; Dong, R.; Tian, D.; Liu, M.; Liao, J. Epstein-Barr Virus Is Associated with Gastric Cancer Precursor: Atrophic Gastritis. Int. J. Med. Sci. 2022, 19, 924–931. [Google Scholar] [CrossRef]
- Cheng, Y.; Zhou, X.; Xu, K.; Huang, J.; Huang, Q. Very Low Risk of Lymph Node Metastasis in Epstein-Barr Virus-Associated Early Gastric Carcinoma with Lymphoid Stroma. BMC Gastroenterol. 2020, 20, 273. [Google Scholar] [CrossRef] [PubMed]
- Sun, K.; Jia, K.; Lv, H.; Wang, S.Q.; Wu, Y.; Lei, H.; Chen, X. EBV-Positive Gastric Cancer: Current Knowledge and Future Perspectives. Front. Oncol. 2020, 10, 583463. [Google Scholar] [CrossRef] [PubMed]
- Rodriquenz, M.G.; Roviello, G.; D’angelo, A.; Lavacchi, D.; Roviello, F.; Polom, K. MSI and EBV Positive Gastric Cancer’s Subgroups and Their Link With Novel Immunotherapy. J. Clin. Med. 2020, 9, 1427. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Li, J.; Hao, Y.; Nie, Y.; Li, Z.; Qian, M.; Liang, Q.; Yu, J.; Zeng, M.; Wu, K. Differentiated Tumor Immune Microenvironment of Epstein-Barr Virus-Associated and Negative Gastric Cancer: Implication in Prognosis and Immunotherapy. Oncotarget 2017, 8, 67094–67103. [Google Scholar] [CrossRef] [PubMed]
- Lima, Á.; Sousa, H.; Medeiros, R.; Nobre, A.; Machado, M. PD-L1 Expression in EBV Associated Gastric Cancer: A Systematic Review and Meta-Analysis. Discov. Oncol. 2022, 13, 19. [Google Scholar] [CrossRef]
- Osumi, H.; Kawachi, H.; Murai, K.; Kusafuka, K.; Inoue, S.; Kitamura, M.; Yoshio, T.; Kakusima, N.; Ishihara, R.; Ono, H.; et al. Risk Stratification for Lymph Node Metastasis Using Epstein-Barr Virus Status in Submucosal Invasive (PT1) Gastric Cancer without Lymphovascular Invasion: A Multicenter Observational Study. Gastric. Cancer 2019, 22, 1176–1182. [Google Scholar] [CrossRef]
- Liu, X.; Choi, M.G.; Kim, K.; Kim, K.M.; Kim, S.T.; Park, S.H.; Cristescu, R.; Peter, S.; Lee, J. High PD-L1 Expression in Gastric Cancer (GC) Patients and Correlation with Molecular Features. Pathol. Res. Pract. 2020, 216, 152881. [Google Scholar] [CrossRef]
- Huang, Y.H.; Zhang, C.Z.Y.; Huang, Q.S.; Yeong, J.; Wang, F.; Yang, X.; He, Y.F.; Zhang, X.L.; Zhang, H.; Chen, S.L.; et al. Clinicopathologic Features, Tumor Immune Microenvironment and Genomic Landscape of Epstein-Barr Virus-Associated Intrahepatic Cholangiocarcinoma. J. Hepatol. 2021, 74, 838–849. [Google Scholar] [CrossRef]
- Chan, A.W.H.; Tong, J.H.M.; Sung, M.Y.M.; Lai, P.B.S.; To, K.F. Epstein-Barr Virus-Associated Lymphoepithelioma-like Cholangiocarcinoma: A Rare Variant of Intrahepatic Cholangiocarcinoma with Favourable Outcome. Histopathology 2014, 65, 674–683. [Google Scholar] [CrossRef]
- Egawa, N.; Egawa, K.; Griffin, H.; Doorbar, J. Human Papillomaviruses; Epithelial Tropisms, and the Development of Neoplasia. Viruses 2015, 7, 3863–3890. [Google Scholar] [CrossRef]
- Bansal, A.; Singh, M.; Rai, B. Human Papillomavirus-Associated Cancers: A Growing Global Problem. Int. J. Appl. Basic. Med. Res. 2016, 6, 84. [Google Scholar] [CrossRef] [PubMed]
- Hübbers, C.U.; Akgül, B. HPV and Cancer of the Oral Cavity. Virulence 2015, 6, 244. [Google Scholar] [CrossRef] [PubMed]
- de Abreu, P.M.; Có, A.C.G.; Azevedo, P.L.; do Valle, I.B.; de Oliveira, K.G.; Gouvea, S.A.; Cordeiro-Silva, M.F.; Louro, I.D.; de Podestá, J.R.V.; Lenzi, J.; et al. Frequency of HPV in Oral Cavity Squamous Cell Carcinoma. BMC Cancer 2018, 18, 324. [Google Scholar] [CrossRef] [PubMed]
- Wickenhauser, C.; Bethmann, D.; Kappler, M.; Eckert, A.W.; Steven, A.; Bukur, J.; Fox, B.A.; Beer, J.; Seliger, B. Tumor Microenvironment, HLA Class I and APM Expression in HPV-Negative Oral Squamous Cell Carcinoma. Cancers 2021, 13, 620. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, T.; Yoshikawa, K.; Suzuki, S.; Gosho, M.; Ueda, R.; Kazaoka, Y. Tumor-Infiltrating FoxP3+ T Cells Are Associated with Poor Prognosis in Oral Squamous Cell Carcinoma. Clin. Exp. Dent. Res. 2022, 8, 152–159. [Google Scholar] [CrossRef] [PubMed]
- BlattID, S.; Krüger, M.; Rump, C.; Zimmer, S.; Sagheb, K.; Kü nzelID, J. Differences in PD-L1 Expression between Oral and Oropharyngeal Squamous Cell Carcinoma. PLoS ONE 2022, 17, e0269136. [Google Scholar] [CrossRef]
- Martin, D.; Balermpas, P.; Winkelmann, R.; Rödel, F.; Rödel, C.; Fokas, E. Anal Squamous Cell Carcinoma—State of the Art Management and Future Perspectives. Cancer Treat. Rev. 2018, 65, 11–21. [Google Scholar] [CrossRef]
- McBride, A.A. Oncogenic Human Papillomaviruses. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2017, 372, 20160273. [Google Scholar] [CrossRef]
- Faber, M.T.; Frederiksen, K.; Palefsky, J.M.; Kjaer, S.K. Risk of Anal Cancer Following Benign Anal Disease and Anal Cancer Precursor Lesions: A Danish Nationwide Cohort Study. Cancer Epidemiol. Biomarkers Prev. 2020, 29, 185–192. [Google Scholar] [CrossRef]
- Palefsky, J.M.; Lee, J.Y.; Jay, N.; Goldstone, S.E.; Darragh, T.M.; Dunlevy, H.A.; Rosa-Cunha, I.; Arons, A.; Pugliese, J.C.; Vena, D.; et al. Treatment of Anal High-Grade Squamous Intraepithelial Lesions to Prevent Anal Cancer. N. Engl. J. Med. 2022, 386, 2273–2282. [Google Scholar] [CrossRef]
- Liu, Y.; Gaisa, M.M.; Wang, X.; Swartz, T.H.; Arens, Y.; Dresser, K.A.; Sigel, C.; Sigel, K. Differences in the Immune Microenvironment of Anal Cancer Precursors by HIV Status and Association With Ablation Outcomes. J. Infect. Dis. 2018, 217, 703–709. [Google Scholar] [CrossRef] [PubMed]
- Shamseddine, A.A.; Burman, B.; Lee, N.Y.; Zamarin, D.; Riaz, N. Tumor Immunity and Immunotherapy for HPV-Related Cancers. Cancer Discov. 2021, 11, 1896–1912. [Google Scholar] [CrossRef] [PubMed]
- Bian, J.J.; Almhanna, K. Anal Cancer and Immunotherapy-Are We There Yet? Transl. Gastroenterol. Hepatol. 2019, 4, 57. [Google Scholar] [CrossRef] [PubMed]
- Ciardiello, D.; Pio Guerrera, L.; Anna Maiorano, B.; Parente, P.; Pia Latiano, T.; di Maio, M.; Ciardiello, F.; Troiani, T.; Martinelli, E.; Maiello, E. Immunotherapy in Advanced Anal Cancer: Is the Beginning of a New Era? Cancer Treat. Rev. 2022, 105, 102373. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, T.A.; Rajdev, L.; Rhodes, A.; Dantanarayana, A.; Tennakoon, S.; Chea, S.; Spelman, T.; Lensing, S.; Rutishauser, R.; Bakkour, S.; et al. Impact of Anti-PD-1 and Anti-CTLA-4 on the Human Immunodeficiency Virus (HIV) Reservoir in People Living with HIV with Cancer on Antiretroviral Therapy: The AIDS Malignancy Consortium 095 Study. Clin. Infect. Dis. 2021, 73, e1973–e1981. [Google Scholar] [CrossRef] [PubMed]
- Miles, B.A.; Monk, B.J.; Safran, H.P. Mechanistic Insights into ADXS11-001 Human Papillomavirus-Associated Cancer Immunotherapy. Gynecol. Oncol. Res. Pract. 2017, 4, 9. [Google Scholar] [CrossRef]
- Phase 1-2 Study of ADXS11-001 or MEDI4736 Alone or Combo in Cervical or HPV+ Head & Neck Cancer—Full Text View—ClinicalTrials.Gov. Available online: https://clinicaltrials.gov/ct2/show/NCT02291055 (accessed on 23 October 2022).




{kind=link}
{kind=link}
{kind=link}
| HCV Non-Structural and Structural Proteins | Effect |
|---|---|
| Core protein, NS2, NS3, NS5A | Inhibition of apoptosis |
| Core, NS5A | Reactive oxygen species → TGF-β → HSCs activated → fibrogenesis |
| E1, E2, NS5A | Chronic inflammation via cytokine secretion |
| E2, core, NS2, NS3, NS5A | Cell proliferation |
| Core | EMT Mutagenesis and genome instability Angiogenesis via VEGF Lipid metabolism modification → steatosis |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sarantis, P.; Trifylli, E.-M.; Koustas, E.; Papavassiliou, K.A.; Karamouzis, M.V.; Papavassiliou, A.G. Immune Microenvironment and Immunotherapeutic Management in Virus-Associated Digestive System Tumors. Int. J. Mol. Sci. 2022, 23, 13612. https://doi.org/10.3390/ijms232113612
Sarantis P, Trifylli E-M, Koustas E, Papavassiliou KA, Karamouzis MV, Papavassiliou AG. Immune Microenvironment and Immunotherapeutic Management in Virus-Associated Digestive System Tumors. International Journal of Molecular Sciences. 2022; 23(21):13612. https://doi.org/10.3390/ijms232113612
Chicago/Turabian StyleSarantis, Panagiotis, Eleni-Myrto Trifylli, Evangelos Koustas, Kostas A. Papavassiliou, Michalis V. Karamouzis, and Athanasios G. Papavassiliou. 2022. "Immune Microenvironment and Immunotherapeutic Management in Virus-Associated Digestive System Tumors" International Journal of Molecular Sciences 23, no. 21: 13612. https://doi.org/10.3390/ijms232113612
APA StyleSarantis, P., Trifylli, E.-M., Koustas, E., Papavassiliou, K. A., Karamouzis, M. V., & Papavassiliou, A. G. (2022). Immune Microenvironment and Immunotherapeutic Management in Virus-Associated Digestive System Tumors. International Journal of Molecular Sciences, 23(21), 13612. https://doi.org/10.3390/ijms232113612