Abstract
Muscarinic acetylcholine receptors are membrane receptors involved in many physiological processes. Malfunction of muscarinic signaling is a cause of various internal diseases, as well as psychiatric and neurologic conditions. Cholesterol, neurosteroids, neuroactive steroids, and steroid hormones are molecules of steroid origin that, besides having well-known genomic effects, also modulate membrane proteins including muscarinic acetylcholine receptors. Here, we review current knowledge on the allosteric modulation of muscarinic receptors by these steroids. We give a perspective on the research on the non-genomic effects of steroidal compounds on muscarinic receptors and drug development, with an aim to ultimately exploit such knowledge.
1. Introduction
Muscarinic acetylcholine receptors (mAChRs) are members of the G-protein-coupled receptor (GPCR) family, and are represented by five distinct receptor subtypes, M1–M5 [1]. When activated by their endogenous agonist acetylcholine (ACh), mAChRs exert their functions through second messenger cascades by coupling to specific classes of G-proteins (Figure 1). The M1, M3 and M5 subtypes preferentially activate phospholipase C (PLC) and promote calcium mobilization through Gq/11, while M2 and M4 receptors inhibit the activity of adenylyl cyclase and thus cAMP synthesis via the Gi/o family of G-proteins [2].
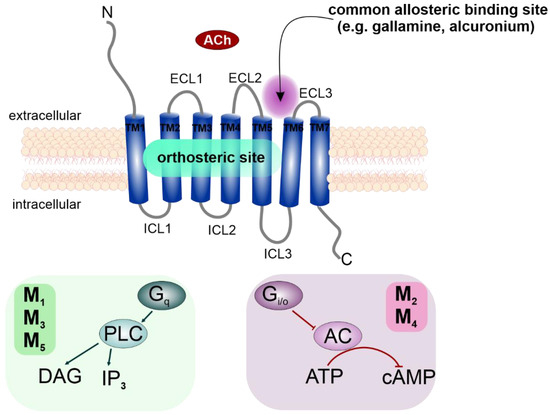
Figure 1.
Schematic structure of mAChRs and their preferential signaling pathways. Classical allosteric modulators of mAChRs, such as gallamine or alcuronium, bind to the extracellular part of the receptor between ECL2 and ECL3 [8]. Ach—acetylcholine; ECL—extracellular loop; TM—transmembrane α-helix; ICL—intracellular loop; DAG—diacylglycerol; IP3—inositol triphosphate; AC—adenylyl cyclase; PLC—phospholipase A.
Given the distribution of individual mAChRs subtypes, their expression levels and activation of distinct signaling cascades, these receptors play an important role in mediating a wide range of physiological functions in the central and peripheral nervous systems [3]. Malfunction or dysregulation of cholinergic signaling mediated by these receptors is strongly associated with the development of multiple pathological conditions and, as a consequence, targeting individual mAChRs subtypes represents a promising therapeutic approach for the treatment of neurologic and psychiatric conditions, e.g., Alzheimer’s disease, Parkinson’s disease, schizophrenia, substance abuse (for review see [4]) and diseases such as type 2 diabetes, asthma, cardiovascular diseases, and incontinence [5,6,7].
The mAChRs are also characterized by the highly conserved structure of their orthosteric binding sites; thus, it is virtually impossible to selectively activate individual subtypes of the receptor. This issue directed the research toward the development of compounds which act as allosteric modulators of mAChRs. By definition, allosteric modulators bind to a site spatially distinct from that of the endogenous transmitter; they change receptor conformation, leading to alterations in binding properties of the specific ligand (ACh for mAChRs), i.e., its affinity and ultimately potency and efficacy [9,10]. Allosteric binding sites of mAChR are far less conserved in their structures and thus offer a possibility for the development of mAChRs subtype-specific compounds [11]. Consequently, in the past decades, a great effort has been dedicated to the research and development of allosteric modulators that bind to these less conserved sites.
The most studied allosteric modulators of muscarinic receptors are neuromuscular blockers, such as gallamine [12,13], alcuronium and pancuronium [14]. According to the literature, many other allosteric modulators have been described. For example, thiochrome [15], verapamil [16], strychnine, [17], (−)-eburnamonine [18], fangicholine and tetrandrine [19], and 9-methoxy-α-lapachone [20], along with many others. The allosteric modulation of muscarinic receptors has been studied in a variety of pathological states [21,22,23]. As result of an enormous study, selective allosteric modulators have been identified. For example, benzylquinoline carboxylic acid (M1-selective allosteric modulators) [24,25] and compounds VU0010010, VU0152099, VU0152100, and LY2033298 (M4-selective allosteric modulators) have all been determined as such [26,27,28]. The selective positive allosteric modulation is considered a druggable target for the potential treatment of psychiatric and neurologic disorders, like Alzheimer’s disease or schizophrenia.
Multiple reports have shown that the interaction of steroids with mAChRs affects the ligand binding and functional responses of these receptors. It was demonstrated that molecules of membrane cholesterol (CHOL) change the affinity of muscarinic ligands and affect mAChRs activation dynamics [29,30,31]. CHOL represents a structural building block for all endogenous steroids, including steroid hormones (SHs) [32,33,34,35]. Typically, SHs (corticosteroids, sex steroids, bile acids) exert their well-known regulatory effects via activation of nuclear receptors (NRs), resulting in gene transcription and protein synthesis. This accounts for their long-lasting genomic effects from hours to days. It is noteworthy that SHs are also known to directly bind to and activate their specific membrane receptors, such as the progesterone receptor (mPR) or androgen receptor (mAR), along with estrogen membrane receptors α and β (mERα and mERβ), as well as the GPCR receptor GPR30 [36,37]. However, direct binding and modulation of actions of membrane-located receptors is a much faster process than the classic genomic effects, and lasts from milliseconds to minutes [36].
The non-genomic properties are characteristic of steroids synthesized de novo from the CHOL in the nervous system, in particular for neurosteroids (NSs) and also SHs [38]. Their synthetic analogues, which employ the same mode of action, are called neuroactive steroids (NASs). By acting through numerous ligand-gated ion channels, voltage-gated ion channels, or GPCRs, NSs produce various effects on the central and peripheral nervous system (CNS and PNS) [33,39]. Among NSs, SHs are also known to affect ACh release and cholinergic neurotransmission via interaction with mAChRs, improving memory and cognition [40,41]. Multiple reports describe that SHs bind to mAChRs and inhibit the interaction with muscarinic ligands at micromolar or higher concentrations [42,43,44,45], suggesting that steroids present in the body at physiological, i.e., nanomolar concentrations, cannot modulate mAChRs. However, it was recently proven otherwise. SHs can also act as NSs, i.e., progesterone and corticosterone, which have been shown to bind to mAChRs and modulate them in an allosteric manner at nanomolar concentrations [46]. Moreover, presumed binding sites of NSs and NAS at mAChRs were also identified [47].
There is a growing need for summarization and discussion of previous reports concerning the effects of steroids on mAChRs. Until now, an exact mechanism of action and a manner of steroids binding to these receptors has not been fully understood. In this review, therefore, we will focus on the mAChRs in the CNS and the allosteric modulation of their activity by endogenous and exogenous steroids. In particular, we describe the direct and indirect modulation of mAChRs by CHOL, SHs, NSs and NASs.
2. Cholesterol, Neurosteroids and Neuroactive Steroids
2.1. Cholesterol
A steroidal molecule is characterized by a tetracyclic cyclopenta[a]phenanthrene skeleton that has a specific position numbering and ring letters (Figure 2). The primary function of CHOL is structural. It serves as the main building block for synthesizing various SHs (gonadal sex hormones and adrenal glucocorticoids and mineralocorticoids), vitamin D, bile acids, and also NSs. A simplified scheme of steroid biosynthesis is summarized in Figure 3.
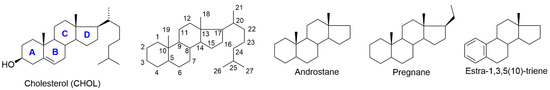
Figure 2.
Structure of cholesterol with ring letters (A–D), ring numbering (1–27) and trivial names of basic skeletons relevant for this review.
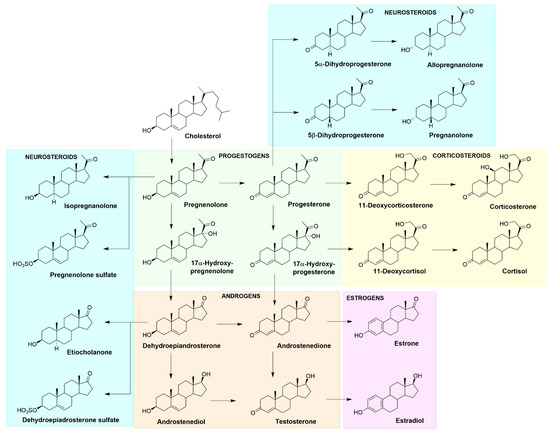
Figure 3.
Simplified scheme of steroid biosynthesis, including the major classes of steroid hormones and examples of neurosteroids.
CHOL is also an essential component of the cell membranes, maintaining their fluidity and integrity. Within the membrane, a polar C-3 hydroxyl group of CHOL interacts with surrounding phospholipids and proteins, while the tetracyclic steroid skeleton with the lipophilic C-17 substituent interacts with the fatty acids. This enables the integration of CHOL into the lipid bilayer and secures its integrity [48]. Molecules of CHOL are distributed throughout the plasma membranes and may form dimers, as detected in X-ray crystal structures of membrane proteins [49], or concentrate in specialized sphingolipid-rich domains known as lipid rafts [50]. Fractions of membranes rich in CHOL are thicker and more rigid. Near the receptors, lipid rafts interfere with the machinery of signal transduction [50], diminishing the availability of signaling molecules and affecting the activity of the receptors [51]. Thus, the close interaction of CHOL with membrane proteins, including ion channels and GPCRs, affects processes of ligand binding, receptor activation and signal transduction [52,53,54].
Lipid molecules are frequently found in X-ray and cryo-EM structures of GPCRs, indicating that these lipids may specifically interact with GPCRs in their membrane environment [54]. If so, then they may allosterically modulate ligand binding to the receptors and the functional response of receptors to agonists. As CHOL binds to multiple specific binding sites on many GPCRs, it can be considered their allosteric modulator [55,56,57]. Indeed, membrane CHOL was found to co-crystallize with various GPCRs for distinct classes of agonists as published in the RCSB database (https://www.rcsb.org/ (accessed on 20 February 2022)). As for the mAChRs, CHOL was not found in the crystal structures of the receptors. However, its binding site was revealed using molecular docking [31]. The molecules of CHOL can directly influence GPCR activity by altering the binding of a specific ligand, affecting receptor activation as well as a receptor-to-G-protein coupling. CHOL can also affect GPCRs indirectly through changes in the membrane organization, such as alterations in the fluidity of the membrane surrounding the receptor and thus effectors available for signaling (signal trafficking). For review, see [10,58,59].
2.2. NSs, NASs–Functions and Their Genomic and Non-Genomic Effects
As mentioned previously, NSs represent a class of endogenous compounds synthesized de novo in CNS from CHOL or steroidal precursors imported from peripheral endocrine glands. NSs are known to modulate neuronal excitability by acting through various ligand-gated ion channels and GPCRs [38,54], both, positively and/or negatively. The best-known function of NSs in CNS is the modulation of γ-aminobutyric acid (GABAA) receptors responsible for inhibitory neurotransmission in the brain [60]. Further, some NSs modulate the N-methyl-D-aspartate (NMDA) glutamate receptors, α-amino-3-hydroxy-5-methyl-4-isoxazolepropionate (AMPA)/kainate receptors, glycine or nicotinic acetylcholine receptors (nAChRs) [33]. Consequently, NSs are involved in the regulation of various CNS functions such as cognition and memory processes [61]. Moreover, NSs also modulate pain pathways [62,63,64,65], and exert neuroprotective effects [66,67,68], among a myriad of other actions. For review, see [69].
In contrast, SHs, by definition, should exert multiple functions via the activation of nuclear receptors (NRs) specific for steroid hormones [70]. Typically, activation of NRs induces gene transcription and protein synthesis, therefore their action is relatively slow (hours to days) [36]. Nevertheless, there is a piece of strong evidence that, in addition to the classical genomic mechanism of action, SHs can exert rapid, non-genomic signaling via interaction with membrane receptors [36,71,72]. Consequently, the literature describes the crucial role of SHs in the development and functioning of the CNS [72,73,74,75]. For example, synthesized locally within CNS, progesterone [76] and estradiol [77] influence neuronal functions and produce a variety of effects that are unrelated to reproduction [78,79,80,81]. Acting as NSs, estradiol, progesterone as well as corticosterone, regulate cognition [82,83,84], memory [85,86], brain development [87] and behaviour [88].
On the other hand, some steroids share both hormonal and neurosteroid activity. For example, allopregnanolone—a well-known example of a potent allosteric modulator of GABAA receptors—was demonstrated to also exert genomic effects via activation of mPRs [89,90]. Similarly, dehydroepiandrosterone (DHEA), a metabolic intermediate in the biosynthesis of many SHs and the most abundant hormone in mammals, is secreted by the adrenal gland and ovary. Its hormonal effects are mediated through androgen and estrogen receptors, peroxisome proliferator-activated receptor (PPAR), pregnane X receptor (PXR), and the constitutive androstane receptor (CAR) [91]. Regarding neurosteroid activity of DHEA, it has been described as a ligand of GABAA, NMDA, sigma-1 receptors and L-type calcium channels. This explains the effects of DHEA on physiological functions and pathological abnormalities in the brain [92,93].
Taken together, the current literature shows that the effects of steroids are complex and cannot be assigned to a single mode of action. Such a multi-target mode of action may explain their unique drug-likeness in a variety of neurological and psychiatric conditions. When interacting with membrane receptors, the effects of NSs are exerted in a manner of non-genomic signaling. However, chronic exposure to NSs may indirectly (non-genomic pathways) induce genomic action (e.g., changes in receptor expression) [33,94]. Therefore, the crosstalk between genomic and non-genomic steroid effects needs to be taken into consideration. The interplay of genomic and non-genomic actions of NASs is summarised in Figure 4.
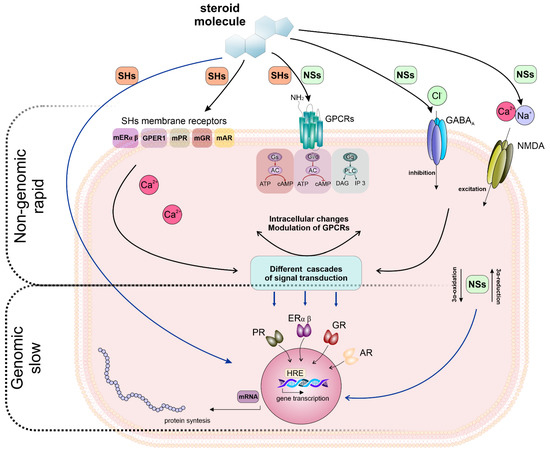
Figure 4.
The interplay between genomic and non-genomic effects of steroids. Top, non-genomic, rapid (seconds to minutes) signaling: the activation of membrane-localized receptors for estrogen (mERα; mERβ; GPER1/GPR30), progesterone (mPR), glucocorticoids (mGR) and androgens (mAR) by a specific hormone modulates numerous signaling cascades and produces different cellular effects [95,96]. NSs exert their rapid, non-genomic effects via modulation of membrane ionotropic receptors and channels, e.g., γ-aminobutyric acid receptors, GABAA or NMDA receptors, resulting in excitability changes within neurons. Activation of Gs protein results in stimulation of adenylyl cyclase (AC) and cAMP synthesis; activation of Gq/11 protein results in the activation of phospholipase C (PLC) and production of inositol 1,4,5-trisphosphate (IP3) and diacylglycerol (DAG); activation of Gi/o proteins inhibits AC and cAMP synthesis. Middle, metabolites of neurosteroid (NSs) produced by intracellular oxidation bind to steroid receptors [33,35]. Bottom, slow genomic effects (minutes to hours): steroid hormones (SHs) bind to their specific intracellular class I nuclear receptors (progesterone receptor (PR), oestrogen receptors (ERα and β), glucocorticoid receptors (GR) or androgen receptors (AR)) which, in the absence of the ligand, reside in the cytosol. The binding of the ligand to these receptors results in the translocation of the receptor–ligand complex to the nucleus where it binds to specific hormone response elements (HREs) and regulates gene transcription. The figure was prepared according to [33,36,94].
3. Muscarinic Receptors
Muscarinic acetylcholine receptors (mAChR) are members of class A, Rhodopsine-like GPCRs, and are represented by five distinct receptor subtypes, M1–M5 [1]. Like all GPCRs, mAChRs are integral membrane proteins consisting of seven transmembrane α-helices (TM1 to TM7) connected via three intracellular (ICL1 to ICL3) and three extracellular (ECL1 to ECL3) loops (Figure 1). Individual TM helixes form a hydrophilic pocket (orthosteric binding site) accessible from the extracellular side for endogenous signaling molecules. mAChRs activation by their endogenous agonist ACh results in subsequent G-protein activation, and depending on the G-protein class, mediates various cellular responses [97,98].
Activated mAChRs trigger distinct second messenger cascades coupled to designated G-protein classes and thus mediate a wide range of physiological functions throughout the body. M1, M3, and M5 mAChR subtypes preferentially activate Gq/11 G-proteins to stimulate phospholipase C (PLC) and induce the mobilisation of intracellular calcium. M2 and M4 receptors activate Gi/o G-proteins, the α-subunit of which inhibits adenylyl cyclase (AC), decreasing the production of cAMP, while the βγ-dimer of G-proteins modulates conductance of K+ and Ca2+ channels [3,99]. Besides these preferential signaling pathways, muscarinic agonists may activate also other ones, termed non-preferential signaling pathways [100,101].
Specific targeting of mAChRs subtypes, and thus selective regulation of their signaling pathways, might be of great value in seeking the treatment for the CNS [4] and diseases affecting internal organs [5,6,7].
4. Direct Effects of Steroids on mAChRs
4.1. Direct Effects of Cholesterol
Molecules of CHOL can directly influence GPCRs by altering the binding of a specific ligand, activation of a receptor as well as preferences of receptor-to-G-protein coupling. In contrast, CHOL can affect GPCRs indirectly through changes in the membrane organization, such as alterations in the fluidity of the membranes surrounding GPCRs and thus signal trafficking [58,59].
The membrane CHOL modulates GPCRs by acting on their allosteric binding sites. CHOL-binding motifs were predicted based on analyses of X-ray and cryo-EM structures of various GPCRs. Three CHOL-binding motifs were described so far. The motif common to all membrane proteins is the Cholesterol Recognition Amino acid Consensus (CRAC) [102] and its inverse variant (CARC) [103]. The so-called Cholesterol Consensus Motif (CCM), the groove formed by the transmembrane domains TM2, TM3 and TM4, was identified in the structure of the β2-adrenergic receptor [56]. As CHOL-binding sites on GPCRs are distinct from the binding sites of endogenous transmitters, CHOL can be considered an allosteric modulator [56]. Allosteric binding sites on mAChRs represent far less conserved structures compared to their orthosteric binding site, and offer a possibility to target specific receptor subtypes [11].
Initially, it was demonstrated that CHOL directly affects the affinity of muscarinic ligands. An increase in the content of CHOL within the membrane resulted in a reduced affinity for the muscarinic agonist carbachol at M2, but increased its affinity at M1 and M3 receptors. On the other hand, CHOL depletion increased the affinity of carbachol to M1, M2, and M3 subtypes. In contrast, CHOL depletion was shown to diminish the affinity of the muscarinic antagonist N-methylscopolamine (NMS) at these receptors. Enrichment of membranes with CHOL caused a decrease in affinity for NMS at M1 and M3, and an increase in affinity at the M2 receptor [29,30].
Changes in the content of the membrane CHOL also affect preferential and non-preferential signaling through the M2 as well as M1, and M3 expressed in CHO cells [29,30]. CHOL-dependent changes in preferential mAChRs signaling are presented in Figure 5. Regarding M2, CHOL depletion significantly strengthens the preferential signaling pathway Gi/o and reinforces the maximal effect of inhibition of cAMP synthesis. It also stimulated non-preferential Gs and Gq/11 signaling pathways, as shown in Figure 5B. Additionally, in the case of M1 and M3 receptors, both gradually increase and decrease in membrane CHOL concentration, resulting in a concentration-dependent inhibition of the preferential signaling pathway via Gq/11 and a decrease in accumulation of inositol trisphosphate(Figure 5B,C). As for the non-preferential Gs-mediated signaling, an increase in membrane CHOL concentration inhibited the cAMP accumulation, while a decrease in membrane CHOL concentration stimulated cAMP synthesis.
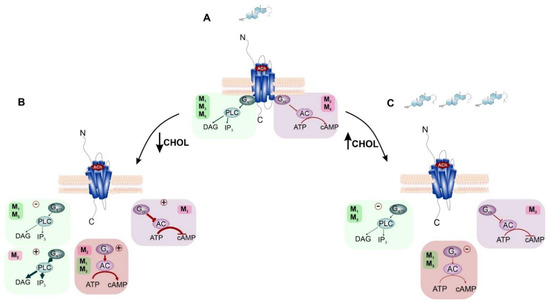
Figure 5.
Influence of membrane CHOL content on preferential and non-preferential signaling of mAChRs. (A) standard mAChRs signaling in the cell membrane with a natural level of CHOL (light blue steroid molecule). Preferential coupling of M1, M3, M5 receptors to Gq/11 protein (green box) and M2, M4 receptors coupling to Gi/o protein (purple box). (B) depletion of membrane CHOL diminishes preferential Gq/11 signaling and enhances preferential Gi/o and non-preferential Gs (red box) and Gq/11 signaling. (C) increase in membrane CHOL level attenuates signaling via preferential G-proteins.
Based on molecular modeling, the CHOL allosteric binding site was found in the intracellular leaflet of the membrane between TM6 and TM7 of mAChRs [31]. This binding site presumably also represents a site of binding for various steroidal compounds. In addition, subtype specificity of some ligands was shown to be affected by the content of membrane CHOL. Specifically, binding of CHOL at the TM6 and TM7 interface attenuates activation of M1, M4 and M5 receptors [31].
Xanomeline (3-(hexyloxy)-4-(1-methyl-1,2,5,6-tetrahydropyridin-3-yl)-1,2,5-thiadiazole) is a muscarinic agonist considered functionally selective for M1 and M4 receptors, developed for treatment of Alzheimer’s disease [104,105]. Xanomeline binding to mAChRs is partially resistant to washing [106]. Wash-resistant xanomeline steadily activates mAChRs with an exception of the M5 subtype [107]. Mutation of leucine 6.46 to isoleucine at the CHOL binding site in M1 and M4 receptors resulted in receptors insensitive to activation by wash-resistant xanomeline. On the other hand, the mutation of isoleucine 6.46 to leucine in the M5 receptor made it sensitive to activation by wash-resistant xanomeline. Decreasing membrane CHOL content reversed the effects of mutations, indicating that xanomeline functional selectivity is rather the result of specific receptor–membrane interactions than agonist–receptor interactions [31]. Thus, changing membrane CHOL level or interaction of a receptor with the membrane might represent a novel possibility to achieve pharmacological selectivity for mAChRs.
4.2. Non-Genomic Effects of NSs and NASs on mAChRs
Historically, multiple reports showed that the interaction of SHs with mAChRs alters the binding dynamics of various ligands [42,43,44,108,109,110,111]. Early research conducted on the direct effects of SHs on mAChRs was focused on changes in the binding of radiolabelled muscarinic antagonists like [3H]-quinuclidinyl benzilate ([3H](-)QNB), N-methyl-[3H]-4-piperidyl benzilate ([3H]4NMPB) or N-methyl-[3H]-scopolamine ([3H]NMS) in the cell membranes prepared from rat brain tissues. In competitive binding studies with [3H]4NMPB, the steroids progesterone and estradiol affected the binding properties of the mAChR agonist, oxotremorine. Both steroids decreased the affinity and proportion of the high-affinity binding sites [112]. Later in experiments with [3H]NMS, other researchers confirmed that progesterone and estradiol (but not testosterone) bind to mAChRs in the membranes prepared from the rat hypothalamus and amygdala tissues [113].
In the study by Klangkagaya and Chan [44], the effects of 50 steroid compounds on [3H](-)QNB binding to mAChRs in hypothalamic and pituitary membranes was reported. The structures of active pregnane and androstane compounds, including their IC50 values in inhibiting the [3H](-)QNB binding, are summarized in Figure 6 and Figure 7, respectively. The results of this study demonstrated that the pregnane skeleton is considerably more relevant for further development than the androstane skeleton. Further, it was determined that incomplete inhibition of [3H](-)QNB binding by tested steroidal compounds indicates allosteric modulation of [3H](-)QNB binding [44].
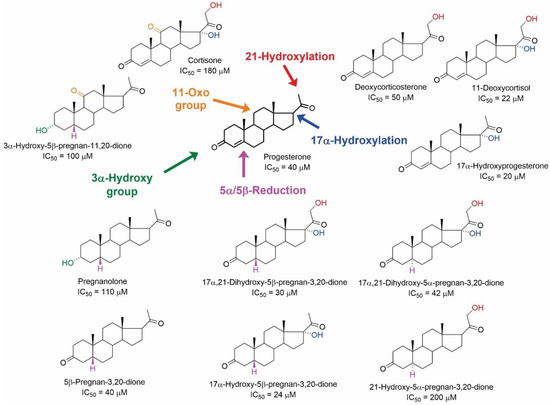
Figure 6.
Structures of pregnane steroids with their IC50 values in inhibiting the [3H](-)QNB binding from the study of Klangkalya and Chan [44].
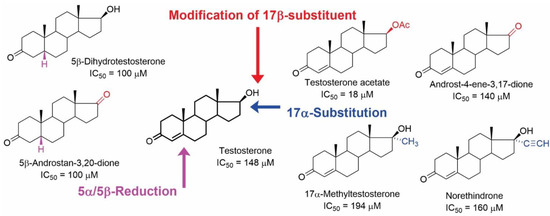
Figure 7.
Structures of androstane steroids with their IC50 values in inhibiting the [3H](-)QNB binding from the study of Klangkalya and Chan [44].
Regarding the structure–activity relationship, modifications of the progesterone skeleton afforded structures with IC50 values in the tens of µM. In contrast, except for testosterone acetate with an IC50 value of 18 µM, the IC50 values of androstane analogues varied from 100 to 200 µM. Interestingly, the 17α-hydroxy-substituted pregnane skeletons were active, except for hydrocortisone and 17α-hydroxy-5α-pregnan-3,20-dione (IC50 > 200 µM). Similarly, the hydroxylation of the skeleton in position C-21 was tolerated well. In contrast, hydroxylation at position C-11 strongly diminished the affinity for mAChRs (corticosterone and hydrocortisone, IC50 > 200 µM), while the presence of the 11-oxo group decreased the affinity only slightly. Reduction of the Δ3,4-enone to 5β-steroids afforded compounds with higher affinity than the corresponding 5α-analogues. Accordingly, the orientation of the hydrogen atom at the C-5 position was identified as crucial for the inhibition of [3H](-)QNB binding.
As mentioned previously, modification of the testosterone skeleton did not afford active compounds with low micromolar affinities (Figure 7). Out of 50 tested compounds, 16 of them were androstanes, and 7 of them showed the ability to inhibit [3H](-)QNB binding to mAChRs. It should be mentioned that estradiol was also inactive (IC50 > 200 µM) [44].
Further, the allosteric mode of mAChRs modulation by NSs and NASs was described in the study of Shiraishi et al. [111]. The synthetic analgesic neurosteroid alfaxalone (3α-hydroxy-5β-pregnane-11,20-dione) decreased [3H](-)QNB binding to M1 and M3 receptors (IC50 2.6 μM and 4.5 μM, respectively) and inhibited acetylcholine-induced Ca2+-activated Cl− currents in oocytes, expressing M1 and M3 receptors (IC50 values of 1.8 µM and 5.3 µM, respectively). A selective protein kinase C inhibitor GF109203X had a negligible effect on the inhibition of ACh-induced currents by alfaxalone, confirming allosteric modulation of these mAChRs [111].
Similarly, Horishita et al. [42] described voltage clamp experiments showing that pregnenolone and progesterone altered acetylcholine-induced Ca2+-activated Cl− currents, while DHEA did not affect the function of M1 and M3 receptors expressed in Xenopus oocytes. The IC50 values at M1 and M3 for progesterone were 2.5 and 3 μM, while for pregnenolone they were 11.4 and 6 μM, respectively. Further, both steroids were shown to diminish [3H](-)QNB binding to M1 and M3 receptors. Both steroids also affected affinity and binding capacity, indicating non-competitive inhibition, showing that tested steroids bind to M1 and M3 receptors at an allosteric binding site [42].
The literature summarized above describes the effects of steroids on specific ligand binding to muscarinic mAChRs. In general, these binding studies suggested that compounds sharing a steroid scaffold with CHOL bind to mAChRs at the site distinct from their natural agonist ACh or radiolabelled orthosteric ligands used in these experiments. Yet, except for the studies on the alteration of ACh-mediated responses in Xenopus oocytes [42,111], evidence gained from these reports accounted mainly for the manner of ligand–receptor interaction. Moreover, these reports describe the effects of steroids on mAChRs at micromolar or higher concentrations, which exclude their physiological relevance but pointed to their pharmacological potential.
The authors of the recent study examined the allosteric modulation of mAChRs by 20 steroidal compounds (Figure 8) [46]. This study revealed that all tested compounds changed [3H]NMS equilibrium binding at a 10 μM concentration. Moreover, some compounds exerted high-affinity binding with sub-micromolar affinity. Importantly, corticosterone and progesterone were found to bind to the mAChRs with about 100 nM affinity, which is within the physiological range [114,115]. In particular, the structure–activity relationship evaluation of the results [46] has shown that some of the compounds with the highest affinities to mAChRs have an enone group (3-oxo-4-ene structure) in the A-ring. Further, the 5β-steroids generally have higher affinities to all receptor subtypes than their 5α-analogues. The presence of the C-17 acetyl group was shown to represent a key structural element for affinity improvement. Corticosterone with hydroxyl groups at C-11 and C-21 had a higher binding affinity, while the presence and orientation of hydroxyl moiety at C-3 had no significant effect. The aromatization of A-ring, such as the formation of estradiol from testosterone, ended the ability of a compound to affect [3H]NMS binding to mAChR. These findings are in agreement with the structural features of steroids that diminish [3H](-)QNB binding to hypothalamic mAChR [44].
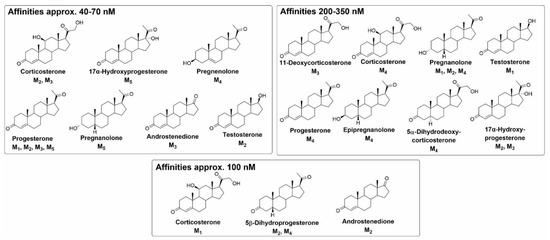
Figure 8.
Overview of binding affinities to individual muscarinic receptor subtypes from [46].
Four compounds, in particular corticosterone, progesterone, estradiol and testosterone, affected the functional response of mAChRs at physiologically relevant concentrations. The influence of steroids on mAChRs functional response to ACh was quantified by changes in specific [35S] GTPγS binding to membranes expressing M2 or M4, or inositol phosphates accumulation in CHO cells expressing M1, M3 or M5 receptors. Corticosterone induced a 3-fold increase in ACh potency at M2, but decreased it 3-fold at the M4 receptor. Progesterone increased the efficacy of ACh receptors by 30% at M1 and by 20% at M3 receptors and decreased it by 30% at M2 receptors. Estradiol increased the efficacy of ACh by 24% at the M1 receptor.
The follow-up study [47] describes the binding site on muscarinic acetylcholine receptors for NASs (Figure 9). It was found that NASs can bind to the two distinct allosteric binding sites on mAChRs, with approximately 100 nM and 10 μM affinities. The high-affinity binding site was investigated in [3H]NMS binding experiments using selected NAS in combination with well-known classic muscarinic allosteric modulators gallamine and alcuronium, and steroid allosteric modulators pancuronium, rapacuronium and WIN-compounds [47]. This high-affinity binding site was shown to be different from the common, extracellularly located allosteric binding site for alcuronium or gallamine, or the aminosteroid-based muscle relaxants pancuronium and rapacuronium. Interestingly, selected NAS bound to the same high-affinity binding site as steroid-based WIN-compounds that do not bind to the classical allosteric binding site located between the ECL2 and ECL3 [116,117,118,119,120]. Compounds 5α-androst-1-en-17β-yl 17-hemisuccinate (MS-96) and 17-methylene-5β-androstan-3α-yl 3-hemiglutarate (MS-112) were able to bind to this site with an affinity of about 50 nM and 16 nM, respectively. The authors have also shown that the membrane CHOL competes with NASs and WIN-compounds for binding to both high- and low-affinity binding sites. It suggests that the high-affinity binding site is rather oriented towards the inner side of the membrane, and that this site may represent a novel target for the allosteric modulation of muscarinic receptors. However, identification of the exact number and location of the CHOL binding sites at mAChRs remains to be determined [47,121].
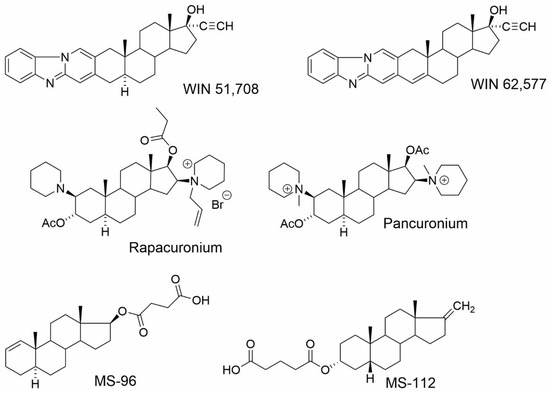
Figure 9.
Structures of WIN compounds, the skeletal muscle relaxants pancuronium and rapacuronium and NASs tested in the study of Dolejsi et al. [47].
5. Conclusions
Steroidal compounds such as cholesterol, neurosteroids, neuroactive steroids and steroid hormones bind to several sites on muscarinic acetylcholine receptors. From these sites, they allosterically modulate the binding of muscarinic ligands and the functional response of muscarinic receptors. They share a common high-affinity binding site that is oriented towards the membrane. Neurosteroids and steroid hormones allosterically modulate muscarinic receptors at physiologically relevant concentrations. The physiological non-genomic effects of neurosteroids and steroid hormones have not been studied in detail so far.
6. Perspectives
Allosteric modulation of muscarinic receptors by steroids proposes two new avenues for future research. One is an exploration of the physiology of the non-genomic effects of neurosteroids and steroid hormones at muscarinic receptors. Besides novel knowledge, an understanding of the non-genomic effects of neurosteroids and steroid hormones may bring new ways for the treatment of diseases resulting from a malfunction of muscarinic signaling by manipulation with levels of neurosteroids or steroid hormones. The other is exploiting differences in receptor–membrane interactions for the development of selective modulators. These differences can be approached in two ways. First, differences in receptor–membrane interactions among receptor subtypes allow the development of subtype-selective compounds. Second, differences in receptor–membrane interactions among various tissues give the opportunity for tissue-specific modulation. For example, drugs targeting cholesterol binding sites will be more efficient at cholesterol-lean membranes than at cholesterol-rich ones due to competition with membrane cholesterol.
Author Contributions
Funding
Supported by the project National Institute for Research of Metabolic and Cardiovascular Diseases (Programme EXCELES, ID Project No. LX22NPO5104)—Funded by the European Union—Next Generation EU and by the Czech Academy of Sciences institutional supports RVO:61388963 and RVO:67985823.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We thank Esam El-Fakahany, University of Minnesota College of Pharmacy, and Vladimír Doležal, Institute of Physiology CAS, for the critical reading of the manuscript. The Graphical abstract was created with BioRender.com.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the writing of the manuscript, or in the decision to publish.
References
- Bonner, T.I.; Buckley, N.J.; Young, A.C.; Brann, M.R. Identification of a Family of Muscarinic Acetylcholine Receptor Genes. Science 1987, 237, 527–532. [Google Scholar] [CrossRef]
- Caulfield, M.P. Muscarinic Receptors—Characterization, Coupling and Function. Pharmacol. Ther. 1993, 58, 319–379. [Google Scholar] [CrossRef]
- Eglen, R.M. Overview of Muscarinic Receptor Subtypes. In Handb Exp Pharmacol; Fryer, A.D., Arthur Christopoulos, N.N.M., Eds.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 3–28. ISBN 0171-2004. [Google Scholar]
- Scarr, E. Muscarinic Receptors: Their Roles in Disorders of the Central Nervous System and Potential as Therapeutic Targets. CNS Neurosci. Ther. 2012, 18, 369–379. [Google Scholar] [CrossRef]
- Abrams, P.; Andersson, K.-E.; Buccafusco, J.J.; Chapple, C.; De Groat, W.C.; Fryer, A.; Kay, G.; Laties, A.; Nathanson, N.; Pasricha, P.J.; et al. Muscarinic receptors: Their distribution and function in body systems, and the implications for treating overactive bladder. J. Cereb. Blood Flow Metab. 2006, 148, 565–578. [Google Scholar] [CrossRef]
- Gosens, R.; Zaagsma, J.; Meurs, H.; Halayko, A.J. Muscarinic Receptor Signaling in the Pathophysiology of Asthma and COPD. Respir. Res. 2006, 7, 73. [Google Scholar] [CrossRef]
- Gautam, D.; Han, S.J.; Duttaroy, A.; Mears, D.; Hamdan, F.F.; Li, J.H.; Cui, Y.; Jeon, J.; Wess, J. Role of the M3 Muscarinic Acetylcholine Receptor in β-Cell Function and Glucose Homeostasis. Diabetes, Obes. Metab. 2007, 9, 158–169. [Google Scholar] [CrossRef]
- Krejcí, A.; Tucek, S. Changes of Cooperativity between N-Methylscopolamine and Allosteric Modulators Alcuronium and Gallamine Induced by Mutations of External Loops of Muscarinic M(3) Receptors. Mol. Pharmacol. 2001, 60, 761–767. [Google Scholar]
- Mysliveček, J.; Říčný, J.; Kolář, F.; Tuček, S. The Effects of Hydrocortisone on Rat Heart Muscarinic and Adrenergic A1, Β1 and Β2 Receptors, Propranolol-Resistant Binding Sites and on Some Subsequent Steps in Intracellular Signalling. Naunyn. Schmiedebergs. Arch. Pharmacol. 2003, 368, 366–376. [Google Scholar] [CrossRef]
- Jakubik, J.; El-Fakahany, E.E. Current Advances in Allosteric Modulation of Muscarinic Receptors. Biomolecules 2020, 10, 325. [Google Scholar] [CrossRef]
- Zhang, Y.; Doruker, P.; Kaynak, B.; Zhang, S.; Krieger, J.; Li, H.; Bahar, I. Intrinsic Dynamics Is Evolutionarily Optimized to Enable Allosteric Behavior. Curr. Opin. Struct. Biol. 2020, 62, 14–21. [Google Scholar] [CrossRef]
- Clark, A.L.; Mitchelson, F. The Inhibitory Effect of Gallamine on Muscarinic Receptors. Br. J. Pharmacol. 1976, 58, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Stockton, J.M.; Birdsall, N.J.; Burgen, A.S.; Hulme, E.C. Modification of the Binding Properties of Muscarinic Receptors by Gallamine. Mol. Pharmacol. 1983, 23, 551–557. [Google Scholar] [PubMed]
- Nedoma, J.; Dorofeeva, N.A.; Tuček, S.; Shelkovnikov, S.A.; Danilov, A.F. Interaction of the Neuromuscular Blocking Drugs Alcuronium, Decamethonium, Gallamine, Pancuronium, Ritebronium, Tercuronium and d-Tubocurarine with Muscarinic Acetylcholine Receptors in the Heart and Ileum. Naunyn Schmiedebergs Arch. Pharmacol. 1985, 329, 176–181. [Google Scholar] [CrossRef] [PubMed]
- Lazareno, S.; Dolezal, V.; Popham, A.; Birdsall, N.J.M. Thiochrome Enhances Acetylcholine Affinity at Muscarinic M4 Receptors: Receptor Subtype Selectivity via Cooperativity Rather than Affinity. Mol. Pharmacol. 2004, 65, 257–266. [Google Scholar] [CrossRef]
- Waelbroeck, M.; Robberecht, P.; De Neef, P.; Christophe, J. Effects of Verapamil on the Binding Properties of Rat Heart Muscarinic Receptors: Evidence for an Allosteric Site. Biochem. Biophys. Res. Commun. 1984, 121, 340–345. [Google Scholar] [CrossRef]
- Proska, J.; Tucek, S. Competition between Positive and Negative Allosteric Effectors on Muscarinic Receptors. Mol. Pharmacol. 1995, 48, 696–702. [Google Scholar]
- Proška, J.; Tuček, S. Positive Allosteric Action of Eburnamonine on Cardiac Muscarinic Acetylcholine Receptors. Eur. J. Pharmacol. 1996, 305, 201–205. [Google Scholar] [CrossRef]
- Dong, G.Z.; Kameyama, K.; Rinken, A.; Haga, T. Ligand Binding Properties of Muscarinic Acetylcholine Receptor Subtypes (M1-M5) Expressed in Baculovirus-Infected Insect Cells. J. Pharmacol. Exp. Ther. 1995, 274, 378–384. [Google Scholar]
- Dong, G.Z.; Haga, T.; Itokawa, H.; Mizobe, F. Allosteric Binding of 9-Methoxy-Alpha-Lapachone and Alcuronium to the Muscarinic Acetylcholine Receptor M2 Subtype. Biomed. Res. 1995, 16, 327–335. [Google Scholar] [CrossRef]
- Birdsall, N.J.M.; Lazareno, S. Allosterism at Muscarinic Receptors: Ligands and Mechanisms. Mini Rev. Med. Chem. 2005, 5, 523–543. [Google Scholar] [CrossRef]
- Gregory, K.J.; Sexton, P.M.; Christopoulos, A. Allosteric Modulation of Muscarinic Acetylcholine Receptors. Curr. Neuropharmacol. 2007, 5, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Jakubík, J.; El-Fakahany, E.E. Allosteric Modulation of Muscarinic Acetylcholine Receptors. Pharmaceuticals 2010, 3, 2838–2860. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Seager, M.A.; Seager, M.; Wittmann, M.; Jacobson, M.; Bickel, D.; Burno, M.; Jones, K.; Graufelds, V.K.; Xu, G.; et al. Selective Activation of the M1 Muscarinic Acetylcholine Receptor Achieved by Allosteric Potentiation. Proc. Natl. Acad. Sci. USA 2009, 106, 15950–15955. [Google Scholar] [CrossRef] [PubMed]
- Shirey, J.K.; Brady, A.E.; Jones, P.J.; Davis, A.A.; Bridges, T.M.; Kennedy, J.P.; Jadhav, S.B.; Menon, U.N.; Xiang, Z.; Watson, M.L.; et al. A Selective Allosteric Potentiator of the M1 Muscarinic Acetylcholine Receptor Increases Activity of Medial Prefrontal Cortical Neurons and Restores Impairments in Reversal Learning. J. Neurosci. 2009, 29, 14271–14286. [Google Scholar] [CrossRef]
- Shirey, J.K.; Xiang, Z.; Orton, D.; Brady, A.E.; Johnson, K.A.; Williams, R.; Ayala, J.E.; Rodriguez, A.L.; Wess, J.; Weaver, D.; et al. An Allosteric Potentiator of M4 MAChR Modulates Hippocampal Synaptic Transmission. Nat. Chem. Biol. 2008, 4, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Brady, A.E.; Jones, C.K.; Bridges, T.M.; Kennedy, J.P.; Thompson, A.D.; Heiman, J.U.; Breininger, M.L.; Gentry, P.R.; Yin, H.; Jadhav, S.B.; et al. Centrally Active Allosteric Potentiators of the M4 Muscarinic Acetylcholine Receptor Reverse Amphetamine-Induced Hyperlocomotor Activity in Rats. J. Pharmacol. Exp. Ther. 2008, 327, 941–953. [Google Scholar] [CrossRef]
- Chan, W.Y.; McKinzie, D.L.; Bose, S.; Mitchell, S.N.; Witkin, J.M.; Thompson, R.C.; Christopoulos, A.; Lazareno, S.; Birdsall, N.J.M.; Bymaster, F.P.; et al. Allosteric Modulation of the Muscarinic M4 Receptor as an Approach to Treating Schizophrenia. Proc. Natl. Acad. Sci. USA 2008, 105, 10978–10983. [Google Scholar] [CrossRef]
- Michal, P.; Rudajev, V.; El-Fakahany, E.E.; Dolezal, V. Membrane Cholesterol Content Influences Binding Properties of Muscarinic M2 Receptors and Differentially Impacts Activation of Second Messenger Pathways. Eur. J. Pharmacol. 2009, 606, 50–60. [Google Scholar] [CrossRef]
- Michal, P.; El-Fakahany, E.E.; Doležal, V. Changes in Membrane Cholesterol Differentially Influence Preferential and Non-Preferential Signaling of the M1 and M3 Muscarinic Acetylcholine Receptors. Neurochem. Res. 2015, 40, 2068–2077. [Google Scholar] [CrossRef]
- Randáková, A.; Dolejší, E.; Rudajev, V.; Zimčík, P.; Doležal, V.; El-Fakahany, E.E.; Jakubík, J. Role of Membrane Cholesterol in Differential Sensitivity of Muscarinic Receptor Subtypes to Persistently Bound Xanomeline. Neuropharmacology 2018, 133, 129–144. [Google Scholar] [CrossRef]
- Acconcia, F.; Marino, M. Steroid Hormones: Synthesis, Secretion, and Transport. In Principles of Endocrinology and Hormone Action; Belfiore, A., LeRoith, D., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 1–31. ISBN 978-3-319-27318-1. [Google Scholar]
- Rupprecht, R.; Holsboer, F. Neuroactive Steroids: Mechanisms of Action and Neuropsychopharmacological Perspectives. Trends Neurosci. 1999, 22, 410–416. [Google Scholar] [CrossRef]
- Do Rego, J.L.; Seong, J.Y.; Burel, D.; Leprince, J.; Luu-The, V.; Tsutsui, K.; Tonon, M.C.; Pelletier, G.; Vaudry, H. Neurosteroid Biosynthesis: Enzymatic Pathways and Neuroendocrine Regulation by Neurotransmitters and Neuropeptides. Front. Neuroendocrinol. 2009, 30, 259–301. [Google Scholar] [CrossRef]
- Reddy, D.S. Neurosteroids: Endogenous Role in the Human Brain and Therapeutic Potentials. Prog. Brain Res. 2010, 186, 113–137. [Google Scholar] [CrossRef]
- Wilkenfeld, S.R.; Lin, C.; Frigo, D.E. Communication between Genomic and Non-Genomic Signaling Events Coordinate Steroid Hormone Actions. Steroids 2018, 133, 2–7. [Google Scholar] [CrossRef]
- Zhao, X.F. G Protein-Coupled Receptors Function as Cell Membrane Receptors for the Steroid Hormone 20-Hydroxyecdysone. Cell Commun. Signal. 2020, 18, 1–9. [Google Scholar] [CrossRef]
- Baulieu, E.; Robel, P. Neurosteroids: A New Brain Function? J. Steroid Biochem. Mol. Biol. 1990, 37, 395–403. [Google Scholar] [CrossRef]
- Colciago, A.; Bonalume, V.; Melfi, V.; Magnaghi, V. Genomic and Non-Genomic Action of Neurosteroids in the Peripheral Nervous System. Front. Neurosci. 2020, 14, 796. [Google Scholar] [CrossRef]
- Daniel, J.M.; Hulst, J.L.; Lee, C.D. Role of Hippocampal M2 Muscarinic Receptors in the Estrogen-Induced Enhancement of Working Memory. Neuroscience 2005, 132, 57–64. [Google Scholar] [CrossRef]
- Darnaudéry, M.; Koehl, M.; Piazza, P.V.; Le Moal, M.; Mayo, W. Pregnenolone Sulfate Increases Hippocampal Acetylcholine Release and Spatial Recognition. Brain Res. 2000, 852, 173–179. [Google Scholar] [CrossRef]
- Horishita, T.; Minami, K.; Uezono, Y.; Shiraishi, M.; Ogata, J.; Okamoto, T.; Terada, T.; Sata, T. The Effects of the Neurosteroids: Pregnenolone, Progesterone and Dehydroepiandrosterone on Muscarinic Receptor-Induced Responses in Xenopus Oocytes Expressing M1 and M3 Receptors. Naunyn. Schmiedebergs. Arch. Pharmacol. 2005, 371, 221–228. [Google Scholar] [CrossRef]
- Klangkalya, B.; Chan, A. The Effects of Ovarian Hormones on Beta-Adrenergic and Muscarinic Receptors in Rat Heart. Life Sci. 1988, 42, 2307–2314. [Google Scholar] [CrossRef]
- Klangkalya, B.; Chan, A. Structure-Activity Relationships of Steroid Hormones on Muscarinic Receptor Binding. J. Steroid Biochem. 1988, 29, 111–118. [Google Scholar] [CrossRef]
- Klangkalya, B.; Chan, A. Inhibition of Hypothalamic and Pituitary Muscarinic Receptor Binding by Progesterone. Neuroendocrinology 1988, 47, 294–302. [Google Scholar] [CrossRef] [PubMed]
- Dolejší, E.; Szánti-Pintér, E.; Chetverikov, N.; Nelic, D.; Randáková, A.; Doležal, V.; Kudová, E.; Jakubík, J. Neurosteroids and Steroid Hormones Are Allosteric Modulators of Muscarinic Receptors. Neuropharmacology 2021, 199, 108798. [Google Scholar] [CrossRef]
- Dolejší, E.; Chetverikov, N.; Szánti-Pintér, E.; Nelic, D.; Randáková, A.; Doležal, V.; El-Fakahany, E.E.; Kudová, E.; Jakubík, J. Neuroactive Steroids, WIN-Compounds and Cholesterol Share a Common Binding Site on Muscarinic Acetylcholine Receptors. Biochem. Pharmacol. 2021, 192, 114699. [Google Scholar] [CrossRef]
- Fantini, J.; Barrantes, F.J. How Cholesterol Interacts with Membrane Proteins: An Exploration of Cholesterol-Binding Sites Including CRAC, CARC, and Tilted Domains. Front. Physiol. 2013, 4, 31. [Google Scholar] [CrossRef]
- Bandara, A.; Panahi, A.; Pantelopulos, G.A.; Straub, J.E. Exploring the Structure and Stability of Cholesterol Dimer Formation in Multicomponent Lipid Bilayers. J. Comput. Chem. 2017, 38, 1479–1488. [Google Scholar] [CrossRef]
- Simons, K.; Toomre, D. Lipid Rafts and Signal Transduction. Nat. Rev. Mol. Cell Biol. 2000, 1, 31–39. [Google Scholar] [CrossRef]
- Lei, B.; Morris, D.P.; Smith, M.P.; Schwinn, D.A. Lipid Rafts Constrain Basal A1A-Adrenergic Receptor Signaling by Maintaining Receptor in an Inactive Conformation. Cell. Signal. 2009, 21, 1532–1539. [Google Scholar] [CrossRef]
- Niemelä, P.S.; Ollila, S.; Hyvönen, M.T.; Karttunen, M.; Vattulainen, I. Assessing the Nature of Lipid Raft Membranes. PLoS Comput. Biol. 2007, 3, 304–312. [Google Scholar] [CrossRef]
- Levitan, I.; Fang, Y.; Rosenhouse-Dantsker, A.; Romanenko, V. Cholesterol and Ion Channels. Subcell. Biochem. 2010, 51, 509. [Google Scholar] [CrossRef] [PubMed]
- Duncan, A.L.; Song, W.; Sansom, M.S.P. Lipid-Dependent Regulation of Ion Channels and G Protein–Coupled Receptors: Insights from Structures and Simulations. Annu. Rev. Pharmacol. Toxicol. 2020, 60, 31–50. [Google Scholar] [CrossRef] [PubMed]
- Gimpl, G.; Burger, K.; Fahrenholz, F. A Closer Look at the Cholesterol Sensor. Trends Biochem. Sci. 2002, 27, 596–599. [Google Scholar] [CrossRef]
- Hanson, M.A.; Cherezov, V.; Griffith, M.T.; Roth, C.B.; Jaakola, V.-P.; Chien, E.Y.T.; Velasquez, J.; Kuhn, P.; Stevens, R.C. A Specific Cholesterol Binding Site Is Established by the 2.8 A Structure of the Human Beta2-Adrenergic Receptor. Structure 2008, 16, 897–905. [Google Scholar] [CrossRef]
- Paila, Y.D.; Tiwari, S.; Chattopadhyay, A. Are Specific Nonannular Cholesterol Binding Sites Present in G-Protein Coupled Receptors? Biochim. Biophys. Acta Biomembr. 2009, 1788, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Gimpl, G. Interaction of G Protein Coupled Receptors and Cholesterol. Chem. Phys. Lipids 2016, 199, 61–73. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, P.; Chattopadhyay, A. Cholesterol Interaction Motifs in G Protein-Coupled Receptors: Slippery Hot Spots? Wiley Interdiscip. Rev. Syst. Biol. Med. 2020, 12, e1481. [Google Scholar] [CrossRef]
- Reddy, D.S.; Estes, W.A. Clinical Potential of Neurosteroids for CNS Disorders. Trends Pharmacol. Sci. 2016, 37, 543–561. [Google Scholar] [CrossRef]
- Ratner, M.H.; Kumaresan, V.; Farb, D.H. Neurosteroid Actions in Memory and Neurologic/Neuropsychiatric Disorders. Front. Endocrinol. 2019, 10, 169. [Google Scholar] [CrossRef]
- Coronel, M.F.; Labombarda, F.; González, S.L. Neuroactive Steroids, Nociception and Neuropathic Pain: A Flashback to Go Forward. Steroids 2016, 110, 77–87. [Google Scholar] [CrossRef]
- Joksimovic, S.L.; Covey, D.F.; Jevtovic-Todorovic, V.; Todorovic, S.M. Neurosteroids in Pain Management: A New Perspective on an Old Player. Front. Pharmacol. 2018, 9, 1127. [Google Scholar] [CrossRef] [PubMed]
- Meyer, L.; Taleb, O.; Patte-Mensah, C.; Mensah-Nyagan, A.-G. Neurosteroids and Neuropathic Pain Management: Basic Evidence and Therapeutic Perspectives. Front. Neuroendocrinol. 2019, 55, 100795. [Google Scholar] [CrossRef] [PubMed]
- González, S.L.; Meyer, L.; Raggio, M.C.; Taleb, O.; Coronel, M.F.; Patte-Mensah, C.; Mensah-Nyagan, A.G. Allopregnanolone and Progesterone in Experimental Neuropathic Pain: Former and New Insights with a Translational Perspective. Cell. Mol. Neurobiol. 2019, 39, 523–537. [Google Scholar] [CrossRef] [PubMed]
- Borowicz, K.K.; Piskorska, B.; Banach, M.; Czuczwar, S.J. Neuroprotective Actions of Neurosteroids. Front. Endocrinol. 2011, 2, 50. [Google Scholar] [CrossRef]
- Mendell, A.L.; MacLusky, N.J. Neurosteroid Metabolites of Gonadal Steroid Hormones in Neuroprotection: Implications for Sex Differences in Neurodegenerative Disease. Front. Mol. Neurosci. 2018, 11, 359. [Google Scholar] [CrossRef]
- Yilmaz, C.; Karali, K.; Fodelianaki, G.; Gravanis, A.; Chavakis, T.; Charalampopoulos, I.; Alexaki, V.I. Neurosteroids as Regulators of Neuroinflammation. Front. Neuroendocrinol. 2019, 55, 100788. [Google Scholar] [CrossRef]
- Kudova, E. Rapid Effects of Neurosteroids on Neuronal Plasticity and Their Physiological and Pathological Implications. Neurosci. Lett. 2021, 750, 135771. [Google Scholar] [CrossRef]
- Klinge, C.M. Steroid Hormone Receptors and Signal Transduction Processes. In Principles of Endocrinology and Hormone Action; Springer: Cham, Switzerland, 2018; pp. 187–232. [Google Scholar]
- Reddy, D.S. Catamenial Epilepsy: Discovery of an Extrasynaptic Molecular Mechanism for Targeted Therapy. Front. Cell. Neurosci. 2016, 10, 101. [Google Scholar] [CrossRef]
- Baulieu, E.E. Neurosteroids: A Novel Function of the Brain. Psychoneuroendocrinology 1998, 23, 963–987. [Google Scholar] [CrossRef]
- Reddy, D.S. Mass Spectrometric Assay and Physiological-Pharmacological Activity of Androgenic Neurosteroids. Neurochem. Int. 2008, 52, 541–553. [Google Scholar] [CrossRef]
- Reddy, D.S. Role of Hormones and Neurosteroids in Epileptogenesis. Front. Cell. Neurosci. 2013, 7, 115. [Google Scholar] [CrossRef] [PubMed]
- Reddy, D.S. Neurosteroids. In Progress in Brain Research; Elsevier: Amsterdam, The Netherlands, 2010; Volume 186, pp. 113–137. ISBN 9780444536303. [Google Scholar]
- Baulieu, E.E.; Schumacher, M. Progesterone as a Neuroactive Neurosteroid, with Special Reference to the Effect of Progesterone on Myelination. Hum. Reprod. 2000, 15 (Suppl. 1), 1–13. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Almey, A.; Milner, T.A.; Brake, W.G. Estrogen Receptors in the Central Nervous System and Their Implication for Dopamine-Dependent Cognition in Females. Horm. Behav. 2015, 74, 125–138. [Google Scholar] [CrossRef] [PubMed]
- Pang, Z.P.; Han, W. Regulation of Synaptic Functions in Central Nervous System by Endocrine Hormones and the Maintenance of Energy Homoeostasis. Biosci. Rep. 2012, 32, 423–432. [Google Scholar] [CrossRef]
- Karpinski, M.; Mattina, G.F.; Steiner, M. Effect of Gonadal Hormones on Neurotransmitters Implicated in the Pathophysiology of Obsessive-Compulsive Disorder: A Critical Review. Neuroendocrinology 2017, 105, 1–16. [Google Scholar] [CrossRef]
- Barth, C.; Villringer, A.; Sacher, J. Sex Hormones Affect Neurotransmitters and Shape the Adult Female Brain during Hormonal Transition Periods. Front. Neurosci. 2015, 9, 1–20. [Google Scholar] [CrossRef]
- Rudolph, L.M.; Cornil, C.A.; Mittelman-Smith, M.A.; Rainville, J.R.; Remage-Healey, L.; Sinchak, K.; Micevych, P.E. Actions of Steroids: New Neurotransmitters. J. Neurosci. 2016, 36, 11449–11458. [Google Scholar] [CrossRef]
- Belanoff, J.K.; Gross, K.; Yager, A.; Schatzberg, A.F. Corticosteroids and Cognition. J. Psychiatr. Res. 2001, 35, 127–145. [Google Scholar] [CrossRef]
- Wolf, O.T. Cognitive Functions and Sex Steroids. Ann. Endocrinol. 2003, 64, 158–161. [Google Scholar]
- Ali, S.A.; Begum, T.; Reza, F. Hormonal Influences on Cognitive Function. Malaysian J. Med. Sci. 2018, 25, 31–41. [Google Scholar] [CrossRef]
- Frick, K.M.; Kim, J. Mechanisms Underlying the Rapid Effects of Estradiol and Progesterone on Hippocampal Memory Consolidation in Female Rodents. Horm. Behav. 2018, 104, 100–110. [Google Scholar] [CrossRef] [PubMed]
- Ouanes, S.; Popp, J. High Cortisol and the Risk of Dementia and Alzheimer’s Disease: A Review of the Literature. Front. Aging Neurosci. 2019, 11, 43. [Google Scholar] [CrossRef] [PubMed]
- McEwen, B.S. Steroid Hormones: Effect on Brain Development and Function. Horm. Res. 1992, 37, 1–10. [Google Scholar] [CrossRef]
- Rubinow, D.R.; Schmidt, P.J. Gonadal Steroids, Brain, and Behavior: Role of Context. Dialogues Clin. Neurosci. 2002, 4, 123–137. [Google Scholar] [CrossRef]
- Thomas, P.; Pang, Y. Membrane Progesterone Receptors: Evidence for Neuroprotective, Neurosteroid Signaling and Neuroendocrine Functions in Neuronal Cells. Neuroendocrinology 2012, 96, 162–171. [Google Scholar] [CrossRef]
- Thomas, P.; Pang, Y. Anti-Apoptotic Actions of Allopregnanolone and Ganaxolone Mediated Through Membrane Progesterone Receptors (PAQRs) in Neuronal Cells. Front. Endocrinol. 2020, 11, 417. [Google Scholar] [CrossRef] [PubMed]
- Webb, S.J.; Geoghegan, T.E.; Prough, R.A.; Michael Miller, K.K. The Biological Actions of Dehydroepiandrosterone Involves Multiple Receptors. Drug Metab. Rev. 2006, 38, 89–116. [Google Scholar] [CrossRef]
- Zheng, P. Neuroactive Steroid Regulation of Neurotransmitter Release in the CNS: Action, Mechanism and Possible Significance. Prog. Neurobiol. 2009, 89, 134–152. [Google Scholar] [CrossRef]
- Yadid, G.; Sudai, E.; Maayan, R.; Gispan, I.; Weizman, A. The Role of Dehydroepiandrosterone (DHEA) in Drug-Seeking Behavior. Neurosci. Biobehav. Rev. 2010, 35, 303–314. [Google Scholar] [CrossRef]
- Lösel, R.; Wehling, M. Nongenomic Actions of Steroid Hormones. Nat. Rev. Mol. Cell Biol. 2003, 4, 46–56. [Google Scholar] [CrossRef]
- Tuem, K.B.; Atey, T.M. Neuroactive Steroids: Receptor Interactions and Responses. Front. Neurol. 2017, 8, 442. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Liu, Y.; Cao, J.-M. G Protein-Coupled Receptors: Extranuclear Mediators for the Non-Genomic Actions of Steroids. Int. J. Mol. Sci. 2014, 15, 15412–15425. [Google Scholar] [CrossRef] [PubMed]
- Rosenbaum, D.M.; Rasmussen, S.G.F.; Kobilka, B.K. The Structure and Function of G-Protein-Coupled Receptors. Nature 2009, 459, 356–363. [Google Scholar] [CrossRef] [PubMed]
- Hauser, A.S.; Attwood, M.M.; Rask-Andersen, M.; Schiöth, H.B.; Gloriam, D.E. Trends in GPCR Drug Discovery: New Agents, Targets and Indications. Nat. Rev. Drug Discov. 2017, 16, 829–842. [Google Scholar] [CrossRef] [PubMed]
- Dascal, N.; Kahanovitch, U. The Roles of Gβγ and Gα in Gating and Regulation of GIRK Channels. Int. Rev. Neurobiol. 2015, 123, 27–85. [Google Scholar] [CrossRef] [PubMed]
- Randáková, A.; Nelic, D.; Ungerová, D.; Nwokoye, P.; Su, Q.; Doležal, V.; El-Fakahany, E.E.; Boulos, J.; Jakubík, J. Novel M 2 -selective, G i -biased Agonists of Muscarinic Acetylcholine Receptors. Br. J. Pharmacol. 2020, 177, 2073–2089. [Google Scholar] [CrossRef]
- Randáková, A.; Jakubík, J. Functionally Selective and Biased Agonists of Muscarinic Receptors. Pharmacol. Res. 2021, 169, 105641. [Google Scholar] [CrossRef]
- Li, H.; Papadopoulos, V. Peripheral-Type Benzodiazepine Receptor Function in Cholesterol Transport. Identification of a Putative Cholesterol Recognition/Interaction Amino Acid Sequence and Consensus Pattern. Endocrinology 1998, 139, 4991–4997. [Google Scholar] [CrossRef]
- Jafurulla, M.; Tiwari, S.; Chattopadhyay, A. Identification of Cholesterol Recognition Amino Acid Consensus (CRAC) Motif in G-Protein Coupled Receptors. Biochem. Biophys. Res. Commun. 2011, 404, 569–573. [Google Scholar] [CrossRef]
- Bymaster, F.P.; Carter, P.A.; Peters, S.C.; Zhang, W.; Ward, J.S.; Mitch, C.H.; Calligaro, D.O.; Whitesitt, C.A.; DeLapp, N.; Shannon, H.E.; et al. Xanomeline Compared to Other Muscarinic Agents on Stimulation of Phosphoinositide Hydrolysis in Vivo and Other Cholinomimetic Effects. Brain Res 1998, 795, 179–190. [Google Scholar] [CrossRef]
- DeLapp, N.; Wu, S.; Belagaje, R.; Johnstone, E.; Little, S.; Shannon, H.; Bymaster, F.; Calligaro, D.; Mitch, C.; Whitesitt, C.; et al. Effects of the M1 Agonist Xanomeline on Processing of Human Beta-Amyloid Precursor Protein (FAD, Swedish Mutant) Transfected into Chinese Hamster Ovary-M1 Cells. Biochem. Biophys. Res. Commun. 1998, 244, 156–160. [Google Scholar] [CrossRef] [PubMed]
- Christopoulos, A.; Pierce, T.L.; Sorman, J.L.; El-Fakahany, E.E. On the Unique Binding and Activating Properties of Xanomeline at the M1 Muscarinic Acetylcholine Receptor. Mol. Pharmacol. 1998, 53, 1120–1130. [Google Scholar] [PubMed]
- Grant, M.K.O.; El-Fakahany, E.E. Persistent Binding and Functional Antagonism by Xanomeline at the Muscarinic M5 Receptor. J. Pharmacol. Exp. Ther. 2005, 315, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Avissar, S.; Egozi, Y.; Sokolovsky, M. Studies on Muscarinic Receptors in Mouse and Rat Hypothalamus: A Comparison of Sex and Cyclical Differences. Neuroendocrinology 1981, 32, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, M.; Giles, A.; Wilkinson, D.A. M 2 Muscarinic ([3 H] N -Methyl Scopolamine) Binding in Micropunches of Rat Ventricular Myocardium: Characterization and Modification by Progesterone. Can. J. Physiol. Pharmacol. 1992, 70, 943–948. [Google Scholar] [CrossRef]
- Wilkinson, M.; Siauw, M.; Horackova, M. Modulation of cardiac M2 muscarinic receptor binding by progesterone-related steroids. J. Mol. Cell. Cardiol. 1995, 27, 1831–1839. [Google Scholar] [CrossRef]
- Shiraishi, M.; Minami, K.; Shibuya, I.; Uezono, Y.; Ogata, J.; Okamoto, T.; Murasaki, O.; Kaibara, M.; Ueta, Y.; Shigematsu, A. The Inhibitory Effects of Alphaxalone on M1 and M3 Muscarinic Receptors Expressed in Xenopus Oocytes. Anesth. Analg. 2003, 97, 449–455. [Google Scholar] [CrossRef]
- Sokolovsky, M.; Egozi, Y.; Avissar, S. Molecular Regulation of Receptors: Interaction of Beta-Estradiol and Progesterone with the Muscarinic System. Proc. Natl. Acad. Sci. USA 1981, 78, 5554–5558. [Google Scholar] [CrossRef]
- Al-Daham, M.I.M.; Thomas, P.J. Contrasting Effects of Testicular and Ovarian Steroids upon Muscarinic Binding Sites in the Brain. Pharmacology 1987, 34, 250–258. [Google Scholar] [CrossRef]
- Bae, Y.J.; Zeidler, R.; Baber, R.; Vogel, M.; Wirkner, K.; Loeffler, M.; Ceglarek, U.; Kiess, W.; Körner, A.; Thiery, J.; et al. Reference Intervals of Nine Steroid Hormones over the Life-Span Analyzed by LC-MS/MS: Effect of Age, Gender, Puberty, and Oral Contraceptives. J. Steroid Biochem. Mol. Biol. 2019, 193, 105409. [Google Scholar] [CrossRef]
- Hill, M.; Hána, V.; Velíková, M.; Pařízek, A.; Kolátorová, L.; Vítků, J.; Škodová, T.; Šimková, M.; Šimják, P.; Kancheva, R.; et al. A Method for Determination of One Hundred Endogenous Steroids in Human Serum by Gas Chromatography-Tandem Mass Spectrometry. Physiol. Res. 2019, 68, 179–207. [Google Scholar] [CrossRef] [PubMed]
- Lazareno, S.; Popham, A.; Birdsall, N.J. Allosteric Interactions of Staurosporine and Other Indolocarbazoles with N-[Methyl-(3)H]Scopolamine and Acetylcholine at Muscarinic Receptor Subtypes: Identification of a Second Allosteric Site. Mol. Pharmacol. 2000, 58, 194–207. [Google Scholar] [CrossRef] [PubMed]
- Lazareno, S.; Popham, A.; Birdsall, N.J.M. Analogs of WIN 62,577 Define a Second Allosteric Site on Muscarinic Receptors. Mol. Pharmacol. 2002, 62, 1492–1505. [Google Scholar] [CrossRef] [PubMed]
- Kruse, A.C.; Ring, A.M.; Manglik, A.; Hu, J.; Hu, K.; Eitel, K.; Hübner, H.; Pardon, E.; Valant, C.; Sexton, P.M.; et al. Activation and Allosteric Modulation of a Muscarinic Acetylcholine Receptor. Nature 2013, 504, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.-P.; Prilla, S.; Mohr, K.; Ellis, J. Critical Amino Acid Residues of the Common Allosteric Site on the M2 Muscarinic Acetylcholine Receptor: More Similarities than Differences between the Structurally Divergent Agents Gallamine and Bis(Ammonio)Alkane-Type Hexamethylene-Bis-[Dimethyl-(3-Phthalimidopropyl)ammonium]dibromide. Mol. Pharmacol. 2005, 68, 769–778. [Google Scholar] [CrossRef]
- Leppik, R.A.; Miller, R.C.; Eck, M.; Paquet, J.L. Role of Acidic Amino Acids in the Allosteric Modulation by Gallamine of Antagonist Binding at the M2 Muscarinic Acetylcholine Receptor. Mol. Pharmacol. 1994, 45, 983–990. [Google Scholar]
- Jakubík, J.; El-Fakahany, E.E. Allosteric Modulation of GPCRs of Class A by Cholesterol. Int. J. Mol. Sci. 2021, 22, 1953. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).