
Application of Advanced Non-Linear Spectral Decomposition and Regression Methods for Spectroscopic Analysis of Targeted and Non-Targeted Irradiation Effects in an In-Vitro Model


 ,
, 
Abstract
1. Introduction
2. Results
2.1. Visualisation of Spectra
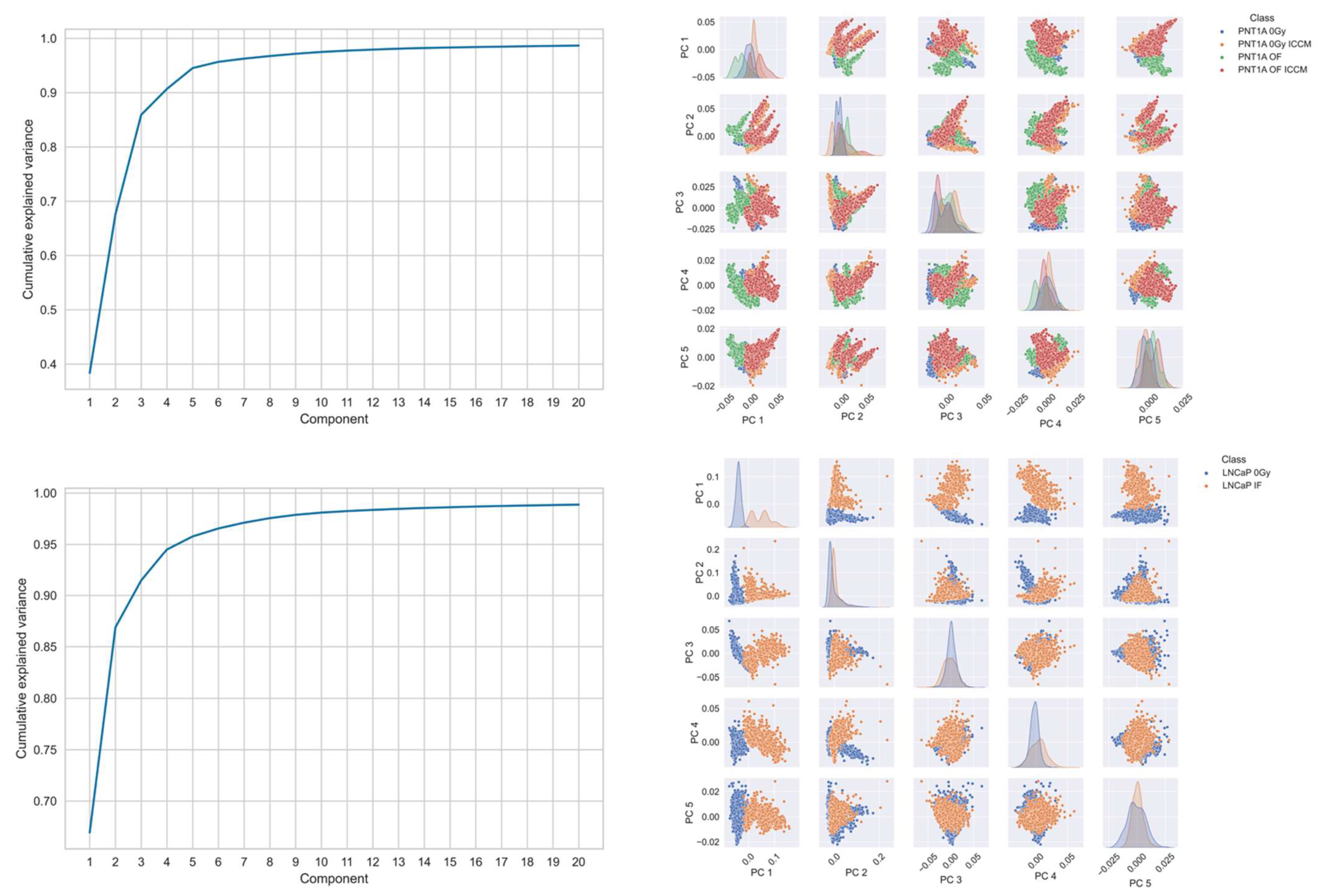
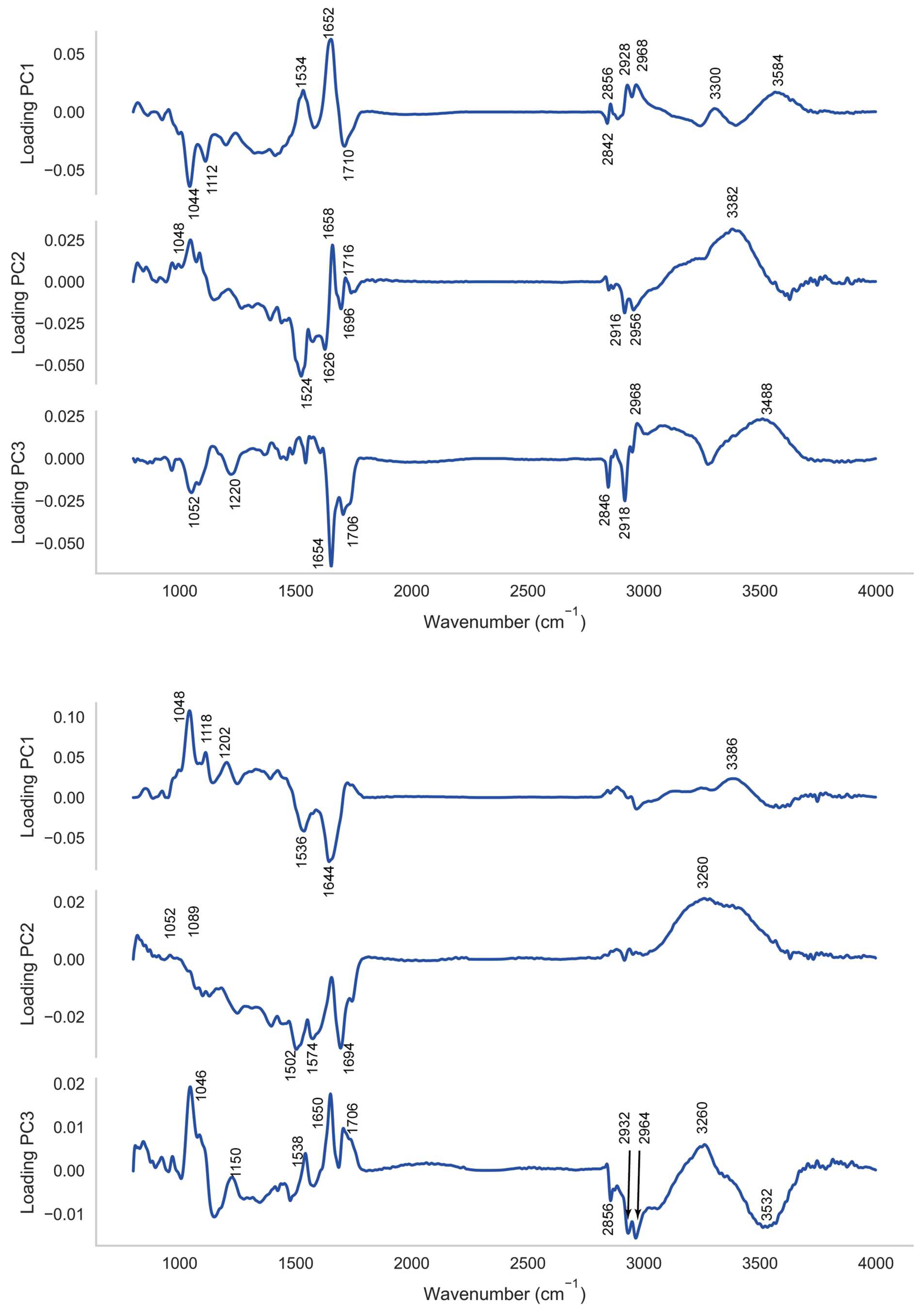
2.2. PCA-SVM Modelling
2.3. CLS Spectral Fitting
2.4. XGBoost Regression against Cell Volume
3. Discussion
- A reduction in colony volume between the PNT1A cells exposed to 0.2 Gy (PNT1A OF) and in unirradiated cells exposed to secreted factors from the tumour cells (PNT1A 0 Gy ICCM) when compared to cells irradiated with a low dose prior to exposure to secreted factors from the tumour (PNT1A OF ICCM);
- An increase in DNA double strand break (DSB) damage foci (γH2AX fluorescence measured via confocal microscopy) for all exposure modes, with the exposure of PNT1A cells to both a dose of 0.2 Gy and secreted factors from the tumour cells (PNT1A OF ICCM) producing a statistically significant increase in damage relative to the other exposure modes.
4. Materials and Methods
4.1. Sample Preparation and Characteristics
4.1.1. Cell Culture, Irradiation, Exposure to Irradiated Cell Culture Medium (ICCM) and Parallel Biological Assays
4.1.2. FTIR Spectral Acquisition and Pre-Processing
4.2. Chemometrics and Machine Learning
4.2.1. Principal Components Analysis
4.2.2. Support Vector Machine
4.2.3. t-SNE
4.2.4. Extreme-Gradient-Boosted Regression (XGBR)
4.2.5. Classic Least Squares Spectral Fitting (CLS)
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rawla, P. Epidemiology of Prostate Cancer. World J. Oncol. 2019, 10, 63–89. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef]
- Litwin, M.S.; Tan, H.-J. The Diagnosis and Treatment of Prostate Cancer. JAMA 2017, 317, 2532–2542. [Google Scholar] [CrossRef] [PubMed]
- Suchowerska, N.; Ebert, M.A.; Zhang, M.; Jackson, M. In vitroresponse of tumour cells to non-uniform irradiation. Phys. Med. Biol. 2005, 50, 3041–3051. [Google Scholar] [CrossRef] [PubMed]
- Shields, L.; Vega-Carrascal, I.; Singleton, S.; Lyng, F.L.; McClean, B. Cell Survival and DNA Damage in Normal Prostate Cells Irradiated Out-of-Field. Radiat. Res. 2014, 182, 499–506. [Google Scholar] [CrossRef]
- Prise, K.M.; O’Sullivan, J.M. Radiation-induced bystander signalling in cancer therapy. Nat. Cancer 2009, 9, 351–360. [Google Scholar] [CrossRef]
- Daguenet, E.; Louati, S.; Wozny, A.-S.; Vial, N.; Gras, M.; Guy, J.-B.; Vallard, A.; Rodriguez-Lafrasse, C.; Magné, N. Radiation-induced bystander and abscopal effects: Important lessons from preclinical models. Br. J. Cancer 2020, 123, 339–348. [Google Scholar] [CrossRef]
- Butler, H.J.; Brennan, P.M.; Cameron, J.M.; Finlayson, D.; Hegarty, M.G.; Jenkinson, M.D.; Palmer, D.S.; Smith, B.R.; Baker, M.J. Development of high-throughput ATR-FTIR technology for rapid triage of brain cancer. Nat. Commun. 2019, 10, 4501. [Google Scholar] [CrossRef]
- Mitchell, A.L.; Gajjar, K.B.; Theophilou, G.; Martin, F.L.; Martin-Hirsch, P.L. Vibrational spectroscopy of biofluids for disease screening or diagnosis: Translation from the laboratory to a clinical setting. J. Biophotonics 2014, 7, 153–165. [Google Scholar] [CrossRef]
- Medipally, D.K.R.; Cullen, D.; Untereiner, V.; Sockalingum, G.D.; Maguire, A.; Nguyen, T.N.Q.; Bryant, J.; Noone, E.; Bradshaw, S.; Finn, M.; et al. Vibrational spectroscopy of liquid biopsies for prostate cancer diagnosis. Ther. Adv. Med. Oncol. 2020, 12, 1758835920918499. [Google Scholar] [CrossRef]
- Harder, S.J.; Matthews, Q.; Isabelle, M.; Brolo, A.G.; Lum, J.J.; Jirasek, A. A Raman Spectroscopic Study of Cell Response to Clinical Doses of Ionizing Radiation. Appl. Spectrosc. 2015, 69, 193–204. [Google Scholar] [CrossRef] [PubMed]
- Matthews, Q.; Jirasek, A.; Lum, J.J.; Brolo, A.G. Biochemical signatures of in vitro radiation response in human lung, breast and prostate tumour cells observed with Raman spectroscopy. Phys. Med. Biol. 2011, 56, 6839–6855. [Google Scholar] [CrossRef] [PubMed]
- Delfino, I.; Perna, G.; Lasalvia, M.; Capozzi, V.; Manti, L.; Camerlingo, C.; Lepore, M. Visible micro-Raman spectroscopy of single human mammary epithelial cells exposed to x-ray radiation. J. Biomed. Opt. 2015, 20, 035003. [Google Scholar] [CrossRef] [PubMed]
- Delfino, I.; Perna, G.; Ricciardi, V.; Lasalvia, M.; Manti, L.; Capozzi, V.; Lepore, M. X-ray irradiation effects on nuclear and membrane regions of single SH-SY5Y human neuroblastoma cells investigated by Raman micro-spectroscopy. J. Pharm. Biomed. Anal. 2019, 164, 557–573. [Google Scholar] [CrossRef] [PubMed]
- Roman, M.; Wrobel, T.P.; Panek, A.; Paluszkiewicz, C.; Kwiatek, W.M. Exploring subcellular responses of prostate cancer cells to clinical doses of X-rays by Raman microspectroscopy. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2021, 255, 119653. [Google Scholar] [CrossRef]
- Meade, A.D.; Clarke, C.; Byrne, H.J.; Lyng, F.M. Fourier Transform Infrared Microspectroscopy and Multivariate Methods for Radiobiological Dosimetry. Radiat. Res. 2010, 173, 225–237. [Google Scholar] [CrossRef]
- Ainsbury, E.; Badie, C.; Barnard, S.; Manning, G.; Moquet, J.; Abend, M.; Antunes, A.C.; Barrios, L.; Bassinet, C.; Beinke, C.; et al. Integration of new biological and physical retrospective dosimetry methods into EU emergency response plans—Joint RENEB and EURADOS inter-laboratory comparisons. Int. J. Radiat. Biol. 2016, 93, 99–109. [Google Scholar] [CrossRef]
- Meade, A.D.; Howe, O.; Unterreiner, V.; Sockalingum, G.D.; Byrne, H.J.; Lyng, F.M. Vibrational spectroscopy in sensing radiobiological effects: Analyses of targeted and non-targeted effects in human keratinocytes. Faraday Discuss. 2016, 187, 213–234. [Google Scholar] [CrossRef]
- Lipiec, E.; Bambery, K.; Lekki, J.; Tobin, M.J.; Vogel, C.; Whelan, D.; Wood, B.R.; Kwiatek, W. SR-FTIR Coupled with Principal Component Analysis Shows Evidence for the Cellular Bystander Effect. Radiat. Res. 2015, 184, 73–82. [Google Scholar] [CrossRef]
- Harder, S.J.; Isabelle, M.; DeVorkin, L.; Smazynski, J.; Beckham, W.; Brolo, A.G.; Lum, J.J.; Jirasek, A. Raman spectroscopy identifies radiation response in human non-small cell lung cancer xenografts. Sci. Rep. 2016, 6, 21006. [Google Scholar] [CrossRef]
- Van Nest, S.J.; Nicholson, L.M.; DeVorkin, L.; Brolo, A.G.; Lum, J.J.; Jirasek, A. Raman Spectroscopic Signatures Reveal Distinct Biochemical and Temporal Changes in Irradiated Human Breast Adenocarcinoma Xenografts. Radiat. Res. 2018, 189, 497–504. [Google Scholar] [CrossRef] [PubMed]
- Maguire, A.; Vegacarrascal, I.; White, L.; McClean, B.; Howe, O.; Lyng, F.M.; Meade, A.D. Analyses of Ionizing Radiation EffectsIn Vitroin Peripheral Blood Lymphocytes with Raman Spectroscopy. Radiat. Res. 2015, 183, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Meade, A.D.; Maguire, A.; Bryant, J.; Cullen, D.; Medipally, D.; White, L.; McClean, B.; Shields, L.; Armstrong, J.; Dunne, M.; et al. Prediction of DNA damage and G2 chromosomal radio-sensitivity ex vivo in peripheral blood mononuclear cells with label-free Raman micro-spectroscopy. Int. J. Radiat. Biol. 2018, 95, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.N.Q.; Maguire, A.; Mooney, C.; Jackson, N.; Lynam-Lennon, N.; Weldon, V.; Muldoon, C.; Maguire, A.A.; O’Toole, D.; Ravi, N.; et al. Prediction of pathological response to neo-adjuvant chemoradiotherapy for oesophageal cancer using vibrational spectroscopy. Transl. Biophotonics 2020, 3, e202000014. [Google Scholar] [CrossRef]
- Medipally, D.K.; Nguyen, T.N.Q.; Bryant, J.; Untereiner, V.; Sockalingum, G.D.; Cullen, D.; Noone, E.; Bradshaw, S.; Finn, M.; Dunne, M.; et al. Monitoring Radiotherapeutic Response in Prostate Cancer Patients Using High Throughput FTIR Spectroscopy of Liquid Biopsies. Cancers 2019, 11, 925. [Google Scholar] [CrossRef] [PubMed]
- Cullen, D.; Bryant, J.; Maguire, A.; Medipally, D.; McClean, B.; Shields, L.; Noone, E.; Bradshaw, S.; Finn, M.; Dunne, M.; et al. Raman spectroscopy of lymphocytes for the identification of prostate cancer patients with late radiation toxicity following radiotherapy. Transl. Biophotonics 2020, 2, e201900035. [Google Scholar] [CrossRef]
- Wang, R.; Zhou, T.; Liu, W.; Zuo, L. Molecular mechanism of bystander effects and related abscopal/cohort effects in cancer therapy. Oncotarget 2018, 9, 18637–18647. [Google Scholar] [CrossRef] [PubMed]
- Sun, R.; Sbai, A.; Ganem, G.; Boudabous, M.; Collin, F.; Marcy, P.-Y.; Doglio, A.; Thariat, J. Effets non ciblés (bystander, abscopal) de la radiothérapie externe: Potentielles implications pour le clinicien? Cancer/Radiothérapie 2014, 18, 770–778. [Google Scholar] [CrossRef]
- Pouget, J.-P.; Georgakilas, A.G.; Ravanat, J.-L. Targeted and Off-Target (Bystander and Abscopal) Effects of Radiation Therapy: Redox Mechanisms and Risk/Benefit Analysis. Antioxid. Redox Signal. 2018, 29, 1447–1487. [Google Scholar] [CrossRef]
- Butterworth, K.T.; Ghita, M.; McMahon, S.J.; McGarry, C.K.; Griffin, R.J.; Hounsell, A.R.; Prise, K.M. Modelling responses to spatially fractionated radiation fields using preclinical image-guided radiotherapy. Br. J. Radiol. 2017, 90, 20160485. [Google Scholar] [CrossRef]
- Welch, B.L. On the Comparison of Several Mean Values: An Alternative Approach. Biometrika 1951, 38, 330–336. [Google Scholar] [CrossRef]
- Games, P.A.; Howell, J.F. Pairwise Multiple Comparison Procedures with Unequal N’s and/or Variances: A Monte Carlo Study. J. Educ. Stat. 1976, 1, 113. [Google Scholar] [CrossRef]
- Meade, A.D.; Lyng, F.M.; Knief, P.; Byrne, H.J. Growth substrate induced functional changes elucidated by FTIR and Raman spectroscopy in in–vitro cultured human keratinocytes. Anal. Bioanal. Chem. 2007, 387, 1717–1728. [Google Scholar] [CrossRef]
- Maguire, P.; Mothersill, C.; McClean, B.; Seymour, C.; Lyng, F.M. Modulation of Radiation Responses by Pre-exposure to Irradiated Cell Conditioned Medium. Radiat. Res. 2007, 167, 485–492. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Wu, L.; Chen, S.; Zhu, L.; Huang, P.; Tong, L.; Zhao, Y.; Zhao, G.; Wang, J.; Mei, T.; et al. Mitochondrial dysfunction resulting from loss of cytochrome c impairs radiation-induced bystander effect. Br. J. Cancer 2009, 100, 1912–1916. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Peixoto, P.M.; Ryu, S.-Y.; Pruzansky, D.P.; Kuriakose, M.; Gilmore, A.; Kinnally, K.W. Mitochondrial apoptosis is amplified through gap junctions. Biochem. Biophys. Res. Commun. 2009, 390, 38–43. [Google Scholar] [CrossRef]
- Hei, T.K.; Zhou, H.; Chai, Y.; Ponnaiya, B.; Ivanov, V.N. Radiation Induced Non-targeted Response: Mechanism and Potential Clinical Implications. Curr. Mol. Pharmacol. 2011, 4, 96–105. [Google Scholar] [CrossRef]
- Jiang, Y.; Chen, X.; Tian, W.; Yin, X.; Wang, J.; Yang, H. The role of TGF-β1–miR-21–ROS pathway in bystander responses induced by irradiated non-small-cell lung cancer cells. Br. J. Cancer 2014, 111, 772–780. [Google Scholar] [CrossRef]
- Temme, J.; Bauer, G. Low-Dose Gamma Irradiation Enhances Superoxide Anion Production by Nonirradiated Cells Through TGF-β1-Dependent Bystander Signaling. Radiat. Res. 2012, 179, 422–432. [Google Scholar] [CrossRef]
- Ivanov, V.N.; Zhou, H.; Ghandhi, S.A.; Karasic, T.B.; Yaghoubian, B.; Amundson, S.A.; Hei, T.K. Radiation-induced bystander signaling pathways in human fibroblasts: A role for interleukin-33 in the signal transmission. Cell. Signal. 2010, 22, 1076–1087. [Google Scholar] [CrossRef]
- Zhou, H.; Ivanov, V.N.; Gillespie, J.; Geard, C.R.; Amundson, S.A.; Brenner, D.J.; Yu, Z.; Lieberman, H.B.; Hei, T.K. Mechanism of radiation-induced bystander effect: Role of the cyclooxygenase-2 signaling pathway. Proc. Natl. Acad. Sci. USA 2005, 102, 14641–14646. [Google Scholar] [CrossRef] [PubMed]
- Lorimore, S.A.; McIlrath, J.M.; Coates, P.J.; Wright, E.G. Chromosomal Instability in Unirradiated Hemopoietic Cells Resulting from a Delayed In vivo Bystander Effect of γ Radiation. Cancer Res. 2005, 65, 5668–5673. [Google Scholar] [CrossRef] [PubMed]
- Lorimore, S.A.; Coates, P.J.; E Scobie, G.; Milne, G.; Wright, E.G. Inflammatory-type responses after exposure to ionizing radiation in vivo: A mechanism for radiation-induced bystander effects? Oncogene 2001, 20, 7085–7095. [Google Scholar] [CrossRef] [PubMed]
- Seymour, C.; Mothersill, C. Medium from irradiated human epithelial cells but not human fibroblasts reduces the clonogenic survival of unirradiated cells. Int. J. Radiat. Biol. 1997, 71, 421–427. [Google Scholar] [CrossRef]
- Troein, C.; Siregar, S.; De Beeck, M.O.; Peterson, C.; Tunlid, A.; Persson, P. OCTAVVS: A Graphical Toolbox for High-Throughput Preprocessing and Analysis of Vibrational Spectroscopy Imaging Data. Methods Protoc. 2020, 3, 34. [Google Scholar] [CrossRef]
- Bassan, P.; Kohler, A.; Martens, H.; Lee, J.; Jackson, E.; Lockyer, N.; Dumas, P.; Brown, M.; Clarke, N.; Gardner, P. RMieS-EMSC correction for infrared spectra of biological cells: Extension using full Mie theory and GPU computing. J. Biophotonics 2010, 3, 609–620. [Google Scholar] [CrossRef]
- Vallat, R. Pingouin: Statistics in Python. J. Open Source Softw. 2018, 3, 1026. [Google Scholar] [CrossRef]
- Ferguson, D.; Henderson, A.; McInnes, E.F.; Lind, R.; Wildenhain, J.; Gardner, P. Infrared micro-spectroscopy coupled with multivariate and machine learning techniques for cancer classification in tissue: A comparison of classification method, performance, and pre-processing technique. Analyst 2022, 147, 3709–3722. [Google Scholar] [CrossRef]
- Jolliffe, I.T.; Cadima, J. Principal component analysis: A review and recent developments. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 2016, 374, 20150202. [Google Scholar] [CrossRef]
- Cruz, J.A.; Wishart, D.S. Applications of Machine Learning in Cancer Prediction and Prognosis. Cancer Inform. 2007, 2, 59–77. [Google Scholar] [CrossRef]
- Gareth, J.; Daniela, W.; Trevor, H.; Robert, T. An Introduction to Statistical Learning: With Applications in R; Springer Science & Business Media: New York, NY, USA, 2013. [Google Scholar]
- Cristianini, N.; Shawe-Taylor, J. An Introduction to Support Vector Machines and Other Kernel-Based Learning Methods; Cambridge University Press: Cambridge, UK, 2000. [Google Scholar]
- Furey, T.; Cristianini, N.; Duffy, N.; Bednarski, D.W.; Schummer, M.; Haussler, D. Support vector machine classification and validation of cancer tissue samples using microarray expression data. Bioinformatics 2000, 16, 906–914. [Google Scholar] [CrossRef] [PubMed]
- Schölkopf, B.; Smola, A.J.; Bach, F. Learning with Kernels: Support Vector Machines, Regularization, Optimization, and Beyond Adaptive Computation and Machine Learning; MIT Press: Cambridge, MA, USA, 2002; p. 626. [Google Scholar]
- Van der Maaten, L.; Hinton, G. Visualizing Data using t-SNE. J. Mach. Learn. Res. 2008, 9, 2579–2605. [Google Scholar]
- Kobak, D.; Berens, P. The art of using t-SNE for single-cell transcriptomics. Nat. Commun. 2019, 10, 5416. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.-C.; Xu, J.; Brinkhof, B.; Wang, H.; Cui, Z.; Huang, W.E.; Ye, H. A single-cell Raman-based platform to identify developmental stages of human pluripotent stem cell-derived neurons. Proc. Natl. Acad. Sci. USA 2020, 117, 18412–18423. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Zhao, Z.; Qian, N.; Wei, S.; Hu, F.; Min, W. Multiplexed live-cell profiling with Raman probes. Nat. Commun. 2021, 12, 3405. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Guestrin, C. XGBoost: A Scalable Tree Boosting System. In Proceedings of the 22nd ACM SIGKDD International Conference on Knowledge Discovery and Data Mining, San Francisco, CA, USA, 13–17 August 2016; pp. 785–794. [Google Scholar] [CrossRef]
- Paraskevaidi, M.; Matthew, B.J.; Holly, B.J.; Hugh, B.J.; Thulya, C.P.V.; Loren, C.; StJohn, C.; Peter, G.; Callum, G.; Sergei, K.G.; et al. Clinical applications of infrared and Raman spectroscopy in the fields of cancer and infectious diseases. Appl. Spectrosc. Rev. 2021, 56, 804–868. [Google Scholar] [CrossRef]
- Guang, P.; Huang, W.; Guo, L.; Yang, X.; Huang, F.; Yang, M.; Wen, W.; Li, L. Blood-based FTIR-ATR spectroscopy coupled with extreme gradient boosting for the diagnosis of type 2 diabetes. Medicine 2020, 99, e19657. [Google Scholar] [CrossRef]
- Mwanga, E.P.; Minja, E.G.; Mrimi, E.; Jiménez, M.G.; Swai, J.K.; Abbasi, S.; Ngowo, H.S.; Siria, D.J.; Mapua, S.; Stica, C.; et al. Detection of malaria parasites in dried human blood spots using mid-infrared spectroscopy and logistic regression analysis. Malar. J. 2019, 18, 341. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Fang, T.; Li, Y.; Guo, L.; Li, F.; Huang, F.; Li, L. Pre-diabetes diagnosis based on ATR-FTIR spectroscopy combined with CART and XGBoots. Optik 2019, 180, 189–198. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| F | p-Value | Molecule |
|---|---|---|
| 3246 | <0.001 | Actin |
| 3836 | <0.001 | Cytochrome C |
| 4143 | <0.001 | Glycogen |
| 418 | <0.001 | IL8 |
| 2352 | <0.001 | Phosphatidyl-choline |
| 1531 | <0.001 | Phosphatidyl-inositol |
| 1006 | <0.001 | RNA |
| 336 | <0.001 | TGF-β2 |
| 1092 | <0.001 | Vitamin-C |
| 2657 | <0.001 | Vitamin-E |
| A | B | SE | T | p-Value | Molecule |
|---|---|---|---|---|---|
| PNT1A 0 Gy | PNT1A 0 Gy ICCM | 1.964 × 10−3 | −2.4 | 0.074 | Actin |
| PNT1A 0 Gy | PNT1A OF | 1.713 × 10−3 | 32.4 | <0.001 | Actin |
| PNT1A 0 Gy | PNT1A OF ICCM | 1.630 × 10−3 | −30.2 | <0.001 | Actin |
| PNT1A 0 Gy | PNT1A 0 Gy ICCM | 3.004 × 10−2 | 6.4 | <0.001 | Cytochrome C |
| PNT1A 0 Gy | PNT1A OF | 2.772 × 10−2 | −20.9 | <0.001 | Cytochrome C |
| PNT1A 0 Gy | PNT1A OF ICCM | 2.255 × 10−2 | 42.9 | <0.001 | Cytochrome C |
| PNT1A 0 Gy | PNT1A 0 Gy ICCM | 5.284 × 10−3 | 35.9 | <0.001 | Glycogen |
| PNT1A 0 Gy | PNT1A OF | 7.196 × 10−3 | −73.6 | <0.001 | Glycogen |
| PNT1A 0 Gy | PNT1A OF ICCM | 5.642 × 10−3 | 1.0 | 0.757 | Glycogen |
| PNT1A 0 Gy | PNT1A 0 Gy ICCM | 2.950 × 10−2 | −0.2 | 0.997 | IL8 |
| PNT1A 0 Gy | PNT1A OF | 2.188 × 10−2 | −16.9 | <0.001 | IL8 |
| PNT1A 0 Gy | PNT1A OF ICCM | 3.463 × 10−2 | −33.0 | <0.001 | IL8 |
| PNT1A 0 Gy | PNT1A 0 Gy ICCM | 4.661 × 10−3 | −16.7 | <0.001 | Phosphatidyl-choline |
| PNT1A 0 Gy | PNT1A OF | 3.810 × 10−3 | 37.9 | <0.001 | Phosphatidyl-choline |
| PNT1A 0 Gy | PNT1A OF ICCM | 3.966 × 10−3 | −17.7 | <0.001 | Phosphatidyl-choline |
| PNT1A 0 Gy | PNT1A 0 Gy ICCM | 4.236 × 10−3 | 51.3 | <0.001 | Phosphatidyl-inositol |
| PNT1A 0 Gy | PNT1A OF | 7.814 × 10−3 | −34.7 | <0.001 | Phosphatidyl-inositol |
| PNT1A 0 Gy | PNT1A OF ICCM | 5.190 × 10−3 | 18.6 | <0.001 | Phosphatidyl-inositol |
| PNT1A 0 Gy | PNT1A 0 Gy ICCM | 1.706 × 10−3 | 51.5 | <0.001 | RNA |
| PNT1A 0 Gy | PNT1A OF | 1.721 × 10−3 | 12.1 | <0.001 | RNA |
| PNT1A 0 Gy | PNT1A OF ICCM | 1.933 × 10−3 | 23.5 | <0.001 | RNA |
| PNT1A 0 Gy | PNT1A 0 Gy ICCM | 1.507 × 10−3 | 11.5 | <0.001 | TGF-β2 |
| PNT1A 0 Gy | PNT1A OF | 1.429 × 10−3 | −15.3 | <0.001 | TGF-β2 |
| PNT1A 0 Gy | PNT1A OF ICCM | 1.486 × 10−3 | −5.6 | <0.001 | TGF-β2 |
| PNT1A 0 Gy | PNT1A 0 Gy ICCM | 4.315 × 10−2 | −23.6 | <0.001 | Vitamin-C |
| PNT1A 0 Gy | PNT1A OF | 2.655 × 10−2 | −33.8 | <0.001 | Vitamin-C |
| PNT1A 0 Gy | PNT1A OF ICCM | 2.770 × 10−2 | 17.5 | <0.001 | Vitamin-C |
| PNT1A 0 Gy | PNT1A 0 Gy ICCM | 7.249 × 10−3 | 23.2 | <0.001 | Vitamin-E |
| PNT1A 0 Gy | PNT1A OF | 7.169 × 10−3 | −45.8 | <0.001 | Vitamin-E |
| PNT1A 0 Gy | PNT1A OF ICCM | 9.688 × 10−3 | 43.5 | <0.001 | Vitamin-E |
| F | p-Value | Molecule |
|---|---|---|
| 1167 | <0.001 | Actin |
| 1180 | <0.001 | Cytochrome C |
| 3938 | <0.001 | Glycogen |
| 106 | <0.001 | IL8 |
| 272 | <0.001 | Phosphatidyl-choline |
| 7020 | <0.001 | Phosphatidyl-inositol |
| 389 | <0.001 | RNA |
| 1 | <0.001 | TGF-β2 |
| 5246 | <0.001 | Vitamin-C |
| 5833 | <0.001 | Vitamin-E |
| A | B | SE | T | p-Value | Molecule |
|---|---|---|---|---|---|
| LNCaP 0 Gy | LNCaP IF | 0.004 | 34.2 | <0.001 | Actin |
| LNCaP 0 Gy | LNCaP IF | 0.112 | −34.4 | <0.001 | Cytochrome C |
| LNCaP 0 Gy | LNCaP IF | 0.052 | −62.8 | <0.001 | Glycogen |
| LNCaP 0 Gy | LNCaP IF | 0.064 | 10.3 | <0.001 | IL8 |
| LNCaP 0 Gy | LNCaP IF | 0.006 | −16.5 | <0.001 | Phosphatidyl-choline |
| LNCaP 0 Gy | LNCaP IF | 0.034 | −83.8 | <0.001 | Phosphatidyl-inositol |
| LNCaP 0 Gy | LNCaP IF | 0.004 | 19.7 | <0.001 | RNA |
| LNCaP 0 Gy | LNCaP IF | 0.004 | −1.1 | <0.001 | TGF-β2 |
| LNCaP 0 Gy | LNCaP IF | 1.348 | −72.4 | <0.001 | Vitamin-C |
| LNCaP 0 Gy | LNCaP IF | 0.135 | −76.4 | <0.001 | Vitamin-E |
| Irradiation | Volume Average (μm3) | Std Dev |
|---|---|---|
| 0 Gy | 699,592 | 105,269 |
| Out of field (OF) | 345,629 | 192,621 |
| 0 Gy + ICCM | 514,930 | 83,311 |
| Out of field + ICCM (OF + ICCM) | 1,062,630 | 254,959 |
| Sample | Exposure Mode | Protein | Carbohydrate | RNA | Lipid | Cytokine | Antioxidants | Cytochrome C | DNA Damage | Cell Survival |
|---|---|---|---|---|---|---|---|---|---|---|
| PNT1A OF | Low dose | Decrease | Increase | No change | Increase | Increase | No change | Increase | Increase | Increase |
| PNT1A OF ICCM | Low dose plus exposure to secreted factors | Increase | No change | Decrease | No change | No change | Decrease | Decrease | Increase | Increase |
| PNTT1A ICCM | Exposure to secreted factors | No change | Decrease | Decrease | Increase | No change | Decrease | No change | No change | Decrease |
| LNCAP 2 Gy | Exposure to high doses | Decrease | Increase | Decrease | Increase | Decrease | Increase | Increase | Increase | Not measured |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Slattery, C.; Nguyen, K.; Shields, L.; Vega-Carrascal, I.; Singleton, S.; Lyng, F.M.; McClean, B.; Meade, A.D. Application of Advanced Non-Linear Spectral Decomposition and Regression Methods for Spectroscopic Analysis of Targeted and Non-Targeted Irradiation Effects in an In-Vitro Model. Int. J. Mol. Sci. 2022, 23, 12986. https://doi.org/10.3390/ijms232112986
Slattery C, Nguyen K, Shields L, Vega-Carrascal I, Singleton S, Lyng FM, McClean B, Meade AD. Application of Advanced Non-Linear Spectral Decomposition and Regression Methods for Spectroscopic Analysis of Targeted and Non-Targeted Irradiation Effects in an In-Vitro Model. International Journal of Molecular Sciences. 2022; 23(21):12986. https://doi.org/10.3390/ijms232112986
Chicago/Turabian StyleSlattery, Ciara, Khanh Nguyen, Laura Shields, Isabel Vega-Carrascal, Sean Singleton, Fiona M. Lyng, Brendan McClean, and Aidan D. Meade. 2022. "Application of Advanced Non-Linear Spectral Decomposition and Regression Methods for Spectroscopic Analysis of Targeted and Non-Targeted Irradiation Effects in an In-Vitro Model" International Journal of Molecular Sciences 23, no. 21: 12986. https://doi.org/10.3390/ijms232112986
APA StyleSlattery, C., Nguyen, K., Shields, L., Vega-Carrascal, I., Singleton, S., Lyng, F. M., McClean, B., & Meade, A. D. (2022). Application of Advanced Non-Linear Spectral Decomposition and Regression Methods for Spectroscopic Analysis of Targeted and Non-Targeted Irradiation Effects in an In-Vitro Model. International Journal of Molecular Sciences, 23(21), 12986. https://doi.org/10.3390/ijms232112986