
P-Selectin is a Critical Factor for Platelet-Mediated Protection on Restraint Stress-Induced Gastrointestinal Injury in Mice

 and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
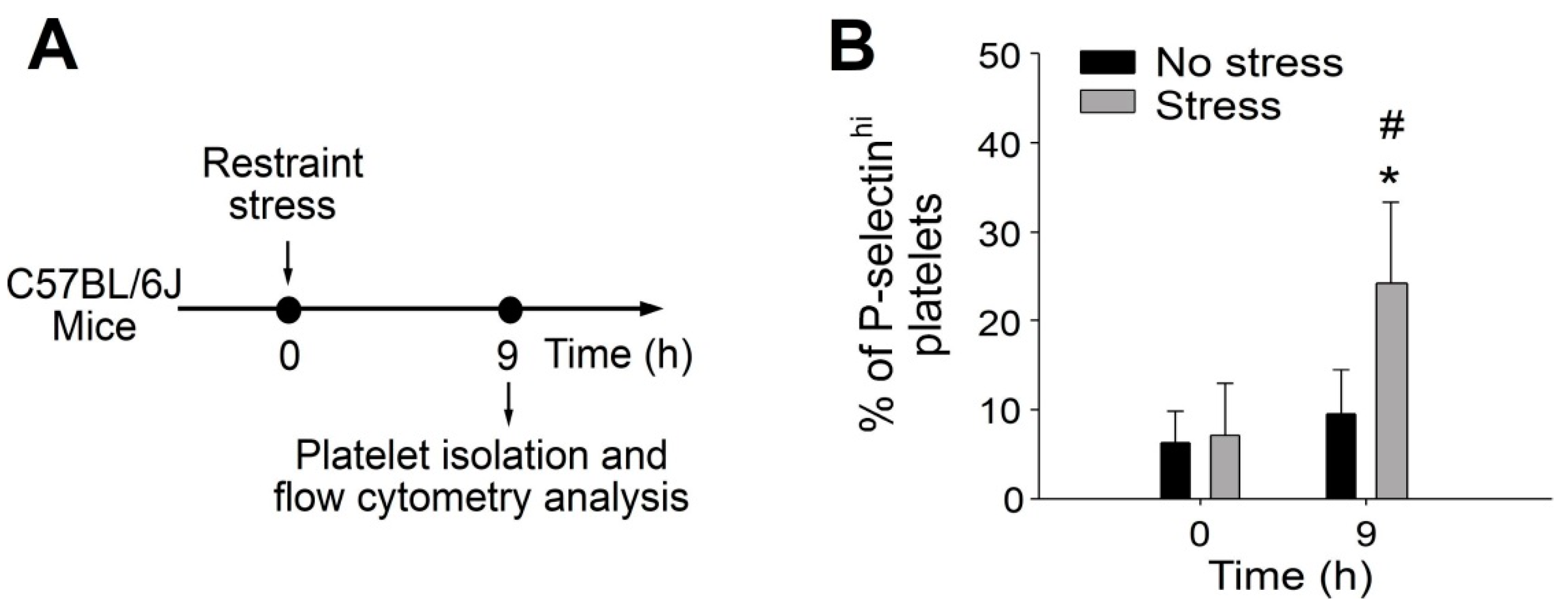
2.1. Involvement of P-Selectin+ Platelets in GI Protection during Stress-Induced GI Injury
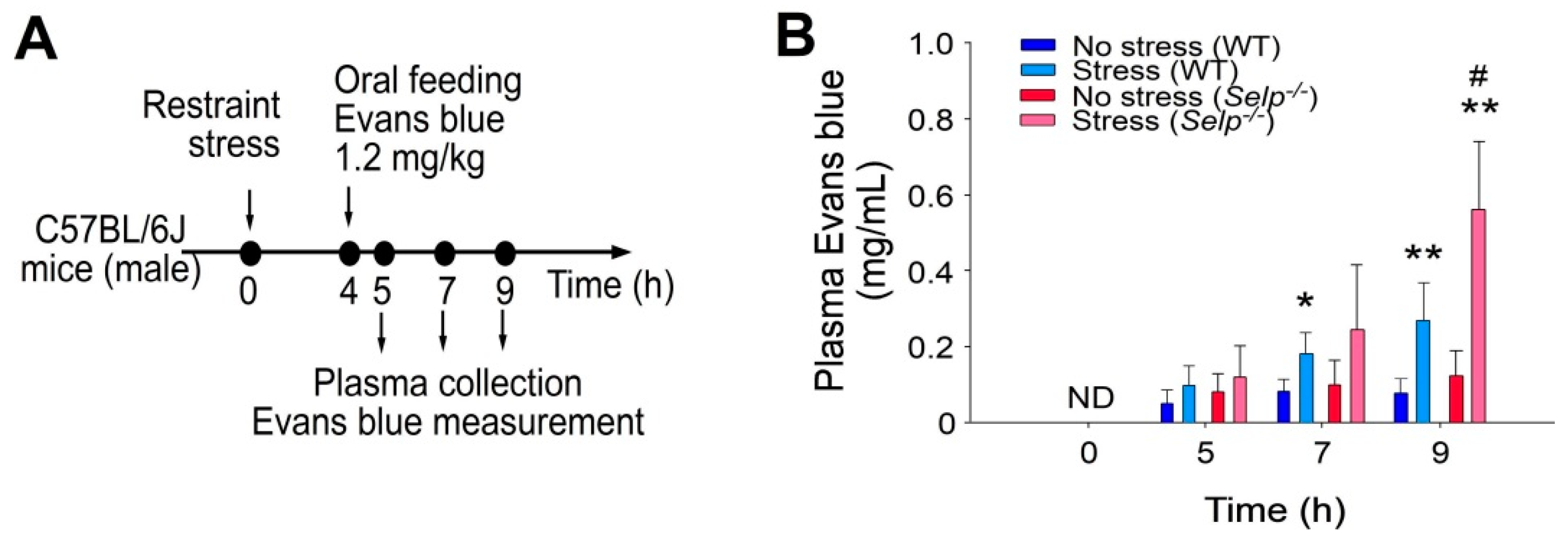
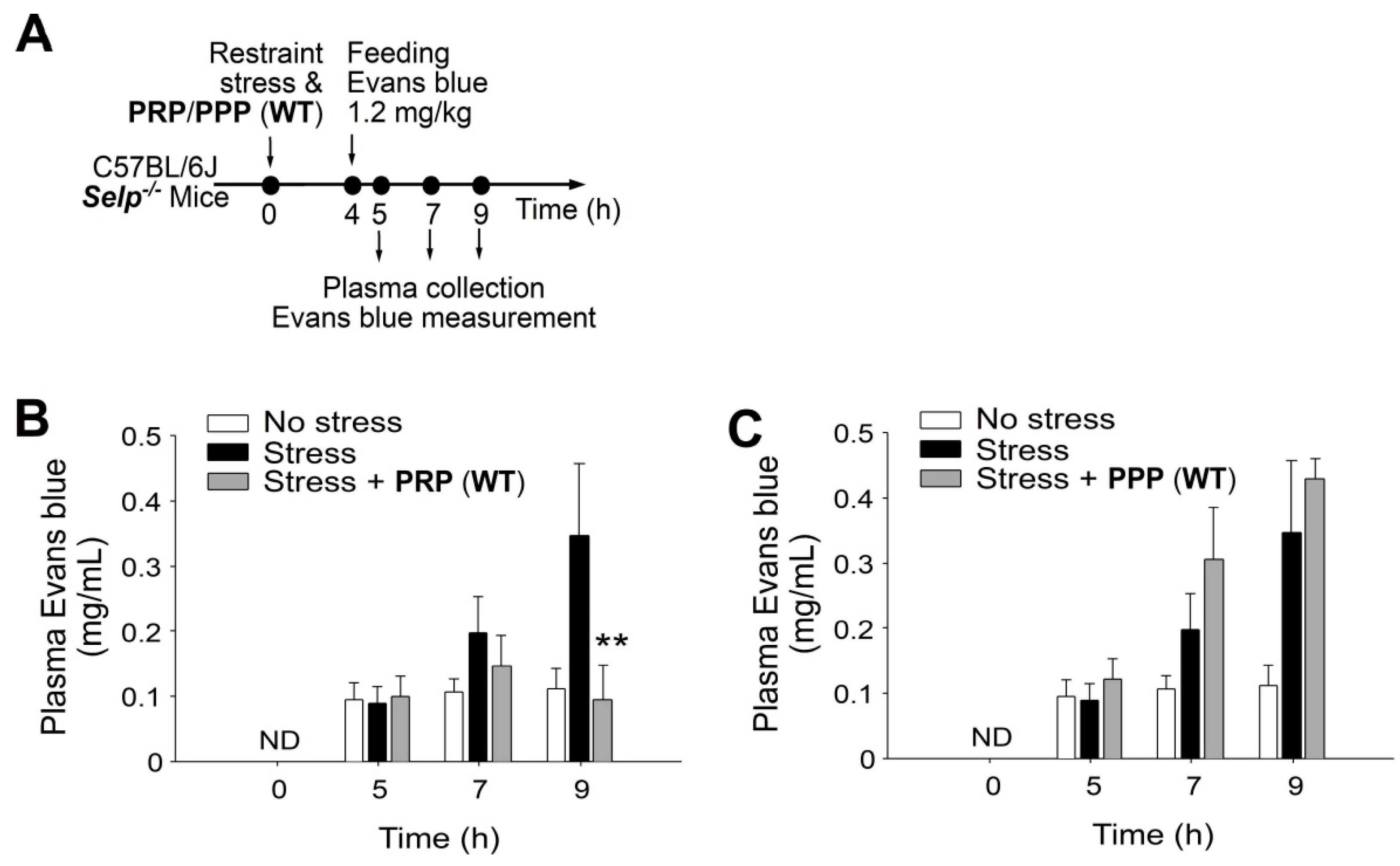
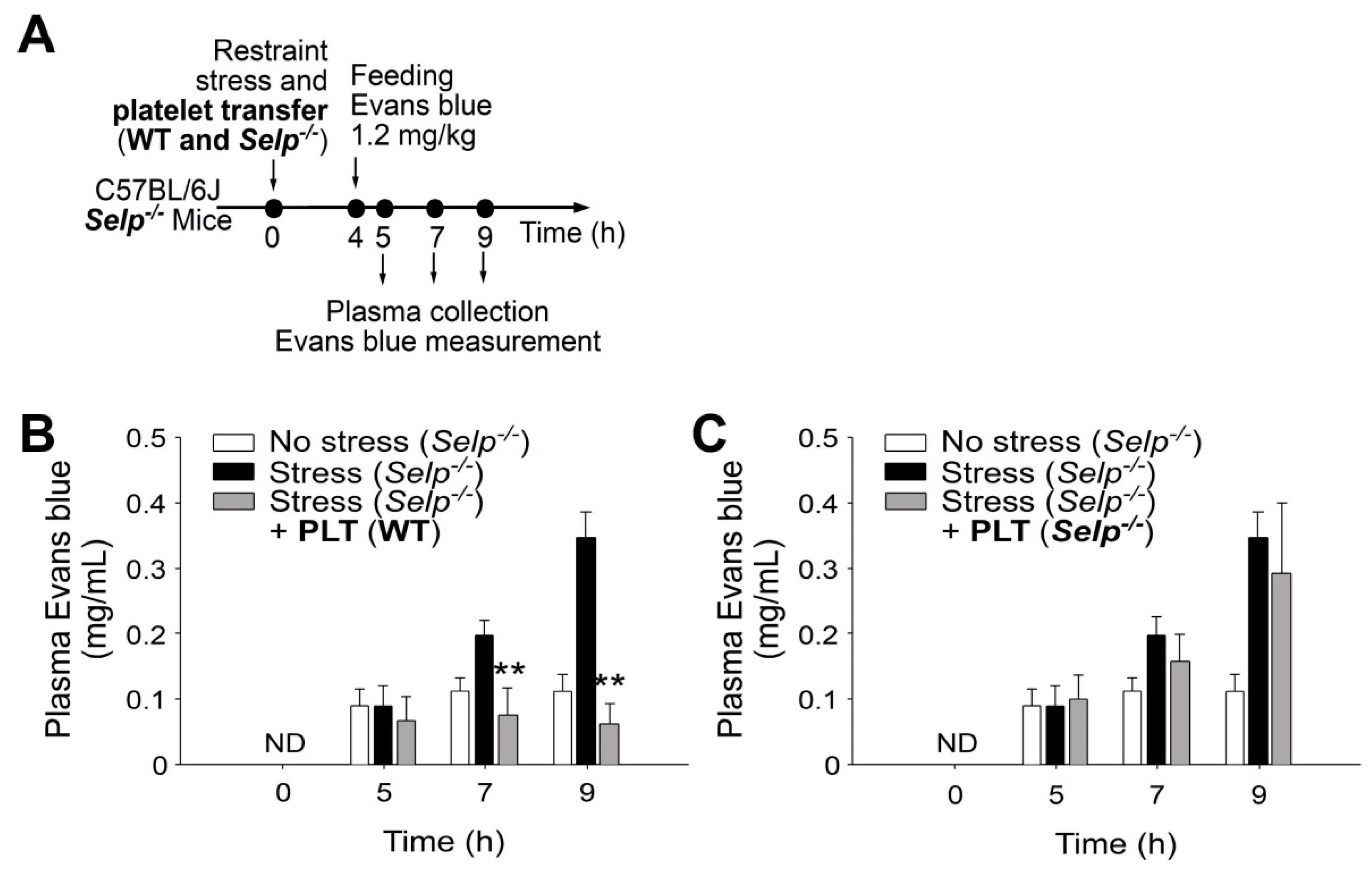
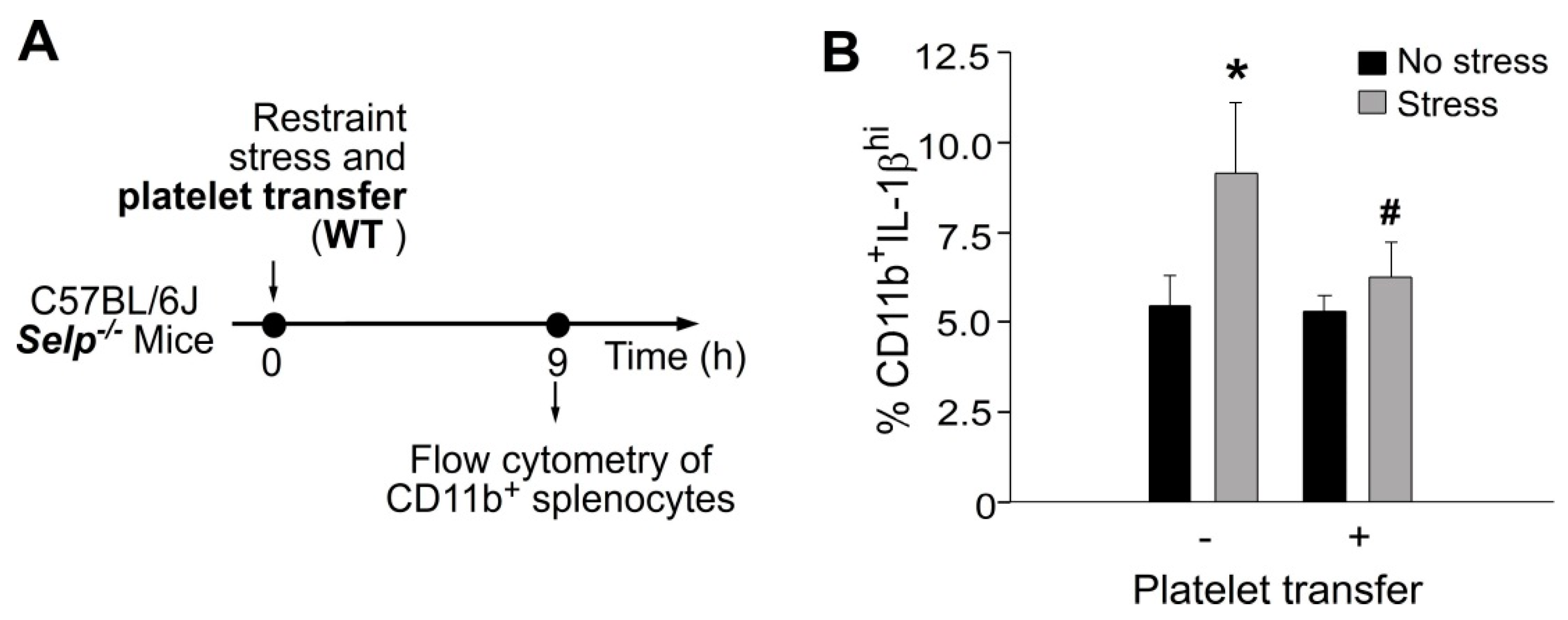
2.2. Platelet-Transfer Ameliorated Stress-Induced GI Leakage and Inflammation, in which P-Selection Is a Critical Factor
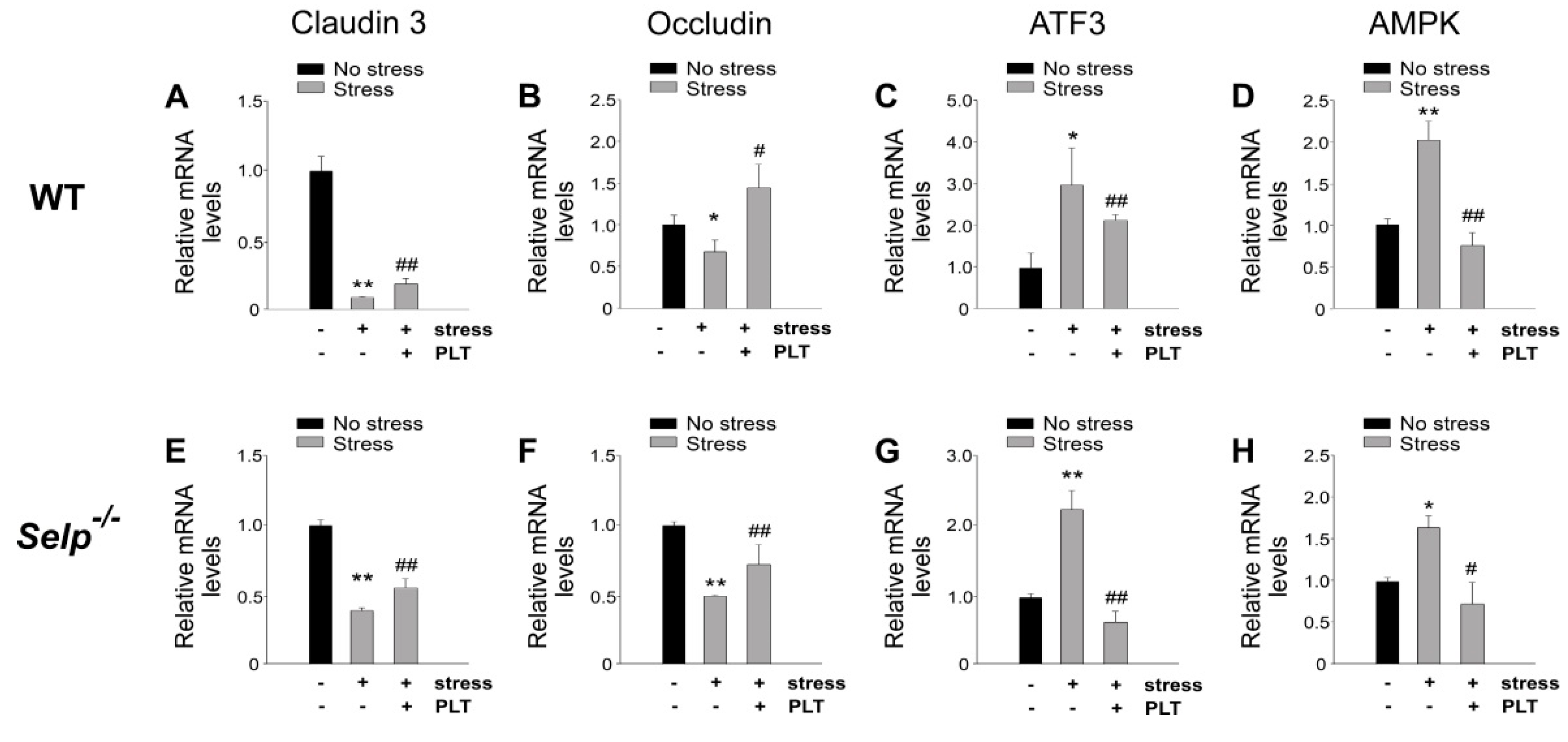
2.3. Platelet-Mediated Protection Is Associated with Corrections of Stress-Induced Aberrant GI mRNA Expression
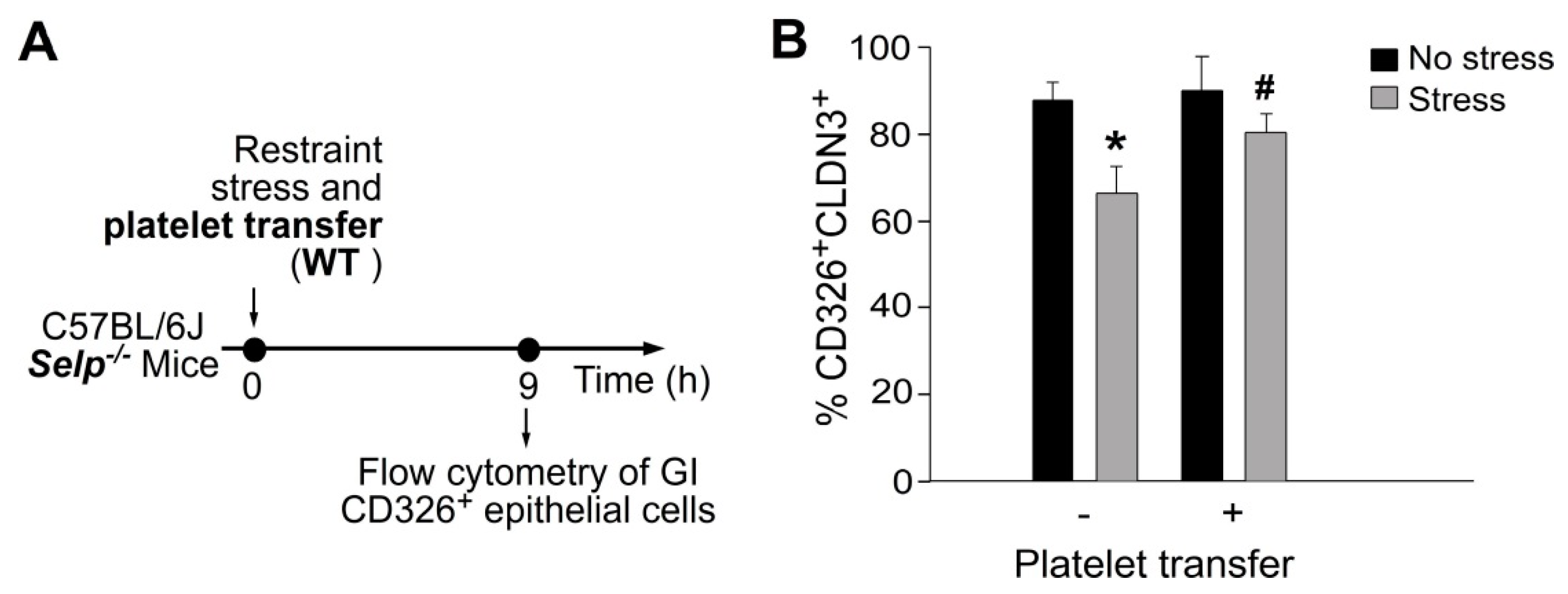
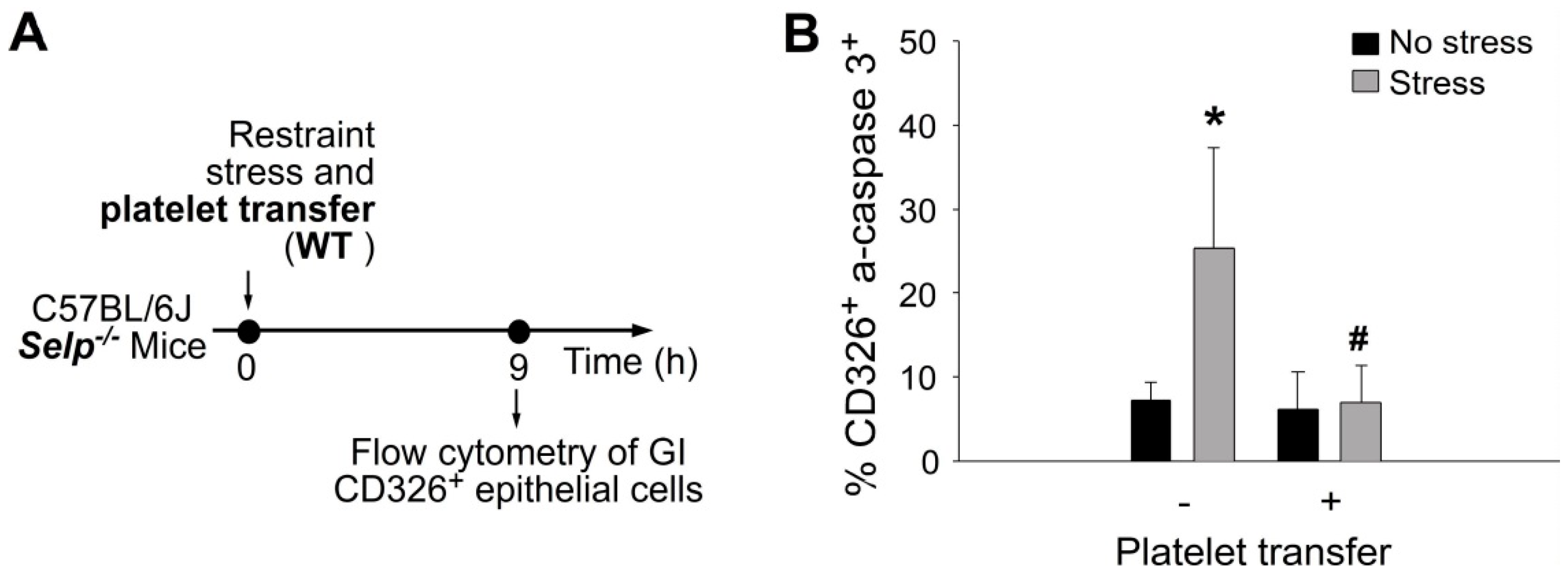
2.4. Platelet-Mediated Protection Is Associated with Rescue of Stress-Mediated Suppression on GI Claudin 3 (CLDN3) Expression and Stress-Induced GI Epithelial Cell Apoptosis
3. Discussion
4. Materials and Methods
4.1. Laboratory Mice
4.2. Induction and Measurement of Stress-Induced GI Leakage
4.3. Platelet, PRP and PPP Transfer
4.4. Bleeding Time Experiment
4.5. qRT-PCR Analysis
4.5.1. RNA Isolation and cDNA Preparation
4.5.2. qRT-PCR
4.6. Flow Cytometry Analysis
4.7. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sapolsky, R.M. Why Zebras Don’t Get Ulcers, 3rd ed.; W.H. Freeman: New York, NY, USA, 2004. [Google Scholar]
- Oligschlaeger, Y.; Yadati, T.; Houben, T.; Condello Olivan, C.M.; Shiri-Sverdlov, R. Inflammatory Bowel Disease: A Stressed “Gut/Feeling”. Cells 2019, 8, 659. [Google Scholar] [CrossRef]
- Rao, M.; Gershon, M.D. The bowel and beyond: The enteric nervous system in neurological disorders. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 517–528. [Google Scholar] [CrossRef]
- GBD 2017 Disease and Injury Incidence and Prevalence Collaborators. Global, regional, and national incidence, prevalence, and years lived with disability for 354 diseases and injuries for 195 countries and territories, 1990–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet 2018, 392, 1789–1858. [Google Scholar] [CrossRef]
- Severance, E.G.; Prandovszky, E.; Castiglione, J.; Yolken, R.H. Gastroenterology issues in schizophrenia: Why the gut matters. Curr. Psychiatry Rep. 2015, 17, 27. [Google Scholar] [CrossRef]
- Karling, P.; Maripuu, M.; Wikgren, M.; Adolfsson, R.; Norrback, K.-F. Association between gastrointestinal symptoms and affectivity in patients with bipolar disorder. World J. Gastroenterol. 2016, 22, 8540–8548. [Google Scholar] [CrossRef]
- Zhang, B.; Wang, H.E.; Bai, Y.-M.; Tsai, S.-J.; Su, T.-P.; Chen, T.-J.; Wang, Y.-P.; Chen, M.-H. Inflammatory bowel disease is associated with higher dementia risk: A nationwide longitudinal study. Gut 2021, 70, 85–91. [Google Scholar] [CrossRef]
- Alkasir, R.; Li, J.; Li, X.; Jin, M.; Zhu, B. Human gut microbiota: The links with dementia development. Protein Cell 2017, 8, 90–102. [Google Scholar] [CrossRef] [PubMed]
- Wasilewska, J.; Klukowski, M. Gastrointestinal symptoms and autism spectrum disorder: Links and risks—A possible new overlap syndrome. Pediatr. Health Med. Ther. 2015, 6, 153–166. [Google Scholar] [CrossRef]
- Long-Smith, C.; O’Riordan, K.J.; Clarke, G.; Stanton, C.; Dinan, T.G.; Cryan, J.F. Microbiota-Gut-Brain Axis: New Therapeutic Opportunities. Annu. Rev. Pharmacol. Toxicol. 2020, 60, 477–502. [Google Scholar] [CrossRef]
- Cryan, J.F.; O’Riordan, K.J.; Cowan, C.S.M.; Sandhu, K.V.; Bastiaanssen, T.F.S.; Boehme, M.; Codagnone, M.G.; Cussotto, S.; Fulling, C.; Golubeva, A.V.; et al. The Microbiota-Gut-Brain Axis. Physiol. Rev. 2019, 99, 1877–2013. [Google Scholar] [CrossRef]
- Campos, A.C.; Fogaca, M.V.; Aguiar, D.C.; Guimaraes, F.S. Animal models of anxiety disorders and stress. Rev. Bras. Psiquiatr. 2013, 35 (Suppl. 2), S101–S111. [Google Scholar] [CrossRef]
- Paré, W.P.; Glavin, G.B. Restraint stress in biomedical research: A review. Neurosci. Biobehav. Rev. 1986, 10, 339–370. [Google Scholar] [CrossRef]
- Glavin, G.B.; Paré, W.P.; Sandbak, T.; Bakke, H.-K.; Murison, R. Restraint stress in biomedical research: An update. Neurosci. Biobehav. Rev. 1994, 18, 223–249. [Google Scholar] [CrossRef]
- Chuang, D.-J.; Pethaperumal, S.; Siwakoti, B.; Chien, H.-J.; Cheng, C.-F.; Hung, S.-C.; Lien, T.-S.; Sun, D.-S.; Chang, H.-H. Activating Transcription Factor 3 Protects against Restraint Stress-Induced Gastrointestinal Injury in Mice. Cells 2021, 10, 3530. [Google Scholar] [CrossRef] [PubMed]
- McEver, R.P. Selectins: Initiators of leucocyte adhesion and signalling at the vascular wall. Cardiovasc. Res. 2015, 107, 331–339. [Google Scholar] [CrossRef]
- Eisinger, F.; Patzelt, J.; Langer, H.F. The Platelet Response to Tissue Injury. Front. Med. 2018, 5, 317. [Google Scholar] [CrossRef]
- Perkins, L.A.; Anderson, C.J.; Novelli, E.M. Targeting P-Selectin Adhesion Molecule in Molecular Imaging: P-Selectin Expression as a Valuable Imaging Biomarker of Inflammation in Cardiovascular Disease. J. Nucl. Med. 2019, 60, 1691–1697. [Google Scholar] [CrossRef]
- Sun, D.-S.; Chang, Y.-W.; Kau, J.-H.; Huang, H.-H.; Ho, P.-H.; Tzeng, Y.-J.; Chang, H.-H. Soluble P-selectin rescues mice from anthrax lethal toxin-induced mortality through PSGL-1 pathway-mediated correction of hemostasis. Virulence 2017, 8, 1216–1228. [Google Scholar] [CrossRef]
- Kapupara, K.; Wen, Y.-T.; Tsai, R.-K.; Huang, S.-P. Soluble P-selectin promotes retinal ganglion cell survival through activation of Nrf2 signaling after ischemia injury. Cell Death Dis. 2017, 8, e3172. [Google Scholar] [CrossRef]
- Sun, D.-S.; Ho, P.-H.; Chang, H.-H. Soluble P-selectin rescues viper venom-induced mortality through anti-inflammatory properties and PSGL-1 pathway-mediated correction of hemostasis. Sci. Rep. 2016, 6, 35868. [Google Scholar] [CrossRef]
- Huang, H.-S.; Chang, H.-H. Platelets in inflammation and immune modulations: Functions beyond hemostasis. Arch. Immunol. Ther. Exp. 2012, 60, 443–451. [Google Scholar] [CrossRef]
- Zhao, X.; Zeng, H.; Lei, L.; Tong, X.; Yang, L.; Yang, Y.; Li, S.; Zhou, Y.; Luo, L.; Huang, J.; et al. Tight junctions and their regulation by non-coding RNAs. Int. J. Biol. Sci. 2021, 17, 712–727. [Google Scholar] [CrossRef]
- Gawaz, M.; Vogel, S. Platelets in tissue repair: Control of apoptosis and interactions with regenerative cells. Blood 2013, 122, 2550–2554. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Mhaskar, R.; Grossman, B.J.; Kaufman, R.M.; Tobian, A.A.; Kleinman, S.; Gernsheimer, T.; Tinmouth, A.T.; Djulbegovic, B.; Panel, A.P.T.G. Platelet transfusion: A systematic review of the clinical evidence. Transfusion 2015, 55, 1116–1127. [Google Scholar] [CrossRef] [PubMed]
- Ding, Z.-Y.; Tan, Y.; Peng, Q.; Zuo, J.; Li, N. Novel applications of platelet concentrates in tissue regeneration (Review). Exp. Ther. Med. 2021, 21, 226. [Google Scholar] [CrossRef] [PubMed]
- Alves, R.; Grimalt, R. A Review of Platelet-Rich Plasma: History, Biology, Mechanism of Action, and Classification. Skin Appendage Disord. 2018, 4, 18–24. [Google Scholar] [CrossRef]
- Everts, P.; Onishi, K.; Jayaram, P.; Lana, J.F.; Mautner, K. Platelet-Rich Plasma: New Performance Understandings and Therapeutic Considerations in 2020. Int. J. Mol. Sci. 2020, 21, 7794. [Google Scholar] [CrossRef]
- Geropoulos, G.; Psarras, K.; Giannis, D.; Martzivanou, E.C.; Papaioannou, M.; Kakos, C.D.; Pavlidis, E.T.; Symeonidis, N.; Koliakos, G.; Pavlidis, T.E. Platelet rich plasma effectiveness in bowel anastomoses: A systematic review. World J. Gastrointest. Surg. 2021, 13, 1736–1753. [Google Scholar] [CrossRef]
- Daradka, M.; Alardah, M.M.; Ismail, Z.B. Effects of autologous platelet-rich plasma coated sutures on intestinal anastomotic healing in rabbits. Heliyon 2019, 5, e02713. [Google Scholar] [CrossRef]
- Fresno, L.; Fondevila, D.; Bambo, O.; Chacaltana, A.; García, F.; Andaluz, A. Effects of platelet-rich plasma on intestinal wound healing in pigs. Vet. J. 2010, 185, 322–327. [Google Scholar] [CrossRef]
- Daglioglu, Y.K.; Duzgun, O.; Sarici, I.S.; Ulutaş, K.T. Comparison of platelet rich plasma versus fibrin glue on colonic anastomoses in rats. Acta Cir. Bras. 2018, 33, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, G.; Tanrikulu, Y. Short-term Results of Platelet-Rich Plasma in the Treatment of Chronic Anal Fissure: Randomized Controlled Clinical Study. Dis. Colon Rectum 2021, 64, 714–723. [Google Scholar] [CrossRef] [PubMed]
- Imam, S.S.; Al-Abbasi, F.A.; Hosawi, S.; Afzal, M.; Nadeem, M.S.; Ghoneim, M.M.; Alshehri, S.; Alzarea, S.I.; Alquraini, A.; Gupta, G.; et al. Role of platelet rich plasma mediated repair and regeneration of cell in early stage of cardiac injury. Regen. Ther. 2022, 19, 144–153. [Google Scholar] [CrossRef] [PubMed]
- Ruan, D.; Tang, C.; Fei, Y.; Qian, S.; Xiang, X.; Xu, J.; Huang, Z.; Chen, X.; Heng, B.C.; Yin, Z.; et al. Early-Stage Primary Anti-inflammatory Therapy Enhances the Regenerative Efficacy of Platelet-Rich Plasma in a Rabbit Achilles Tendinopathy Model. Am. J. Sports Med. 2021, 49, 3357–3371. [Google Scholar] [CrossRef]
- Kuffler, D.P. Mechanisms for Reducing Neuropathic Pain. Mol. Neurobiol. 2020, 57, 67–87. [Google Scholar] [CrossRef]
- Evangelista, V.; Manarini, S.; Sideri, R.; Rotondo, S.; Martelli, N.; Piccoli, A.; Totani, L.; Piccardoni, P.; Vestweber, D.; de Gaetano, G.; et al. Platelet/polymorphonuclear leukocyte interaction: P-selectin triggers protein-tyrosine phosphorylation-dependent CD11b/CD18 adhesion: Role of PSGL-1 as a signaling molecule. Blood 1999, 93, 876–885. [Google Scholar] [CrossRef]
- Rossaint, J.; Margraf, A.; Zarbock, A. Role of Platelets in Leukocyte Recruitment and Resolution of Inflammation. Front. Immunol. 2018, 9, 2712. [Google Scholar] [CrossRef]
- Impellizzeri, D.; Cuzzocrea, S. Targeting selectins for the treatment of inflammatory diseases. Expert Opin. Ther. Targets 2014, 18, 55–67. [Google Scholar] [CrossRef]
- Lorant, D.E.; Topham, M.K.; Whatley, R.E.; McEver, R.P.; McIntyre, T.M.; Prescott, S.M.; Zimmerman, G.A. Inflammatory roles of P-selectin. J. Clin. Investig. 1993, 92, 559–570. [Google Scholar] [CrossRef]
- Blann, A.D.; Lip, G.Y. Hypothesis: Is soluble P-selectin a new marker of platelet activation? Atherosclerosis 1997, 128, 135–138. [Google Scholar] [CrossRef]
- Ho, Y.-Y.; Sun, D.-S.; Chang, H.-H. Silver Nanoparticles Protect Skin from Ultraviolet B-Induced Damage in Mice. Int. J. Mol. Sci. 2020, 21, 7082. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.-L.; Sun, D.-S.; Su, M.-T.; Lien, T.-S.; Chen, Y.-H.; Lin, C.-Y.; Huang, C.-H.; King, C.-C.; Li, C.-R.; Chen, T.-H.; et al. Suppressed humoral immunity is associated with dengue nonstructural protein NS1-elicited anti-death receptor antibody fractions in mice. Sci. Rep. 2020, 10, 6294. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.-Y.; Hu, C.-T.; Sun, D.-S.; Lien, T.-S.; Chang, H.-H. Thioacetamide-induced liver damage and thrombocytopenia is associated with induction of antiplatelet autoantibody in mice. Sci. Rep. 2019, 9, 17497. [Google Scholar] [CrossRef] [PubMed]
- Perevedentseva, E.; Krivokharchenko, A.; Karmenyan, A.V.; Chang, H.-H.; Cheng, C.-L. Raman spectroscopy on live mouse early embryo while it continues to develop into blastocyst in vitro. Sci. Rep. 2019, 9, 6636. [Google Scholar] [CrossRef]
- Chan, H.; Huang, H.-S.; Sun, D.-S.; Lee, C.-J.; Lien, T.-S.; Chang, H.-H. TRPM8 and RAAS-mediated hypertension is critical for cold-induced immunosuppression in mice. Oncotarget 2018, 9, 12781–12795. [Google Scholar] [CrossRef]
- Huang, C.-Y.; Yu, W.-S.; Liu, G.-C.; Hung, S.-C.; Chang, J.-H.; Chang, J.-C.; Cheng, C.-L.; Sun, D.-S.; Lin, M.-D.; Lin, W.-Y.; et al. Opportunistic gill infection is associated with TiO2 nanoparticle-induced mortality in zebrafish. PLoS ONE 2021, 16, e0247859. [Google Scholar] [CrossRef]
- Hung, S.-C.; Ke, L.-C.; Lien, T.-S.; Huang, H.-S.; Sun, D.-S.; Cheng, C.-L.; Chang, H.-H. Nanodiamond-Induced Thrombocytopenia in Mice Involve P-Selectin-Dependent Nlrp3 Inflammasome-Mediated Platelet Aggregation, Pyroptosis and Apoptosis. Front. Immunol. 2022, 13, 806686. [Google Scholar] [CrossRef]
- Zimprich, A.; Garrett, L.; Deussing, J.M.; Wotjak, C.T.; Fuchs, H.; Gailus-Durner, V.; de Angelis, M.H.; Wurst, W.; Hölter, S.M. A robust and reliable non-invasive test for stress responsivity in mice. Front. Behav. Neurosci. 2014, 8, 125. [Google Scholar] [CrossRef]
- Chu, X.; Zhou, Y.; Hu, Z.; Lou, J.; Song, W.; Li, J.; Liang, X.; Chen, C.; Wang, S.; Yang, B.; et al. 24-hour-restraint stress induces long-term depressive-like phenotypes in mice. Sci. Rep. 2016, 6, 32935. [Google Scholar] [CrossRef]
- Lien, T.-S.; Sun, D.-S.; Hung, S.-C.; Wu, W.-S.; Chang, H.-H. Dengue Virus Envelope Protein Domain III Induces Nlrp3 Inflammasome-Dependent NETosis-Mediated Inflammation in Mice. Front. Immunol. 2021, 12, 618577. [Google Scholar] [CrossRef]
- Lien, T.-S.; Sun, D.-S.; Wu, C.-Y.; Chang, H.-H. Exposure to Dengue Envelope Protein Domain III Induces Nlrp3 Inflammasome-Dependent Endothelial Dysfunction and Hemorrhage in Mice. Front. Immunol. 2021, 12, 617251. [Google Scholar] [CrossRef] [PubMed]
- Lien, T.-S.; Chan, H.; Sun, D.-S.; Wu, J.-C.; Lin, Y.-Y.; Lin, G.L.; Chang, H.-H. Exposure of Platelets to Dengue Virus and Envelope Protein Domain III Induces Nlrp3 Inflammasome-Dependent Platelet Cell Death and Thrombocytopenia in Mice. Front. Immunol. 2021, 12, 616394. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.-S.; Sun, D.-S.; Lien, T.-S.; Chang, H.-H. Dendritic cells modulate platelet activity in IVIg-mediated amelioration of ITP in mice. Blood 2010, 116, 5002–5009. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Dhurat, R.; Sukesh, M. Principles and Methods of Preparation of Platelet-Rich Plasma: A Review and Author’s Perspective. J. Cutan. Aesthet. Surg. 2014, 7, 189–197. [Google Scholar] [CrossRef]
- Bergmeier, W.; Boulaftali, Y. Adoptive transfer method to study platelet function in mouse models of disease. Thromb. Res. 2014, 133 (Suppl. 1), S3–S5. [Google Scholar] [CrossRef]
- Kau, J.-H.; Shih, Y.-L.; Lien, T.-S.; Lee, C.-C.; Huang, H.-H.; Lin, H.-C.; Sun, D.-S.; Chang, H.-H. Activated protein C ameliorates Bacillus anthracis lethal toxin-induced lethal pathogenesis in rats. J. Biomed. Sci. 2012, 19, 98. [Google Scholar] [CrossRef]
- Tsuchida-Straeten, N.; Ensslen, S.; Schafer, C.; Woltje, M.; Denecke, B.; Moser, M.; Graber, S.; Wakabayashi, S.; Koide, T.; Jahnen-Dechent, W. Enhanced blood coagulation and fibrinolysis in mice lacking histidine-rich glycoprotein (HRG). J. Thromb. Haemost. 2005, 3, 865–872. [Google Scholar] [CrossRef]
- Wu, M.-S.; Sun, D.-S.; Lin, Y.-C.; Cheng, C.-L.; Hung, S.-C.; Chen, P.-K.; Yang, J.-H.; Chang, H.-H. Nanodiamonds protect skin from ultraviolet B-induced damage in mice. J. Nanobiotechnology 2015, 13, 35. [Google Scholar] [CrossRef]
- Odenwald, M.A.; Turner, J.R. The intestinal epithelial barrier: A therapeutic target? Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 9–21. [Google Scholar] [CrossRef]
- Groschwitz, K.R.; Hogan, S.P. Intestinal barrier function: Molecular regulation and disease pathogenesis. J. Allergy Clin. Immunol. 2009, 124, 3–20. [Google Scholar] [CrossRef]
- Rath, E.; Moschetta, A.; Haller, D. Mitochondrial function—Gatekeeper of intestinal epithelial cell homeostasis. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 497–516. [Google Scholar] [CrossRef] [PubMed]
- Sheng, H.; Shao, J.; Townsend, C.M., Jr.; Evers, B.M. Phosphatidylinositol 3-kinase mediates proliferative signals in intestinal epithelial cells. Gut 2003, 52, 1472–1478. [Google Scholar] [CrossRef] [PubMed]
- Ouchi, T.; Morimura, S.; Dow, L.E.; Miyoshi, H.; Udey, M.C. EpCAM (CD326) Regulates Intestinal Epithelial Integrity and Stem Cells via Rho-Associated Kinase. Cells 2021, 10, 256. [Google Scholar] [CrossRef] [PubMed]
- Balfe, A.; Lennon, G.; Lavelle, A.; Docherty, N.G.; Coffey, J.C.; Sheahan, K.; Winter, D.C.; O’Connell, P.R. Isolation and gene expression profiling of intestinal epithelial cells: Crypt isolation by calcium chelation from in vivo samples. Clin. Exp. Gastroenterol. 2018, 11, 29–37. [Google Scholar] [CrossRef]
- Chen, Y.-S.; Lin, H.-H.; Hsueh, P.-T.; Ni, W.-F.; Liu, P.-J.; Chen, P.-S.; Chang, H.-H.; Sun, D.-S.; Chen, Y.-L. Involvement of L-selectin expression in Burkholderia pseudomallei-infected monocytes invading the brain during murine melioidosis. Virulence 2017, 8, 751–766. [Google Scholar] [CrossRef]
- Chang, Y.-S.; Ko, B.-H.; Ju, J.-C.; Chang, H.-H.; Huang, S.-H.; Lin, C.-W. SARS Unique Domain (SUD) of Severe Acute Respiratory Syndrome Coronavirus Induces NLRP3 Inflammasome-Dependent CXCL10-Mediated Pulmonary Inflammation. Int. J. Mol. Sci. 2020, 21, 3179. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pethaperumal, S.; Hung, S.-C.; Lien, T.-S.; Sun, D.-S.; Chang, H.-H. P-Selectin is a Critical Factor for Platelet-Mediated Protection on Restraint Stress-Induced Gastrointestinal Injury in Mice. Int. J. Mol. Sci. 2022, 23, 11909. https://doi.org/10.3390/ijms231911909
Pethaperumal S, Hung S-C, Lien T-S, Sun D-S, Chang H-H. P-Selectin is a Critical Factor for Platelet-Mediated Protection on Restraint Stress-Induced Gastrointestinal Injury in Mice. International Journal of Molecular Sciences. 2022; 23(19):11909. https://doi.org/10.3390/ijms231911909
Chicago/Turabian StylePethaperumal, Subhashree, Shih-Che Hung, Te-Sheng Lien, Der-Shan Sun, and Hsin-Hou Chang. 2022. "P-Selectin is a Critical Factor for Platelet-Mediated Protection on Restraint Stress-Induced Gastrointestinal Injury in Mice" International Journal of Molecular Sciences 23, no. 19: 11909. https://doi.org/10.3390/ijms231911909
APA StylePethaperumal, S., Hung, S.-C., Lien, T.-S., Sun, D.-S., & Chang, H.-H. (2022). P-Selectin is a Critical Factor for Platelet-Mediated Protection on Restraint Stress-Induced Gastrointestinal Injury in Mice. International Journal of Molecular Sciences, 23(19), 11909. https://doi.org/10.3390/ijms231911909