
Proteomic, Transcriptomic, Mutational, and Functional Assays Reveal the Involvement of Both THF and PLP Sites at the GmSHMT08 in Resistance to Soybean Cyst Nematode

 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
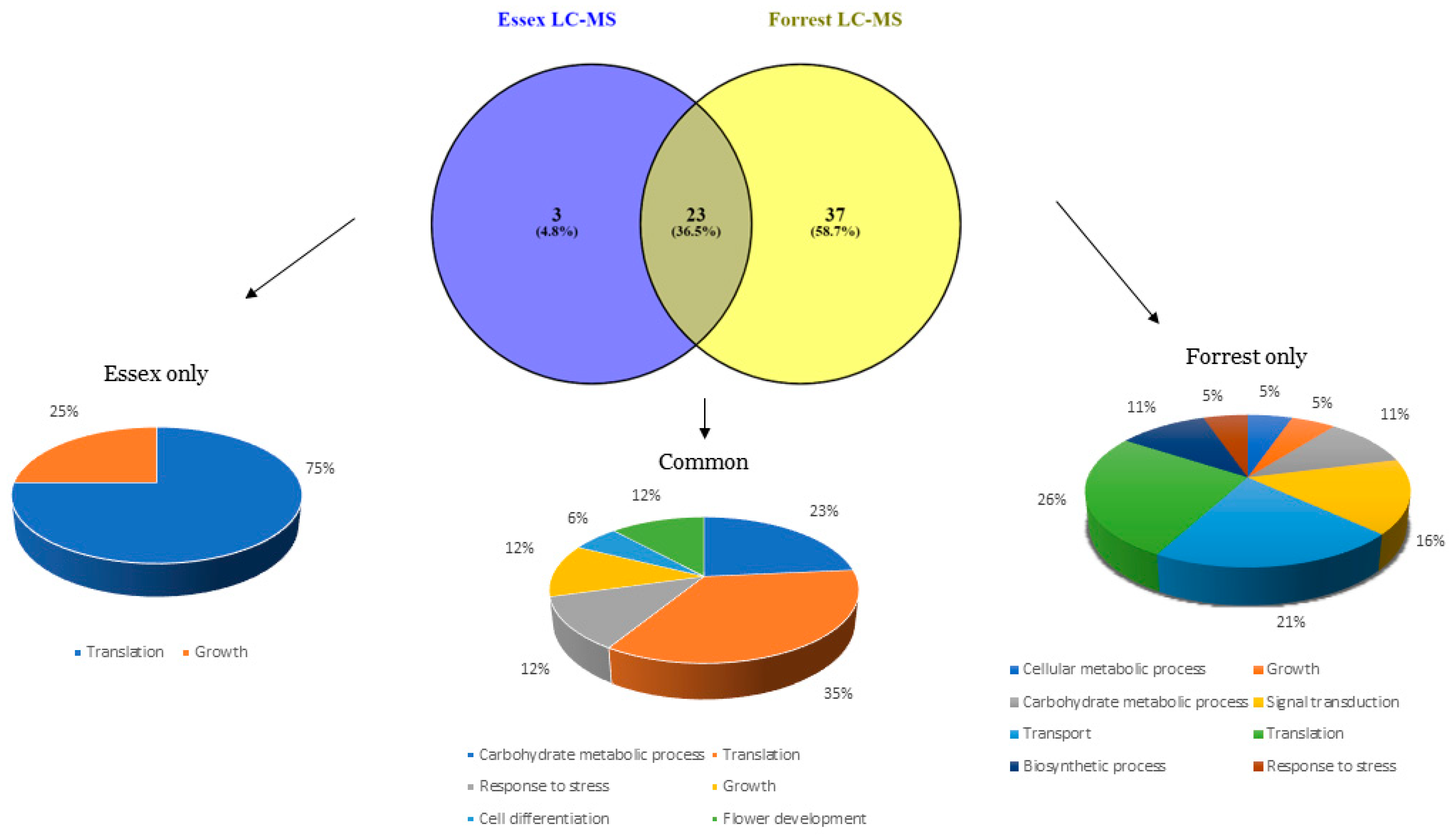
2.1. Mass Spectrometry Identifies the Presence of Proteins That Use Serine and Glycine as Substrates/Precursors
2.2. Identification of Induced Gene Expression in Response to SCN Infection
2.3. Correlation between the Identified Genes and the Previously Reported QTLs for SCN Resistance
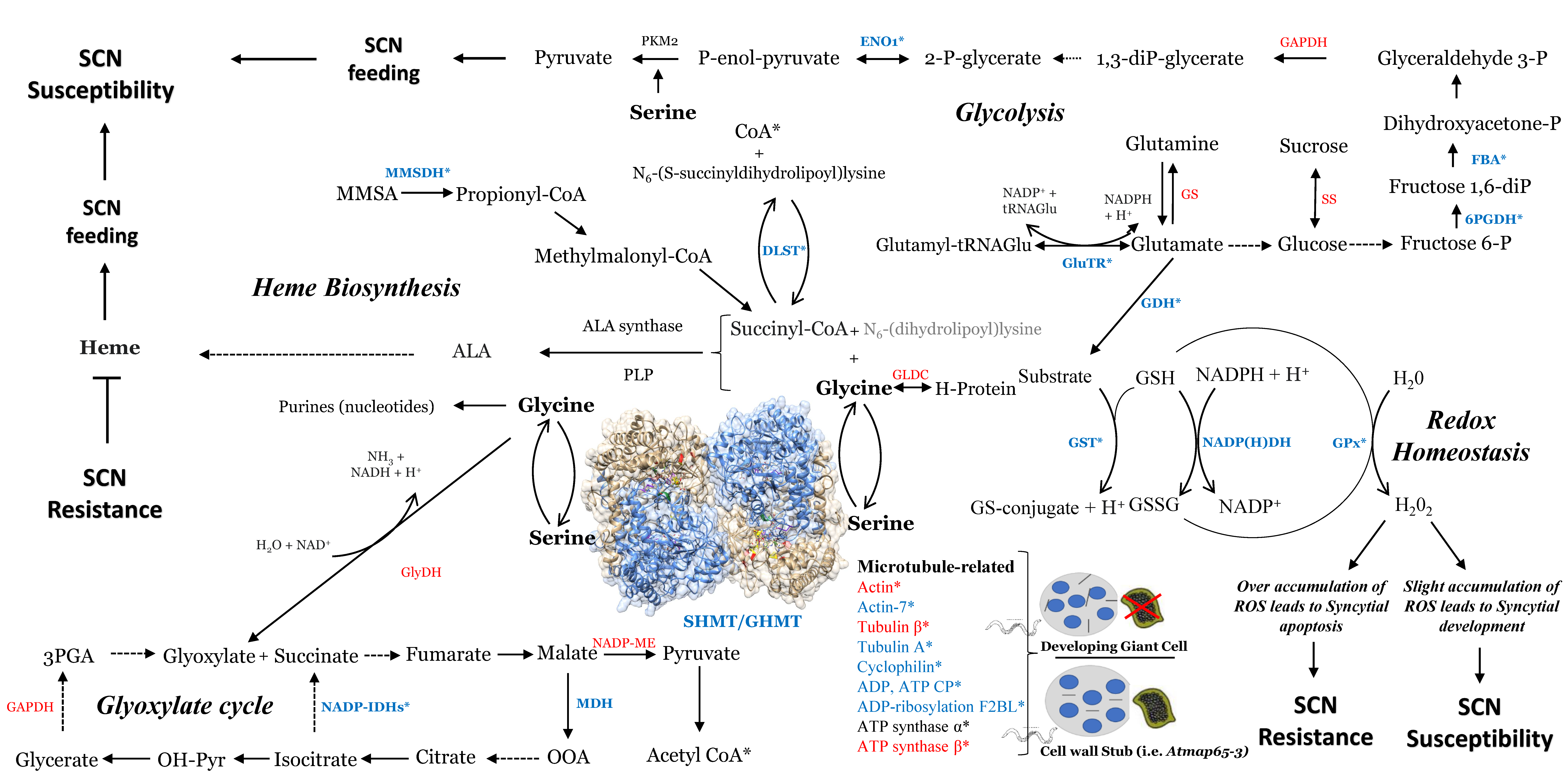
2.4. Identification of Genes Related to Redox Homeostasis
2.5. Glycolysis Cycle in Response to SCN Infection
2.6. Identification of Gene-Related Glyoxylate Cycle
2.7. Identification of Succinyl-CoA, Serine/Glycine, and Heme-Related Genes
2.8. Identification of Cytoskeleton-Related and ATP-Mitochondrial-Related Genes
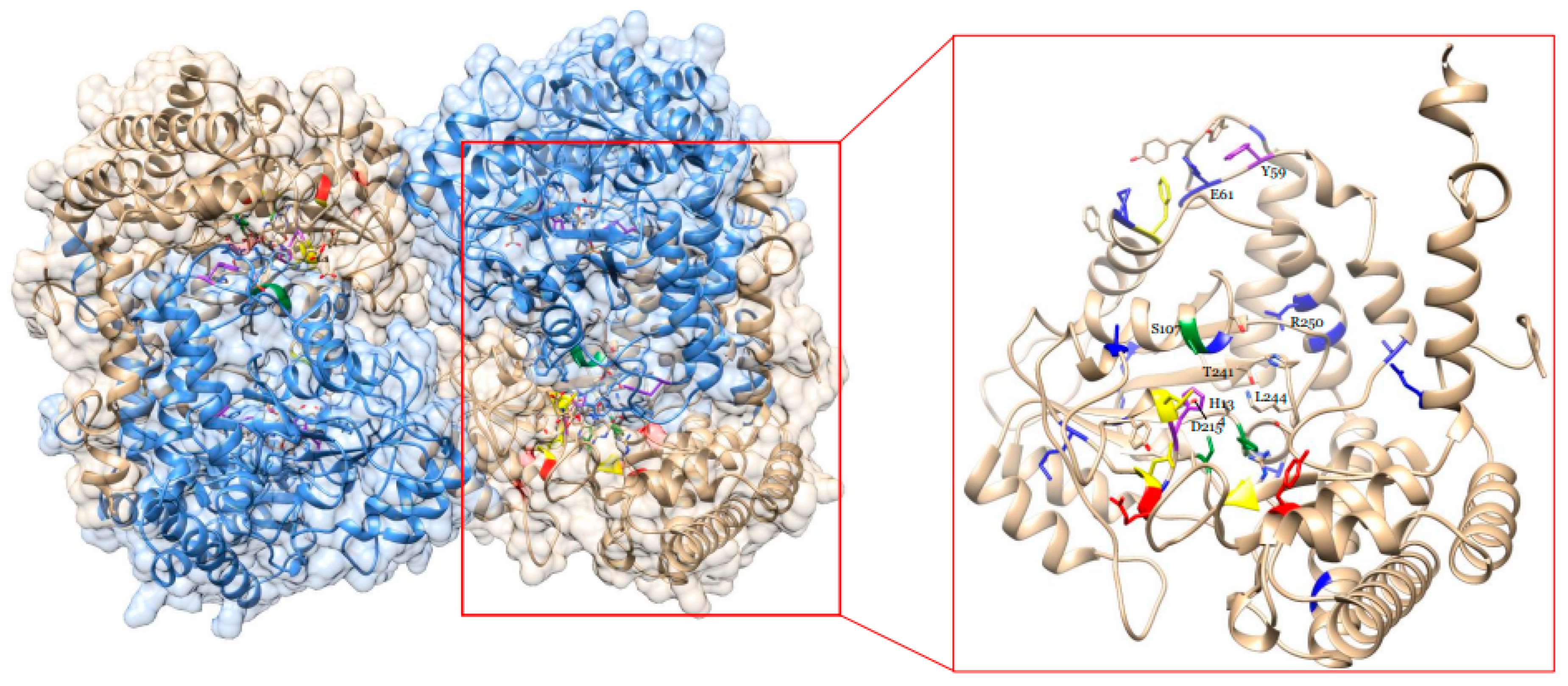
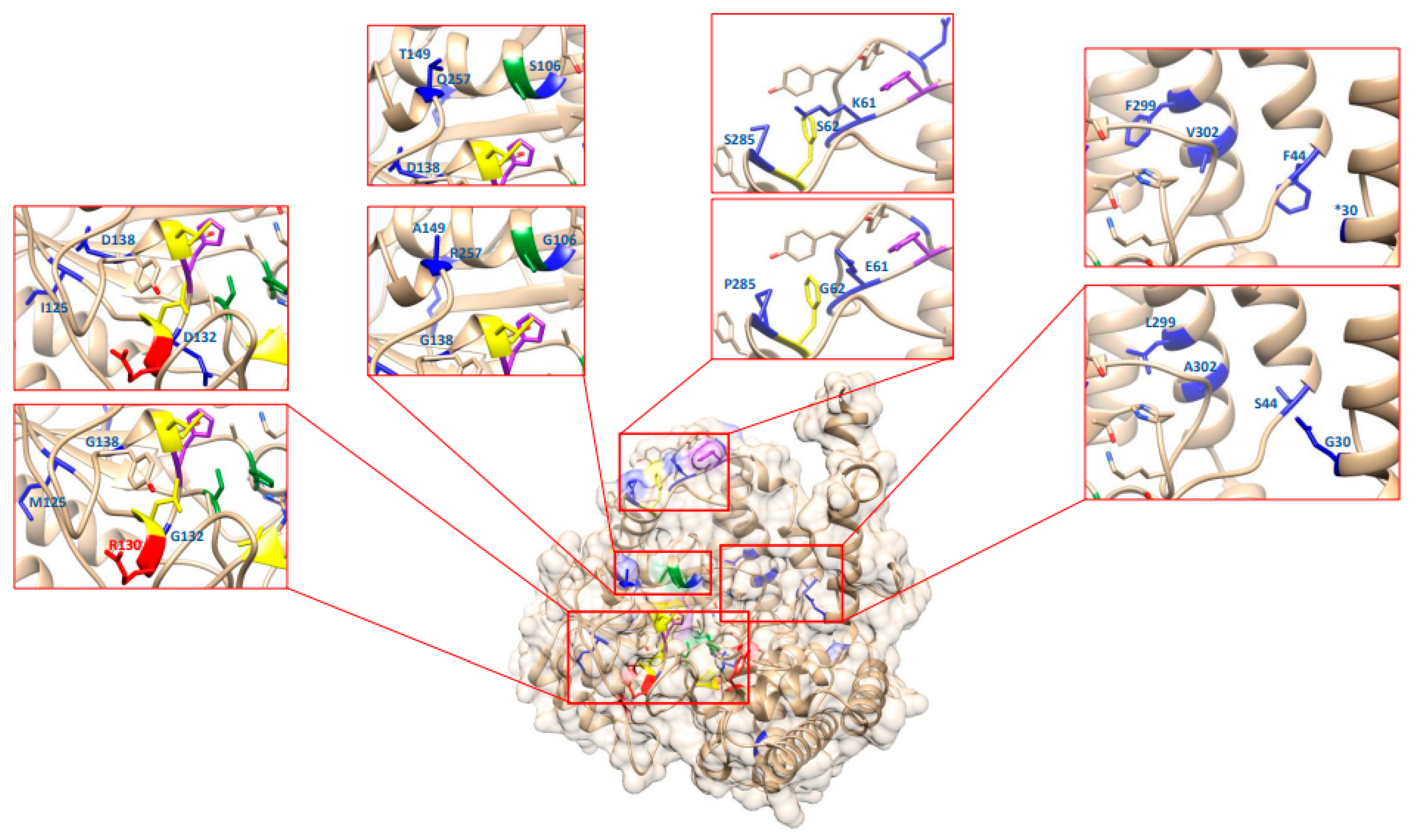
2.9. In silico Analysis of the GmSHMT08 THF Cofactor Binding Sites and PLP Cofactor Binding and Catalysis Sites
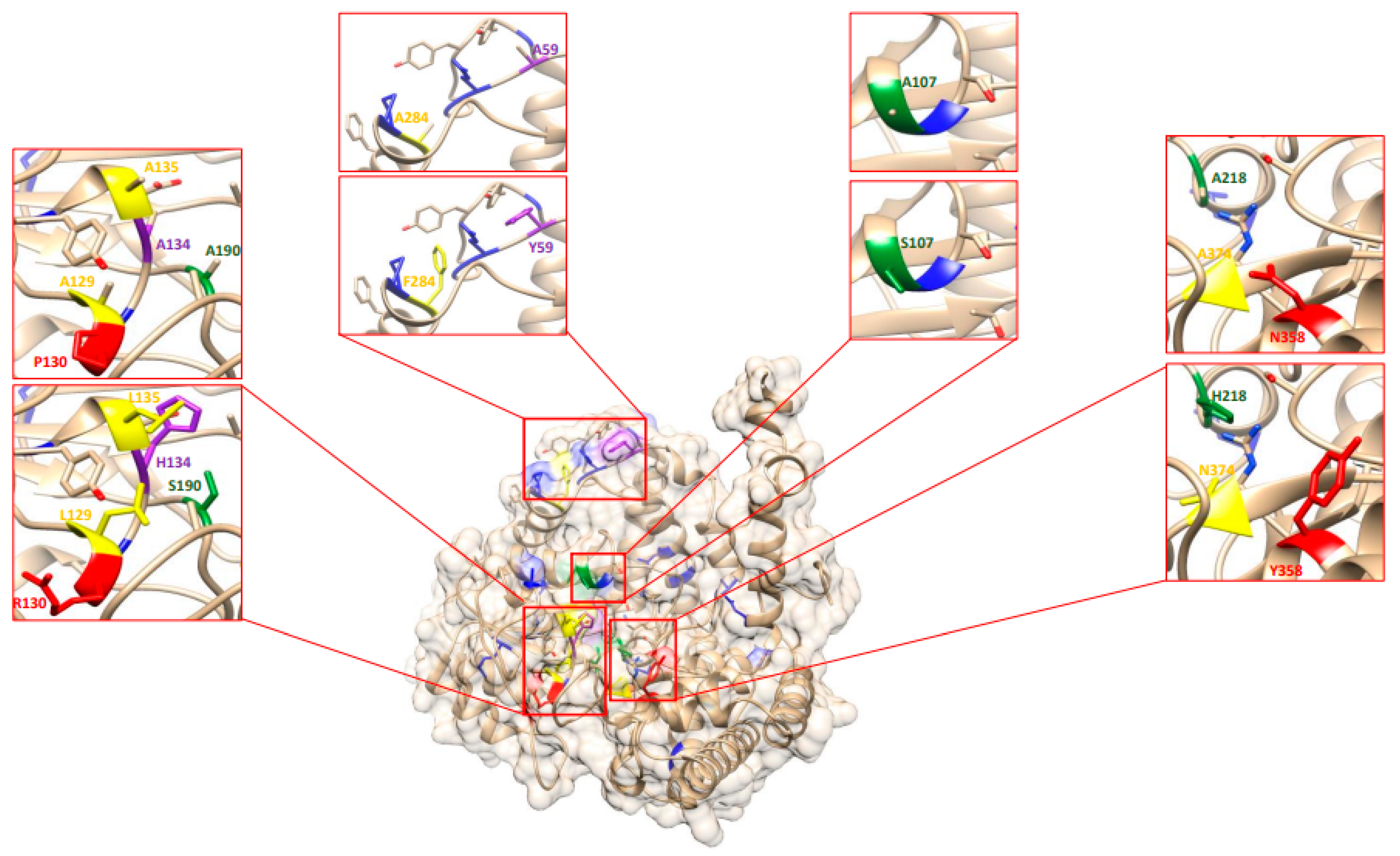
2.10. Re-Analysis of the EMS-Induced GmSHMT08 Mutations Reveal Their Potential Impact on PLP/THF Cofactor Binding and Catalysis
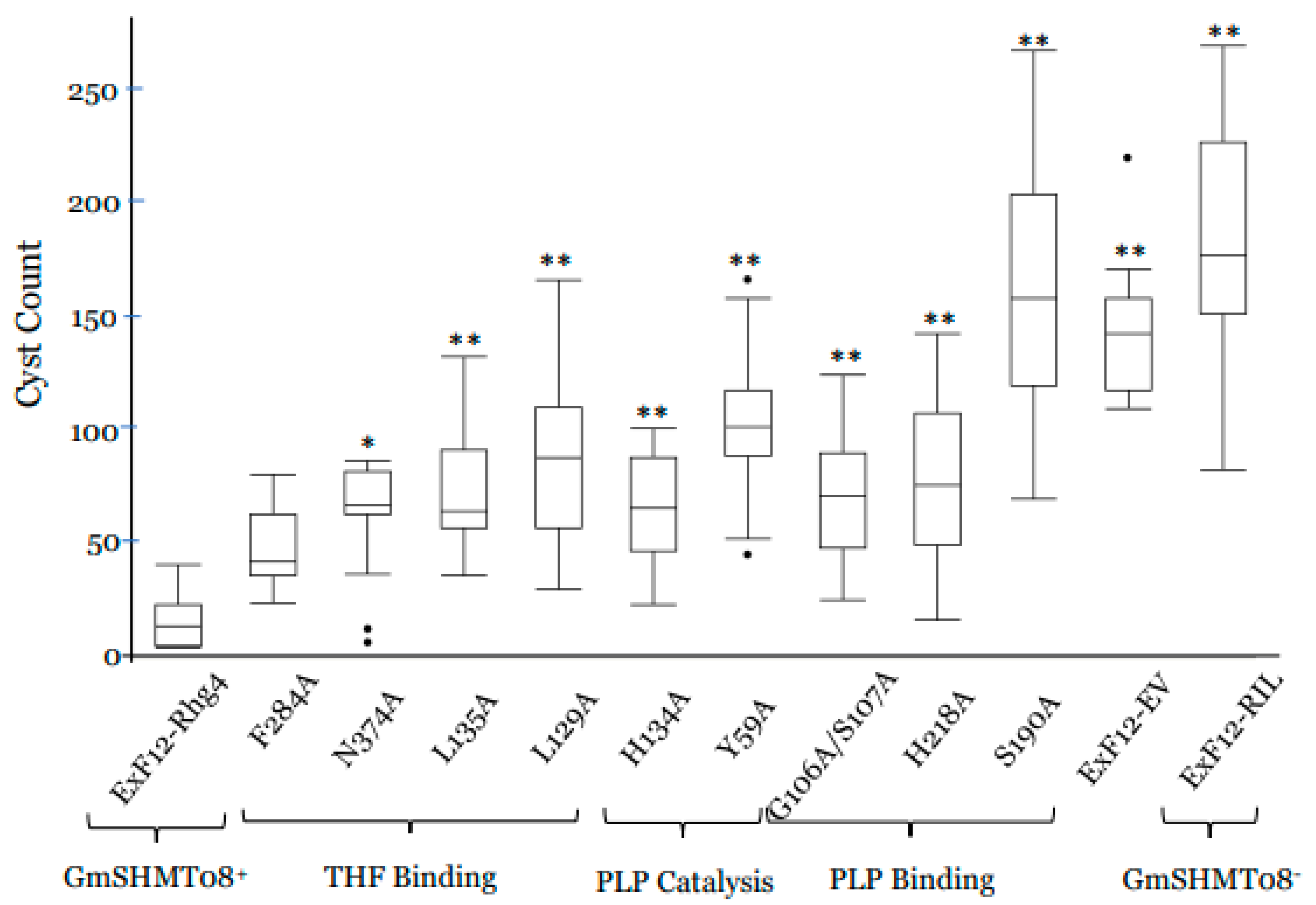
2.11. Functional Validation of the GmSHMT08 THF Cofactor Binding Sites and Their Role in SCN Resistance
2.12. Functional Validation of the GmSHMT08 PLP Cofactor Binding and Catalysis Sites Points to Their Involvement in SCN Resistance
3. Discussion
4. Material and Methods
4.1. Protein Extractions and Immunoprecipitation Using GmSHMT08 Antibodies
4.2. Mass Spectrometry Analysis
4.3. RNA-seq Library Preparation and Analysis
4.4. Cloning the Forrest GmSHMT08 WT and Site Directed Mutagenesis
4.5. Genotyping of ExF RIL Population
4.6. Transgenic Soybean Composite Hairy Root
4.7. GmSHMT08 TILLING Mutants
4.8. Modeling of GmSHMT08 Protein, PLP and THF Cofactor Sites
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shahbandeh, M. Production Value of Soybeans in the U.S. 2000–2020; Statista: Hamburg, Germany, 2021. [Google Scholar]
- Liu, S.; Kandoth, P.K.; Warren, S.D.; Yeckel, G.; Heinz, R.; Alden, J.; Yang, C.; Jamai, A.; El-Mellouki, T.; Juvale, P.S.; et al. A soybean cyst nematode resistance gene points to a new mechanism of plant resistance to pathogens. Nature 2012, 492, 256–260. [Google Scholar] [CrossRef]
- Mitchum, M.G. Soybean Resistance to the Soybean Cyst Nematode Heterodera glycines: An Update. Phytopathology 2016, 106, 1444–1450. [Google Scholar] [CrossRef]
- Howland, A.; Monnig, N.; Mathesius, J.; Nathan, M.; Mitchum, M.G. Survey of Heterodera glycines Population Densities and Virulence Phenotypes During 2015–2016 in Missouri. Plant Dis. 2018, 102, 2407–2410. [Google Scholar] [CrossRef] [PubMed]
- Meksem, K.; Pantazopoulos, P.; Njiti, V.N.; Hyten, L.D.; Arelli, P.R.; Lightfoot, D.A. ’Forrest’ resistance to the soybean cyst nematode is bigenic: Saturation mapping of the Rhg1and Rhg4 loci. Theor. Appl. Genet. 2001, 103, 710–717. [Google Scholar] [CrossRef]
- Kandoth, P.K.; Liu, S.; Prenger, E.; Ludwig, A.; Lakhssassi, N.; Heinz, R.; Zhou, Z.; Howland, A.; Gunther, J.; Eidson, S.; et al. Systematic Mutagenesis of Serine Hydroxymethyltransferase Reveals an Essential Role in Nematode Resistance. Plant Physiol. 2017, 175, 1370–1380. [Google Scholar] [CrossRef] [PubMed]
- Lakhssassi, N.; Patil, G.; Piya, S.; Zhou, Z.; Baharlouei, A.; Kassem, M.A.; Lightfoot, D.A.; Hewezi, T.; Barakat, A.; Nguyen, H.T.; et al. Genome reorganization of the GmSHMT gene family in soybean showed a lack of functional redundancy in resistance to soybean cyst nematode. Sci. Rep. 2019, 9, 1506. [Google Scholar] [CrossRef] [PubMed]
- Patil, G.B.; Lakhssassi, N.; Wan, J.; Song, L.; Zhou, Z.; Klepadlo, M.; Vuong, T.D.; Stec, A.O.; Kahil, S.S.; Colantonio, V.; et al. Whole-genome re-sequencing reveals the impact of the interaction of copy number variants of the rhg1 and Rhg4 genes on broad-based resistance to soybean cyst nematode. Plant Biotechnol. J. 2019, 17, 1595–1611. [Google Scholar] [CrossRef]
- Stover, P.; Schirch, V. Serine hydroxymethyltransferase catalyzes the hydrolysis of 5,10-methenyltetrahydrofolate to 5-formyltetrahydrofolate. J. Biol. Chem. 1990, 265, 14227–14233. [Google Scholar] [CrossRef]
- Hanson, A.D.; Gage, D.A.; Shachar-Hill, Y. Plant one-carbon metabolism and its engineering. Trends Plant Sci. 2000, 5, 206–213. [Google Scholar] [CrossRef]
- Appaji Rao, N.; Ambili, M.; Jala, V.R.; Subramanya, H.S.; Savithri, H.S. Structure-function relationship in serine hydroxymethyltransferase. Biochim. Biophys. Acta 2003, 1647, 24–29. [Google Scholar] [CrossRef]
- Schirch, L. Serine hydroxymethyltransferase. Adv. Enzym. Relat. Areas Mol. Biol. 1982, 53, 83–112. [Google Scholar] [CrossRef]
- Shuvalov, O.; Petukhov, A.; Daks, A.; Fedorova, O.; Vasileva, E.; Barlev, N.A. One-carbon metabolism and nucleotide biosynthesis as attractive targets for anticancer therapy. Oncotarget 2017, 8, 23955–23977. [Google Scholar] [CrossRef]
- Parra, M.; Stahl, S.; Hellmann, H. Vitamin B₆ and Its Role in Cell Metabolism and Physiology. Cells 2018, 7, 84. [Google Scholar] [CrossRef] [PubMed]
- Narkewicz, M.R.; SAULS, S.D.; TJOA, S.S.; TENG, C.; FENNESSEY, P.V. Evidence for intracellular partitioning of serine and glycine metabolism in Chinese hamster ovary cells. Biochem. J. 1996, 313, 991–996. [Google Scholar] [CrossRef]
- Voll, L.M.; Jamai, A.; Renneé, P.; Voll, H.; McClung, C.R.; Weber, A.P.M. The Photorespiratory Arabidopsis shm1 Mutant Is Deficient in SHM1. Plant Physiol. 2006, 140, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Skibola, C.F.; Smith, M.T.; Hubbard, A.; Shane, B.; Roberts, A.C.; Law, G.R.; Rollinson, S.; Roman, E.; Cartwright, R.A.; Morgan, G.J. Polymorphisms in the thymidylate synthase and serine hydroxymethyltransferase genes and risk of adult acute lymphocytic leukemia. Blood 2002, 99, 3786–3791. [Google Scholar] [CrossRef]
- Lim, U.; Peng, K.; Shane, B.; Stover, P.J.; Litonjua, A.A.; Weiss, S.T.; Gaziano, J.M.; Strawderman, R.L.; Raiszadeh, F.; Selhub, J.; et al. Polymorphisms in Cytoplasmic Serine Hydroxymethyltransferase and Methylenetetrahydrofolate Reductase Affect the Risk of Cardiovascular Disease in Men. J. Nutr. 2005, 135, 1989–1994. [Google Scholar] [CrossRef][Green Version]
- Amelio, I.; Cutruzzolá, F.; Antonov, A.; Agostini, M.; Melino, G. Serine and glycine metabolism in cancer. Trends Biochem. Sci 2014, 39, 191–198. [Google Scholar] [CrossRef]
- Korasick, D.A.; Kandoth, P.K.; Tanner, J.J.; Mitchum, M.G.; Beamer, L.J. Impaired folate binding of serine hydroxymethyltransferase 8 from soybean underlies resistance to the soybean cyst nematode. J. Biol. Chem. 2020, 295, 3708–3718. [Google Scholar] [CrossRef]
- Lakhssassi, N.; Piya, S.; Bekal, S.; Liu, S.; Zhou, Z.; Bergounioux, C.; Miao, L.; Meksem, J.; Lakhssassi, A.; Jones, K.; et al. A pathogenesis-related protein GmPR08-Bet VI promotes a molecular interaction between the GmSHMT08 and GmSNAP18 in resistance to Heterodera glycines. Plant Biotechnol. J. 2020, 18, 1810–1829. [Google Scholar] [CrossRef]
- Babicki, S.; Arndt, D.; Marcu, A.; Liang, Y.; Grant, J.R.; Maciejewski, A.; Wishart, D.S. Heatmapper: Web-enabled heat mapping for all. Nucleic Acids Res. 2016, 44, W147–W153. [Google Scholar] [CrossRef]
- Lakhssassi, N.; Piya, S.; Knizia, D.; El Baze, A.; Cullen, M.A.; Meksem, J.; Lakhssassi, A.; Hewezi, T.; Meksem, K. Mutations at the Serine Hydroxymethyltransferase Impact its Interaction with a Soluble NSF Attachment Protein and a Pathogenesis-Related Protein in Soybean. Vaccines 2020, 8, 349. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Lasker, K.; Schneidman-Duhovny, D.; Webb, B.; Huang, C.C.; Pettersen, E.F.; Goddard, T.D.; Meng, E.C.; Sali, A.; Ferrin, T.E. UCSF Chimera, MODELLER, and IMP: An integrated modeling system. J. Struct. Biol. 2012, 179, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Kandoth, P.K.; Mitchum, M.G. War of the worms: How plants fight underground attacks. Curr. Opin. Plant Biol. 2013, 16, 457–463. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; You, Y.; Fan, H.; Zhu, X.; Wang, Y.; Duan, Y.; Xuan, Y.; Chen, L. The Role of Sugar Transporter Genes during Early Infection by Root-Knot Nematodes. Int. J. Mol. Sci. 2018, 19, 302. [Google Scholar] [CrossRef]
- Hofmann, J.; Hess, P.H.; Szakasits, D.; Blöchl, A.; Wieczorek, K.; Daxböck-Horvath, S.; Bohlmann, H.; van Bel, A.J.E.; Grundler, F.M.W. Diversity and activity of sugar transporters in nematode-induced root syncytia. J. Exp. Bot. 2009, 60, 3085–3095. [Google Scholar] [CrossRef]
- Sijmons, P.C.; Grundler, F.M.; von Mende, N.; Burrows, P.R.; Wyss, U. Arabidopsis thaliana as a new model host for plant-parasitic nematodes. Plant J. 1991, 1, 245–254. [Google Scholar] [CrossRef]
- Braeckman, B.; Houthoofd, K.; Vanfleteren, J. Intermediary metabolism. In WormBook: The Online Review of C. elegans Biology; WormBook: Pasadena, CA, USA, 2009. [Google Scholar]
- Zečić, A.; Dhondt, I.; Braeckman, B.P. The nutritional requirements of Caenorhabditis elegans. Genes Nutr. 2019, 14, 15. [Google Scholar] [CrossRef]
- Hofmann, J.; Wieczorek, K.; Blöchl, A.; Grundler, F.M. Sucrose supply to nematode-induced syncytia depends on the apoplasmic and symplasmic pathways. J. Exp. Bot. 2007, 58, 1591–1601. [Google Scholar] [CrossRef]
- Hofmann, J.; Szakasits, D.; Blöchl, A.; Sobczak, M.; Daxböck-Horvath, S.; Golinowski, W.; Bohlmann, H.; Grundler, F.M.W. Starch serves as carbohydrate storage in nematode-induced syncytia. Plant Physiol. 2008, 146, 228–235. [Google Scholar] [CrossRef]
- Hofmann, J.; Grundler, F.M. Females and males of root-parasitic cyst nematodes induce different symplasmic connections between their syncytial feeding cells and the phloem in Arabidopsis thaliana. Plant Physiol. Biochem. 2006, 44, 430–433. [Google Scholar] [CrossRef]
- Hoth, S.; Schneidereit, A.; Lauterbach, C.; Scholz-Starke, J.; Sauer, N. Nematode infection triggers the de novo formation of unloading phloem that allows macromolecular trafficking of green fluorescent protein into syncytia. Plant Physiol. 2005, 138, 383–392. [Google Scholar] [CrossRef]
- Hoth, S.; Stadler, R.; Sauer, N.; Hammes, U.Z. Differential vascularization of nematode-induced feeding sites. Proc. Natl. Acad. Sci. USA 2008, 105, 12617–12622. [Google Scholar] [CrossRef]
- Ko, M.P.; Huang, P.Y.; Huang, J.S.; Barker, K.R. The occurrence of phytoferritin and its relationship to effectiveness of soybean nodules. Plant Physiol. 1987, 83, 299–305. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Goheen, S.; Campbell, J.; Donal, P. Nutritional Requirements of Soybean Cyst Nematodes. In Soybean—Pest Resistance; El-Shemy, H., Ed.; IntechOpen: London, UK, 2013. [Google Scholar]
- Sheehan, D.; Meade, G.; Foley, V.M.; Dowd, C.A. Structure, function and evolution of glutathione transferases: Implications for classification of non-mammalian members of an ancient enzyme superfamily. Biochem. J. 2001, 360, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Udomsinprasert, R.; Pongjaroenkit, S.; Wongsantichon, J.; Oakley, A.J.; Prapanthadara, L.-a.; Wilce, M.C.J.; Ketterman, A.J. Identification, characterization and structure of a new Delta class glutathione transferase isoenzyme. Biochem. J. 2005, 388, 763–771. [Google Scholar] [CrossRef]
- Allocati, N.; Federici, L.; Masulli, M.; Di Ilio, C. Glutathione transferases in bacteria. FEBS J. 2009, 276, 58–75. [Google Scholar] [CrossRef]
- Hayes, J.D.; Flanagan, J.U.; Jowsey, I.R. Glutathione transferases. Annu. Rev. Pharmacol. Toxicol. 2005, 45, 51–88. [Google Scholar] [CrossRef] [PubMed]
- Josephy, P.D. Genetic variations in human glutathione transferase enzymes: Significance for pharmacology and toxicology. Hum. Genom. Proteom. 2010, 2010, 876940. [Google Scholar] [CrossRef]
- Kandoth, P.K.; Ithal, N.; Recknor, J.; Maier, T.; Nettleton, D.; Baum, T.J.; Mitchum, M.G. The Soybean Rhg1 locus for resistance to the soybean cyst nematode Heterodera glycines regulates the expression of a large number of stress- and defense-related genes in degenerating feeding cells. Plant Physiol. 2011, 155, 1960–1975. [Google Scholar] [CrossRef] [PubMed]
- Vivoli, M.; Angelucci, F.; Ilari, A.; Morea, V.; Angelaccio, S.; di Salvo, M.L.; Contestabile, R. Role of a conserved active site cation-pi interaction in Escherichia coli serine hydroxymethyltransferase. Biochemistry 2009, 48, 12034–12046. [Google Scholar] [CrossRef] [PubMed]
- De Rossi, M.C.; Levi, V.; Bruno, L. Retraction of rod-like mitochondria during microtubule-dependent transport. Biosci. Rep. 2018, 38, BSR20180208. [Google Scholar] [CrossRef] [PubMed]
- Melkov, A.; Abdu, U. Regulation of long-distance transport of mitochondria along microtubules. Cell Mol. Life Sci. 2018, 75, 163–176. [Google Scholar] [CrossRef]
- Rostovtseva, T.K.; Gurnev, P.A.; Chen, M.Y.; Bezrukov, S.M. Membrane lipid composition regulates tubulin interaction with mitochondrial voltage-dependent anion channel. J. Biol. Chem. 2012, 287, 29589–29598. [Google Scholar] [CrossRef]
- Kuznetsov, A.V.; Javadov, S.; Grimm, M.; Margreiter, R.; Ausserlechner, M.J.; Hagenbuchner, J. Crosstalk between Mitochondria and Cytoskeleton in Cardiac Cells. Cells 2020, 9, 222. [Google Scholar] [CrossRef]
- West, A.P.; Shadel, G.S.; Ghosh, S. Mitochondria in innate immune responses. Nat. Rev. Immunol. 2011, 11, 389–402. [Google Scholar] [CrossRef]
- Lisowski, P.; Kannan, P.; Mlody, B.; Prigione, A. Mitochondria and the dynamic control of stem cell homeostasis. EMBO Rep. 2018, 19, e45432. [Google Scholar] [CrossRef] [PubMed]
- Dan Dunn, J.; Alvarez, L.A.; Zhang, X.; Soldati, T. Reactive oxygen species and mitochondria: A nexus of cellular homeostasis. Redox Biol. 2015, 6, 472–485. [Google Scholar] [CrossRef]
- Li, Z.; Ding, B.; Zhou, X.; Wang, G.L. The Rice Dynamin-Related Protein OsDRP1E Negatively Regulates Programmed Cell Death by Controlling the Release of Cytochrome c from Mitochondria. PLoS Pathog. 2017, 13, e1006157. [Google Scholar] [CrossRef]
- Tang, C.; Deng, L.; Chang, D.; Chen, S.; Wang, X.; Kang, Z. TaADF3, an Actin-Depolymerizing Factor, Negatively Modulates Wheat Resistance Against Puccinia striiformis. Front. Plant Sci. 2016, 6, 1214. [Google Scholar] [CrossRef] [PubMed]
- Day, B.; Henty, J.L.; Porter, K.J.; Staiger, C.J. The Pathogen-Actin Connection: A Platform for Defense Signaling in Plants. Annu. Rev. Phytopathol. 2011, 49, 483–506. [Google Scholar] [CrossRef]
- Miklis, M.; Consonni, C.; Bhat, R.A.; Lipka, V.; Schulze-Lefert, P.; Panstruga, R. Barley MLO Modulates Actin-Dependent and Actin-Independent Antifungal Defense Pathways at the Cell Periphery. Plant Physiol. 2007, 144, 1132–1143. [Google Scholar] [CrossRef] [PubMed]
- Singh, H.; Kaur, K.; Singh, M.; Kaur, G.; Singh, P. Plant Cyclophilins: Multifaceted Proteins With Versatile Roles. Front. Plant Sci. 2020, 11, 585212. [Google Scholar] [CrossRef] [PubMed]
- Mainali, H.R.; Vadivel, A.K.A.; Li, X.; Gijzen, M.; Dhaubhadel, S. Soybean cyclophilin GmCYP1 interacts with an isoflavonoid regulator GmMYB176. Sci. Rep. 2017, 7, 39550. [Google Scholar] [CrossRef] [PubMed]
- Piya, S.; Kihm, C.; Rice, J.H.; Baum, T.J.; Hewezi, T. Cooperative Regulatory Functions of miR858 and MYB83 during Cyst Nematode Parasitism. Plant Physiol. 2017, 174, 1897–1912. [Google Scholar] [CrossRef] [PubMed]
- Lei, P.; Qi, N.; Zhou, Y.; Wang, Y.; Zhu, X.; Xuan, Y.; Liu, X.; Fan, H.; Chen, L.; Duan, Y. Soybean miR159-GmMYB33 Regulatory Network Involved in Gibberellin-Modulated Resistance to Heterodera glycines. Int. J. Mol. Sci. 2021, 22, 13172. [Google Scholar] [CrossRef]
- Bane, F.T.; Bannon, J.H.; Pennington, S.R.; Campiani, G.; Williams, D.C.; Zisterer, D.M.; Mc Gee, M.M. The Microtubule-Targeting Agents, PBOX-6 [Pyrrolobenzoxazepine 7-[(dimethylcarbamoyl)oxy]-6-(2-naphthyl)pyrrolo-[2,1-d] (1,5)-benzoxazepine] and Paclitaxel, Induce Nucleocytoplasmic Redistribution of the Peptidyl-Prolyl Isomerases, Cyclophilin A and Pin1, in Malignant Hematopoietic Cells. J. Pharmacol. Exp. Ther. 2009, 329, 38. [Google Scholar] [CrossRef] [PubMed]
- Caillaud, M.-C.; Lecomte, P.; Jammes, F.; Quentin, M.; Pagnotta, S.; Andrio, E.; de Almeida Engler, J.; Marfaing, N.; Gounon, P.; Abad, P.; et al. MAP65-3 microtubule-associated protein is essential for nematode-induced giant cell ontogenesis in Arabidopsis. Plant Cell 2008, 20, 423–437. [Google Scholar] [CrossRef]
- Lakhssassi, N.; Liu, S.; Bekal, S.; Zhou, Z.; Colantonio, V.; Lambert, K.; Barakat, A.; Meksem, K. Characterization of the Soluble NSF Attachment Protein gene family identifies two members involved in additive resistance to a plant pathogen. Sci. Rep. 2017, 7, 45226. [Google Scholar] [CrossRef]
- Moraes, J.G.N.; Behura, S.K.; Bishop, J.V.; Hansen, T.R.; Geary, T.W.; Spencer, T.E. Analysis of the uterine lumen in fertility-classified heifers: II. Proteins and metabolites†. Biol. Reprod. 2020, 102, 571–587. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Dobin, A.; Gingeras, T.R. Mapping RNA-seq Reads with STAR. Curr. Protoc. Bioinform. 2015, 51, 11.14.11–11.14.19. [Google Scholar] [CrossRef]
- Trapnell, C.; Pachter, L.; Salzberg, S.L. TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq--a Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera--a visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef]
- Leon, J.; Lawton, M.A.; Raskin, I. Hydrogen Peroxide Stimulates Salicylic Acid Biosynthesis in Tobacco. Plant Physiol. 1995, 108, 1673–1678. [Google Scholar] [CrossRef] [PubMed]
- Moreno, J.I.; Martín, R.; Castresana, C. Arabidopsis SHMT1, a serine hydroxymethyltransferase that functions in the photorespiratory pathway influences resistance to biotic and abiotic stress. Plant J. 2005, 41, 451–463. [Google Scholar] [CrossRef]
- Melillo, M.T.; Leonetti, P.; Bongiovanni, M.; Castagnone-Sereno, P.; Bleve-Zacheo, T. Modulation of reactive oxygen species activities and H2O2 accumulation during compatible and incompatible tomato-root-knot nematode interactions. New Phytol. 2006, 170, 501–512. [Google Scholar] [CrossRef]
- Siddique, S.; Matera, C.; Radakovic, Z.S.; Hasan, M.S.; Gutbrod, P.; Rozanska, E.; Sobczak, M.; Torres, M.A.; Grundler, F.M. Parasitic worms stimulate host NADPH oxidases to produce reactive oxygen species that limit plant cell death and promote infection. Sci. Signal. 2014, 7, ra33. [Google Scholar] [CrossRef] [PubMed]
- Hewezi, T. Cellular Signaling Pathways and Posttranslational Modifications Mediated by Nematode Effector Proteins. Plant Physiol. 2015, 169, 1018–1026. [Google Scholar] [CrossRef]
- Webb, D.M.; Baltazar, B.M.; Rao-Arelli, A.P.; Schupp, J.; Clayton, K.; Keim, P.; Beavis, W.D. Genetic mapping of soybean cyst nematode race-3 resistance loci in the soybean PI 437.654. Theoret. Appl. Genet. 1995, 91, 574–581. [Google Scholar] [CrossRef]
- Ferreira, M.; Cervigni, G.; Ferreira, A.; Schuster, I.; Santana, F.; Pereira, W.; Barros, E.; Moreira, M. QTL for resistance to soybean cyst nematode races 3, 9, and 14 in cultivar Hartwig. Pesq. Agropec. Bras. 2011, 46, 420–428. [Google Scholar] [CrossRef]
- Qiu, B.X.; Arelli, P.R.; Sleper, D.A. RFLP markers associated with soybean cyst nematode resistance and seed composition in a ‘Peking’ × ‘Essex’ population. Theor. Appl. Genet. 1999, 98, 356–364. [Google Scholar] [CrossRef]
- Chang, W.; Dong, L.; Wang, Z.; Hu, H.; Han, Y.; Teng, W.; Zhang, H.; Guo, M.; Li, W. QTL underlying resistance to two HG types of Heterodera glycines found in soybean cultivar ‘L-10′. BMC Genom. 2011, 12, 233. [Google Scholar] [CrossRef]
- Yue, P.; Arelli, P.; Sleper, D. Molecular characterization of resistance to Heterodera glycines in soybean PI 438489B. Theor. Appl. Genet. 2001, 102, 921–928. [Google Scholar] [CrossRef]
- Abdelmajid, K.; Ramos, L.; Hyten, D.; Bond, J.; Bendahmane, A.; Arelli, P.; Njiti, V.; Cianzio, S.; Kantartzi, S.; Meksem, K. Quantitative Trait Loci (QTL) that Underlie SCN Resistance in Soybean [Glycine max (L.) Merr.] PI438489B by ‘Hamilton’ Recombinant Inbred Line (RIL) Population. Atlas J. Plant Biol. 2014, 1, 29–38. [Google Scholar] [CrossRef][Green Version]
- Concibido, V.C.; Denny, R.L.; Boutin, S.R.; Hautea, R.; Orf, J.H.; Young, N.D. DNA marker analysis of loci underlying resistance to soybean cyst nematode (Heterodera glycines Ichinohe). Crop Sci. 1994, 34, 240–246. [Google Scholar] [CrossRef]
- Guo, B.; Sleper, D.; Nguyen, H.; Arelli, P.; Shannon, J. Quantitative Trait Loci Underlying Resistance to Three Soybean Cyst Nematode Populations in Soybean PI 404198A. Crop Sci. 2006, 46, 224–233. [Google Scholar] [CrossRef]
- Wu, X.; Blake, S.; Sleper, D.; Shannon, J.; Cregan, P.; Nguyen, H. QTL, additive and epistatic effects for SCN resistance in PI 437654. Theor. Appl. Genet. 2009, 118, 1093–1105. [Google Scholar] [CrossRef]
- Chang, S.J.C.; Doubler, T.W.; Kilo, V.Y.; Abu-Thredeih, J.; Prabhu, R.; Freire, V.; Suttner, R.; Klein, J.; Schmidt, M.E.; Gibson, P.T.; et al. Association of loci underlying field-resistance to soybean sudden-death syndrome (SDS) and cyst-nematode (SCN) RACE-3. Crop Sci. 1997, 37, 966–971. [Google Scholar] [CrossRef]
- Vuong, T.; Sleper, D.; Shannon, J.; Nguyen, H. Novel quantitative trait loci for broad-based resistance to soybean cyst nematode (Heterodera glycines Ichinohe) in soybean PI 567516C. Theor. Appl. Genet. 2010, 121, 1253–1266. [Google Scholar] [CrossRef]
- Arriagada, O.; Mora, F.; Dellarosa, J.; Ferreira, M.; Cervigni, G.; Schuster, I. Bayesian mapping of quantitative trait loci (QTL) controlling soybean cyst nematode resistant. Euphytica 2012, 186, 907–917. [Google Scholar] [CrossRef]
- Jiao, Y.; Vuong, T.; Liu, Y.; Meinhardt, C.; Liu, Y.; Joshi, T.; Cregan, P.; Xu, D.; Shannon, J.; Nguyen, H. Identification and evaluation of quantitative trait loci underlying resistance to multiple HG types of soybean cyst nematode. Theor. Appl. Genet. 2015, 128, 15–23. [Google Scholar] [CrossRef]
- Silva, M.; Schuster, I.; Silva, J.; Ferreira, A.; Barros, E.; Moreira, M. Validation of microsatellite markers for assisted selection of soybean resistance to cyst nematode races 3 and 14. Pesq. Agropec. Bras. 2007, 42, 1143–1150. [Google Scholar] [CrossRef]
- Vaghchhipawala, Z.; Bassuner, R.; Clayton, K.; Lewers, K.S.; Shoemaker, R.C.; Mackenzie, S. Modulations in gene expression and mapping of genes associated with cyst nematode infection of soybean. Mol. Plant Microbe Interac. 2001, 14, 42–54. [Google Scholar] [CrossRef]
- Valdes-Lopez, O.; Thibivilliers, S.; Qiu, J.; Xu, W.; Nguyen, T.; Libault, M.; Le, B.; Goldberg, R.; Hill, C.; Hartman, G.; et al. Identification of Quantitative Trait Loci Controlling Gene Expression during the Innate Immunity Response of Soybean. Plant Physiol. 2011, 157, 1975–1986. [Google Scholar] [CrossRef] [PubMed]
- Carlson, S.R.; Diers, B.W.; Kabelka, E.A. Localization of Two Loci that Confer Resistance to Soybean Cyst Nematode from Glycine soja PI 468916. Crop Sci. 2005, 45, 2473–2481. [Google Scholar]
- Yue, P.; Sleper, D.A.; Arelli, P.R. Mapping Resistance to Multiple Races of Heterodera glycines in Soybean PI 89772. Crop Sci. 2001, 41, 1589–1595. [Google Scholar] [CrossRef]
- Guo, B.; Sleper, D.; Arelli, P.; Shannon, J.; Nguyen, H. Identification of QTLs associated with resistance to soybean cyst nematode races 2, 3 and 5 in soybean PI 90763. Theor. Appl. Genet. 2005, 111, 965–971. [Google Scholar] [CrossRef]
- Swaminathan, S.; Abeysekara, N.; Knight, J.; Liu, M.; Dong, J.; Hudson, M.; Bhattacharyya, M.; Cianzio, S. Mapping of new quantitative trait loci for sudden death syndrome and soybean cyst nematode resistance in two soybean populations. Theor. Appl. Genet. 2018, 131, 1047–1062. [Google Scholar] [CrossRef]
- Vuong, T.; Sleper, D.; Shannon, J.; Wu, X.; Nguyen, H. Confirmation of quantitative trait loci for resistance to multiple-HG types of soybean cyst nematode (Heterodera glycines Ichinohe). Euphytica 2011, 181, 101–113. [Google Scholar] [CrossRef]
- Mahalingam, R.; Skorupska, H.T. DNA markers for resistance to Heterodera glycines I. race 3 in soybean cultivar Peking. Breed. Sci. 1995, 45, 435–443. [Google Scholar]
- Wang, D.; Diers, B.W.; Arelli, P.R.; Shoemaker, R.C. Loci underlying resistance to Race 3 of soybean cyst nematode in Glycine soja plant introduction 468916. Theor. Appl. Genet. 2001, 103, 561–566. [Google Scholar] [CrossRef]
- Kazi, S.; Shultz, J.; Afzal, J.; Hashmi, R.; Jasim, M.; Bond, J.; Arelli, P.; Lightfoot, D. Iso-lines and inbred-lines confirmed loci that underlie resistance from cultivar ‘Hartwig’ to three soybean cyst nematode populations. Theor. Appl. Genet. 2010, 120, 633–644. [Google Scholar] [CrossRef][Green Version]
- Schuster, I.; Abdelnoor, R.V.; Marin, S.R.R.; Carvalho, V.P.; Kiihl, R.A.S.; Silva, J.F.V.; Sediyama, C.S.; Barros, E.G.; Moreira, M.A. Identification of a new major QTL associated with resistance to soybean cyst nematode (Heterodera glycines). Theor. Appl. Genet. 2001, 102, 91–96. [Google Scholar] [CrossRef]
- Winter, S.; Shelp, B.; Anderson, T.; Welacky, T.; Rajcan, I. QTL associated with horizontal resistance to soybean cyst nematode in Glycine soja PI464925B. Theor. Appl. Genet. 2007, 114, 461–472. [Google Scholar] [CrossRef]
- Prabhu, R.R.; Njiti, V.N.; Bell-Johnson, B.; Johnson, J.E.; Schmidt, M.E.; Klein, J.H.; Lightfoot, D.A. Selecting soybean cultivars for dual resistance to soybean cyst nematode and sudden death syndrome using two DNA markers. Crop Sci. 1999, 39, 982–987. [Google Scholar] [CrossRef]
- Concibido, V.C.; Lange, D.A.; Denny, R.L.; Orf, J.H.; Young, N.D. Genome mapping of soybean cyst nematode resistance genes in Peking, PI90763, and PI88788 using DNA markers. Crop Sci. 1997, 37, 258–264. [Google Scholar] [CrossRef]
- Vierling, R.A.; Faghihi, J.; Ferris, V.R.; Ferris, J.M. Association of RFLP markers with loci conferring broad-based resistance to the soybean cyst nematode (Heterodera glycines). Theor. Appl. Genet. 1996, 92, 83–86. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Njiti, V.N.; Meksem, K.; Iqbal, M.J.; Triwitayakorn, K.; Kassem, M.A.; Davis, G.T.; Schmidt, M.E.; Lightfoot, D.A. Quantitative trait loci in Two Soybean Recombinant Inbred Line Populations Segregating for Yield and Disease Resistance. Crop Sci. 2002, 42, 271–277. [Google Scholar] [CrossRef] [PubMed]
- Ferdous, S.; Watanabe, S.; Suzuki-Orihara, C.; Tanaka, Y.; Kamiya, M.; Yamanaka, N.; Harada, K. QTL Analysis of Resistance to Soybean Cyst Nematode Race 3 in Soybean Cultivar Toyomusume. Breed. Sci. 2006, 56, 155–163. [Google Scholar] [CrossRef][Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lakhssassi, N.; Knizia, D.; El Baze, A.; Lakhssassi, A.; Meksem, J.; Meksem, K. Proteomic, Transcriptomic, Mutational, and Functional Assays Reveal the Involvement of Both THF and PLP Sites at the GmSHMT08 in Resistance to Soybean Cyst Nematode. Int. J. Mol. Sci. 2022, 23, 11278. https://doi.org/10.3390/ijms231911278
Lakhssassi N, Knizia D, El Baze A, Lakhssassi A, Meksem J, Meksem K. Proteomic, Transcriptomic, Mutational, and Functional Assays Reveal the Involvement of Both THF and PLP Sites at the GmSHMT08 in Resistance to Soybean Cyst Nematode. International Journal of Molecular Sciences. 2022; 23(19):11278. https://doi.org/10.3390/ijms231911278
Chicago/Turabian StyleLakhssassi, Naoufal, Dounya Knizia, Abdelhalim El Baze, Aicha Lakhssassi, Jonas Meksem, and Khalid Meksem. 2022. "Proteomic, Transcriptomic, Mutational, and Functional Assays Reveal the Involvement of Both THF and PLP Sites at the GmSHMT08 in Resistance to Soybean Cyst Nematode" International Journal of Molecular Sciences 23, no. 19: 11278. https://doi.org/10.3390/ijms231911278
APA StyleLakhssassi, N., Knizia, D., El Baze, A., Lakhssassi, A., Meksem, J., & Meksem, K. (2022). Proteomic, Transcriptomic, Mutational, and Functional Assays Reveal the Involvement of Both THF and PLP Sites at the GmSHMT08 in Resistance to Soybean Cyst Nematode. International Journal of Molecular Sciences, 23(19), 11278. https://doi.org/10.3390/ijms231911278