

Interplay between Prostate Cancer and Adipose Microenvironment: A Complex and Flexible Scenario

 ,
,  and
and {kind=link}
{kind=link}
Abstract
:1. Introduction
2. Cell Players
2.1. PCa Cells
2.2. Adipocytes and Cancer-Associated-Adipocytes (CAA)
2.3. Other Cells of the TME
3. Molecular Players
3.1. Lipids
3.2. Hormones
3.3. Chemokines, Cytokines, and Proteases
3.4. Persistent Organic Pollutants (POPs)
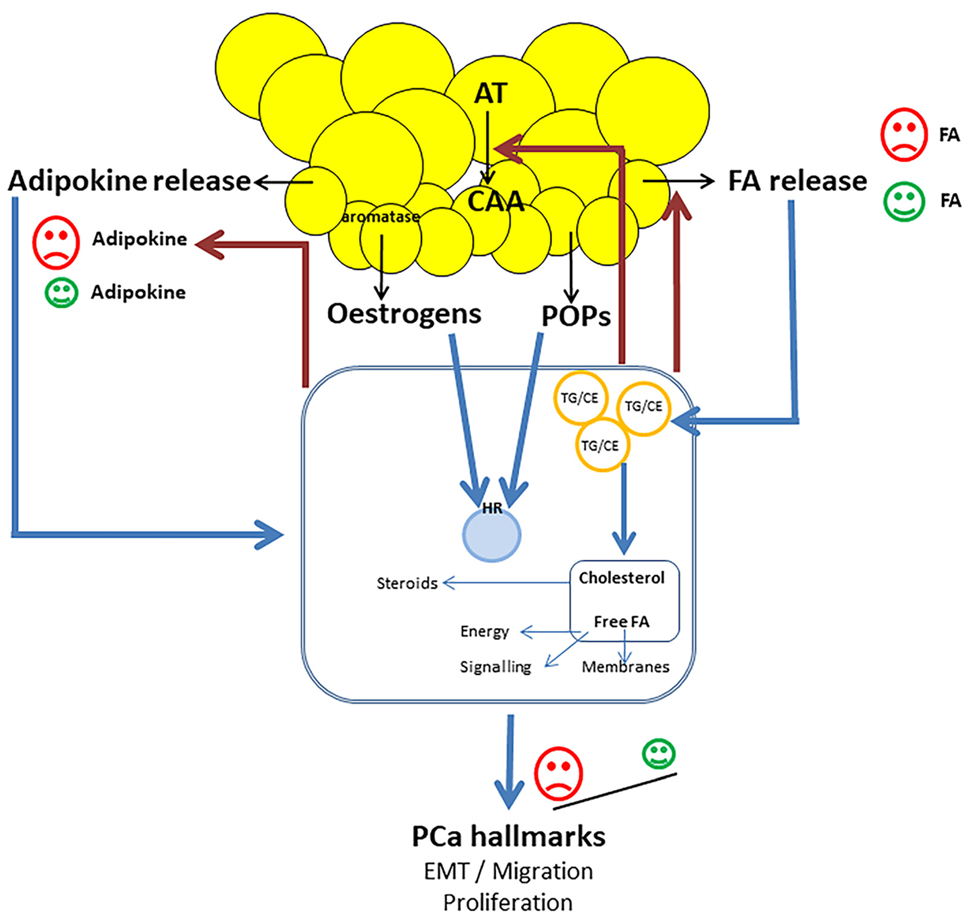
4. Scenario of the Interplay
4.1. On PCa Cells
4.2. On Adipocytes
5. Factors That Can Modulate the Scenario
5.1. Obesity
5.2. Genetic Factors
5.3. Environmental Factors
6. Concluding Remarks
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ADRP | adipocyte differentiation-related protein |
| AR | androgen receptor |
| AT | adipose tissue |
| BMAd | bone marrow adipocyte |
| BMI | body mass index |
| CAA | cancer-associated adipocyte |
| CAF | cancer-associated-fibroblast |
| CE | cholesterol esters |
| CM | conditioned medium |
| EMT | epithelial to mesenchymal transition |
| EPA | ecosapentaenoic acid |
| ER-α | estrogen receptor alpha |
| ER-β | estrogen receptor beta |
| FA | fatty acid |
| FABP | fatty acid binding protein |
| HB-EGF | heparin-binding epidermal growth factor-like growth factor |
| HIF1α | Hypoxia Inducible Factor 1α |
| IL-6 | interleukin-6 |
| LA | linoleic acid |
| LD | lipid droplet |
| MMP | matrix metalloproteinase |
| MSC | mesenchymal stem cell |
| NED | neuroendocrine differentiation |
| NOX5 | NADPH oxidase 5 |
| OCP | organochlorine pesticide |
| PCa | prostate cancer |
| PCB | polychlorobiphenyl |
| POP | persistent organic pollutant |
| PPARγ | peroxisome proliferative-activated receptor γ |
| PPAT | periprostatic adipose tissue |
| PUFA | polyunsaturated fatty acid |
| PXR | pregnane X receptor |
| ROS | reactive oxygen species |
| TAG | triacylglycerol |
| TG | triglycerides |
| TME | tumor microenvironment |
| VEGF | vascular endothelial growth factor |
| WAT | white adipose tissue |
References
- Zhou, C.K.; Check, D.P.; Lortet-Tieulent, J.; Laversanne, M.; Jemal, A.; Ferlay, J.; Bray, F.; Cook, M.B.; Devesa, S.S. Prostate Cancer Incidence in 43 Populations Worldwide: An Analysis of Time Trends Overall and by Age Group. Int. J. Cancer 2016, 138, 1388–1400. [Google Scholar] [CrossRef] [PubMed]
- Sacca, P.A.; Calvo, J.C. Periprostatic Adipose Tissue Microenvironment: Metabolic and Hormonal Pathways During Prostate Cancer Progression. Front. Endocrinol. 2022, 13, 863027. [Google Scholar] [CrossRef] [PubMed]
- Nassar, Z.D.; Aref, A.T.; Miladinovic, D.; Mah, C.Y.; Raj, G.V.; Hoy, A.J.; Butler, L.M. Peri-prostatic Adipose Tissue: The Metabolic Microenvironment of Prostate Cancer. BJU Int. 2018, 121, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Dominiak, A.; Chełstowska, B.; Olejarz, W.; Nowicka, G. Communication in the Cancer Microenvironment as a Target for Therapeutic Interventions. Cancers 2020, 12, 1232. [Google Scholar] [CrossRef] [PubMed]
- Bader, D.A.; McGuire, S.E. Tumour Metabolism and Its Unique Properties in Prostate Adenocarcinoma. Nat. Rev. Urol. 2020, 17, 214–231. [Google Scholar] [CrossRef]
- Laurent, V.; Toulet, A.; Attané, C.; Milhas, D.; Dauvillier, S.; Zaidi, F.; Clement, E.; Cinato, M.; Le Gonidec, S.; Guérard, A.; et al. Periprostatic Adipose Tissue Favors Prostate Cancer Cell Invasion in an Obesity-Dependent Manner: Role of Oxidative Stress. Mol. Cancer Res. 2019, 17, 821–835. [Google Scholar] [CrossRef]
- Diedrich, J.D.; Rajagurubandara, E.; Herroon, M.K.; Mahapatra, G.; Hüttemann, M.; Podgorski, I. Bone Marrow Adipocytes Promote the Warburg Phenotype in Metastatic Prostate Tumors via HIF-1α Activation. Oncotarget 2016, 7, 64854–64877. [Google Scholar] [CrossRef]
- Laurent, V.; Nieto, L.; Valet, P.; Muller, C. Adipose tissue and cancer: A high risk tandem. Med. Sci. MS 2014, 30, 398–404. [Google Scholar] [CrossRef]
- Attané, C.; Muller, C. Drilling for Oil: Tumor-Surrounding Adipocytes Fueling Cancer. Trends Cancer 2020, 6, 593–604. [Google Scholar] [CrossRef]
- Galic, S.; Oakhill, J.S.; Steinberg, G.R. Adipose Tissue as an Endocrine Organ. Mol. Cell. Endocrinol. 2010, 316, 129–139. [Google Scholar] [CrossRef]
- Finley, D.S.; Calvert, V.S.; Inokuchi, J.; Lau, A.; Narula, N.; Petricoin, E.F.; Zaldivar, F.; Santos, R.; Tyson, D.R.; Ornstein, D.K. Periprostatic Adipose Tissue as a Modulator of Prostate Cancer Aggressiveness. J. Urol. 2009, 182, 1621–1627. [Google Scholar] [CrossRef] [PubMed]
- Miladinovic, D.; Cusick, T.; Mahon, K.L.; Haynes, A.-M.; Cortie, C.H.; Meyer, B.J.; Stricker, P.D.; Wittert, G.A.; Butler, L.M.; Horvath, L.G.; et al. Assessment of Periprostatic and Subcutaneous Adipose Tissue Lipolysis and Adipocyte Size from Men with Localized Prostate Cancer. Cancers 2020, 12, 1385. [Google Scholar] [CrossRef] [PubMed]
- Reagan, M.R.; Fairfield, H.; Rosen, C.J. Bone Marrow Adipocytes: A Link between Obesity and Bone Cancer. Cancers 2021, 13, 364. [Google Scholar] [CrossRef] [PubMed]
- Attané, C.; Estève, D.; Chaoui, K.; Iacovoni, J.S.; Corre, J.; Moutahir, M.; Valet, P.; Schiltz, O.; Reina, N.; Muller, C. Human Bone Marrow Is Comprised of Adipocytes with Specific Lipid Metabolism. Cell Rep. 2020, 30, 949–958.e6. [Google Scholar] [CrossRef]
- Arina, A.; Idel, C.; Hyjek, E.M.; Alegre, M.-L.; Wang, Y.; Bindokas, V.P.; Weichselbaum, R.R.; Schreiber, H. Tumor-Associated Fibroblasts Predominantly Come from Local and Not Circulating Precursors. Proc. Natl. Acad. Sci. USA 2016, 113, 7551–7556. [Google Scholar] [CrossRef]
- Bonollo, F.; Thalmann, G.N.; Kruithof-de Julio, M.; Karkampouna, S. The Role of Cancer-Associated Fibroblasts in Prostate Cancer Tumorigenesis. Cancers 2020, 12, 1887. [Google Scholar] [CrossRef]
- Ridge, S.M.; Bhattacharyya, D.; Dervan, E.; Naicker, S.D.; Burke, A.J.; Murphy, J.M.; O’leary, K.; Greene, J.; Ryan, A.E.; Sullivan, F.J.; et al. Secreted Factors from Metastatic Prostate Cancer Cells Stimulate Mesenchymal Stem Cell Transition to a Pro-tumourigenic ‘Activated’ State That Enhances Prostate Cancer Cell Migration. Int. J. Cancer 2018, 142, 2056–2067. [Google Scholar] [CrossRef]
- Xie, H.; Li, L.; Zhu, G.; Dang, Q.; Ma, Z.; He, D.; Chang, L.; Song, W.; Chang, H.-C.; Krolewski, J.J.; et al. Infiltrated Pre-Adipocytes Increase Prostate Cancer Metastasis via Modulation of the MiR-301a/Androgen Receptor (AR)/TGF-Β1/Smad/MMP9 Signals. Oncotarget 2015, 6, 12326–12339. [Google Scholar] [CrossRef]
- Ribeiro, R.; Monteiro, C.; Silvestre, R.; Castela, A.; Coutinho, H.; Fraga, A.; Príncipe, P.; Lobato, C.; Costa, C.; Cordeiro-da-Silva, A.; et al. Human Periprostatic White Adipose Tissue Is Rich in Stromal Progenitor Cells and a Potential Source of Prostate Tumor Stroma. Exp. Biol. Med. Maywood NJ 2012, 237, 1155–1162. [Google Scholar] [CrossRef]
- Gazi, E.; Gardner, P.; Lockyer, N.P.; Hart, C.A.; Brown, M.D.; Clarke, N.W. Direct Evidence of Lipid Translocation between Adipocytes and Prostate Cancer Cells with Imaging FTIR Microspectroscopy. J. Lipid Res. 2007, 48, 1846–1856. [Google Scholar] [CrossRef] [Green Version]
- Tousignant, K.D.; Rockstroh, A.; Poad, B.L.J.; Talebi, A.; Young, R.S.E.; Taherian Fard, A.; Gupta, R.; Zang, T.; Wang, C.; Lehman, M.L.; et al. Therapy-Induced Lipid Uptake and Remodeling Underpin Ferroptosis Hypersensitivity in Prostate Cancer. Cancer Metab. 2020, 8, 11. [Google Scholar] [CrossRef] [PubMed]
- Watt, M.J.; Clark, A.K.; Selth, L.A.; Haynes, V.R.; Lister, N.; Rebello, R.; Porter, L.H.; Niranjan, B.; Whitby, S.T.; Lo, J.; et al. Suppressing Fatty Acid Uptake Has Therapeutic Effects in Preclinical Models of Prostate Cancer. Sci. Transl. Med. 2019, 11, eaau5758. [Google Scholar] [CrossRef] [PubMed]
- Guaita-Esteruelas, S.; Gumà, J.; Masana, L.; Borràs, J. The Peritumoural Adipose Tissue Microenvironment and Cancer. The Roles of Fatty Acid Binding Protein 4 and Fatty Acid Binding Protein 5. Mol. Cell. Endocrinol. 2018, 462, 107–118. [Google Scholar] [CrossRef]
- Huang, M.; Narita, S.; Inoue, T.; Koizumi, A.; Saito, M.; Tsuruta, H.; Numakura, K.; Satoh, S.; Nanjo, H.; Sasaki, T.; et al. Fatty Acid Binding Protein 4 Enhances Prostate Cancer Progression by Upregulating Matrix Metalloproteinases and Stromal Cell Cytokine Production. Oncotarget 2017, 8, 111780–111794. [Google Scholar] [CrossRef]
- Herroon, M.K.; Rajagurubandara, E.; Hardaway, A.L.; Powell, K.; Turchick, A.; Feldmann, D.; Podgorski, I. Bone Marrow Adipocytes Promote Tumor Growth in Bone via FABP4-Dependent Mechanisms. Oncotarget 2013, 4, 2108–2123. [Google Scholar] [CrossRef] [PubMed]
- Tan, N.-S.; Shaw, N.S.; Vinckenbosch, N.; Liu, P.; Yasmin, R.; Desvergne, B.; Wahli, W.; Noy, N. Selective Cooperation between Fatty Acid Binding Proteins and Peroxisome Proliferator-Activated Receptors in Regulating Transcription. Mol. Cell. Biol. 2002, 22, 5114–5127. [Google Scholar] [CrossRef]
- Myers, J.S.; von Lersner, A.K.; Sang, Q.-X.A. Proteomic Upregulation of Fatty Acid Synthase and Fatty Acid Binding Protein 5 and Identification of Cancer- and Race-Specific Pathway Associations in Human Prostate Cancer Tissues. J. Cancer 2016, 7, 1452–1464. [Google Scholar] [CrossRef]
- Al-Jameel, W.; Gou, X.; Jin, X.; Zhang, J.; Wei, Q.; Ai, J.; Li, H.; Al-Bayati, A.; Platt-Higgins, A.; Pettitt, A.; et al. Inactivated FABP5 Suppresses Malignant Progression of Prostate Cancer Cells by Inhibiting the Activation of Nuclear Fatty Acid Receptor PPARγ. Genes Cancer 2019, 10, 80–96. [Google Scholar] [CrossRef]
- Lin, L.-C.; Gao, A.C.; Lai, C.-H.; Hsieh, J.-T.; Lin, H. Induction of Neuroendocrine Differentiation in Castration Resistant Prostate Cancer Cells by Adipocyte Differentiation-Related Protein (ADRP) Delivered by Exosomes. Cancer Lett. 2017, 391, 74–82. [Google Scholar] [CrossRef]
- Balaban, S.; Nassar, Z.D.; Zhang, A.Y.; Hosseini-Beheshti, E.; Centenera, M.M.; Schreuder, M.; Lin, H.-M.; Aishah, A.; Varney, B.; Liu-Fu, F.; et al. Extracellular Fatty Acids Are the Major Contributor to Lipid Synthesis in Prostate Cancer. Mol. Cancer Res. MCR 2019, 17, 949–962. [Google Scholar] [CrossRef]
- Yue, S.; Li, J.; Lee, S.-Y.; Lee, H.J.; Shao, T.; Song, B.; Cheng, L.; Masterson, T.A.; Liu, X.; Ratliff, T.L.; et al. Cholesteryl Ester Accumulation Induced by PTEN Loss and PI3K/AKT Activation Underlies Human Prostate Cancer Aggressiveness. Cell Metab. 2014, 19, 393–406. [Google Scholar] [CrossRef] [PubMed]
- Zechner, R.; Madeo, F.; Kratky, D. Cytosolic Lipolysis and Lipophagy: Two Sides of the Same Coin. Nat. Rev. Mol. Cell Biol. 2017, 18, 671–684. [Google Scholar] [CrossRef] [PubMed]
- Fontaine, A.; Bellanger, D.; Guibon, R.; Bruyère, F.; Brisson, L.; Fromont, G. Lipophagy and Prostate Cancer: Association with Disease Aggressiveness and Proximity to Periprostatic Adipose Tissue. J. Pathol. 2021, 255, 166–176. [Google Scholar] [CrossRef]
- Meng, H.; Shen, Y.; Shen, J.; Zhou, F.; Shen, S.; Das, U.N. Effect of N-3 and n-6 Unsaturated Fatty Acids on Prostate Cancer (PC-3) and Prostate Epithelial (RWPE-1) Cells in Vitro. Lipids Health Dis. 2013, 12, 160. [Google Scholar] [CrossRef] [PubMed]
- Tawadros, T.; Brown, M.D.; Hart, C.A.; Clarke, N.W. Ligand-Independent Activation of EphA2 by Arachidonic Acid Induces Metastasis-like Behaviour in Prostate Cancer Cells. Br. J. Cancer 2012, 107, 1737–1744. [Google Scholar] [CrossRef]
- Wang, X.; Sun, B.; Wei, L.; Jian, X.; Shan, K.; He, Q.; Huang, F.; Ge, X.; Gao, X.; Feng, N.; et al. Cholesterol and Saturated Fatty Acids Synergistically Promote the Malignant Progression of Prostate Cancer. Neoplasia 2022, 24, 86–97. [Google Scholar] [CrossRef]
- Figiel, S.; Bery, F.; Chantôme, A.; Fontaine, D.; Pasqualin, C.; Maupoil, V.; Domingo, I.; Guibon, R.; Fromont, G.; Mahéo, K.; et al. A Novel Calcium-Mediated EMT Pathway Controlled by Lipids: An Opportunity for Prostate Cancer Adjuvant Therapy. Cancers 2019, 11, 1814. [Google Scholar] [CrossRef]
- Figiel, S.; Pinault, M.; Domingo, I.; Guimaraes, C.; Guibon, R.; Besson, P.; Tavernier, E.; Blanchet, P.; Multigner, L.; Bruyère, F.; et al. Fatty Acid Profile in Peri-Prostatic Adipose Tissue and Prostate Cancer Aggressiveness in African–Caribbean and Caucasian Patients. Eur. J. Cancer 2018, 91, 107–115. [Google Scholar] [CrossRef]
- Huang, B.; Song, B.-L.; Xu, C. Cholesterol Metabolism in Cancer: Mechanisms and Therapeutic Opportunities. Nat. Metab. 2020, 2, 132–141. [Google Scholar] [CrossRef]
- Kraemer, F.B.; Khor, V.K.; Shen, W.-J.; Azhar, S. Cholesterol Ester Droplets and Steroidogenesis. Mol. Cell. Endocrinol. 2013, 371, 15–19. [Google Scholar] [CrossRef] [Green Version]
- Lethongsavarn, V.; Pinault, M.; Diedhiou, A.; Guimaraes, C.; Guibon, R.; Bruyère, F.; Mathieu, R.; Rioux-Leclercq, N.; Multigner, L.; Brureau, L.; et al. Tissue Cholesterol Metabolism and Prostate Cancer Aggressiveness: Ethno-geographic Variations. Prostate 2021, 81, 1365–1373. [Google Scholar] [CrossRef] [PubMed]
- Micucci, C.; Valli, D.; Matacchione, G.; Catalano, A. Current Perspectives between Metabolic Syndrome and Cancer. Oncotarget 2016, 7, 38959–38972. [Google Scholar] [CrossRef]
- Massillo, C.; Dalton, G.N.; Porretti, J.; Scalise, G.D.; Farré, P.L.; Piccioni, F.; Secchiari, F.; Pascuali, N.; Clyne, C.; Gardner, K.; et al. CTBP1/CYP19A1/Estradiol Axis Together with Adipose Tissue Impacts over Prostate Cancer Growth Associated to Metabolic Syndrome. Int. J. Cancer 2019, 144, 1115–1127. [Google Scholar] [CrossRef] [PubMed]
- Di Zazzo, E.; Galasso, G.; Giovannelli, P.; Di Donato, M.; Di Santi, A.; Cernera, G.; Rossi, V.; Abbondanza, C.; Moncharmont, B.; Sinisi, A.A.; et al. Prostate Cancer Stem Cells: The Role of Androgen and Estrogen Receptors. Oncotarget 2016, 7, 193–208. [Google Scholar] [CrossRef] [PubMed]
- Buschemeyer, W.C.; Freedland, S.J. Obesity and Prostate Cancer: Epidemiology and Clinical Implications. Eur. Urol. 2007, 52, 331–343. [Google Scholar] [CrossRef]
- Baillargeon, J.; Rose, D.P. Obesity, Adipokines, and Prostate Cancer (Review). Int. J. Oncol. 2006, 28, 737–745. [Google Scholar] [CrossRef]
- Basaria, S.; Muller, D.C.; Carducci, M.A.; Egan, J.; Dobs, A.S. Hyperglycemia and Insulin Resistance in Men with Prostate Carcinoma Who Receive Androgen-Deprivation Therapy. Cancer 2006, 106, 581–588. [Google Scholar] [CrossRef]
- Somasundar, P.; McFadden, D.W.; Hileman, S.M.; Vona-Davis, L. Leptin Is a Growth Factor in Cancer. J. Surg. Res. 2004, 116, 337–349. [Google Scholar] [CrossRef]
- Gorrab, A.; Pagano, A.; Ayed, K.; Chebil, M.; Derouiche, A.; Kovacic, H.; Gati, A. Leptin Promotes Prostate Cancer Proliferation and Migration by Stimulating STAT3 Pathway. Nutr. Cancer 2021, 73, 1217–1227. [Google Scholar] [CrossRef]
- Philp, L.K.; Rockstroh, A.; Sadowski, M.C.; Taherian Fard, A.; Lehman, M.; Tevz, G.; Libério, M.S.; Bidgood, C.L.; Gunter, J.H.; McPherson, S.; et al. Leptin Antagonism Inhibits Prostate Cancer Xenograft Growth and Progression. Endocr. Relat. Cancer 2021, 28, 353–375. [Google Scholar] [CrossRef]
- Gao, Q.; Zheng, J.; Yao, X.; Peng, B. Adiponectin Inhibits VEGF-A in Prostate Cancer Cells. Tumour Biol. J. Int. Soc. Oncodev. Biol. Med. 2015, 36, 4287–4292. [Google Scholar] [CrossRef] [PubMed]
- Philp, L.K.; Rockstroh, A.; Lehman, M.; Sadowski, M.C.; Bartonicek, N.; Wade, J.D.; Otvos, L.; Nelson, C.C. Adiponectin Receptor Activation Inhibits Prostate Cancer Xenograft Growth. Endocr. Relat. Cancer 2020, 27, 711–729. [Google Scholar] [CrossRef] [PubMed]
- Wakui, S.; Furusato, M.; Itoh, T.; Sasaki, H.; Akiyama, A.; Kinoshita, I.; Asano, K.; Tokuda, T.; Aizawa, S.; Ushigome, S. Tumour Angiogenesis in Prostatic Carcinoma with and without Bone Marrow Metastasis: A Morphometric Study. J. Pathol. 1992, 168, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Brawer, M.K.; Deering, R.E.; Brown, M.; Preston, S.D.; Bigler, S.A. Predictors of Pathologic Stage in Prostatic Carcinoma. The Role of Neovascularity. Cancer 1994, 73, 678–687. [Google Scholar] [CrossRef]
- Stefanou, D.; Batistatou, A.; Kamina, S.; Arkoumani, E.; Papachristou, D.J.; Agnantis, N.J. Expression of Vascular Endothelial Growth Factor (VEGF) and Association with Microvessel Density in Benign Prostatic Hyperplasia and Prostate Cancer. In Vivo 2004, 18, 155–160. [Google Scholar]
- Kuniyasu, H.; Troncoso, P.; Johnston, D.; Bucana, C.D.; Tahara, E.; Fidler, I.J.; Pettaway, C.A. Relative Expression of Type IV Collagenase, E-Cadherin, and Vascular Endothelial Growth Factor/Vascular Permeability Factor in Prostatectomy Specimens Distinguishes Organ-Confined from Pathologically Advanced Prostate Cancers. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2000, 6, 2295–2308. [Google Scholar]
- Strohmeyer, D.; Strauss, F.; Rössing, C.; Roberts, C.; Kaufmann, O.; Bartsch, G.; Effert, P. Expression of BFGF, VEGF and c-Met and Their Correlation with Microvessel Density and Progression in Prostate Carcinoma. Anticancer Res. 2004, 24, 1797–1804. [Google Scholar]
- Trojan, L.; Thomas, D.; Knoll, T.; Grobholz, R.; Alken, P.; Michel, M.S. Expression of Pro-Angiogenic Growth Factors VEGF, EGF and BFGF and Their Topographical Relation to Neovascularisation in Prostate Cancer. Urol. Res. 2004, 32, 97–103. [Google Scholar] [CrossRef]
- Freeman, M.R.; Paul, S.; Kaefer, M.; Ishikawa, M.; Adam, R.M.; Renshaw, A.A.; Elenius, K.; Klagsbrun, M. Heparin-Binding EGF-like Growth Factor in the Human Prostate: Synthesis Predominantly by Interstitial and Vascular Smooth Muscle Cells and Action as a Carcinoma Cell Mitogen. J. Cell. Biochem. 1998, 68, 328–338. [Google Scholar] [CrossRef]
- Laurent, V.; Guérard, A.; Mazerolles, C.; Le Gonidec, S.; Toulet, A.; Nieto, L.; Zaidi, F.; Majed, B.; Garandeau, D.; Socrier, Y.; et al. Periprostatic Adipocytes Act as a Driving Force for Prostate Cancer Progression in Obesity. Nat. Commun. 2016, 7, 10230. [Google Scholar] [CrossRef]
- Guérard, A.; Laurent, V.; Fromont, G.; Estève, D.; Gilhodes, J.; Bonnelye, E.; Le Gonidec, S.; Valet, P.; Malavaud, B.; Reina, N.; et al. The Chemokine Receptor CCR3 Is Potentially Involved in the Homing of Prostate Cancer Cells to Bone: Implication of Bone-Marrow Adipocytes. Int. J. Mol. Sci. 2021, 22, 1994. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Tseng, C.; Zhang, Y.; Sirin, O.; Corn, P.G.; Li-Ning-Tapia, E.M.; Troncoso, P.; Davis, J.; Pettaway, C.; Ward, J.; et al. CXCL1 Mediates Obesity-Associated Adipose Stromal Cell Trafficking and Function in the Tumour Microenvironment. Nat. Commun. 2016, 7, 11674. [Google Scholar] [CrossRef] [PubMed]
- Su, F.; Daquinag, A.C.; Ahn, S.; Saha, A.; Dai, Y.; Zhao, Z.; DiGiovanni, J.; Kolonin, M.G. Progression of Prostate Carcinoma Is Promoted by Adipose Stromal Cell-Secreted CXCL12 Signaling in Prostate Epithelium. NPJ Precis. Oncol. 2021, 5, 26. [Google Scholar] [CrossRef] [PubMed]
- Sacca, P.A.; Creydt, V.P.; Choi, H.; Mazza, O.N.; Fletcher, S.J.; Vallone, V.B.F.; Scorticati, C.; Chasseing, N.A.; Calvo, J.C. Human Periprostatic Adipose Tissue: Its Influence on Prostate Cancer Cells. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2012, 30, 113–122. [Google Scholar] [CrossRef]
- Habib, C.N.; Al-Abd, A.M.; Tolba, M.F.; Khalifa, A.E.; Khedr, A.; Mosli, H.A.; Abdel-Naim, A.B. Leptin Influences Estrogen Metabolism and Accelerates Prostate Cell Proliferation. Life Sci. 2015, 121, 10–15. [Google Scholar] [CrossRef]
- Bokobza, E.; Hinault, C.; Tiroille, V.; Clavel, S.; Bost, F.; Chevalier, N. The Adipose Tissue at the Crosstalk between EDCs and Cancer Development. Front. Endocrinol. 2021, 12, 691658. [Google Scholar] [CrossRef]
- Quintana, P.J.E.; Delfino, R.J.; Korrick, S.; Ziogas, A.; Kutz, F.W.; Jones, E.L.; Laden, F.; Garshick, E. Adipose Tissue Levels of Organochlorine Pesticides and Polychlorinated Biphenyls and Risk of Non-Hodgkin’s Lymphoma. Environ. Health Perspect. 2004, 112, 854–861. [Google Scholar] [CrossRef]
- Charazac, A.; Hinault, C.; Dolfi, B.; Hautier, S.; Decondé Le Butor, C.; Bost, F.; Chevalier, N. Low Doses of PFOA Promote Prostate and Breast Cancer Cells Growth through Different Pathways. Int. J. Mol. Sci. 2022, 23, 7900. [Google Scholar] [CrossRef]
- Lemaire, G.; Mnif, W.; Mauvais, P.; Balaguer, P.; Rahmani, R. Activation of Alpha- and Beta-Estrogen Receptors by Persistent Pesticides in Reporter Cell Lines. Life Sci. 2006, 79, 1160–1169. [Google Scholar] [CrossRef]
- Okubo, T.; Yokoyama, Y.; Kano, K.; Soya, Y.; Kano, I. Estimation of Estrogenic and Antiestrogenic Activities of Selected Pesticides by MCF-7 Cell Proliferation Assay. Arch. Environ. Contam. Toxicol. 2004, 46, 445–453. [Google Scholar] [CrossRef]
- Wong, L.I.L.; Labrecque, M.P.; Ibuki, N.; Cox, M.E.; Elliott, J.E.; Beischlag, T.V. P,p’-Dichlorodiphenyltrichloroethane (p,p’-DDT) and p,p’-Dichlorodiphenyldichloroethylene (p,p’-DDE) Repress Prostate Specific Antigen Levels in Human Prostate Cancer Cell Lines. Chem. Biol. Interact. 2015, 230, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Multigner, L.; Ndong, J.R.; Giusti, A.; Romana, M.; Delacroix-Maillard, H.; Cordier, S.; Jégou, B.; Thome, J.P.; Blanchet, P. Chlordecone Exposure and Risk of Prostate Cancer. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2010, 28, 3457–3462. [Google Scholar] [CrossRef] [PubMed]
- Emeville, E.; Giusti, A.; Coumoul, X.; Thomé, J.-P.; Blanchet, P.; Multigner, L. Associations of Plasma Concentrations of Dichlorodiphenyldichloroethylene and Polychlorinated Biphenyls with Prostate Cancer: A Case-Control Study in Guadeloupe (French West Indies). Environ. Health Perspect. 2015, 123, 317–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.; Scheri, R.C.; Zhang, Y.; Curtis, L.R. Chlordecone, a Mixed Pregnane X Receptor (PXR) and Estrogen Receptor Alpha (ERalpha) Agonist, Alters Cholesterol Homeostasis and Lipoprotein Metabolism in C57BL/6 Mice. Toxicol. Appl. Pharmacol. 2008, 233, 193–202. [Google Scholar] [CrossRef]
- Moreira, Â.; Pereira, S.S.; Costa, M.; Morais, T.; Pinto, A.; Fernandes, R.; Monteiro, M.P. Adipocyte Secreted Factors Enhance Aggressiveness of Prostate Carcinoma Cells. PLoS ONE 2015, 10, e0123217. [Google Scholar] [CrossRef]
- Ribeiro, R.; Monteiro, C.; Cunha, V.; Oliveira, M.J.; Freitas, M.; Fraga, A.; Príncipe, P.; Lobato, C.; Lobo, F.; Morais, A.; et al. Human Periprostatic Adipose Tissue Promotes Prostate Cancer Aggressiveness in Vitro. J. Exp. Clin. Cancer Res. 2012, 31, 32. [Google Scholar] [CrossRef]
- Xu, C.-J.; Dong, L.-L.; Kang, X.-L.; Li, Z.-M.; Zhang, H.-Y. Leptin Promotes Proliferation and Inhibits Apoptosis of Prostate Cancer Cells by Regulating ERK1/2 Signaling Pathway. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 8341–8348. [Google Scholar] [CrossRef]
- Noda, T.; Kikugawa, T.; Tanji, N.; Miura, N.; Asai, S.; Higashiyama, S.; Yokoyama, M. Long-term Exposure to Leptin Enhances the Growth of Prostate Cancer Cells. Int. J. Oncol. 2015, 46, 1535–1542. [Google Scholar] [CrossRef]
- Chen, M.; Zhang, J.; Sampieri, K.; Clohessy, J.G.; Mendez, L.; Gonzalez-Billalabeitia, E.; Liu, X.-S.; Lee, Y.-R.; Fung, J.; Katon, J.M.; et al. An Aberrant SREBP-Dependent Lipogenic Program Promotes Metastatic Prostate Cancer. Nat. Genet. 2018, 50, 206–218. [Google Scholar] [CrossRef]
- Labbé, D.P.; Zadra, G.; Yang, M.; Reyes, J.M.; Lin, C.Y.; Cacciatore, S.; Ebot, E.M.; Creech, A.L.; Giunchi, F.; Fiorentino, M.; et al. High-Fat Diet Fuels Prostate Cancer Progression by Rewiring the Metabolome and Amplifying the MYC Program. Nat. Commun. 2019, 10, 4358. [Google Scholar] [CrossRef]
- Dahran, N.; Szewczyk-Bieda, M.; Wei, C.; Vinnicombe, S.; Nabi, G. Normalized Periprostatic Fat MRI Measurements Can Predict Prostate Cancer Aggressiveness in Men Undergoing Radical Prostatectomy for Clinically Localised Disease. Sci. Rep. 2017, 7, 4630. [Google Scholar] [CrossRef] [PubMed]
- Woo, S.; Cho, J.Y.; Kim, S.Y.; Kim, S.H. Periprostatic Fat Thickness on MRI: Correlation with Gleason Score in Prostate Cancer. AJR Am. J. Roentgenol. 2015, 204, W43–W47. [Google Scholar] [CrossRef] [PubMed]
- Roumiguié, M.; Estève, D.; Manceau, C.; Toulet, A.; Gilleron, J.; Belles, C.; Jia, Y.; Houël, C.; Pericart, S.; LeGonidec, S.; et al. Periprostatic Adipose Tissue Displays a Chronic Hypoxic State That Limits Its Expandability. Am. J. Pathol. 2022, 192, 926–942. [Google Scholar] [CrossRef]
- Gucalp, A.; Iyengar, N.M.; Zhou, X.K.; Giri, D.D.; Falcone, D.J.; Wang, H.; Williams, S.; Krasne, M.D.; Yaghnam, I.; Kunzel, B.; et al. Periprostatic Adipose Inflammation Is Associated with High-Grade Prostate Cancer. Prostate Cancer Prostatic Dis. 2017, 20, 418–423. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, A.; Satoh, Y.; Tokuda, Y.; Fujiyama, C.; Udo, K.; Uozumi, J. Effects of Adipocytes on the Proliferation and Differentiation of Prostate Cancer Cells in a 3-D Culture Model. Int. J. Urol. Off. J. Jpn. Urol. Assoc. 2010, 17, 369–376. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, R.J.T.; Monteiro, C.P.D.; Cunha, V.F.P.M.; Azevedo, A.S.M.; Oliveira, M.J.; Monteiro, R.; Fraga, A.M.; Príncipe, P.; Lobato, C.; Lobo, F.; et al. Tumor Cell-Educated Periprostatic Adipose Tissue Acquires an Aggressive Cancer-Promoting Secretory Profile. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2012, 29, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Aaseth, J.; Javorac, D.; Djordjevic, A.B.; Bulat, Z.; Skalny, A.V.; Zaitseva, I.P.; Aschner, M.; Tinkov, A.A. The Role of Persistent Organic Pollutants in Obesity: A Review of Laboratory and Epidemiological Studies. Toxics 2022, 10, 65. [Google Scholar] [CrossRef]
- Adesunloye, B.A. Mechanistic Insights into the Link between Obesity and Prostate Cancer. Int. J. Mol. Sci. 2021, 22, 3935. [Google Scholar] [CrossRef]
- Evans, S.; Metcalfe, C.; Ibrahim, F.; Persad, R.; Ben-Shlomo, Y. Investigating Black-White Differences in Prostate Cancer Prognosis: A Systematic Review and Meta-Analysis. Int. J. Cancer 2008, 123, 430–435. [Google Scholar] [CrossRef]
- Rebbeck, T.R. Prostate Cancer Disparities by Race and Ethnicity: From Nucleotide to Neighborhood. Cold Spring Harb. Perspect. Med. 2018, 8, a030387. [Google Scholar] [CrossRef]
- Khani, F.; Mosquera, J.M.; Park, K.; Blattner, M.; O’Reilly, C.; MacDonald, T.Y.; Chen, Z.; Srivastava, A.; Tewari, A.K.; Barbieri, C.E.; et al. Evidence for Molecular Differences in Prostate Cancer between African American and Caucasian Men. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2014, 20, 4925–4934. [Google Scholar] [CrossRef]
- Meller, S.; Meyer, H.-A.; Bethan, B.; Dietrich, D.; Maldonado, S.G.; Lein, M.; Montani, M.; Reszka, R.; Schatz, P.; Peter, E.; et al. Integration of Tissue Metabolomics, Transcriptomics and Immunohistochemistry Reveals ERG- and Gleason Score-Specific Metabolomic Alterations in Prostate Cancer. Oncotarget 2016, 7, 1421–1438. [Google Scholar] [CrossRef] [PubMed]
- Mathias, R.A.; Sergeant, S.; Ruczinski, I.; Torgerson, D.G.; Hugenschmidt, C.E.; Kubala, M.; Vaidya, D.; Suktitipat, B.; Ziegler, J.T.; Ivester, P.; et al. The Impact of FADS Genetic Variants on Ω6 Polyunsaturated Fatty Acid Metabolism in African Americans. BMC Genet. 2011, 12, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castetbon, K.; Ramalli, L.; Vaidie, A.; Yacou, C.; Merle, S.; Ducros, V.; Deschamps, V.; Blateau, A. Consommations alimentaires et biomarqueurs nutritionnels chez les adultes de 16 ans et plus en Guadeloupe et Martinique. Enquête Kannari 2013–2014. Bull. Épidémiologique Hebd. 2016, 4, 52–62. [Google Scholar]
- Kohlmeier, L.; Kohlmeier, M. Adipose Tissue as a Medium for Epidemiologic Exposure Assessment. Environ. Health Perspect. 1995, 103 (Suppl. S3), 99–106. [Google Scholar] [CrossRef] [PubMed]
- Baldi, I.; Botton, J.; Chevrier, C.; Coumoul, X.; Elbaz, A.; Goujon, S.; Jouzel, J.-N.; Monnereau, A.; Multigner, L.; Salles, B.; et al. Inserm. Pesticides et effets sur la santé: Nouvelles données (2021); Collection Expertise Collective; INSERM: Paris, France; EDP Sciences: Les Ulis, France, 2021. [Google Scholar]
- Friedenreich, C.M.; Wang, Q.; Neilson, H.K.; Kopciuk, K.A.; McGregor, S.E.; Courneya, K.S. Physical Activity and Survival After Prostate Cancer. Eur. Urol. 2016, 70, 576–585. [Google Scholar] [CrossRef]
- Wang, Y.; Jacobs, E.J.; Gapstur, S.M.; Maliniak, M.L.; Gansler, T.; McCullough, M.L.; Stevens, V.L.; Patel, A.V. Recreational Physical Activity in Relation to Prostate Cancer-Specific Mortality Among Men with Nonmetastatic Prostate Cancer. Eur. Urol. 2017, 72, 931–939. [Google Scholar] [CrossRef]
- Capece, M.; Creta, M.; Calogero, A.; La Rocca, R.; Napolitano, L.; Barone, B.; Sica, A.; Fusco, F.; Santangelo, M.; Dodaro, C.; et al. Does Physical Activity Regulate Prostate Carcinogenesis and Prostate Cancer Outcomes? A Narrative Review. Int. J. Environ. Res. Public. Health 2020, 17, 1441. [Google Scholar] [CrossRef] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cancel, M.; Pouillot, W.; Mahéo, K.; Fontaine, A.; Crottès, D.; Fromont, G. Interplay between Prostate Cancer and Adipose Microenvironment: A Complex and Flexible Scenario. Int. J. Mol. Sci. 2022, 23, 10762. https://doi.org/10.3390/ijms231810762
Cancel M, Pouillot W, Mahéo K, Fontaine A, Crottès D, Fromont G. Interplay between Prostate Cancer and Adipose Microenvironment: A Complex and Flexible Scenario. International Journal of Molecular Sciences. 2022; 23(18):10762. https://doi.org/10.3390/ijms231810762
Chicago/Turabian StyleCancel, Mathilde, William Pouillot, Karine Mahéo, Alix Fontaine, David Crottès, and Gaëlle Fromont. 2022. "Interplay between Prostate Cancer and Adipose Microenvironment: A Complex and Flexible Scenario" International Journal of Molecular Sciences 23, no. 18: 10762. https://doi.org/10.3390/ijms231810762
APA StyleCancel, M., Pouillot, W., Mahéo, K., Fontaine, A., Crottès, D., & Fromont, G. (2022). Interplay between Prostate Cancer and Adipose Microenvironment: A Complex and Flexible Scenario. International Journal of Molecular Sciences, 23(18), 10762. https://doi.org/10.3390/ijms231810762