2.1. Analysis of RNA-seq Experiment
RNA-seq libraries were sequenced and 47.9 to 48.3 million reads per lane were generated. A total of 81.8–82.3% of the total Illumina reads aligned to the
Glycine max Wm82.a2.v1 [
47] reference genome (
Table 1).
The expression of 53.2 to 53.6 thousand genes was investigated in this RNA sequencing analysis. An FDR ≤ 0.001 and the absolute value of log
2 ratio ≥1 were used as the threshold to judge the significance of gene expression difference. According to the statistical criteria, we identified genes from the expression profile data with expression levels that were statistically altered by flooding stress. Comparing genotypes, TECIRGA 6070 presented 1494 and 731 genes up- and down-regulated, respectively, in the control condition. Regarding the flooding treatment, 752 and 871 genes were up- and down-regulated, respectively, in the tolerant genotype (
Table 2).
RNA-seq data were validated using qPCR. ERF15, Isoflavone, LOX1, LDOX and two ADH genes were up-regulated in the tolerant cultivar (TECIRGA 6070) 24 h after flooding stress (
Figure 1). In Arabidopsis and soybean, lipoxygenases are involved in increasing stress resistance and boosting defense reactions [
48,
49]. Under flooding stress, most soybean lipoxygenases are decreased in the roots [
50]. Komatsu et al. (2010) [
51] reported that two lipoxygenases were decreased in soybean in response to flooding stress and suggested that these enzymes affected cell wall metabolism due to the suppression of lignification. Taken together, the data indicate that lipoxygenase may regulate hormone pathways for stress tolerance and is involved in defense reactions in soybean under flooding stress. In our work, a lipoxygenase was found up-regulated in the tolerant cultivar. This gene could be involved in the tolerance mechanism.
Anthocyanins, as one of the most important water-soluble pigments in plants, have a wide range of biological functions, such as plant abiotic stress adaptation, defense against pathogen invasion, antioxidation and other health-related functions. Anthocyanins are flavonoids, and their biosynthesis and accumulation are parts of the flavonoid biosynthetic pathway. Leucoanthocyanidin dioxygenase (LDOX) is an enzyme present in this pathway and is responsible to form anthocyanidins, later converted into anthocyanins. Ding et al. (2022) [
52] showed that the increased anthocyanin content could be related to strong waterlogging resistance in rapeseed (
Brassica napus).
Transcription factors (TFs) representing basic helix–loop–helix (bHLH), ethylene response factors (ERFs), myeloblastosis (MYB), no apical meristem (NAC), and WRKY amino acid motif (WRKY) types are major families known to be involved in the mechanisms of stress tolerance.
The AP2/ERF transcription factor superfamily has been identified in many species and is divided into different families based on the number of AP2 domains, including the AP2, ERF, DREB, and RAV families. Ethylene accumulates rapidly under flooding conditions. The production of ethylene increased in waterlogged soybeans, with a significantly greater increase in tolerant lines than in sensitive lines [
53]. The locus Glyma.05G063600 encodes an ethylene-responsive transcription factor 15-related, now named GmERF37, and belongs to Group I [
54]. Chen et al. (2016) [
55] identified 38 soybean ERFs up-regulated in leaf tissue under flooding stress. The authors observed that GmERF37 was up-regulated under flooding and drought stress. This suggests the role of ethylene in both drought and flood-responsive pathways.
Regarding isoflavones, Coutinho et al. (2018) [
56] observed that the amount of isoflavones in soybean genotypes decreased under flooding conditions, but highlighted that the mechanism of the down-regulation of isoflavones in soybeans under flooding stress remains unknown. On the other hand, it is suggested that soybean isoflavone levels are increased when plants are grown in areas subject to intermittent flooding [
57]. In the present study, the increase was observed in the tolerant cultivar after 24 h of flooding. This could be a mechanism for plant tolerance to stress. Regarding alcohol dehydrogenase (ADH), the gene was also induced after 24 h of stress.
Gene Ontology (GO) analyses were performed and DEGs were linked with their pathway (putative biological roles). Among the DEGs found in the tolerant genotypes at control and flooding conditions, 1712 presented pathway annotation. The most representative pathway was related to metabolic pathways (300 genes—17.5%), followed by ribosomes (191 genes—11.2%), the biosynthesis of secondary metabolites (183 genes—10.69%), plant–pathogen interactions (144 genes—8.41%), and plant hormone signal transduction (125 genes—7.3%) (
Supplementary Figure S1).
Ribosomal proteins are essential for protein synthesis, being involved in developmental processes, and act as environmental sensors [
58]. They are commonly decreased in plants in response to abiotic stresses such as flooding [
59], ultraviolet-B [
60], and cold [
61], retarding growth and impairing productivity [
62]. In soybean, proteins related to protein synthesis were also identified in the roots of 2-day-old plants exposed to flooding under light or dark conditions. The 60S acidic ribosomal protein (Glyma12g30800.3 and Glyma03g35080.1) was down-regulated in both conditions (light and dark). However, the ribosomal protein S5/elongation factor G/III/V (Glyma08g18110.1 and Glyma15g40860.1) was up-regulated under light conditions [
48]. Another study performed with soybean analyzed the rough ER fraction isolated from the root tips of 2-day-old plants submitted to flooding for two days and verified that 65% of ribosomal proteins identified were decreased in response to stress [
63]. In the present work, many ribosomal proteins (60S ribosomal protein L3-like, 40S ribosomal protein SA, ribosomal protein S6, 40S ribosomal protein S11, 40S ribosomal protein S2, ribosomal protein S27, and others listed in
Supplementary Table S3) were up-regulated in the tolerant cultivar (TECIRGA 6070) under flooding conditions compared with control conditions. Protein synthesis has previously been shown to be impaired in soybean roots under flooding, and it might be due to changes in ribosomal protein abundance [
18,
63]. Sharmin et al. [
64] performed a protein–protein interaction analysis and identified three hub genes that could be associated with enhanced soybean seed-flooding tolerance: Glyma.01G207700 (ribosomal protein L23/L15e family protein), Glyma.05G016800 (ribosomal protein L23/L15e family protein) and Glyma.08G159800 (40 s ribosomal protein SA B).
Stress signaling events comprise the production of secondary messengers, variation in the intracellular calcium concentration, and the activation of kinase cascades [
65]. The endoplasmic reticulum is the organelle involved in calcium homeostasis and it is crucial for protein quality [
66]. The increase in intracellular Ca
2+ levels in response to stress conditions is perceived by calcium-binding proteins [
67]. These proteins can bind to the promoter region of genes involved in the stress response or interact with DNA-binding proteins that regulate these genes, occasioning in gene activation or suppression [
65]. Calcium encrypts the
stimuli in calcium signatures, which among other sensors are also detected by calmodulin (CaM) protein [
68]. The cytosolic calcium content has been found to be increased in soybean after flooding and drought stresses [
63]. Here, two calmodulin-binding proteins (Glyma07g30990.1 and Glyma08g12100.1), a calmodulin-binding receptor (Glyma17g11810.2) and a calcium-binding protein (Glyma09g40740.1) were detected as up-regulated under flooding conditions in the tolerant genotype (
Supplementary Table S3). Similarly, Sharmin et al. [
64] detected 24 Ca
2+ signaling-related up-regulated DEGs in a flood-tolerant wild soybean genotype relative to the sensitive genotype. Comparisons in soybean gene expression after 6 and 12 h flooding revealed that genes involved in the calcium signaling were significantly up-regulated after 12 h [
69]. A previous study based on protein data reported that calmodulin-binding proteins were decreased in soybean root tips under flooding, whereas calcium-transporting ATPases were increased in response to this stress [
63]. The contradictory response could be accounted to differences in genotypes, developmental stages (2–4-day-old vs. V6-growth-stage plants), analyzed tissue (cotyledon vs. leaves) and tolerance to stress. On the other hand, in the present work, several members of calnexin and calreticulin were also down-regulated in the tolerant genotype. These proteins are chaperones that fold newly synthesized polypeptides and modulate the calcium transport in the endoplasmic reticulum. In the same way, calreticulin and calnexin proteins were found to be differentially abundant in the root tips of soybean submitted to water stress (flooding and drought) [
63]. Nanjo et al. [
69] also observed altered levels of calnexin in soybean under flooding stress. Taken together, these results demonstrate that the protein folding process might be affected by water stress.
To enable the coordination of genes and their regulators involved in stress remediation, there is a crosstalk among different plant hormones [
70]. Nine groups of plant hormones were described to participate in the defense response and signaling pathways for an efficient reaction to environmental stresses [
65]. Abscisic acid (ABA), ehtylene, jasmonic acid (JA) and salicylic acid (SA) are known to perform major roles in intermediating plant defense response against biotic and abiotic stresses [
71]. The expression of some regulatory genes can significantly affect stress tolerance due to their role in enhancing stress signals to regulate downstream responsive genes [
72]. In this scenario, the transcription factors responsive to stress are important tools to manipulate the plant stress response [
73]. Here, regulator molecules, such as transcription factors, phosphatases and kinases, were also differentially expressed when comparing control and flooding conditions. Among the stress-related transcription factors, several members of the NAC, APETALA2/ethylene-responsive factors (AP2/ERFs), bZIP, DOF, WRKY, GRAS and MYB proteins were detected. Members from the NAC, MYB and MYC families are known to function in an ABA-dependent manner. ERFs are the main downstream regulatory factors of ET signaling pathways in response to stress. DELLA proteins (members of the GRAS family of TF) function as integrators of the gibberellic acid (GA) and ABA signaling pathways [
65,
74].
Ten out of eleven WRKY genes were identified and up-regulated in the tolerant cultivar under flooding conditions (GmWRKY8, GmWRKY20, GmWRKY27, GmWRKY36, GmWRKY42, GmWRKY46, GmWRKY52, GmWRKY126 and GmWRKY149). The only WRKY gene down-regulated was GmWRKY144 (
Supplementary Table S3). Similarly, previous studies have reported that the expression of WRKY-encoding genes is quickly and strongly induced in response to flooding in
A. thaliana, and this is correlated with the induction of innate immunity marker genes [
75,
76,
77]. Likewise, maize WRKY6 was induced in two sensitive genotypes more than two-fold at two time points after submergence (24 and 72 h) relative to the control samples. On the other hand, the expression of WRKY6 was down-regulated at both time points relative to controls in the two tolerant genotypes. Two others putative maize WRKY genes (similar to AtWRKY33) were down-regulated after submergence when compared to the sensitive genotypes [
78]. Sharmin et al. [
64] analyzed the transcriptome profile in roots of two contrasting wild soybean genotypes and verified that some transcription factors were uniquely expressed in the tolerant genotype. Among them are RADIALIS, Trihelix, and UNE, whereas E2F, EGL, GLK1, ORG, PIF, SWI/SNF and TGA were uniquely expressed in the sensitive genotype. The authors also suggest that three soybean GmERFVIIs (GmERFVII1-Glyma.02G016100; GmERFVII2-Glyma.09G041500 and GmERFVII3-Glyma.15G152000)) could play a key role in regulating seed-flooding tolerance in soybean, highlighting the importance of AP2/ERF transcription factors.
Nodulins are membrane channel proteins that facilitate the diffusion of water and small uncharged solutes which are involved in the symbiotic processes between legumes and rhizobia. Here, four nodulin-encoding genes (Glyma20g30580; Glyma04g34550; Glyma02g15380 and Glyma17g08110) were up-regulated under flooding stress (
Supplementary Table S3). Similar results were obtained for
A. thaliana. Under anaerobic conditions (such as, e.g., flooded soil), AtNIP2;1 was highly expressed in the root tips [
79].
Concerning the contrasting genotypes present at stress conditions, 1032 DEGs have shown pathway annotation. Of those, 24.8% belong to the metabolic pathways (256 genes), 18.3% to the plant–pathogen interaction pathway (189 genes), 15.3% to the biosynthesis of secondary metabolites (158 genes), and 9.7% to plant hormone signal transduction (100 genes) (
Supplementary Figure S2). The organic substance metabolic process and primary metabolic process were the most abundant categories in the GO process, comprising 47.0 and 44.3% of the sequences, respectively. Those compounds from primary metabolic processes are formed as a part of the normal anabolic and catabolic processes and are necessary to generate biomass precursors.
SNF-1, a SnRK1 (sucrose non-fermenting-1-related protein kinase 1) gene, was up-regulated in the tolerant genotype TECIRGA 6070 under flooding conditions (
Supplementary Table S4). The SnRK1-A pathway induces MYBS1, which activates the RAmy3D starvation-inducible α-amylase gene [
80]. SnRK1 is a central integrator of energy-related signals to coordinate starvation responses in plants, and it is inhibited by T6P (trehalose-6-P-phosphate). In rice, the locus OsTPP7, which encodes a T6P, has been identified as responsible for anaerobic germination tolerance, which enables uniform germination and seedling establishment under submergence [
81]. Sucrose in high levels will result in higher levels of T6P and, therefore, the repression of SnrK1 and the down-regulation of α-amylases [
2].
Considering the up-regulated genes in the tolerant genotype under control conditions, the highest differential expression was detected for ferritin (Glyma07g19060.1 and Glyma01g31300.1), Snakin-1 (Glyma06g04740.1), lectin precursor (Glyma02g18090.1), P21 protein (Glyma05g38130.1) and ribosomal proteins (
Supplementary Table S2).
2.2. Physiological Variables
Chlorophyll fluorescence assessments were performed three times: immediately before the onset of flooding (time 0), during the flooding period (24 and 48 h of flooding) and after tank drainage (24 and 48 h of recovery). The PSII quantum yield evaluated in the period before flooding did not show any difference between the genotypes and this behavior remained constant in the plants maintained in the control condition (
Supplementary Figure S3). Among the plants subjected to stress with 24 h of flooding, the sensitive genotype (FUNDACEP 62) showed significantly lower PSII quantum yield compared to the tolerant one (TECIRGA 6070). After 48 h of flooding, there was no difference between the genotypes for this parameter, probably due to the stress intensity. In the evaluation carried out 24 h after drainage, FUNDACEP 62 again presented lower quantum yield compared to TECIRGA 6070. In the evaluation performed 48 h after drainage, the genotypes did not differ (
Supplementary Figure S3).
The O
2 consumption by roots, fauna and soil microorganisms can result in O
2 soil depletion within 24 h after the beginning of flooding [
82]. As a result, photosynthesis is inhibited. Chlorophyll fluorescence was as an efficient parameter to characterize and differentiate the magnitude of stress caused by excess water between the genotypes at two times: 24 h of flooding and 24 h after drainage. The sensitive genotype, FUNDACEP 62, showed a sharper drop in quantum yield when stressed and a slower recovery after tank drainage (
Supplementary Figure S3).
The occurrence of unfavorable environmental conditions, such as flooding, can induce the excessive production of reactive oxygen species (ROS). In this study, the content of hydrogen peroxide (H
2O
2) and the activity of the ROS detoxifying enzyme ascorbate peroxidase (APX) were evaluated (
Supplementary Figure S4). It was observed that in plants submitted to 24h of flooding, the H
2O
2 content increased in relation to the control in both genotypes. However, this increase was more pronounced for FUNDACEP 62 (sensitive) than for TECIRGA 6070 (tolerant). In the second evaluation carried out after 48h of flooding, despite decreasing the levels of H
2O
2 relative to control, FUNDACEP 62 still showed higher levels of hydrogen peroxide when compared to the control. For TECIRGA 6070, on the other hand, hydrogen peroxide content in plants submitted to 48 h of flooding did not differ from the control condition. In the evaluation carried out 24 h after the tanks were drained, the H
2O
2 content of the two cultivars did not differ from their respective controls (
Supplementary Figure S4A). ROS are often associated with oxidative stress. However, studies have shown that ROS play an important role in stress signaling and perception in plants [
83,
84,
85]. The results of the present study in soybean agree with the results found in Arabidopsis [
27], indicating that in the tolerant genotype TECIRGA 6070, the production of H
2O
2 may act on the perception and signaling of flooding stress.
Regarding APX activity, FUNDACEP 62 showed an increase as a function of flooding for 24 h, whereas in the second evaluation (48 h of flooding), it did not differ between plants under stress and control conditions. Moreover, the genotype TECIRGA 6070 showed lower APX activity in response to flooding for 24 and 48 h compared to its control (
Supplementary Figure S4B). In a study with two other Brazilian soybean genotypes (Embrapa 45 and BR4, considered tolerant and sensitive to flooding, respectively), the inhibition of APX enzyme activity was observed for Embrapa 45 (tolerant) in response to hypoxia [
86]. These results indicate that in hypoxia-tolerant soybean cultivars, the activity of the APX enzyme is initially inhibited when submitted to flooding.
Regarding nitrogen (N), the lack of oxygen in the root system of the soybean plant impairs the nodulation and inhibits the biological N fixation by the symbiotic bacteria in previously formed nodules [
87,
88]. Consequently, the total nitrogen accumulated in the plant is a more sensitive parameter than the accumulation of biomass in the shoot when analyzing flooding responses [
89]. Soybean plants subjected to flooding for 14 days showed a lower accumulation of dry mass in the roots and shoots [
90]. In another study, reductions of up to 55% were observed in the accumulation of shoot dry mass in soybean plants subjected to flooding [
57].
In the present study, N uptake in the shoots was evaluated at two times, namely, 48 h after tank drainage and at flowering. At 48 h after drainage, there was no significant difference in nitrogen uptake between the treatments (flooding and control) for both genotypes. On the other hand, at flowering, the sensitive genotype FUNDACEP 62 showed a marked decrease in N accumulated in shoots when compared to the control. The tolerant genotype TECIRGA 6070 showed the stability of N uptake, and no significant difference was observed between treatments for this genotype (
Supplementary Figure S5).
2.5. Field Experiments and SNP Validation in the Brazilian Soybean Genotypes
To investigate the contribution of the identified polymorphisms to the flooding tolerance, the phenotypic and grain yield impacts were analyzed in a panel of 166 Brazilian soybean genotypes (
Supplementary Table S2). The phenotyping score of the plants (visual damage scale) was performed after the flooding period in four field experiments conducted in two growing seasons.
In Experiment 1, the flooding stress was imposed for five days, and plant phenotyping was performed six days after water drainage. Nine SNPs (GSM0600, GSM0601, GSM0602, GSM0605, GSM0616, GSM0620, GSM0621, GSM0622, GSM0623, and GSM0625) were identified as possibly related to the flooding response (
Table 5 and
Supplementary Table S4). The markers that presented the lowest (7.31) and highest (8.24) mean visual phenotyping scores were GSM0621 (TT allele) and GSM0601 (AA allele), respectively. However, the TT allele (GSM0621) was not very representative within the sampled genotypes, being found in only 2 out of the 96 genotypes investigated in this experiment (
Table 5).
Regarding the second experiment, flooding was imposed for five days, and three phenotyping studies (visual score evaluations) were performed (at 8, 23 and 43 days after water drainage). The first, second and third phenotyping evaluations presented five, five and four significant SNP markers associated with greater flooding tolerance, respectively (
Table 5). The significant SNPs obtained in all phenotyping evaluations were: GSM0601, GSM0604, GSM0612 and GSM0613. The alleles associated with the highest tolerance were AA (GSM0601 and GSM0604), TT (GSM0612) and GG (GSM0613). The markers corresponding to the lowest (4.63 (second phenotyping)) and the highest (6.79 (first phenotyping)) mean value of the visual score were GSM0619 (GG allele) and GSM0600 (CC allele), respectively, with GSM0619 being significant in the second evaluation of visual score. However, the GG (marker GSM0619) and CC (GSM0600) genotypes were not very representative within the investigated genotypes, being found in only 2 and 3 genotypes, respectively, from 32 genotypes (
Supplementary Table S5).
In Experiment 3, the phenotyping was extended for longer periods: up to 60 days after water drainage. One, three, two, and five SNPs were significant in the first, second, third, and fourth phenotyping evaluations, respectively (
Table 6). The SNPs GSM0612 and GSM0613 were significantly associated with a better response to flooding in three of the four evaluations performed. The alleles related to the highest tolerance were TT (GSM0612) and GG (GSM0613). The markers that presented the lowest (5.32 (fourth phenotyping)) and the highest (6.29 (fourth phenotyping)) visual score mean were GSM0599 (GG allele) and GSM0611 (CC allele), respectively. However, the GG allele (GSM0599) was not very representative within the genotypes used in this experiment, being found in only 2 out of the 26 genotypes investigated (
Table 6 and
Supplementary Table S6).
In Experiment 4, only GSM0604 (TT allele) and GSM0605 (TT allele) were associated with greater tolerance to flooding (lower values on the visual assessment scale and significant
p-value), with the GSM0605 marker being significant for the three evaluations performed (
Table 6). According to this experiment, this marker proved to be efficient in discriminating the best response to flooding of the Brazilian soybean genotypes tested. The markers corresponding to the lowest (6.41 (first phenotyping)) and the highest (7.27 (third phenotyping)) visual score mean were, respectively, GSM0605 (TT allele) and GSM0604 (AA allele), which were both significant for visual score values (
Table 6 and
Supplementary Table S7).
Considering the 22 SNPs analyzed in all the field experiments, 17 had a significantly different phenotyping score among the contrasting genotypes (
Table 5 and
Table 6). Furthermore, the SNPs GSM0601, GSM0604, GSM0605, GSM0612, GS0M613, GSM0616 and GSM0625 were significantly related to flooding response in two different experiments. These results demonstrate the viability and efficiency of these SNPs in identifying differential responses of soybean genotypes to flooding.
In order to better comprehend the tolerance of the Brazilian soybean genotypes to flooding, grain yield was also evaluated and correlated with SNP data (
Table 5 and
Table 6). In Experiment 2, six SNPs were significantly related to higher productivity under flooding: GSM0601 (AA allele), GSM0602 (TT allele), GSM0604 (AA allele), GSM0612 (TT allele), GSM0613 (GG allele) and GSM0625 (AA allele) (
Table 5). In Experiment 3, only two SNPs presented an association with higher productivity: GSM0611 (TT allele) and GSM0625 (AA allele) (
Table 7). The AA allele of the GSM0625 marker showed a significant relationship to higher grain yields in both experiments. The markers that presented groups with the highest yields of 1807 and 1214 kg ha
−1 were GSM0601 (Experiment 2) and GSM0625 (Experiment 3), respectively. On the other hand, the lowest average grain yields were 1222.5 and 802 kg ha
−1, obtained in sample groups of the markers GSM0602 (AA alleles) (Experiment 2) and GSM0600 (CC alleles) (Experiment 3), respectively (
Supplementary Table S8).
To further explore the consequences of flooding on plant development and the validation of the molecular markers, evaluations of plant mortality, normalized difference vegetation index (NDVI), and leaf retention were carried out. Among the 22 analyzed SNPs, it was possible to observe an association in only two (GSM0605 and GSM0611) in Experiment 3 (
Table 7). The GG allele (GSM0605) was associated with higher NDVI and higher leaf retention. Regarding the SNP GSM0611, the TT allele showed greater leaf retention. No significant relationship of the SNPs with plant mortality was observed.
On the other hand, in Experiment 4, the SNPs GSM0605 (TT allele), GSM0612 (TT allele), GSM0613 (GG allele) and GSM0614 (CC allele) were significantly associated with lower mortality of soybean plants. In this same experiment, GSM0604 (TT allele), GSM0605 (TT allele) and GSM0618 (TT allele) were significant for a higher NDVI (
Table 7 and
Supplementary Table S9). Based on the results of plant mortality and NDVI, the TT allele of the GSM0605 has potential to predict a greater tolerance to flooding.
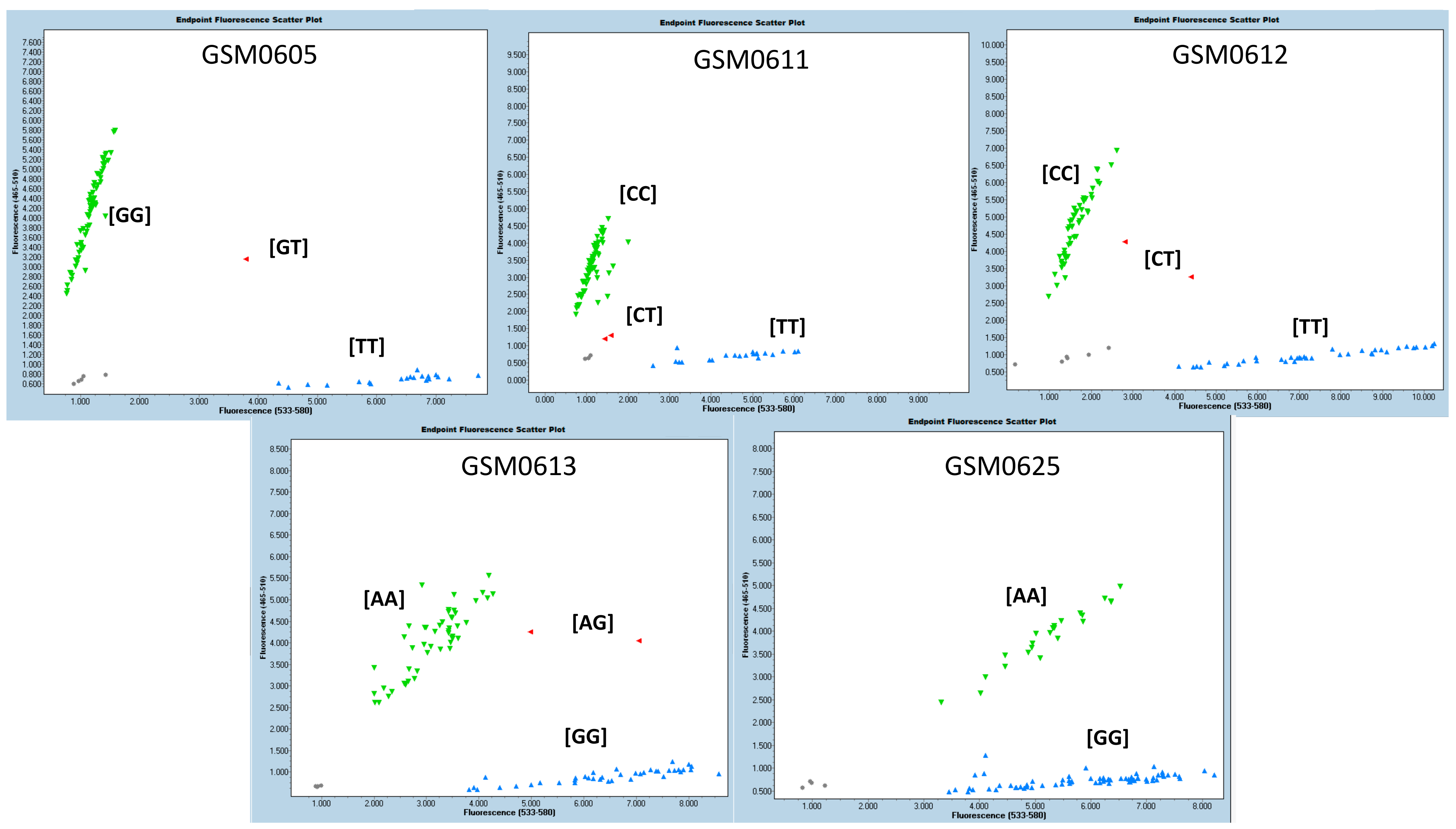
The SNPs that showed statistical significance in at least two characteristics/variables analyzed, or in two experiments, with allelic consistency that exhibited the best performance, were selected. According to the phenotyping (visual score) and grain yield evaluations, five molecular markers were selected as promising for use in assisted selection: GSM0605, GM06011 and GSM0612, GSM0613, and GSM0625 (
Table 8). The methodology used (KASP) precisely grouped the three genotypic classes (two homozygous groups and one heterozygote group) based on these SNPs. In
Figure 2, the graphs generated for the five previously selected markers are shown. The groupings of the three genotypic classes indicate, for each marker, that they can effectively differentiate the evaluated genotypes. The results showed that homozygous genotypes were more frequent than heterozygous ones in all cases. The genotypes with the presence of promising markers were selected through the joint analysis of the five recommended molecular markers. The alleles related to the best performance for the tags GSM0605, GSM0611, GSM0612, GSM0613 and GSM0625 were TT, TT, TT, GG and AA, respectively. Thus, the genotypes BRB 16-253244, HO TERERÊ, SYN 1451946, BRB 16-238251 and NS 6700 simultaneously presented the five alleles associated with greater flooding tolerance. The genotypes BRB 16-236559, TMG 7058, BRB 16-242992, PIO 95R95, BS 1691, DM 66I68, SYN 1561, TEC 7849, M 5838, M 6410, BMX Ponta, CZ 26B42 and M5947 showed four alleles associated with greater tolerance to flooding. The other genotypes analyzed had a maximum of three alleles (including TECIRGA 6070) associated with greater tolerance to flooding (
Table 9).
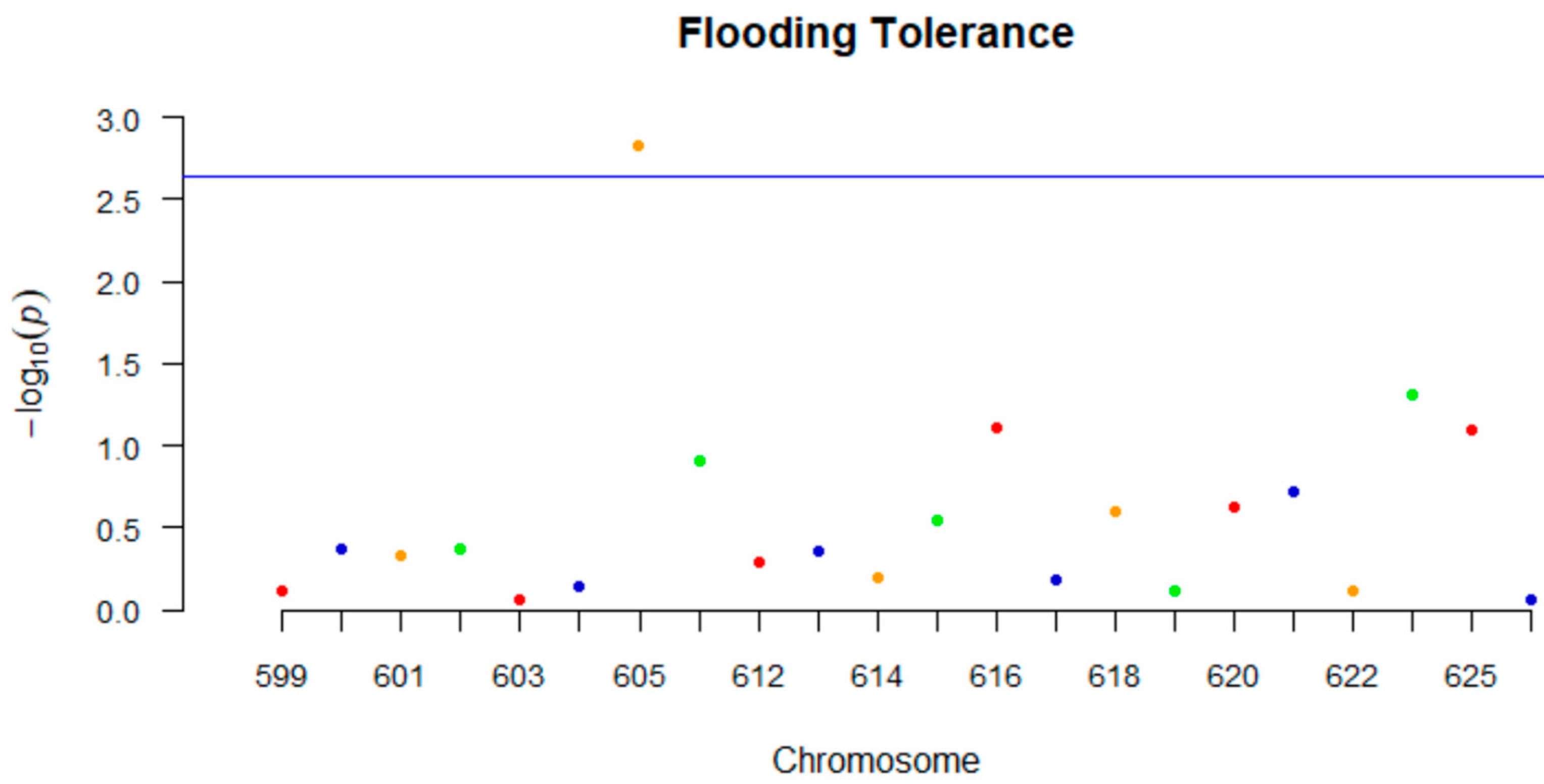
Regarding the GLM analysis, ANOVA indicated that effects of genotype, experiment and replicates were all significant and genotype
x experiment was significant as well (
p < 0.01). Mini-GWAS identified one marker (GSM605) on chromosome 9 that was significant for flooding tolerance (
Figure 3). Single-factor analysis using both BLUPs and fixed effects identified three markers located on chromosomes 9, 10, and 13, respectively, which were significant (
p < 0.05). Of these three markers, one marker was same as the one identified using Mini-GWAS (
Table 10).
SNPs are the most abundant source of DNA polymorphism, with an estimated four to five million SNPs in the soybean genome. A study carried out with the main soybean varieties grown in the United States identified 280 SNPs in more than 76 Kbp of coding sequences [
91]. However, the frequency of SNPs is relatively low in soybean compared to other cultivated crops. This characteristic is mainly due to the low genetic variability of the ancestor and the loss of variability during domestication [
91,
92,
93].
Several studies have sought to understand the genetic and molecular mechanisms of tolerance to flooding in soybean. In this sense, several QTLs have been observed to be associated with greater tolerance to flooding. VanToai et al. [
94] identified a QTL linked to Sat_064, located on chromosome 18, as associated with greater tolerance to flooding using two populations of recombinant strains (RILs). Cornelious et al. [
95] reported five QTL markers associated with flooding tolerance as well. The Satt485 marker is located on chromosome 3, the Satt599 marker on chromosome 5, and three (Satt160, Satt269, and Satt252) markers are located on chromosome 13. They have been associated with flooding tolerance in two populations of recombinant strains. Githiri et al. [
96] identified seven QTLs (ft1 to ft7) associated with grain yield under flooding stress, resulting in a proposed QTL close to Satt100.
Wang et al. [
97] mapped three QTLs-Satt531-A941V (chromosome 1), Satt648-K418_2V (chromosome 5), and Satt038-Satt275 (chromosome 18) that were associated with greater tolerance to flooding in soybean. However, all these previously identified QTLs were discovered using linkage mapping resulting in a restricted mapping resolution due to limited recombination events in a biparental population. To overcome these limitations, high-resolution, high-throughput genomic analysis technologies have been developed, enabling new approaches for marker-assisted selection, such as genomic-wide association studies (GWAS). This type of study aims at genetic mapping through associations between loci and phenotypic characteristics in the population and seeks to detect the effects of genes on the genetic values of individuals [
98].
Based on the above, Wu et al. [
99] performed GWAS using a mapping panel composed of several introductions of soybean plants (PIs) obtained from the USDA germplasm bank to detect SNPs associated with greater tolerance to flooding. The genotypes were originally collected from 17 countries and 9 unknown origins, and were tested for two consecutive years in the field. The plant leaf damage score was used to index the soybean response to flooding stress. Fourteen SNPs were identified as being associated with greater tolerance and five SNPs were in candidate gene coding regions.
Flood-related QTL mapping studies show a wide distribution in the genome, indicating that greater soybean tolerance is a complex quantitative trait controlled by many genes with less effect [
94,
95,
96,
97,
100,
101].
Flooding induces gene expression and phenotypic alterations. Symptoms of flooding stress include leaf chlorosis, defoliation and plant death. The phenotypic variance observed among genotypes is partly due to genetic differences and partly to the difference in the environment, implying the possibility of changes in selection criteria depending on the environment [
102]. Thus, it is important to understand the phenotypic variation that corresponds to the environment and the variation corresponding to the genotype to be able to estimate with better experimental precision the response of genotypes in different environments.
Phenotyping through visual score has been used to select genotypes with greater tolerance to flooding [
103]. It is reported that flooding reduces soybean grain yield by 40 to 50% in genotypes with greater tolerance and by 70% to 80% in genotypes with greater sensitivity to this stress, varying according to the growth stage and duration of stress [
95,
103,
104]. Thus, plants with greater tolerance to flooding are of enormous interest to farmers, as soybean is commonly sensitive to this stress condition [
105].
Previous studies have assessed flooding tolerance in soybeans using field trial methods [
103,
106,
107]. However, a small number of studies have identified QTLs or genes related to traits linked to grain yield. Hu et al. [
108] phenotyped 113 wild soybean accessions related to productive traits, identifying 892 alleles and 18 markers for the evaluated traits. Two markers, namely, sct_010 and satt316, which are associated with grain yield, were expressed stably over two years at two experimental sites. Since these characteristics are complex and quantitative, environmental variations can trigger and modify the expression of genes related to them [
109].
The five SNPs selected in the present study are distributed on chromosomes 3, 4, 9, and 13. Wu et al. [
99] found SNPs associated with a greater tolerance to flooding on chromosomes 3, 4, 7, 13, and 19. Although chromosomes 3, 4, and 13 were found in both studies, no common locus is shared between them. In our study, the validated SNPs were in genes likely to be involved in the variation of tolerance to the flooding conditions. Wu et al. [
99] found SNPs in candidate genes that perform diverse biological functions, including heat stress regulation, protein phosphorylation, DNA repair, histone methylation, and protein degradation.
Among the genes with SNPs associated with higher flooding tolerance, Glyma.03G00410, which presents two SNPs (
Table 3), is located on chromosome 3 and encodes a calcium-binding protein, which converts signals in a wide variety of biochemical alterations [
110]. Once bound to calcium, calmodulin acts as part of a signal transduction pathway, with various target proteins, such as kinases or phosphatases [
65,
111]. Thus, calmodulin can act as a mediator of soybean growth during stress [
63].
In the adaptation of plants to flooding, the ethylene response factor (ERF) genes play an essential role in plant survival strategy through the regulation of gene expression [
112]. The GSM0625 marker is in the Glyma.13G274100 gene, located on chromosome 13 (
Table 3). This gene encodes transcription factors of the APETALA2/ethylene response factor (AP2/ERF) family. These transcription factors play key roles in signal transduction to activate or suppress the expression of defense genes, as well as act in growth regulation and the interaction between different signaling pathways and hormones in plants [
113,
114,
115,
116,
117].
The AP2/ERF family has been characterized and its genes participate in the regulation of flooding tolerance [
118]. After the onset of flooding, AP2/ERFs are induced and, in turn, interact with the
cis elements present in the promoter of stress-responsive genes, providing greater tolerance to this condition [
119]. Microarray studies have shown that several Arabidopsis AP2/ERF genes are induced during hypoxia [
120]. Thus, AP2/ERF is regulated at different levels of transcription and translation to achieve homeostasis during adverse conditions, and is thus a promising candidate for the study of several networks involved in plant development, metabolic and stress responses.
The SNP GSM0605 is located in the Glyma.09G02100 gene, and this locus encodes a Cysteine desulfurase-like protein which is involved in the assembly of iron–sulfur (Fe–S) clusters [
121]. These cofactors are needed in all clades of biology to perform a wide variety of unique functions, including nitrogen fixation, ribosome assembly, DNA repair, mitochondrial respiration, and metabolite catabolism. Fe–S proteins catalyze the conversion of the amino acid cysteine into alanine and elemental sulfur, and are known for their vital role in redox reactions during mitochondrial electron transport [
122], biotin and thiamine formation, gene expression, and other cellular processes [
123,
124].
In addition to robust flooding tolerance assays to generate phenotypic data, genotypic assessments are also important for the development and selection of molecular markers. The outcomes of plant protein function and gene expression in response to a changing environment may result from SNPs in coding regions and regulatory sequences, respectively. The present study demonstrates that Brazilian soybean genotypes present allelic variation (
Figure 2 and
Table 9).
The increase in the commercial value of soybeans on the international market has resulted in an increase in the cultivated area of this crop under different climatic and soil conditions in many regions of the world, including soils subject to periods of flooding [
125]. A great challenge is the difficult adaptation of soybean to these conditions, since most genotypes are sensitive to excess water. A possible strategy is the identification and development of soybean genotypes more adapted to areas subject to flooding and that have a high grain yield potential. For this, it is important to recognize and understand the mechanisms of soybean response to flooding to improve grain productivity when grown in these areas.
In Rio Grande do Sul state, Brazil, soybean cultivation is strongly associated with crop rotation with irrigated rice in lowland areas. In the 2021/2022 growing season, the area cultivated with soybeans in this environment in this state was approximately 380,000 ha. Thus, considering this important agricultural frontier for the expansion of soybean-cultivated areas, the obtention of genotypes with greater tolerance to flooding should be one of the goals of breeding programs [
103,
106,
126,
127,
128]. This work provides some potential molecular markers that could be used for soybean genetic improvement and plant breeding. One of the advantages of SNP use is that the location of the variants in coding regions associated with biological and agronomic characteristics can be recognized, and phenotypes can be projected by genotypes, accelerating the selection of more tolerant plants to flooding.


 ,
, 


{kind=link}
{kind=link}
{kind=link}
{kind=link}