
Metabolism and Signaling of Plant Mitochondria in Adaptation to Environmental Stresses

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
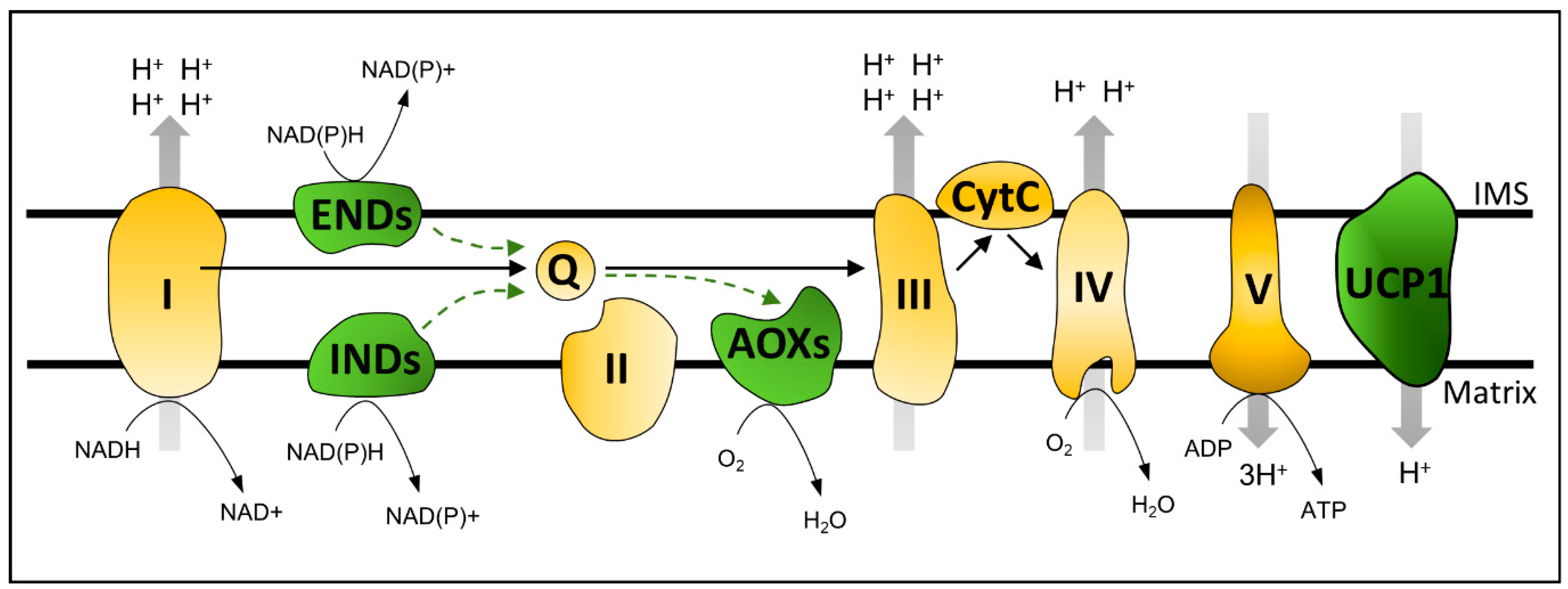
2. Plant Oxidative Phosphorylation Bypasses: Importance for Mitochondrial Signaling and Stress Response
2.1. UNCOUPLING PROTEINS
2.2. ALTERNATIVE OXIDASES and ALTERNATIVE NAD(P)H DEHYDROGENASES
2.2.1. ALTERNATIVE OXIDASES
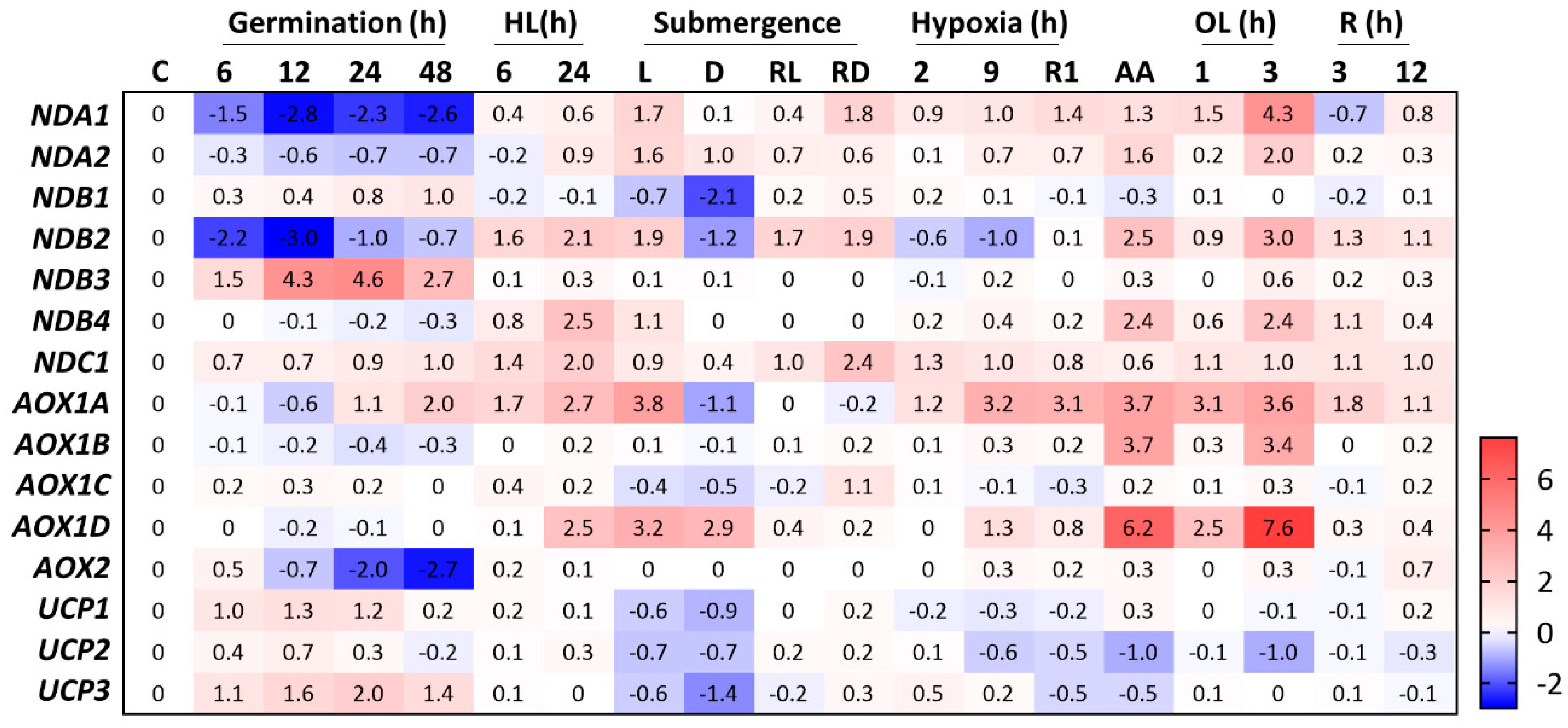
2.2.2. ALTERNATIVE NAD(P)H DEHYDROGENASES
Internal NADP(H) DEHYDROGENASES
External NAD(P)H DEHYDROGENASEs
3. Mitochondrial Retrograde Signaling and Crosstalk with Chloroplasts
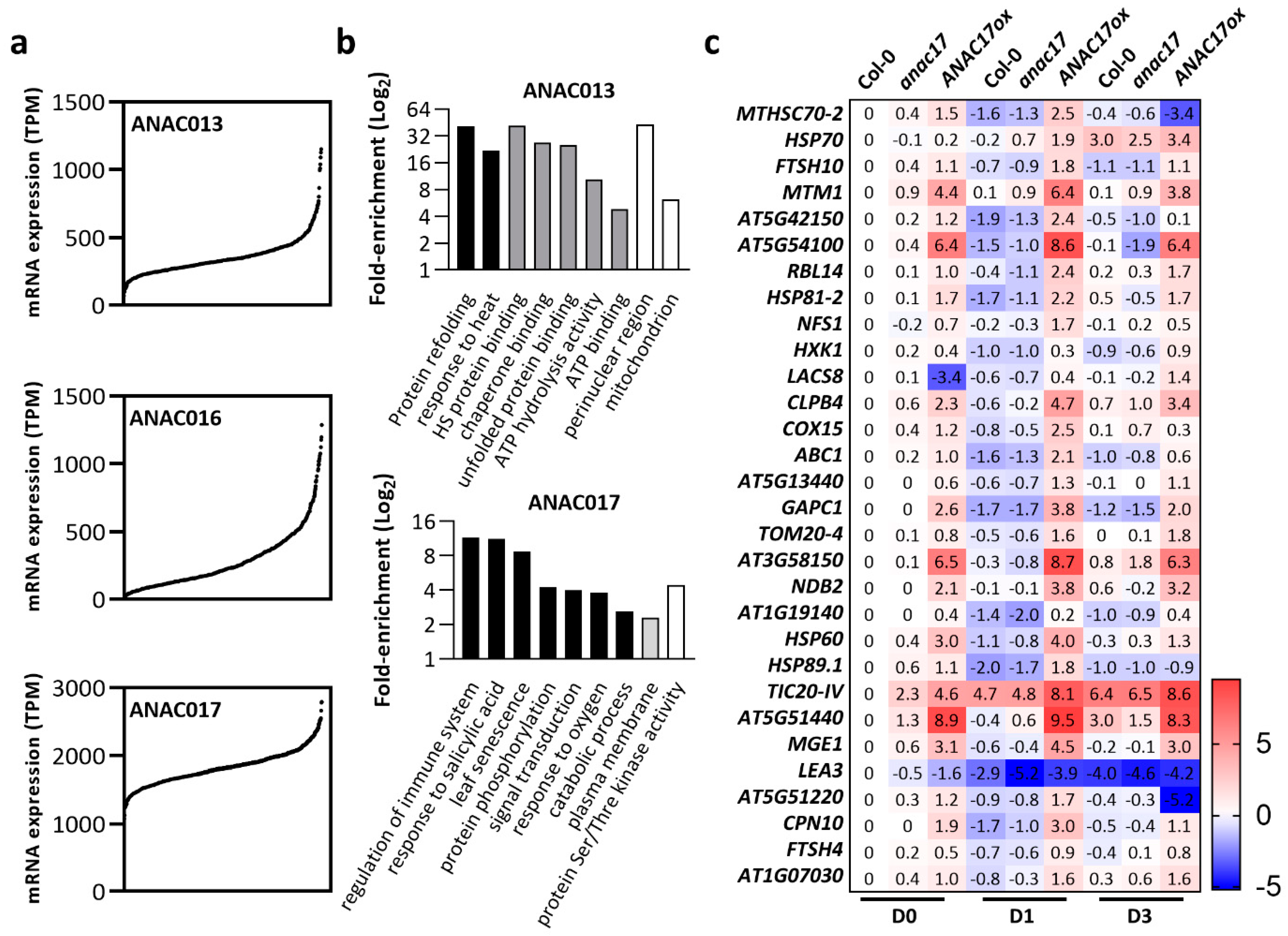
3.1. Arabidopsis NO APICAL MERISTEM (ANAC) Transcription Factors
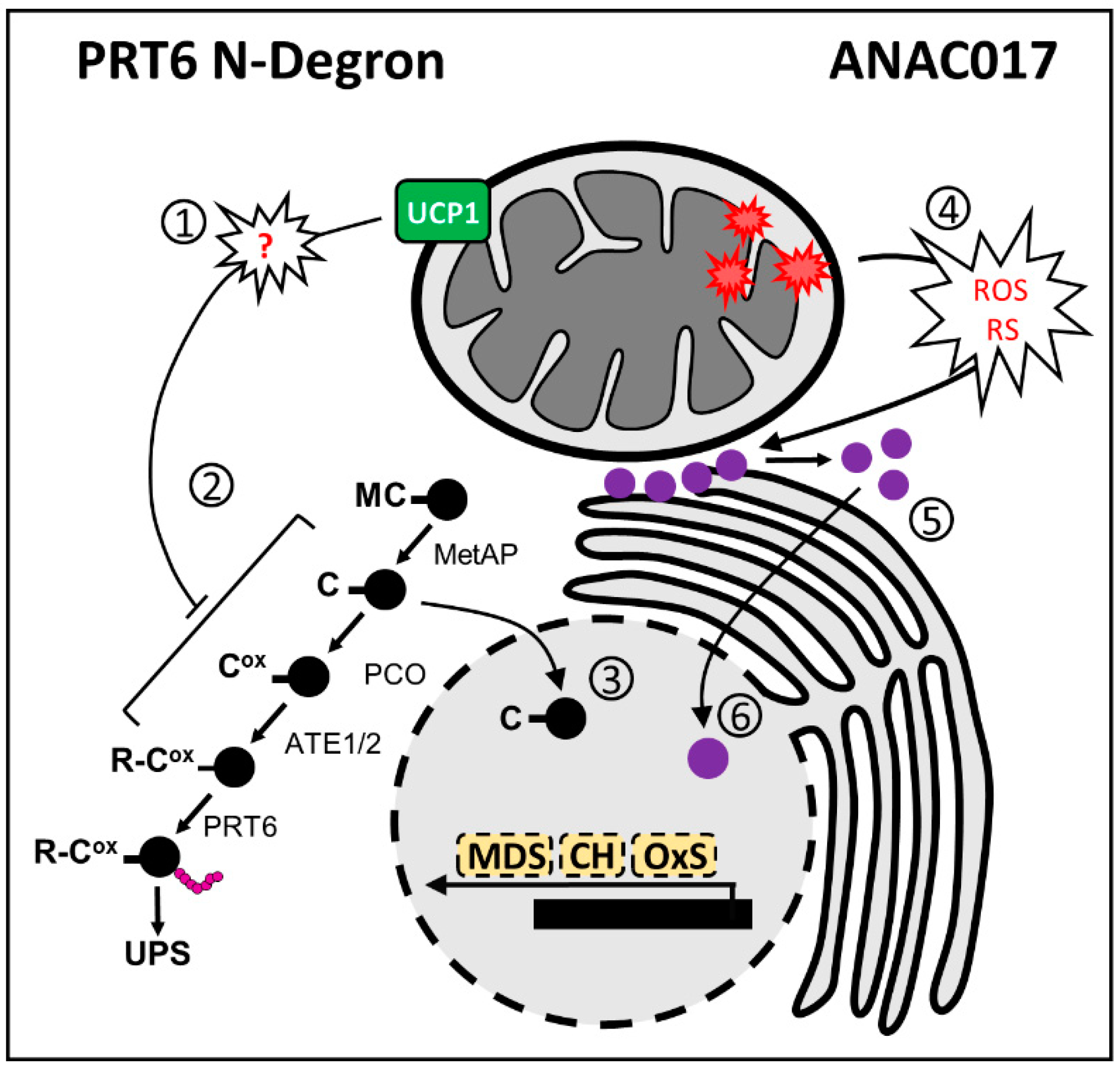
3.2. PRT6 N-Degron
3.3. Crosstalk between ANAC and PRT6 N-Degron in Mitochondrial Retrograde Signaling
4. Mitochondria–Chloroplast Crosstalk
5. Inner Membrane Transporters
6. Manipulating Mitochondrial Metabolism to Improve Plant Resistance against Environmental Stresses
7. Conclusions and Perspectives
Supplementary Materials
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chandel, N.S. Evolution of Mitochondria as Signaling Organelles. Cell Metab. 2015, 22, 204–206. [Google Scholar] [CrossRef] [PubMed]
- Dourmap, C.; Morin, A.; Caubrière, D.; Kerdiles, M.; Le Moullec, J.; Couée, I. Stress Signalling Dynamics of the Mitochondrial Electron Transport Chain and Oxidative Phosphorylation System in Higher Plants. Ann. Bot. 2020, 125, 721–736. [Google Scholar] [CrossRef]
- van Aken, O. Mitochondrial Redox Systems as Central Hubs in Plant Metabolism and Signaling. Plant Physiol. 2021, 186, 36–52. [Google Scholar] [CrossRef] [PubMed]
- Paszkiewicz, G.; Gualberto, J.M.; Benamar, A.; Macherel, D.; Logan, D.C. Arabidopsis Seed Mitochondria Are Bioenergetically Active Immediately upon Imbibition and Specialize via Biogenesis in Preparation for Autotrophic Growth. Plant Cell 2017, 29, 109–128. [Google Scholar] [CrossRef] [PubMed]
- Eme, L.; Spang, A.; Lombard, J.; Stairs, C.W.; Ettema, T.J.G. Archaea and the Origin of Eukaryotes. Nat. Rev. Microbiol. 2017, 15, 711–723. [Google Scholar] [CrossRef] [PubMed]
- Møller, I.M.; Rasmusson, A.G.; van Aken, O. Plant Mitochondria—Past, Present and Future. Plant J. 2021, 108, 912–959. [Google Scholar] [CrossRef]
- Martin, W.; Müller, M. The Hydrogen Hypothesis for the First Eukaryote. Nature 1998, 392, 37–41. [Google Scholar] [CrossRef]
- Ueda, M.; Kadowaki, K.I. Gene Content and Gene Transfer from Mitochondria to the Nucleus During Evolution. In Advances in Botanical Research; Academic Press Inc.: Cambridge, MA, USA, 2012; Volume 63, pp. 21–40. [Google Scholar]
- Schwarzländer, M.; Logan, D.C.; Johnston, I.G.; Jones, N.S.; Meyer, A.J.; Fricker, M.D.; Sweetlove, L.J. Pulsing of Membrane Potential in Individual Mitochondria: A Stress-Induced Mechanism to Regulate Respiratory Bioenergetics in Arabidopsis. Plant Cell 2012, 24, 1188–1201. [Google Scholar] [CrossRef]
- Vercesi, A.E.; Martins, I.S.; Silva, M.A.P.; Leite, H.M.F. PUMPing Plants. Nature 1995, 375, 24. [Google Scholar] [CrossRef]
- Fuchs, P.; Rugen, N.; Carrie, C.; Elsasser, M.; Finkemeier, I.; Giese, J.; Hildebrandt, T.M.; Kühn, K.; Maurino, V.G.; Ruberti, C.; et al. Single Organelle Function and Organization as Estimated from Arabidopsis Mitochondrial Proteomics. Plant J. 2019, 101, 420–441. [Google Scholar] [CrossRef]
- Vercesi, A.E.; Borecký, J.; Maia, I.d.G.; Arruda, P.; Cuccovia, I.M.; Chaimovich, H. Plant Uncoupling Mitochondrial Proteins. Annu. Rev. Plant Biol. 2006, 57, 383–404. [Google Scholar] [CrossRef] [PubMed]
- Maia, I.G.; Benedetti, C.E.; Leite, A.; Turcinelli, S.R.; Vercesi, A.E.; Arruda, P. AtPUMP: An Arabidopsis Gene Encoding a Plant Uncoupling Mitochondrial Protein. FEBS Lett. 1998, 429, 403–406. [Google Scholar] [CrossRef]
- Monné, M.; Daddabbo, L.; Gagneul, D.; Obata, T.; Hielscher, B.; Palmieri, L.; Miniero, D.V.; Fernie, A.R.; Weber, A.P.M.; Palmieri, F. Uncoupling Proteins 1 and 2 (UCP1 and UCP2) From Arabidopsis Thaliana Are Mitochondrial Transporters of Aspartate, Glutamate, and Dicarboxylates. J. Biol. Chem. 2018, 293, 4213–4227. [Google Scholar] [CrossRef] [PubMed]
- Barreto, P.; Couñago, R.M.; Arruda, P. Mitochondrial Uncoupling Protein-Dependent Signaling in Plant Bioenergetics and Stress Response. Mitochondrion 2020, 53, 109–120. [Google Scholar] [CrossRef]
- Pastore, D.; Trono, D.; Laus, M.N.; di Fonzo, N.; Flagella, Z. Possible Plant Mitochondria Involvement in Cell Adaptation to Drought Stress—A Case Study: Durum Wheat Mitochondria. J. Exp. Bot. 2007, 58, 195–210. [Google Scholar] [CrossRef] [PubMed]
- Garmash, E.V.; Belykh, E.S.; Velegzhaninov, I.O. The Gene Expression Profiles of Mitochondrial Respiratory Components in Arabidopsis Plants with Differing Amounts of ALTERNATIVE OXIDASE1a under High Intensity Light. Plant Signal. Behav. 2021, 16, 1864962. [Google Scholar] [CrossRef]
- Hruz, T.; Laule, O.; Szabo, G.; Wessendorp, F.; Bleuler, S.; Oertle, L.; Widmayer, P.; Gruissem, W.; Zimmermann, P. Genevestigator V3: A Reference Expression Database for the Meta-Analysis of Transcriptomes. Adv. Bioinform. 2008, 2008, 420747. [Google Scholar] [CrossRef]
- Begcy, K.; Mariano, E.D.; Mattiello, L.; Nunes, A.V.; Mazzafera, P.; Maia, I.G.; Menossi, M. An Arabidopsis Mitochondrial Uncoupling Protein Confers Tolerance to Drought and Salt Stress in Transgenic Tobacco Plants. PLoS ONE 2011, 6, e23776. [Google Scholar] [CrossRef]
- Barreto, P.; Okura, V.K.; Neshich, I.A.P.; Maia, I.D.G.; Arruda, P. Overexpression of UCP1 in Tobacco Induces Mitochondrial Biogenesis and Amplifies a Broad Stress Response. BMC Plant Biol. 2014, 14, 144. [Google Scholar] [CrossRef]
- Barreto, P.; Yassitepe, J.E.C.T.; Wilson, Z.A.; Arruda, P. Mitochondrial Uncoupling Protein 1 Overexpression Increases Yield in Nicotiana Tabacum under Drought Stress by Improving Source and Sink Metabolism. Front. Plant Sci. 2017, 8, 1836. [Google Scholar] [CrossRef]
- Fuchs, P.; Bohle, F.; Lichtenauer, S.; Ugalde, J.M.; Feitosa Araujo, E.; Mansuroglu, B.; Ruberti, C.; Wagner, S.; Müller-Schüssele, S.J.; Meyer, A.J.; et al. Reductive Stress Triggers ANAC017-Mediated Retrograde Signaling to Safeguard the Endoplasmic Reticulum by Boosting Mitochondrial Respiratory Capacity. Plant Cell 2022, 34, 1375–1395. [Google Scholar] [CrossRef] [PubMed]
- Sweetlove, L.J.; Lytovchenko, A.; Morgan, M.; Nunes-Nesi, A.; Taylor, N.L.; Baxter, C.J.; Eickmeier, I.; Fernie, A.R. Mitochondrial Uncoupling Protein Is Required for Efficient Photosynthesis. Proc. Natl. Acad. Sci. USA 2006, 103, 19587–19592. [Google Scholar] [CrossRef]
- Lima, R.P.M.; Nunes-Laitz, A.V.; Arcuri, M.d.L.C.; Campos, F.G.; Joca, T.A.C.; Monteiro, G.C.; Kushima, H.; Lima, G.P.P.; de Almeida, L.F.R.; Barreto, P.; et al. The Double Knockdown of the Mitochondrial Uncoupling Protein Isoforms Reveals Partial Redundant Roles During Arabidopsis Thaliana Vegetative and Reproductive Development. Plant Sci. 2022, 322, 111365. [Google Scholar] [CrossRef] [PubMed]
- Ng, S.; Giraud, E.; Duncan, O.; Law, S.R.; Wang, Y.; Xu, L.; Narsai, R.; Carrie, C.; Walker, H.; Day, D.A.; et al. Cyclin-Dependent Kinase E1 (CDKE1) Provides a Cellular Switch in Plants between Growth and Stress Responses. J. Biol. Chem. 2013, 288, 3449–3459. [Google Scholar] [CrossRef] [PubMed]
- Geisler, D.A.; Päpke, C.; Obata, T.; Nunes-Nesi, A.; Matthes, A.; Schneitz, K.; Maximova, E.; Araújo, W.L.; Fernie, A.R.; Persson, S. Downregulation of the δ-Subunit Reduces Mitochondrial ATP Synthase Levels, Alters Respiration, and Restricts Growth and Gametophyte Development in Arabidopsis. Plant Cell 2012, 24, 2792–2811. [Google Scholar] [CrossRef]
- Garmier, M.; Carroll, A.J.; Delannoy, E.; Vallet, C.; Day, D.A.; Small, I.D.; Millar, A.H. Complex I Dysfunction Redirects Cellular and Mitochondrial Metabolism in Arabidopsis. Plant Physiol. 2008, 148, 1324–1341. [Google Scholar] [CrossRef]
- Welchen, E.; García, L.; Mansilla, N.; Gonzalez, D.H. Coordination of Plant Mitochondrial Biogenesis: Keeping Pace with Cellular Requirements. Front. Plant Sci. 2014, 4, 551. [Google Scholar] [CrossRef]
- Barreto, P.; Dambire, C.; Sharma, G.; Vicente, J.; Osborne, R.; Yassitepe, J.; Gibbs, D.J.; Maia, I.G.; Holdsworth, M.J.; Arruda, P. Mitochondrial Retrograde Signaling through UCP1-Mediated Inhibition of the Plant Oxygen-Sensing Pathway. Curr. Biol. 2022, 32, 1403–1411. [Google Scholar] [CrossRef]
- Arcuri, M.D.L.C.; Nunes-Laitz, A.V.; Lima, R.P.M.; Barreto, P.; Marinho, A.N.; Arruda, P.; Maia, I.G. Knockdown of Mitochondrial Uncoupling Proteins 1 and 2 (AtUCP1 and 2) in Arabidopsis Thaliana Impacts Vegetative Development and Fertility. Plant Cell Physiol. 2021, 62, 1630–1644. [Google Scholar] [CrossRef]
- Rasmusson, A.G.; Geisler, D.A.; Møller, I.M. The Multiplicity of Dehydrogenases in the Electron Transport Chain of Plant Mitochondria. Mitochondrion 2008, 8, 47–60. [Google Scholar] [CrossRef]
- Vanlerberghe, C.C.; Mclntosh, L. Signals Regulating the Expression of the Nuclear Gene Encoding Alternative Oxidase of Plant Mitochondria. Plant Physiol. 1996, 11, 589–595. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, K.; Noguchi, K. Differential Gene Expression Profiles of the Mitochondrial Respiratory Components in Illuminated Arabidopsis Leaves. Plant Cell Physiol. 2009, 50, 1449–1462. [Google Scholar] [CrossRef] [PubMed]
- Rasmusson, A.G.; Agius, S.C. Rotenone-Insensitive NAD(P)H Dehydrogenases in Plants: Immunodetection and Distribution of Native Proteins in Mitochondria. Plant Physiol. Biochem. 2001, 39, 1057–1066. [Google Scholar] [CrossRef]
- Ho, L.H.M.; Giraud, E.; Lister, R.; Thirkettle-Watts, D.; Low, J.; Clifton, R.; Howell, K.A.; Carrie, C.; Donald, T.; Whelan, J. Characterization of the Regulatory and Expression Context of an Alternative Oxidase Gene Provides Insights into Cyanide-Insensitive Respiration during Growth and Development. Plant Physiol. 2007, 143, 1519–1533. [Google Scholar] [CrossRef] [PubMed]
- Elhafez, D.; Murcha, M.W.; Clifton, R.; Soole, K.L.; Day, D.A.; Whelan, J. Characterization of Mitochondrial Alternative NAD(P)H Dehydrogenases in Arabidopsis: Intraorganelle Location and Expression. Plant Cell Physiol. 2006, 47, 43–54. [Google Scholar] [CrossRef]
- Escobar, M.A.; Franklin, K.A.; Svensson, Å.S.; Salter, M.G.; Whitelam, G.C.; Rasmusson, A.G. Light Regulation of the Arabidopsis Respiratory Chain. Multiple Discrete Photoreceptor Responses Contribute to Induction of Type II NAD(P)H Dehydrogenase Genes. Plant Physiol. 2004, 136, 2710–2721. [Google Scholar] [CrossRef]
- Clifton, R.; Lister, R.; Parker, K.L.; Sappl, P.G.; Elhafez, D.; Millar, A.H.; Day, D.A.; Whelan, J. Stress-Induced Co-Expression of Alternative Respiratory Chain Components in Arabidopsis Thaliana. Plant Mol. Biol. 2005, 58, 193–212. [Google Scholar] [CrossRef]
- Florez-Sarasa, I.; Ribas-Carbo, M.; Del-Saz, N.F.; Schwahn, K.; Nikoloski, Z.; Fernie, A.R.; Flexas, J. Unravelling the in Vivo Regulation and Metabolic Role of the Alternative Oxidase Pathway in C3 Species under Photoinhibitory Conditions. New Phytol. 2016, 212, 66–79. [Google Scholar] [CrossRef]
- Dahal, K.; Vanlerberghe, G.C. Alternative Oxidase Respiration Maintains Both Mitochondrial and Chloroplast Function during Drought. New Phytol. 2017, 213, 560–571. [Google Scholar] [CrossRef]
- Dahal, K.; Wang, J.; Martyn, G.D.; Rahimy, F.; Vanlerberghe, G.C. Mitochondrial Alternative Oxidasemaintains Respiration and Preserves Photosynthetic Capacity during Moderate Drought in Nicotiana Tabacum. Plant Physiol. 2014, 166, 1560–1574. [Google Scholar] [CrossRef]
- Del-Saz, N.F.; Iglesias-Sanchez, A.; Alonso-Forn, D.; López-Gómez, M.; Palma, F.; Clemente-Moreno, M.J.; Fernie, A.R.; Ribas-Carbo, M.; Florez-Sarasa, I. The Lack of Alternative Oxidase 1a Restricts in Vivo Respiratory Activity and Stress-Related Metabolism for Leaf Osmoprotection and Redox Balancing Under Sudden Acute Water and Salt Stress in Arabidopsis Thaliana. Front Plant Sci. 2022, 13, 833113. [Google Scholar] [CrossRef] [PubMed]
- Vanlerberghe, G.C.; Dahal, K.; Alber, N.A.; Chadee, A. Photosynthesis, Respiration and Growth: A Carbon and Energy Balancing Act for Alternative Oxidase. Mitochondrion 2020, 52, 197–211. [Google Scholar] [CrossRef] [PubMed]
- Polidoros, A.N.; Mylona, P.V.; Arnholdt-Schmitt, B. Aox Gene Structure, Transcript Variation and Expression in Plants. Physiol. Plant. 2009, 137, 342–353. [Google Scholar] [CrossRef]
- Clifton, R.; Millar, A.H.; Whelan, J. Alternative Oxidases in Arabidopsis: A Comparative Analysis of Differential Expression in the Gene Family Provides New Insights into Function of Non-Phosphorylating Bypasses. Biochim. Biophys. Acta Bioenerg. 2006, 1757, 730–741. [Google Scholar] [CrossRef]
- Oh, G.G.K.; O’Leary, B.M.; Signorelli, S.; Harvey Millar, A. Alternative Oxidase (AOX) 1a and 1d Limit Prolineinduced Oxidative Stress and Aid Salinity Recovery in Arabidopsis. Plant Physiol. 2022, 188, 1521–1536. [Google Scholar] [CrossRef] [PubMed]
- Narsai, R.; Law, S.R.; Carrie, C.; Xu, L.; Whelan, J. In-Depth Temporal Transcriptome Profiling Reveals a Crucial Developmental Switch with Roles for RNA Processing and Organelle Metabolism That Are Essential for Germination in Arabidopsis. Plant Physiol. 2011, 157, 1342–1362. [Google Scholar] [CrossRef]
- Strodtkötter, I.; Padmasree, K.; Dinakar, C.; Speth, B.; Niazi, P.S.; Wojtera, J.; Voss, I.; Do, P.T.; Nunes-Nesi, A.; Fernie, A.R.; et al. Induction of the AOX1D Isoform of Alternative Oxidase in A. Thaliana T-DNA Insertion Lines Lacking Isoform AOX1A Is Insufficient to Optimize Photosynthesis When Treated with Antimycin a. Mol. Plant 2009, 2, 284–297. [Google Scholar] [CrossRef]
- Amor, Y.; Chevion, M.; Levine, A. Anoxia Pretreatment Protects Soybean Cells against H2O2-Induced Cell Death: Possible Involvement of Peroxidases and of Alternative Oxidase. FEBS Lett. 2000, 477, 175–180. [Google Scholar] [CrossRef]
- van Dongen, J.T.; Fröhlich, A.; Ramírez-Aguilar, S.J.; Schauer, N.; Fernie, A.R.; Erban, A.; Kopka, J.; Clark, J.; Langer, A.; Geigenberger, P. Transcript and Metabolite Profiling of the Adaptive Response to Mild Decreases in Oxygen Concentration in the Roots of Arabidopsis Plants. Ann. Bot. 2009, 103, 269–280. [Google Scholar] [CrossRef]
- Christianson, J.A.; Llewellyn, D.J.; Dennis, E.S.; Wilson, I.W. Comparisons of Early Transcriptome Responses to Low-Oxygen Environments in Three Dicotyledonous Plant Species. Plant Signal. Behav. 2010, 5, 1006–1009. [Google Scholar] [CrossRef]
- Branco-Price, C.; Kaiser, K.A.; Jang, C.J.H.; Larive, C.K.; Bailey-Serres, J. Selective MRNA Translation Coordinates Energetic and Metabolic Adjustments to Cellular Oxygen Deprivation and Reoxygenation in Arabidopsis Thaliana. Plant J. 2008, 56, 743–755. [Google Scholar] [CrossRef]
- Meng, X.; Li, L.; Narsai, R.; de Clercq, I.; Whelan, J.; Berkowitz, O. Mitochondrial Signalling Is Critical for Acclimation and Adaptation to Flooding in Arabidopsis Thaliana. Plant J. 2020, 103, 227–247. [Google Scholar] [CrossRef] [PubMed]
- Voesenek, L.A.C.J.; Bailey-Serres, J. Flood Adaptive Traits and Processes: An Overview. New Phytol. 2015, 206, 57–73. [Google Scholar] [CrossRef]
- Gupta, K.J.; Kumari, A.; Florez-Sarasa, I.; Fernie, A.R.; Igamberdiev, A.U. Interaction of Nitric Oxide with the Components of the Plant Mitochondrial Electron Transport Chain. J. Exp. Bot. 2018, 69, 3413–3424. [Google Scholar] [CrossRef] [PubMed]
- Gupta, K.J.; Stoimenova, M.; Kaiser, W.M. In Higher Plants, Only Root Mitochondria, but Not Leaf Mitochondria Reduce Nitrite to NO, in Vitro and in Situ. J. Exp. Bot. 2005, 56, 2601–2609. [Google Scholar] [CrossRef] [PubMed]
- Poyton, R.O.; Ball, K.A.; Castello, P.R. Mitochondrial Generation of Free Radicals and Hypoxic Signaling. Trends Endocrinol. Metab. 2009, 20, 332–340. [Google Scholar] [CrossRef]
- Millar, A.H.; Day, D. Alternative Solutions to Radical Problems. Trends Plant Sci. 1997, 2, 290. [Google Scholar] [CrossRef]
- Cvetkovska, M.; Vanlerberghe, G.C. Alternative Oxidase Modulates Leaf Mitochondrial Concentrations of Superoxide and Nitric Oxide. New Phytol. 2012, 195, 32–39. [Google Scholar] [CrossRef]
- Jayawardhane, J.; Cochrane, D.W.; Vyas, P.; Bykova, N.V.; Vanlerberghe, G.C.; Igamberdiev, A.U. Roles for Plant Mitochondrial Alternative Oxidase Under Normoxia, Hypoxia, and Reoxygenation Conditions. Front. Plant Sci. 2020, 11, 566. [Google Scholar] [CrossRef]
- Popov, V.N.; Syromyatnikov, M.Y.; Fernie, A.R.; Chakraborty, S.; Gupta, K.J.; Igamberdiev, A.U. The Uncoupling of Respiration in Plant Mitochondria: Keeping Reactive Oxygen and Nitrogen Species under Control. J. Exp. Bot. 2021, 72, 793–807. [Google Scholar] [CrossRef]
- Vishwakarma, A.; Kumari, A.; Mur, L.A.J.; Gupta, K.J. A Discrete Role for Alternative Oxidase under Hypoxia to Increase Nitric Oxide and Drive Energy Production. Free Radic. Biol. Med. 2018, 122, 40–51. [Google Scholar] [CrossRef] [PubMed]
- Zeidler, D.; Zä Hringer, U.; Gerber, I.; Dubery, I.; Hartung, T.; Bors, W.; Hutzler, P.; Rg Durner, J. Innate Immunity in Arabidopsis Thaliana: Lipopolysaccharides Activate Nitric Oxide Synthase (NOS) and Induce Defense Genes. Proc. Natl. Acad. Sci. USA 2004, 101, 15811–15816. [Google Scholar] [CrossRef] [PubMed]
- Kumari, A.; Pathak, P.K.; Bulle, M.; Igamberdiev, A.U.; Gupta, K.J. Alternative Oxidase Is an Important Player in the Regulation of Nitric Oxide Levels under Normoxic and Hypoxic Conditions in Plants. J. Exp. Bot. 2019, 70, 4345–4354. [Google Scholar] [CrossRef] [PubMed]
- Sweetman, C.; Waterman, C.D.; Rainbird, B.M.; Smith, P.M.C.; Jenkins, C.D.; Day, D.A.; Soolea, K.L. Atndb2 Is the Main External NADH Dehydrogenase in Mitochondria and Is Important for Tolerance to Environmental Stress. Plant Physiol. 2019, 181, 774–788. [Google Scholar] [CrossRef] [PubMed]
- Michalecka, A.M.; Svensson, Å.S.; Johansson, F.I.; Agius, S.C.; Johanson, U.; Brennicke, A.; Binder, S.; Rasmusson, A.G. Arabidopsis Genes Encoding Mitochondrial Type II NAD(P)H Dehydrogenases Have Different Evolutionary Origin and Show Distinct Responses to Light. Plant Physiol. 2003, 133, 642–652. [Google Scholar] [CrossRef]
- Wallström, S.V.; Florez-Sarasa, I.; Araújo, W.L.; Escobar, M.A.; Geisler, D.A.; Aidemark, M.; Lager, I.; Fernie, A.R.; Ribas-Carbó, M.; Rasmusson, A.G. Suppression of NDA-Type Alternative Mitochondrial NAD(P)H Dehydrogenases in Arabidopsis Thaliana Modifies Growth and Metabolism, but Not High Light Stimulation of Mitochondrial Electron Transport. Plant Cell Physiol. 2014, 55, 881–896. [Google Scholar] [CrossRef]
- Ferro, M.; Brugière, S.; Salvi, D.; Seigneurin-Berny, D.; Court, M.; Moyet, L.; Ramus, C.; Miras, S.; Mellal, M.; le Gall, S.; et al. AT-CHLORO, a Comprehensive Chloroplast Proteome Database with Subplastidial Localization and Curated Information on Envelope Proteins. Mol. Cell. Proteom. 2010, 9, 1063–1084. [Google Scholar] [CrossRef]
- Zybailov, B.; Rutschow, H.; Friso, G.; Rudella, A.; Emanuelsson, O.; Sun, Q.; van Wijk, K.J. Sorting Signals, N-Terminal Modifications and Abundance of the Chloroplast Proteome. PLoS ONE 2008, 3, e1994. [Google Scholar] [CrossRef]
- Rasmusson, A.G.; Soole, K.L.; Elthon, T.E. Alternative NAD(P)H Dehydrogenases of Plant Mitochondria. Annu. Rev. Plant Biol. 2004, 55, 23–39. [Google Scholar] [CrossRef]
- Svensson, A.S.; Rasmusson, A.G. Light-Dependent Gene Expression for Proteins in the Respiratory Chain of Potato Leaves. Plant J. 2001, 28, 73–82. [Google Scholar] [CrossRef]
- van Aken, O.; Zhang, B.; Law, S.; Narsai, R.; Whelan, J. AtWRKY40 and AtWRKY63 Modulate the Expression of Stress-Responsive Nuclear Genes Encoding Mitochondrial and Chloroplast Proteins. Plant Physiol. 2013, 162, 254–271. [Google Scholar] [CrossRef] [PubMed]
- Kleine, T.; Kindgren, P.; Benedict, C.; Hendrickson, L.; Strand, Å. Genome-Wide Gene Expression Analysis Reveals a Critical Role for CRYPTOCHROME1 in the Response of Arabidopsis to High Irradiance. Plant Physiol. 2007, 144, 1391–1406. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Leydon, A.R.; Manziello, A.; Pandey, R.; Mount, D.; Denic, S.; Vasic, B.; Johnson, M.A.; Palanivelu, R. Penetration of the Stigma and Style Elicits a Novel Transcriptome in Pollen Tubes, Pointing to Genes Critical for Growth in a Pistil. PLoS Genet. 2009, 5, e1000621. [Google Scholar] [CrossRef]
- Huang, J.; Zhao, X.; Chory, J. The Arabidopsis Transcriptome Responds Specifically and Dynamically to High Light Stress. Cell Rep. 2019, 29, 4186–4199. [Google Scholar] [CrossRef] [PubMed]
- Jethva, J.; Schmidt, R.R.; Sauter, M.; Selinski, J. Try or Die: Dynamics of Plant Respiration and How to Survive Low Oxygen Conditions. Plants 2022, 11, 205. [Google Scholar] [CrossRef]
- Giuntoli, B.; Shukla, V.; Maggiorelli, F.; Giorgi, F.M.; Lombardi, L.; Perata, P.; Licausi, F. Age-Dependent Regulation of ERF-VII Transcription Factor Activity in Arabidopsis Thaliana. Plant Cell Environ. 2017, 40, 2333–2346. [Google Scholar] [CrossRef] [PubMed]
- Gibbs, D.J.; Lee, S.C.; Md Isa, N.; Gramuglia, S.; Fukao, T.; Bassel, G.W.; Correia, C.S.; Corbineau, F.; Theodoulou, F.L.; Bailey-Serres, J.; et al. Homeostatic Response to Hypoxia Is Regulated by the N-End Rule Pathway in Plants. Nature 2011, 479, 415–418. [Google Scholar] [CrossRef]
- Geisler, D.A.; Broselid, C.; Hederstedt, L.; Rasmusson, A.G. Ca2+-Binding and Ca2+-Independent Respiratory NADH and NADPH Dehydrogenases of Arabidopsis Thaliana. J. Biol. Chem. 2007, 282, 28455–28464. [Google Scholar] [CrossRef]
- Michalecka, A.M.; Agius, S.C.; Møller, I.M.; Rasmusson, A.G. Identification of a Mitochondrial External NADPH Dehydrogenase by Overexpression in Transgenic Nicotiana Sylvestris. Plant J. 2004, 37, 415–425. [Google Scholar] [CrossRef]
- Kawakatsu, T.; Huang, S.-S.C.; Jupe, F.; Sasaki, E.; Schmitz, R.J.J.; Urich, M.A.A.; Castanon, R.; Nery, J.R.R.; Barragan, C.; He, Y.; et al. Epigenomic Diversity in a Global Collection of Arabidopsis Thaliana Accessions. Cell 2016, 166, 492–505. [Google Scholar] [CrossRef]
- Klepikova, A.V.; Kasianov, A.S.; Gerasimov, E.S.; Logacheva, M.D.; Penin, A.A. A High Resolution Map of the Arabidopsis Thaliana Developmental Transcriptome Based on RNA-Seq Profiling. Plant J. 2016, 88, 1058–1070. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.; Barthet, M.; Melino, V.; Smith, P.; Day, D.; Soole, K. Alterations in the Mitochondrial Alternative NAD(P)H Dehydrogenase NDB4 Lead to Changes in Mitochondrial Electron Transport Chain Composition, Plant Growth and Response to Oxidative Stress. Plant Cell Physiol. 2011, 52, 1222–1237. [Google Scholar] [CrossRef] [PubMed]
- Wallström, S.V.; Florez-Sarasa, I.; Araújo, W.L.; Aidemark, M.; Fernández-Fernández, M.; Fernie, A.R.; Ribas-Carbó, M.; Rasmusson, A.G. Suppression of the External Mitochondrial NADPH Dehydrogenase, NDB1, in Arabidopsis Thaliana Affects Central Metabolism and Vegetative Growth. Mol. Plant 2014, 7, 356–368. [Google Scholar] [CrossRef] [PubMed]
- Podgórska, A.; Ostaszewska-Bugajska, M.; Borysiuk, K.; Tarnowska, A.; Jakubiak, M.; Burian, M.; Rasmusson, A.G.; Szal, B. Suppression of External NADPH Dehydrogenase—NDB1 in Arabidopsis Thaliana Confers Improved Tolerance to Ammonium Toxicity via Efficient Glutathione/Redox Metabolism. Int. J. Mol. Sci. 2018, 19, 1412. [Google Scholar] [CrossRef] [PubMed]
- Giraud, E.; van Aken, O.; Ho, L.H.M.; Whelan, J. The Transcription Factor ABI4 Is a Regulator of Mitochondrial Retrograde Expression of ALTERNATIVE OXIDASE1a. Plant Physiol. 2009, 150, 1286–1296. [Google Scholar] [CrossRef] [PubMed]
- Atkin, O.K.; Macherel, D. The Crucial Role of Plant Mitochondria in Orchestrating Drought Tolerance. Ann. Bot. 2009, 103, 581–597. [Google Scholar] [CrossRef] [PubMed]
- Yamada, S.; Ozaki, H.; Noguchi, K. The Mitochondrial Respiratory Chain Maintains the Photosynthetic Electron Flow in Arabidopsis Thaliana Leaves under High-Light Stress. Plant Cell Physiol. 2019, 61, 283–295. [Google Scholar] [CrossRef]
- de Clercq, I.; Vermeirssen, V.; van Aken, O.; Vandepoele, K.; Murcha, M.W.; Law, S.R.; Inzé, A.; Ng, S.; Ivanova, A.; Rombaut, D.; et al. The Membrane-Bound NAC Transcription Factor ANAC013 Functions in Mitochondrial Retrograde Regulation of the Oxidative Stress Response in Arabidopsis. Plant Cell 2013, 25, 3472–3490. [Google Scholar] [CrossRef]
- van Aken, O.; Whelan, J. Comparison of Transcriptional Changes to Chloroplast and Mitochondrial Perturbations Reveals Common and Specific Responses in Arabidopsis. Front. Plant Sci. 2012, 3, 281. [Google Scholar] [CrossRef]
- Ng, S.; Ivanova, A.; Duncan, O.; Law, S.R.; van Aken, O.; de Clercq, I.; Wang, Y.; Carrie, C.; Xu, L.; Kmiec, B.; et al. A Membrane-Bound NAC Transcription Factor, ANAC017, Mediates Mitochondrial Retrograde Signaling in Arabidopsis. Plant Cell 2013, 25, 3450–3471. [Google Scholar] [CrossRef]
- Broda, M.; Khan, K.; O’Leary, B.; Pruzinska, A.; Pong Lee, C.; Harvey Millar, A.; van Aken, O. Increased Expression of ANAC017 Primes for Accelerated Senescence. Plant Physiol. 2021, 186, 2205–2221. [Google Scholar] [CrossRef] [PubMed]
- van Aken, O.; Ford, E.; Lister, R.; Huang, S.; Millar, A.H. Retrograde Signalling Caused by Heritable Mitochondrial Dysfunction Is Partially Mediated by ANAC017 and Improves Plant Performance. Plant J. 2016, 88, 542–558. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Li, L.; de Clercq, I.; Narsai, R.; Xu, Y.; Hartmann, A.; Claros, D.L.; Custovic, E.; Lewsey, M.G.; Whelan, J.; et al. ANAC017 Coordinates Organellar Functions and Stress Responses by Reprogramming Retrograde Signaling. Plant Physiol. 2019, 180, 634–653. [Google Scholar] [CrossRef]
- Kim, Y.S.; Sakuraba, Y.; Han, S.H.; Yoo, S.C.; Paek, N.C. Mutation of the Arabidopsis NAC016 Transcription Factor Delays Leaf Senescence. Plant Cell Physiol. 2013, 54, 1660–1672. [Google Scholar] [CrossRef]
- Kim, J.; Woo, H.R.R.; Nam, H.G.G. Toward Systems Understanding of Leaf Senescence: An Integrated Multi-Omics Perspective on Leaf Senescence Research. Mol. Plant 2016, 9, 813–825. [Google Scholar] [CrossRef]
- Kim, H.J.; Park, J.H.; Kim, J.; Kim, J.J.; Hong, S.; Kim, J.; Kim, J.H.; Woo, H.R.; Hyeon, C.; Lim, P.O.; et al. Time-Evolving Genetic Networks Reveal a Nac Troika That Negatively Regulates Leaf Senescence in Arabidopsis. Proc. Natl. Acad. Sci. USA 2018, 115, E4930–E4939. [Google Scholar] [CrossRef] [PubMed]
- van der Graaff, E.; Schwacke, R.; Schneider, A.; Desimone, M.; Flügge, U.I.; Kunze, R. Transcription Analysis of Arabidopsis Membrane Transporters and Hormone Pathways during Developmental and Induced Leaf Senescence. Plant Physiol. 2006, 141, 776–792. [Google Scholar] [CrossRef] [PubMed]
- Pascual, J.; Rahikainen, M.; Angeleri, M.; Alegre, S.; Gossens, R.; Shapiguzov, A.; Heinonen, A.; Trotta, A.; Durian, G.; Winter, Z.; et al. ACONITASE 3 Is Part of TheANAC017 Transcription Factor-Dependent Mitochondrial Dysfunction Response. Plant Physiol. 2021, 186, 1859–1877. [Google Scholar] [CrossRef]
- Holdsworth, M.J.; Vicente, J.; Sharma, G.; Abbas, M.; Zubrycka, A. The Plant N-Degron Pathways of Ubiquitin-Mediated Proteolysis. J. Integr. Plant Biol. 2020, 62, 70–89. [Google Scholar] [CrossRef]
- Gibbs, D.J.; MdIsa, N.; Movahedi, M.; Lozano-Juste, J.; Mendiondo, G.M.; Berckhan, S.; Marín-delaRosa, N.; VicenteConde, J.; SousaCorreia, C.; Pearce, S.P.; et al. Nitric Oxide Sensing in Plants Is Mediated by Proteolytic Control of Group VII ERF Transcription Factors. Mol. Cell 2014, 53, 369–379. [Google Scholar] [CrossRef]
- Licausi, F.; Kosmacz, M.; Weits, D.A.; Giuntoli, B.; Giorgi, F.M.; Voesenek, L.A.C.J.; Perata, P.; van Dongen, J.T. Oxygen Sensing in Plants Is Mediated by an N-End Rule Pathway for Protein Destabilization. Nature 2011, 479, 419–422. [Google Scholar] [CrossRef]
- Gibbs, D.J.; Conde, J.V.; Berckhan, S.; Prasad, G.; Mendiondo, G.M.; Holdsworth, M.J. Group VII Ethylene Response Factors Coordinate Oxygen and Nitric Oxide Signal Transduction and Stress Responses in Plants. Plant Physiol. 2015, 169, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Lamichhane, S.; Alpuerto, J.B.; Han, A.; Fukao, T. The Central Negative Regulator of Flooding Tolerance, the PROTEOLYSIS 6 Branch of the n-Degron Pathway, Adversely Modulates Salinity Tolerance in Arabidopsis. Plants 2020, 9, 1415. [Google Scholar] [CrossRef] [PubMed]
- Mendiondo, G.M.; Gibbs, D.J.; Szurman-Zubrzycka, M.; Korn, A.; Marquez, J.; Szarejko, I.; Maluszynski, M.; King, J.; Axcell, B.; Smart, K.; et al. Enhanced Waterlogging Tolerance in Barley by Manipulation of Expression of the N-End Rule Pathway E3 Ligase PROTEOLYSIS6. Plant Biotechnol. J. 2016, 14, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Vicente, J.; Mendiondo, G.M.; Movahedi, M.; Peirats-Llobet, M.; Juan, Y.-T.; Shen, Y.-Y.; Dambire, C.; Smart, K.; Rodriguez, P.L.; Charng, Y.-Y.; et al. The Cys-Arg/N-End Rule Pathway Is a General Sensor of Abiotic Stress in Flowering Plants. Curr. Biol. 2017, 27, 3183–3190. [Google Scholar] [CrossRef]
- Holman, T.J.; Jones, P.D.; Russell, L.; Medhurst, A.; Ubeda Tomá, S.D.S.; Talloji, P.; Marquez, J.; Schmuths, H.; Tung, S.-A.; Taylor, I.; et al. The N-End Rule Pathway Promotes Seed Germination and Establishment through Removal of ABA Sensitivity in Arabidopsis. Proc. Natl. Acad. Sci. USA 2009, 106, 4549–4554. [Google Scholar] [CrossRef]
- Labandera, A.M.; Tedds, H.M.; Bailey, M.; Sprigg, C.; Etherington, R.D.; Akintewe, O.; Kalleechurn, G.; Holdsworth, M.J.; Gibbs, D.J. The PRT6 N-Degron Pathway Restricts VERNALIZATION 2 to Endogenous Hypoxic Niches to Modulate Plant Development. New Phytol. 2021, 229, 126–139. [Google Scholar] [CrossRef]
- Barreto, P.; Arcuri, M.L.C.; Lima, R.P.M.; Marino, C.L.; Maia, I.G. Comprehensive In Silico Analysis and Transcriptional Profiles Highlight the Importance of Mitochondrial Dicarboxylate Carriers (DICs) on Hypoxia Response in Both Arabidopsis Thaliana and Eucalyptus Grandis. Plants 2022, 11, 181. [Google Scholar] [CrossRef]
- Wang, Y.; Selinski, J.; Mao, C.; Zhu, Y.; Berkowitz, O.; Whelan, J. Linking Mitochondrial and Chloroplast Retrograde Signalling in Plants. Philos. Trans. Royal Soc. 2020, 375, 20190410. [Google Scholar] [CrossRef]
- Li, L.; Wurtele, E.S. The QQS Orphan Gene of Arabidopsis Modulates Carbon and Nitrogen Allocation in Soybean. Plant Biotechnol. J. 2015, 13, 177–187. [Google Scholar] [CrossRef]
- Bai, Y.; Shen, Y.; Zhang, Z.; Jia, Q.; Xu, M.; Zhang, T.; Fang, H.; Yu, X.; Li, L.; Liu, D.; et al. A GPAT1 Mutation in Arabidopsis Enhances Plant Height but Impairs Seed Oil Biosynthesis. Int. J. Mol. Sci. 2021, 22, 785. [Google Scholar] [CrossRef] [PubMed]
- Nietzsche, M.; Schießl, I.; Börnke, F. The Complex Becomes More Complex: Protein-Protein Interactions of SnRK1 with DUF581 Family Proteins Provide a Framework for Cell- and Stimulus Type-Specific SnRK1 Signaling in Plants. Front. Plant Sci 2014, 5, 54. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.; Chandel, N.S.; Simon, M.C. Cellular Adaptation to Hypoxia through Hypoxia Inducible Factors and Beyond. Nat. Rev. Mol. Cell Biol. 2020, 21, 268–283. [Google Scholar] [CrossRef]
- Ratcliffe, P.J.; O’Rourke, J.F.; Maxwel, P.H.; Pugh, C.W. Oxygen Sensing, Hypoxia-Inducible Factor-1 and the Regulation of Mammalian Gene Expression. J. Exp. Biol. 1998, 201, 1153–1162. [Google Scholar] [CrossRef] [PubMed]
- Holdsworth, M.J.; Gibbs, D.J. Comparative Biology of Oxygen Sensing in Plants and Animals. Curr. Biol. 2020, 30, R362–R369. [Google Scholar] [CrossRef]
- Meléndez-Rodríguez, F.; Urrutia, A.A.; Lorendeau, D.; Rinaldi, G.; Roche, O.; Böğürcü-Seidel, N.; Ortega Muelas, M.; Mesa-Ciller, C.; Turiel, G.; Bouthelier, A.; et al. HIF1α Suppresses Tumor Cell Proliferation through Inhibition of Aspartate Biosynthesis. Cell Rep. 2019, 26, 2257–2265. [Google Scholar] [CrossRef]
- Wagner, S.; van Aken, O.; Elsässer, M.; Schwarzländer, M. Mitochondrial Energy Signaling and Its Role in the Low-Oxygen Stress Response of Plants. Plant Physiol. 2018, 176, 1156–1170. [Google Scholar] [CrossRef]
- Barreto, P.; Okura, V.; Pena, I.A.; Maia, R.; Maia, I.G.; Arruda, P. Overexpression of Mitochondrial Uncoupling Protein 1 (UCP1) Induces a Hypoxic Response in Nicotiana Tabacum Leaves. J. Exp. Bot. 2016, 67, 301–313. [Google Scholar] [CrossRef]
- Schwarzländer, M.; König, A.C.; Sweetlove, L.J.; Finkemeier, I. The Impact of Impaired Mitochondrial Function on Retrograde Signalling: A Meta-Analysis of Transcriptomic Responses. J. Exp. Bot. 2012, 63, 1735–1750. [Google Scholar] [CrossRef]
- Mustroph, A.; Lee, S.C.; Oosumi, T.; Zanetti, M.E.; Yang, H.; Ma, K.; Yaghoubi-Masihi, A.; Fukao, T.; Bailey-Serres, J. Cross-Kingdom Comparison of Transcriptomic Adjustments to Low-Oxygen Stress Highlights Conserved and Plant-Specific Responses. Plant Physiol. 2010, 152, 1484–1500. [Google Scholar] [CrossRef]
- Bui, L.T.; Shukla, V.; Giorgi, F.M.; Trivellini, A.; Perata, P.; Licausi, F.; Giuntoli, B. Differential Submergence Tolerance between Juvenile and Adult Arabidopsis Plants Involves the ANAC017 Transcription Factor. Plant J. 2020, 104, 979–994. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Liew, L.C.; Yin, L.; Lewsey, M.G.; Whelan, J.; Berkowitz, O. The Retrograde Signalling Regulator ANAC017 Recruits the MKK9-MPK3/6, Ethylene, and Auxin Signalling Pathways to Balance Mitochondrial Dysfunction with Growth. Plant Cell 2022. online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Hartman, S.; Liu, Z.; van Veen, H.; Vicente, J.; Reinen, E.; Martopawiro, S.; Zhang, H.; van Dongen, N.; Bosman, F.; Bassel, G.W.; et al. Ethylene-Mediated Nitric Oxide Depletion Pre-Adapts Plants to Hypoxia Stress. Nature Comm. 2019, 10, 4020. [Google Scholar] [CrossRef] [PubMed]
- Jurdak, R.; Launay-Avon, A.; Paysant-Le Roux, C.; Bailly, C. Retrograde Signalling from the Mitochondria to the Nucleus Translates the Positive Effect of Ethylene on Dormancy Breaking of Arabidopsis Thaliana Seeds. New Phytol. 2021, 229, 2192–2205. [Google Scholar] [CrossRef]
- Jeong, J.; Cohu, C.; Kerkeb, L.; Pilon, M.; Connolly, E.L.; Guerinot, M. lou Chloroplast Fe(III) Chelate Reductase Activity Is Essential for Seedling Viability under Iron Limiting Conditions. Proc. Natl. Acad. Sci. USA 2008, 105, 10619–10624. [Google Scholar] [CrossRef]
- Shapiguzov, A.; Vainonen, J.P.; Hunter, K.; Tossavainen, H.; Tiwari, A.; Järvi, S.; Hellman, M.; Aarabi, F.; Alseekh, S.; Wybouw, B.; et al. Arabidopsis RCD1 Coordinates Chloroplast and Mitochondrial Functions through Interaction with ANAC Transcription Factors. eLife 2019, 8, e43284. [Google Scholar] [CrossRef]
- Jaspers, P.; Blomster, T.; Brosché, M.; Salojärvi, J.; Ahlfors, R.; Vainonen, J.P.; Reddy, R.A.; Immink, R.; Angenent, G.; Turck, F.; et al. Unequally Redundant RCD1 and SRO1 Mediate Stress and Developmental Responses and Interact with Transcription Factors. Plant J. 2009, 60, 268–279. [Google Scholar] [CrossRef]
- Hiltscher, H.; Rudnik, R.; Shaikhali, J.; Heiber, I.; Mellenthin, M.; Meirelles Duarte, I.; Schuster, G.; Kahmann, U.; Baier, M. The Radical Induced Cell Death Protein 1 (RCD1) Supports Transcriptional Activation of Genes for Chloroplast Antioxidant Enzymes. Front. Plant Sci. 2014, 5, 475. [Google Scholar] [CrossRef]
- Brosché, M.; Blomster, T.; Salojärvi, J.; Cui, F.; Sipari, N.; Leppälä, J.; Lamminmäki, A.; Tomai, G.; Narayanasamy, S.; Reddy, R.A.; et al. Transcriptomics and Functional Genomics of ROS-Induced Cell Death Regulation by RADICAL-INDUCED CELL DEATH1. PLoS Genet. 2014, 10, e1004112. [Google Scholar] [CrossRef]
- Abbas, M.; Berckhan, S.; Rooney, D.J.; Gibbs, D.J.; Vicente Conde, J.; Sousa Correia, C.; Bassel, G.W.; Marín-De La Rosa, N.; León, J.; Alabadí, D.; et al. Oxygen Sensing Coordinates Photomorphogenesis to Facilitate Seedling Survival. Curr. Biol. 2015, 25, 1483–1488. [Google Scholar] [CrossRef]
- Abbas, M.; Sharma, G.; Dambire, C.; Marquez, J.; Alonso-Blanco, C.; Proaño, K.; Holdsworth, M.J. An Oxygen-Sensing Mechanism for Angiosperm Adaptation to Altitude. Nature 2022, 606, 565–569. [Google Scholar] [CrossRef] [PubMed]
- Araújo, W.L.; Nunes-Nesi, A.; Fernie, A.R. On the Role of Plant Mitochondrial Metabolism and Its Impact on Photosynthesis in Both Optimal and Sub-Optimal Growth Conditions. Photosynth. Res. 2014, 119, 141–156. [Google Scholar] [CrossRef] [PubMed]
- Nunes-Nesi, A.; Araújo, W.L.; Fernie, A.R. Targeting Mitochondrial Metabolism and Machinery as a Means to Enhance Photosynthesis. Plant Physiol. 2011, 155, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Batista-Silva, W.; da Fonseca-Pereira, P.; Martins, A.O.; Zsögön, A.; Nunes-Nesi, A.; Araújo, W.L. Engineering Improved Photosynthesis in the Era of Synthetic Biology. Plant Commun. 2020, 1, 100032. [Google Scholar] [CrossRef] [PubMed]
- Geigenberger, P.; Thormählen, I.; Daloso, D.M.; Fernie, A.R. The Unprecedented Versatility of the Plant Thioredoxin System. Trends Plant Sci. 2017, 22, 249–262. [Google Scholar] [CrossRef]
- Yoshida, K.; Noguchi, K.; Motohashi, K.; Hisabori, T. Systematic Exploration of Thioredoxin Target Proteins in Plant Mitochondria. Plant Cell Physiol. 2013, 54, 875–892. [Google Scholar] [CrossRef]
- Balmer, Y.; Vensel, W.H.; Tanaka, C.K.; Hurkman, W.J.; Gelhaye, E.; Rouhier, N.; Jacquot, J.-P.; Manieri, W.; Schü, P.; Droux, M.; et al. Thioredoxin Links Redox to the Regulation of Fundamental Processes of Plant Mitochondria. Proc. Natl. Acad. Sci. USA 2004, 101, 2642–2647. [Google Scholar] [CrossRef]
- Daloso, D.M.; Müller, K.; Obata, T.; Florian, A.; Tohge, T.; Bottcher, A.; Riondet, C.; Bariat, L.; Carrari, F.; Nunes-Nesi, A.; et al. Thioredoxin, a Master Regulator of the Tricarboxylic Acid Cycle in Plant Mitochondria. Proc. Natl. Acad. Sci. USA 2015, 112, E1392–E1400. [Google Scholar] [CrossRef]
- da Fonseca-Pereira, P.; Daloso, D.M.; Gago, J.; Nunes-Nesi, A.; Araújo, W.L. On the Role of the Plant Mitochondrial Thioredoxin System during Abiotic Stress. Plant Signal. Behav. 2019, 14, 1592536. [Google Scholar] [CrossRef]
- da Fonseca-Pereira, P.; Souza, P.V.L.; Hou, L.Y.; Schwab, S.; Geigenberger, P.; Nunes-Nesi, A.; Timm, S.; Fernie, A.R.; Thormählen, I.; Araújo, W.L.; et al. Thioredoxin H2 Contributes to the Redox Regulation of Mitochondrial Photorespiratory Metabolism. Plant Cell Environ. 2020, 43, 188–208. [Google Scholar] [CrossRef]
- Hou, L.Y.; Lehmann, M.; Geigenberger, P. Thioredoxin H2 and O1 Show Different Subcellular Localizations and Redox-Active Functions, and Are Extrachloroplastic Factors Influencing Photosynthetic Performance in Fluctuating Light. Antioxidants 2021, 10, 705. [Google Scholar] [CrossRef] [PubMed]
- Geigenberger, P.; Fernie, A.R. Metabolic Control of Redox and Redox Control of Metabolism in Plants. Antioxid. Redox Signal. 2014, 21, 1389–1421. [Google Scholar] [CrossRef] [PubMed]
- König, A.C.; Hartl, M.; Boersema, P.J.; Mann, M.; Finkemeier, I. The Mitochondrial Lysine Acetylome of Arabidopsis. Mitochondrion 2014, 19, 252–260. [Google Scholar] [CrossRef] [PubMed]
- Havelund, J.F.; Thelen, J.J.; Møller, I.M. Biochemistry, Proteomics, and Phosphoproteomics of Plant Mitochondria from Non-Photosynthetic Cells. Front. Plant Sci. 2013, 4, 51. [Google Scholar] [CrossRef] [PubMed]
- da Fonseca-Pereira, P.; Neri-Silva, R.; Cavalcanti, J.H.F.; Brito, D.S.; Weber, A.P.M.; Araújo, W.L.; Nunes-Nesi, A. Data-Mining Bioinformatics: Connecting Adenylate Transport and Metabolic Responses to Stress. Trends Plant Sci. 2018, 23, 961–974. [Google Scholar] [CrossRef]
- Nunes-Nesi, A.; Cavalcanti, J.H.F.; Fernie, A.R. Characterization of in Vivo Function(s) of Members of the Plant Mitochondrial Carrier Family. Biomolecules 2020, 10, 1226. [Google Scholar] [CrossRef]
- Pfaff, E.; Klingenberg, M.; Ritt, E.; Vogell, W. Correlation of unspecific permeable mitochondrial spaces with intermembrane spaces. Eur. J, Biochem 1968, 5, 222–232. [Google Scholar] [CrossRef]
- Benz, R.; Kottke, M.; Brdiczka, D. The Cationically Selective State of the Mitochondrial Outer Membrane Pore: A Study with Intact Mitochondria and Reconstituted Mitochondrial Porin. Biochim. Biophys. Acta 1990, 1022, 311–318. [Google Scholar] [CrossRef]
- Colombini, M. A Candidate for the Permeability Pathway of the Outer Mitochondrial Membrane. Nature 1979, 279, 643–645. [Google Scholar] [CrossRef]
- Lee, C.P.; Millar, A.H. The Plant Mitochondrial Transportome: Balancing Metabolic Demands with Energetic Constraints. Trends Plant Sci. 2016, 21, 662–676. [Google Scholar] [CrossRef]
- Fernie, A.R.; Cavalcanti, J.H.F.; Nunes-Nesi, A. Metabolic Roles of Plant Mitochondrial Carriers. Biomolecules 2020, 10, 1013. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.P.; Elsässer, M.; Fuchs, P.; Fenske, R.; Schwarzländer, M.; Millar, A.H. The Versatility of Plant Organic Acid Metabolism in Leaves Is Underpinned by Mitochondrial Malate-Citrate Exchange. Plant Cell 2021, 33, 3700–3720. [Google Scholar] [CrossRef] [PubMed]
- Steinbeck, J.; Fuchs, P.; Negroni, Y.L.; Elsässer, M.; Lichtenauer, S.; Stockdreher, Y.; Feitosa-Araujo, E.; Kroll, J.B.; Niemeier, J.O.; Humberg, C.; et al. In Vivo Nadh/Nad1 Biosensing Reveals the Dynamics of Cytosolic Redox Metabolism in Plants. Plant Cell 2020, 32, 3324–3345. [Google Scholar] [CrossRef] [PubMed]
- Schwarzländer, M.; Dick, T.P.; Meyer, A.J.; Morgan, B. Dissecting Redox Biology Using Fluorescent Protein Sensors. Antioxid. Redox Signal. 2016, 24, 680–712. [Google Scholar] [CrossRef] [PubMed]
- de Souza Chaves, I.; Feitosa-Araújo, E.; Florian, A.; Medeiros, D.B.; da Fonseca-Pereira, P.; Charton, L.; Heyneke, E.; Apfata, J.A.C.; Pires, M.V.; Mettler-Altmann, T.; et al. The Mitochondrial NAD+ Transporter (NDT1) Plays Important Roles in Cellular NAD+ Homeostasis in Arabidopsis Thaliana. Plant J. 2019, 100, 487–504. [Google Scholar] [CrossRef]
- Feitosa-Araujo, E.; da Fonseca-Pereira, P.; Knorr, L.S.; Schwarzländer, M.; Nunes-Nesi, A. NAD Meets ABA: Connecting Cellular Metabolism and Hormone Signaling. Trends Plant Sci. 2022, 27, 16–28. [Google Scholar] [CrossRef]
- Moseler, A.; Kruse, I.; Maclean, A.E.; Pedroletti, L.; Franceschetti, M.; Wagner, S.; Wehler, R.; Fischer-Schrader, K.; Poschet, G.; Wirtz, M.; et al. The function of glutaredoxin GRXS15 is required for lipoyl-dependent dehydrogenases in mitochondria. Plant Physiol. 2022, 186, 1507–1525. [Google Scholar] [CrossRef]
- Feitosa-Araujo, E.; de Souza Chaves, I.; Florian, A.; da Fonseca-Pereira, P.; Condori Apfata, J.A.; Heyneke, E.; Medeiros, D.B.; Pires, M.V.; Mettler-Altmann, T.; Neuhaus, H.E.; et al. Downregulation of a Mitochondrial NAD+Transporter (NDT2) Alters Seed Production and Germination in Arabidopsis. Plant Cell Physiol. 2020, 61, 897–908. [Google Scholar] [CrossRef]
- António, C.; Päpke, C.; Rocha, M.; Diab, H.; Limami, A.M.; Obata, T.; Fernie, A.R.; van Dongen, J.T. Regulation of Primary Metabolism in Response to Low Oxygen Availability as Revealed by Carbon and Nitrogen Isotope Redistribution. Plant Physiol. 2016, 170, 43–56. [Google Scholar] [CrossRef]
- Challabathula, D.; Analin, B.; Mohanan, A.; Bakka, K. Differential Modulation of Photosynthesis, ROS and Antioxidant Enzyme Activities in Stress-Sensitive and -Tolerant Rice Cultivars during Salinity and Drought upon Restriction of COX and AOX Pathways of Mitochondrial Oxidative Electron Transport. J. Plant Physiol. 2022, 268, 153583. [Google Scholar] [CrossRef]
- Del-Saz, N.F.; Ribas-Carbo, M.; McDonald, A.E.; Lambers, H.; Fernie, A.R.; Florez-Sarasa, I. An In Vivo Perspective of the Role(s) of the Alternative Oxidase Pathway. Trends Plant Sci. 2018, 23, 206–219. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Watanabe, C.K.A.; Miyagi, A.; Kawai-Yamada, M.; Terashima, I.; Noguchi, K. Mitochondrial Aox Supports Redox Balance of Photosynthetic Electron Transport, Primary Metabolite Balance, and Growth in Arabidopsis Thaliana under High Light. Int. J. Mol. Sci. 2019, 20, 3067. [Google Scholar] [CrossRef] [PubMed]
- Fiorani, F.; Umbach, A.L.; Siedow, J.N. The Alternative Oxidase of Plant Mitochondria Is Involved in the Acclimation of Shoot Growth at Low Temperature. A Study of Arabidopsis AOX1a Transgenic Plants. Plant Physiol. 2005, 139, 1795–1805. [Google Scholar] [CrossRef] [PubMed]
- Giraud, E.; Ho, L.H.M.; Clifton, R.; Carroll, A.; Estavillo, G.; Tan, Y.F.; Howell, K.A.; Ivanova, A.; Pogson, B.J.; Millar, A.H.; et al. The Absence of Alternative Oxidase1a in Arabidopsis Results in Acute Sensitivity to Combined Light and Drought Stress. Plant Physiol. 2008, 147, 595–610. [Google Scholar] [CrossRef] [PubMed]
- Gandin, A.; Duffes, C.; Day, D.A.; Cousins, A.B. The Absence of Alternative Oxidase AOX1A Results in Altered Response of Photosynthetic Carbon Assimilation to Increasing CO2 in Arabidopsis Thaliana. Plant Cell Physiol. 2012, 53, 1627–1637. [Google Scholar] [CrossRef] [PubMed]
- Demircan, N.; Cucun, G.; Uzilday, B. Mitochondrial Alternative Oxidase (AOX1a) Is Required for the Mitigation of Arsenic-Induced Oxidative Stress in Arabidopsis Thaliana. Plant Biotechnol. Rep. 2020, 14, 235–245. [Google Scholar] [CrossRef]
- Garmash, E.V.; Velegzhaninov, I.O.; Ermolina, K.V.; Rybak, A.V.; Malyshev, R.V. Altered Levels of AOX1a Expression Result in Changes in Metabolic Pathways in Arabidopsis Thaliana Plants Acclimated to Low Dose Rates of Ultraviolet B Radiation. Plant Science 2020, 291, 110332. [Google Scholar] [CrossRef]
- Vishwakarma, A.; Bashyam, L.; Senthilkumaran, B.; Scheibe, R.; Padmasree, K. Physiological Role of AOX1a in Photosynthesis and Maintenance of Cellular Redox Homeostasis under High Light in Arabidopsis Thaliana. Plant Physiol. Biochem. 2014, 81, 44–53. [Google Scholar] [CrossRef]
- Vishwakarma, A.; Tetali, S.D.; Selinski, J.; Scheibe, R.; Padmasree, K. Importance of the Alternative Oxidase (AOX) Pathway in Regulating Cellular Redox and ROS Homeostasis to Optimize Photosynthesis during Restriction of the Cytochrome Oxidase Pathway in Arabidopsis Thaliana. Ann. Bot. 2015, 116, 555–569. [Google Scholar] [CrossRef]
- Dinakar, C.; Vishwakarma, A.; Raghavendra, A.S.; Padmasree, K. Alternative Oxidase Pathway Optimizes Photosynthesis during Osmotic and Temperature Stress by Regulating Cellular Ros, Malate Valve and Antioxidative Systems. Front. Plant Sci. 2016, 7, 68. [Google Scholar] [CrossRef]
- Watanabe, C.K.A.; Yamori, W.; Takahashi, S.; Terashima, I.; Noguchi, K. Mitochondrial Alternative Pathway-Associated Photoprotection of Photosystem II Is Related to the Photorespiratory Pathway. Plant Cell Physiol. 2016, 57, 1426–1431. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.W.; Kahlen, K.; Stützel, H. Disentangling the Contributions of Osmotic and Ionic Effects of Salinity on Stomatal, Mesophyll, Biochemical and Light Limitations to Photosynthesis. Plant Cell Environ. 2015, 38, 1528–1542. [Google Scholar] [CrossRef] [PubMed]
- Savvides, A.; Ali, S.; Tester, M.; Fotopoulos, V. Chemical Priming of Plants Against Multiple Abiotic Stresses: Mission Possible? Trends Plant Sci. 2016, 21, 329–340. [Google Scholar] [CrossRef]
- Sako, K.; Futamura, Y.; Shimizu, T.; Matsui, A.; Hirano, H.; Kondoh, Y.; Muroi, M.; Aono, H.; Tanaka, M.; Honda, K.; et al. Inhibition of Mitochondrial Complex I by the Novel Compound FSL0260 Enhances High Salinity-Stress Tolerance in Arabidopsis Thaliana. Sci. Rep. 2020, 10, 8691. [Google Scholar] [CrossRef]
- Dissanayake, B.M.; Staudinger, C.; Munns, R.; Taylor, N.L.; Millar, A.H. Distinct Salinity-Induced Changes in Wheat Metabolic Machinery in Different Root Tissue Types. J. Proteom. 2022, 256, 104502. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Fan, Y.; Shabala, S.; Li, C.; Lv, C.; Guo, B.; Xu, R.; Zhou, M. Understanding Mechanisms of Salinity Tolerance in Barley by Proteomic and Biochemical Analysis of Near-Isogenic Lines. Int. J. Mol. Sci. 2020, 21, 1516. [Google Scholar] [CrossRef]
- Jiang, D.; Chen, J.; Zhang, Z.; Hou, X. Mitochondrial Transcription Termination Factor 27 Is Required for Salt Tolerance in Arabidopsis Thaliana. Int. J. Mol. Sci. 2021, 22, 1466. [Google Scholar] [CrossRef]
- Che-Othman, M.H.; Millar, A.H.; Taylor, N.L. Connecting Salt Stress Signalling Pathways with Salinity-Induced Changes in Mitochondrial Metabolic Processes in C3 Plants. Plant Cell Environ. 2017, 40, 2875–2905. [Google Scholar] [CrossRef]
- Che-Othman, M.H.; Jacoby, R.P.; Millar, A.H.; Taylor, N.L. Wheat Mitochondrial Respiration Shifts from the Tricarboxylic Acid Cycle to the GABA Shunt under Salt Stress. New Phytol. 2020, 225, 1166–1180. [Google Scholar] [CrossRef]
- Bandehagh, A.; Taylor, N.L. Can Alternative Metabolic Pathways and Shunts Overcome Salinity Induced Inhibition of Central Carbon Metabolism in Crops? Front. Plant Sci. 2020, 11, 1072. [Google Scholar] [CrossRef]
- Noguchi, K.; Yoshida, K. Interaction between Photosynthesis and Respiration in Illuminated Leaves. Mitochondrion 2008, 8, 87–99. [Google Scholar] [CrossRef] [PubMed]
- Flexas, J.; Bota, J.; Loreto, F.; Cornic, G.; Sharkey, T.D. Diffusive and Metabolic Limitations to Photosynthesis under Drought and Salinity in C3 Plants. Plant Biol. 2004, 6, 269–279. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, C.; Chaves, M.M. Photosynthesis and Drought: Can We Make Metabolic Connections from Available Data? J. Exp. Bot. 2011, 62, 869–882. [Google Scholar] [CrossRef] [PubMed]
- Lawlor, D.W.; Tezara, W. Causes of Decreased Photosynthetic Rate and Metabolic Capacity in Water-Deficient Leaf Cells: A Critical Evaluation of Mechanisms and Integration of Processes. Ann. Bot. 2009, 103, 561–579. [Google Scholar] [CrossRef]
- Cvetkovska, M.; Dahal, K.; Alber, N.A.; Jin, C.; Cheung, M.; Vanlerberghe, G.C. Knockdown of Mitochondrial Alternative Oxidase Induces the “stress State” of Signaling Molecule Pools in Nicotiana Tabacum, with Implications for Stomatal Function. New Phytol. 2014, 203, 449–461. [Google Scholar] [CrossRef] [PubMed]
- Dahal, K.; Martyn, G.D.; Vanlerberghe, G.C. Improved Photosynthetic Performance during Severe Drought in Nicotiana Tabacum Overexpressing a Nonenergy Conserving Respiratory Electron Sink. New Phytol. 2015, 208, 382–395. [Google Scholar] [CrossRef]
- da Fonseca-Pereira, P.; Daloso, D.M.; Gago, J.; de Oliveira Silva, F.M.; Condori-Apfata, J.A.; Florez-Sarasa, I.; Tohge, T.; Reichheld, J.P.; Nunes-Nesi, A.; Fernie, A.R.; et al. The Mitochondrial Thioredoxin System Contributes to the Metabolic Responses under Drought Episodes in Arabidopsis. Plant Cell Physiol. 2019, 60, 213–229. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive Oxygen Species and Antioxidant Machinery in Abiotic Stress Tolerance in Crop Plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Rajput, V.D.; Harish; Singh, R.K.; Verma, K.K.; Sharma, L.; Quiroz-Figueroa, F.R.; Meena, M.; Gour, V.S.; Minkina, T.; Sushkova, S.; et al. Recent Developments in Enzymatic Antioxidant Defence Mechanism in Plants with Special Reference to Abiotic Stress. Biology 2021, 10, 267. [Google Scholar] [CrossRef]
- Tounsi, S.; Jemli, S.; Feki, K.; Brini, F.; Najib Saïdi, M. Superoxide Dismutase (SOD) Family in Durum Wheat: Promising Candidates for Improving Crop Resilience. Protoplasma, 2022; online ahead of print. 2022. [Google Scholar] [CrossRef]
- Block, A.; Guo, M.; Li, G.; Elowsky, C.; Clemente, T.E.; Alfano, J.R. The Pseudomonas Syringae Type III Effector HopG1 Targets Mitochondria, Alters Plant Development and Suppresses Plant Innate Immunity. Cell. Microbiol. 2010, 12, 318–330. [Google Scholar] [CrossRef] [PubMed]
- An, L.; Zhang, S.; Guo, P.; Song, L.; Xie, C.; Guo, H.; Fang, R.; Jia, Y. RIR1 Represses Plant Immunity by Interacting with Mitochondrial Complex I Subunit in Rice. Mol. Plant Path. 2022, 23, 92–103. [Google Scholar] [CrossRef] [PubMed]
- López Sánchez, A.; Hernández Luelmo, S.; Izquierdo, Y.; López, B.; Cascón, T.; Castresana, C. Mitochondrial Stress Induces Plant Resistance Through Chromatin Changes. Front. Plant Sci. 2021, 12, 704964. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Liang, Z.; Zhao, J.; Wang, P.; Ma, W.; Mai, K.K.; Fernandez Andrade, J.A.; Zeng, Y.; Grujic, N.; Jiang, L.; et al. Friendly Mediates Membrane Depolarization-Induced Mitophagy in Arabidopsis. Curr. Biol. 2021, 31, 1931–1944. [Google Scholar] [CrossRef]
- Liu, Q.; Galli, M.; Liu, X.; Federici, S.; Buck, A.; Cody, J.; Labra, M.; Gallavotti, A. NEEDLE1 Encodes a Mitochondria Localized ATP-Dependent Metalloprotease Required for Thermotolerant Maize Growth. Proc. Natl. Acad. Sci. USA 2019, 116, 19736–19742. [Google Scholar] [CrossRef]
- Tivendale, N.D.; Belt, K.; Berkowitz, O.; Whelan, J.; Millar, A.H.; Huang, S. Knockdown of Succinate Dehydrogenase Assembly Factor 2 Induces Reactive Oxygen Species-Mediated Auxin Hypersensitivity Causing PH-Dependent Root Elongation. Plant Cell Physiol. 2021, 62, 1185–1198. [Google Scholar] [CrossRef]
- Qi, W.; Lu, L.; Huang, S.; Song, R. Maize Dek44 Encodes Mitochondrial Ribosomal Protein L9 and Is Required for Seed Development. Plant Physiol. 2019, 180, 2106–2119. [Google Scholar] [CrossRef]
- Xiu, Z.; Peng, L.; Wang, Y.; Yang, H.; Sun, F.; Wang, X.; Cao, S.K.; Jiang, R.; Wang, L.; Chen, B.Y.; et al. Empty Pericarp24 and Empty Pericarp25 Are Required for the Splicing of Mitochondrial Introns, Complex I Assembly, and Seed Development in Maize. Front. Plant Sci. 2020, 11, 608550. [Google Scholar] [CrossRef]
- Hu, Z.; Vanderhaeghen, R.; Cools, T.; Wang, Y.; de Clercq, I.; Leroux, O.; Nguyen, L.; Belt, K.; Millar, H.A.; Audenaert, D.; et al. Mitochondrial Defects Confer Tolerance against Cellulose Deficiency. Plant Cell 2016, 28, 2276–2290. [Google Scholar] [CrossRef]
- Gao, M.; Zhao, H.; Zheng, L.; Zhang, L.; Peng, Y.; Ma, W.; Tian, R.; Yuan, Y.; Ma, F.; Li, M.; et al. Overexpression of Apple Ma12, a Mitochondrial Pyrophosphatase Pump Gene, Leads to Malic Acid Accumulation and the Upregulation of Malate Dehydrogenase in Tomato and Apple Calli. Hort. Res. 2022, 9, uhab053. [Google Scholar] [CrossRef]
- Selinski, J.; Hartmann, A.; Deckers-Hebestreit, G.; Day, D.A.; Whelan, J.; Scheibe, R. Alternative Oxidase Isoforms Are Differentially Activated by Tricarboxylic Acid Cycle Intermediates. Plant Physiol. 2018, 176, 1423–1432. [Google Scholar] [CrossRef] [PubMed]
- Mohanapriya, G.; Bharadwaj, R.; Noceda, C.; Costa, J.H.; Kumar, S.R.; Sathishkumar, R.; Thiers, K.L.L.; Santos Macedo, E.; Silva, S.; Annicchiarico, P.; et al. Alternative Oxidase (AOX) Senses Stress Levels to Coordinate Auxin-Induced Reprogramming From Seed Germination to Somatic Embryogenesis—A Role Relevant for Seed Vigor Prediction and Plant Robustness. Front. Plant Sci. 2019, 10, 1134. [Google Scholar] [CrossRef]
- Huther, C.M.; Ramm, A.; Rombaldi, C.V.; Bacarin, M.A. Physiological Response to Heat Stress of Tomato “Micro-Tom” Plants Expressing High and Low Levels of Mitochondrial SHSP23.6 Protein. Plant Growth Regul. 2013, 70, 175–185. [Google Scholar] [CrossRef]
- Hüther, C.M.; Martinazzo, E.G.; Rombaldi, C.V.; Bacarin, M.A. Effects of flooding stress in ‘Micro-Tom’ tomato plants transformed with different levels of mitochondrial sHSP23.6. Braz. J. Biol. 2017, 77, 43–51. [Google Scholar] [CrossRef]
- Reissig, G.N.; Posso, D.A.; Borella, J.; da Silveira, R.V.D.; Rombaldi, C.V.; Bacarin, M.A. High MT-SHSP23.6 Expression Increases Antioxidant System in ‘Micro-Tom’ Tomato Fruits during Post-Harvest Hypoxia. Sci. Hortic. 2018, 242, 127–136. [Google Scholar] [CrossRef]
- Niemeyer Reissig, G.; Posso, D.A.; Borella, J.; Vieira Dutra da Silveira, R.; Rombaldi, C.V.; Bacarin, M.A. High MT-SHSP23.6 Expression and Moderate Water Deficit Influence the Antioxidant System in “Micro-Tom” Tomato Fruit under Hypoxia. Fruits 2020, 75, 55–70. [Google Scholar] [CrossRef]
- Hüther, C.M.; Borella, J.; Martinazzo, E.G.; Dalberto, D.S.; de Farias, M.E.; Posso, D.A.; Rombaldi, C.V.; Bacarin, M.A. Mitochondrial Sense SHSP23.6 Protein Keeps Photosynthetic Electron Transport during Drought. Theor. Exp. Plant Physiol. 2022, 34, 95–108. [Google Scholar] [CrossRef]
- Muthusamy, S.K.; Dalal, M.; Chinnusamy, V.; Bansal, K.C. Genome-Wide Identification and Analysis of Biotic and Abiotic Stress Regulation of Small Heat Shock Protein (HSP20) Family Genes in Bread Wheat. J. Plant Physiol. 2017, 211, 100–113. [Google Scholar] [CrossRef]
- Yan, K.; Bian, T.; He, W.; Han, G.; Lv, M.; Guo, M.; Lu, M. Root Abscisic Acid Contributes to Defending Photoinibition in Jerusalem Artichoke (Helianthus Tuberosus L.) under Salt Stress. Int. J. Mol. Sci. 2018, 19, 3934. [Google Scholar] [CrossRef]
- Zhang, D.; Jiao, X.; Du, Q.; Song, X.; Li, J. Reducing the Excessive Evaporative Demand Improved Photosynthesis Capacity at Low Costs of Irrigation via Regulating Water Driving Force and Moderating Plant Water Stress of Two Tomato Cultivars. Agric. Water Manag. 2018, 199, 22–33. [Google Scholar] [CrossRef]
- Sinclair, T.R.; Rufty, T.W.; Lewis, R.S. Increasing Photosynthesis: Unlikely Solution For World Food Problem. Trends Plant Sci. 2019, 24, 1032–1039. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.L.; Voon, C.P.; Guan, X.; Yang, Y.; Gardeström, P.; Lim, B.L. In Planta Study of Photosynthesis and Photorespiration Using NADPH and NADH/NAD+ Fluorescent Protein Sensors. Nature Commun. 2020, 11, 3238. [Google Scholar] [CrossRef] [PubMed]
- Timm, S.; Hagemann, M. Photorespiration-How Is It Regulated and How Does It Regulate Overall Plant Metabolism? J. Exp. Bot. 2020, 71, 3955–3965. [Google Scholar] [CrossRef] [PubMed]
- Timm, S.; Nunes-Nesi, A.; Florian, A.; Eisenhut, M.; Morgenthal, K.; Wirtz, M.; Hell, R.; Weckwerth, W.; Hagemann, M.; Fernie, A.R.; et al. Metabolite Profiling in Arabidopsis Thaliana with Moderately Impaired Photorespiration Reveals Novel Metabolic Links and Compensatory Mechanisms of Photorespiration. Metabolites 2021, 11, 391. [Google Scholar] [CrossRef] [PubMed]
- Timm, S.; Wittmiß, M.; Gamlien, S.; Ewald, R.; Florian, A.; Frank, M.; Wirtz, M.; Hell, R.; Fernie, A.R.; Bauwe, H. Mitochondrial Dihydrolipoyl Dehydrogenase Activity Shapes Photosynthesis and Photorespiration of Arabidopsis Thaliana. Plant Cell 2015, 27, 1968–1984. [Google Scholar] [CrossRef]
- Timm, S.; Florian, A.; Arrivault, S.; Stitt, M.; Fernie, A.R.; Bauwe, H. Glycine Decarboxylase Controls Photosynthesis and Plant Growth. FEBS Lett. 2012, 586, 3692–3697. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barreto, P.; Koltun, A.; Nonato, J.; Yassitepe, J.; Maia, I.d.G.; Arruda, P. Metabolism and Signaling of Plant Mitochondria in Adaptation to Environmental Stresses. Int. J. Mol. Sci. 2022, 23, 11176. https://doi.org/10.3390/ijms231911176
Barreto P, Koltun A, Nonato J, Yassitepe J, Maia IdG, Arruda P. Metabolism and Signaling of Plant Mitochondria in Adaptation to Environmental Stresses. International Journal of Molecular Sciences. 2022; 23(19):11176. https://doi.org/10.3390/ijms231911176
Chicago/Turabian StyleBarreto, Pedro, Alessandra Koltun, Juliana Nonato, Juliana Yassitepe, Ivan de Godoy Maia, and Paulo Arruda. 2022. "Metabolism and Signaling of Plant Mitochondria in Adaptation to Environmental Stresses" International Journal of Molecular Sciences 23, no. 19: 11176. https://doi.org/10.3390/ijms231911176
APA StyleBarreto, P., Koltun, A., Nonato, J., Yassitepe, J., Maia, I. d. G., & Arruda, P. (2022). Metabolism and Signaling of Plant Mitochondria in Adaptation to Environmental Stresses. International Journal of Molecular Sciences, 23(19), 11176. https://doi.org/10.3390/ijms231911176