

Viromes of 15 Pepper (Capsicum annuum L.) Cultivars



Abstract
:1. Introduction
2. Results
2.1. Collection of Leaf Samples from 15 Different Pepper Cultivars
2.2. Viral Disease Symptoms in Different Pepper Cultivars
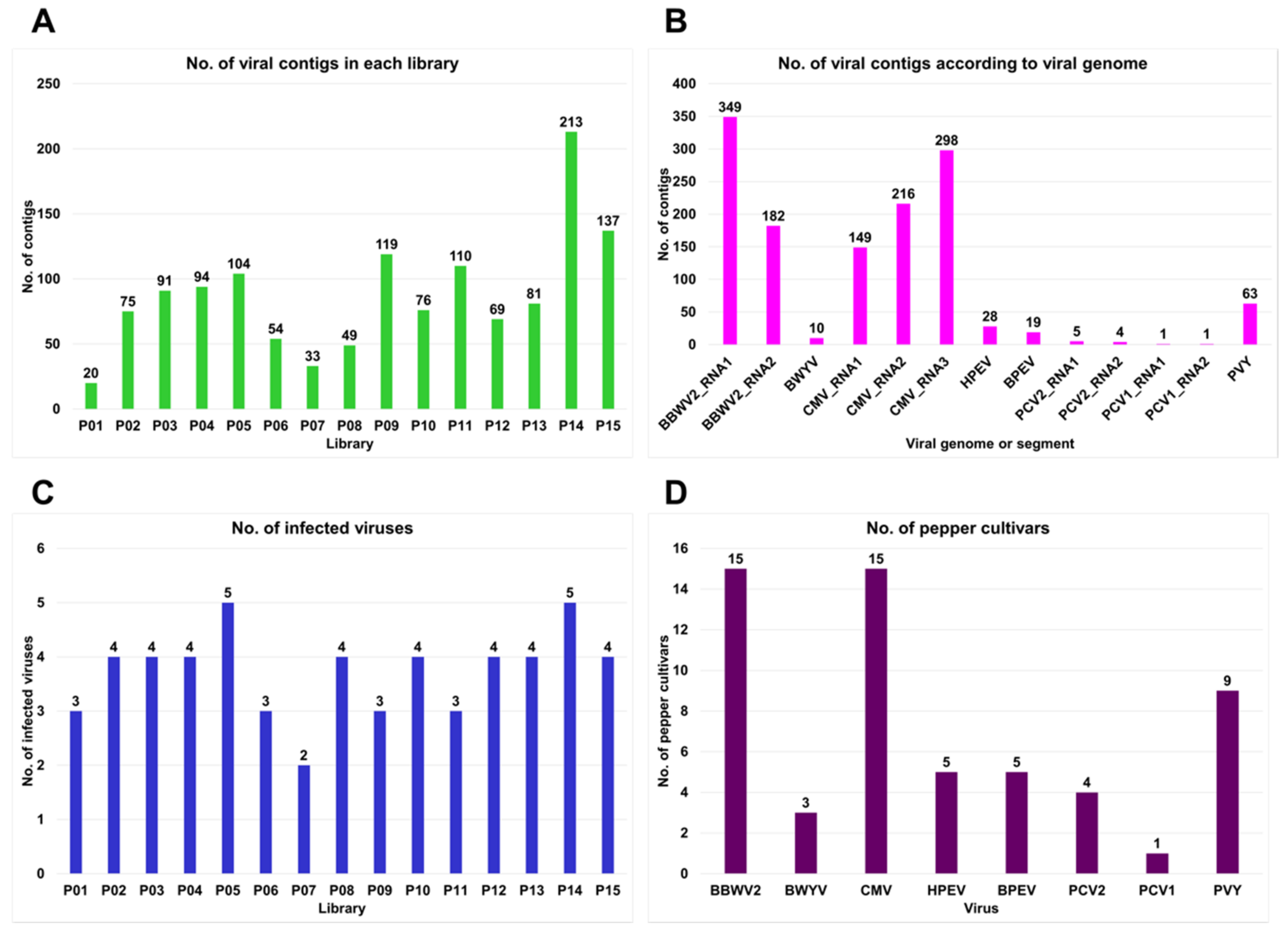
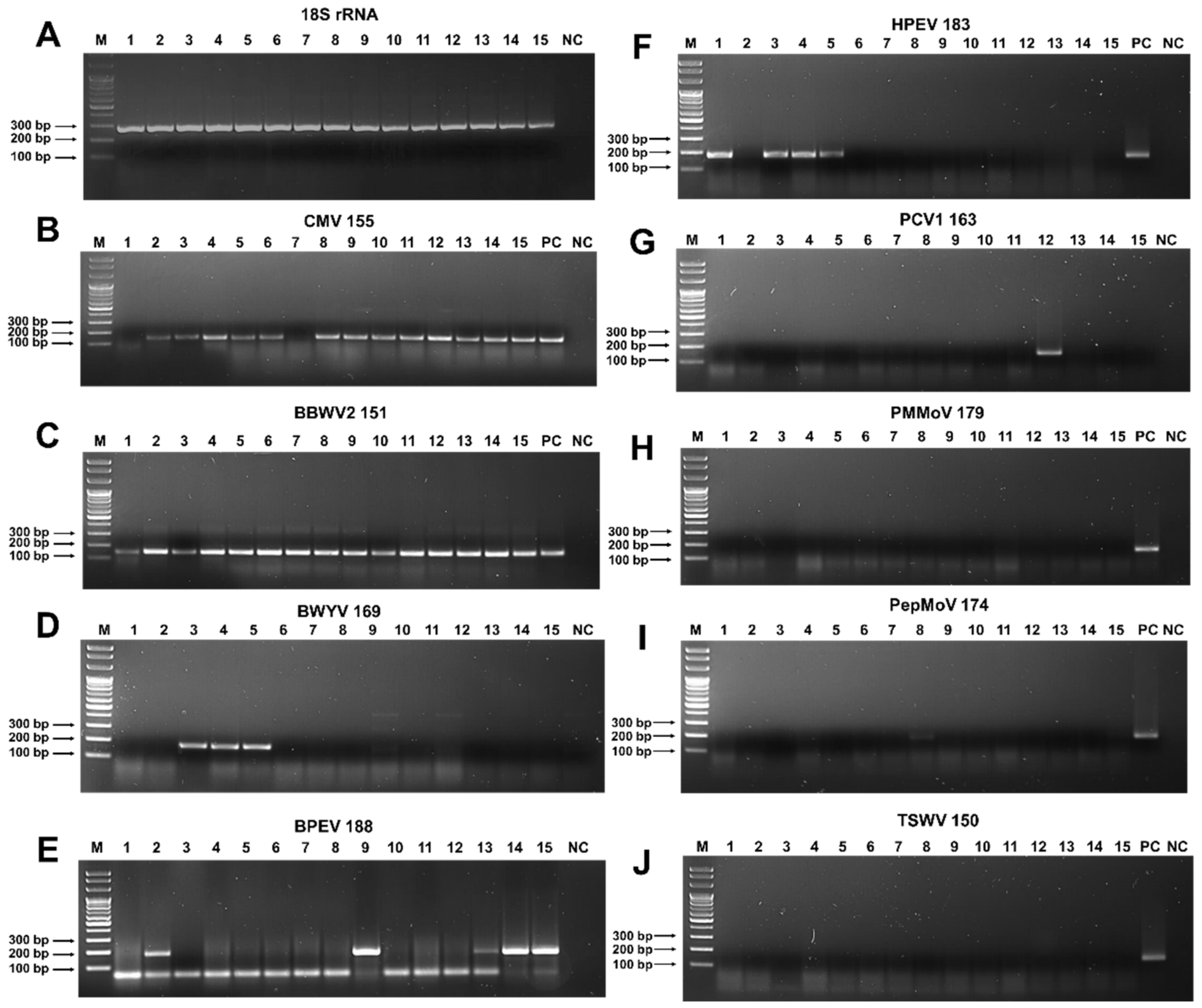
2.3. Identification of Virus-Associated Contigs from 15 Different Pepper Cultivars
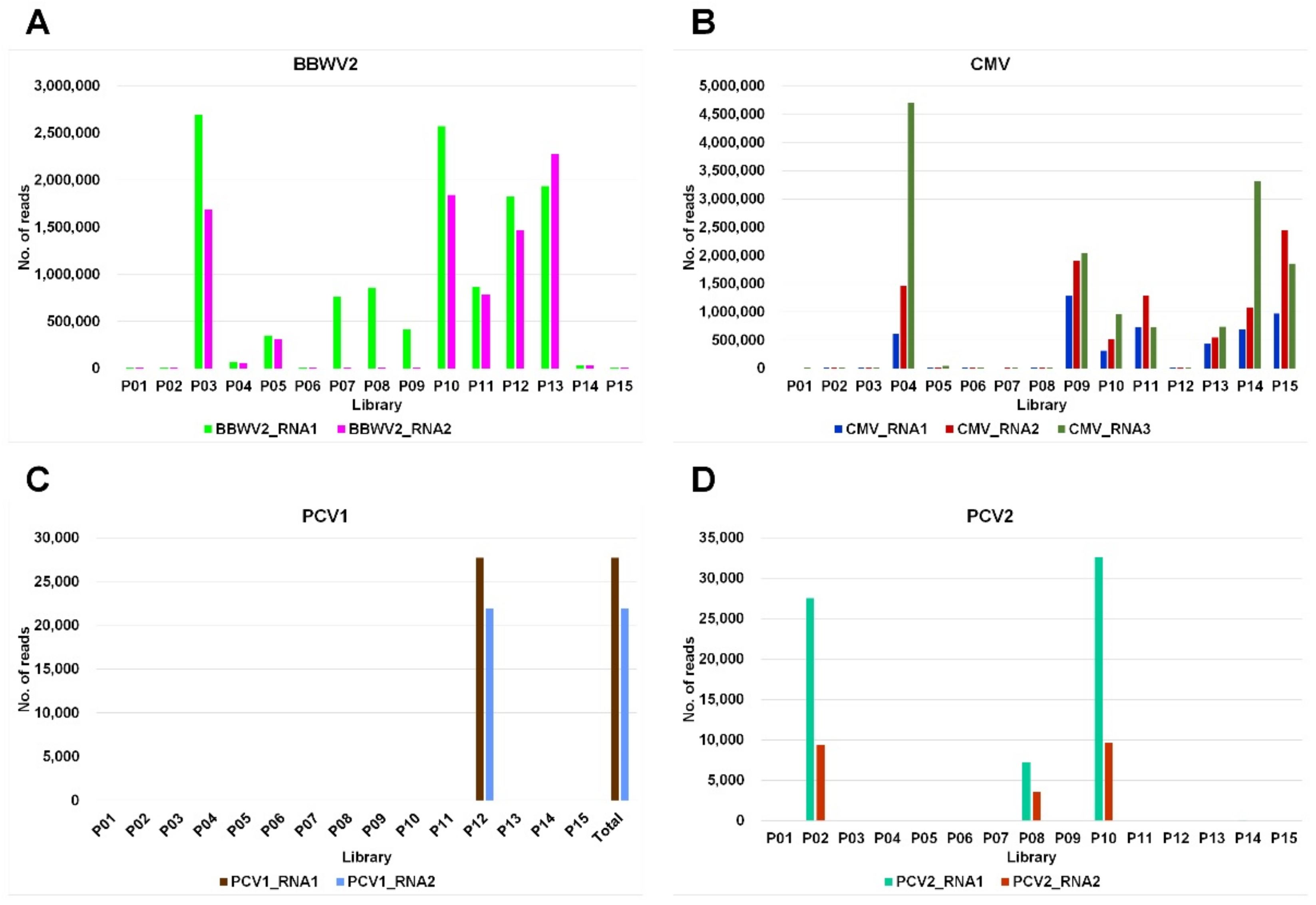
2.4. Viral Genome Assembly
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Viral Disease Symptoms
4.2. Total RNA Extraction and Library Preparation
4.3. RNA Sequencing and De Novo Transcriptome Assembly
4.4. Identification of Viral Contigs and Calculation of Viral Reads
4.5. Assembly of Viral Genome and Virus Genome Annotation
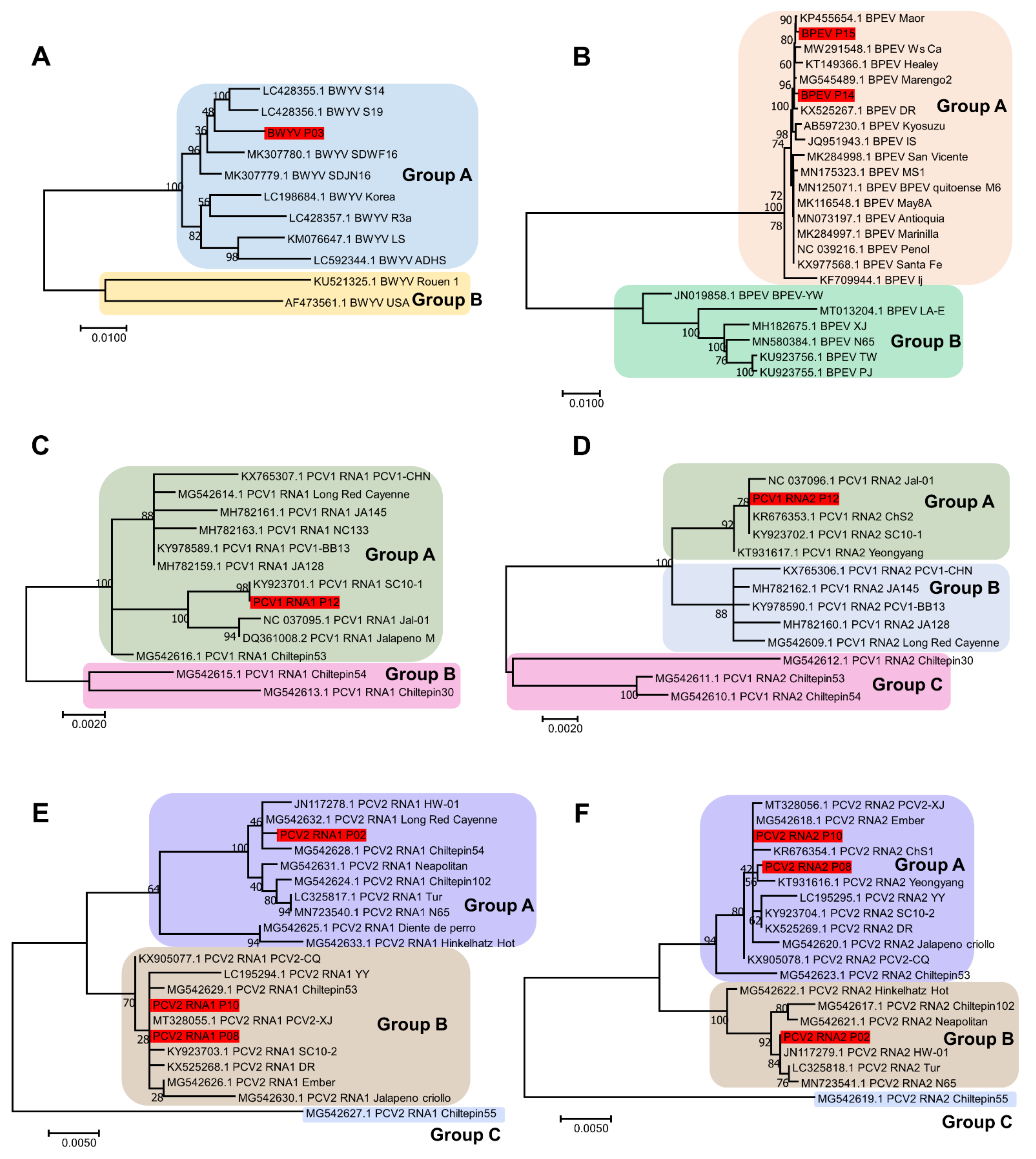
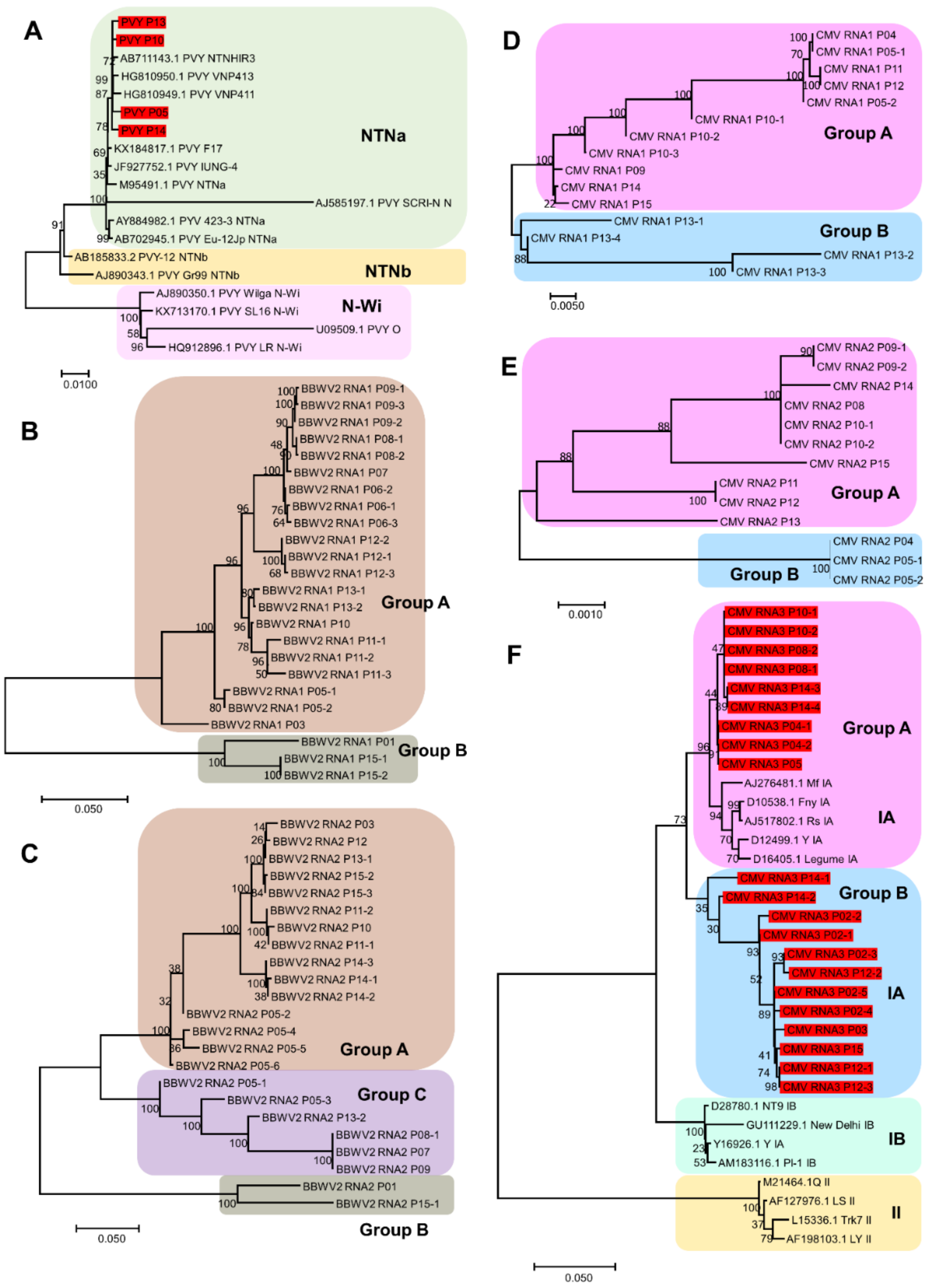
4.6. Construction of Phylogenetic Trees
4.7. Designing of Primers to Detect Viruses Infecting Pepper Plants via RT-PCR
4.8. RT-PCR and Agarose Gel Electrophoresis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Basu, S.K.; De, A.K. Capsicum: Historical and botanical perspectives. In Capsicum; CRC Press: Boca Raton, FL, USA, 2003; pp. 21–35. [Google Scholar]
- Olatunji, T.L.; Afolayan, A.J. The suitability of chili pepper (Capsicum annuum L.) for alleviating human micronutrient dietary deficiencies: A review. Food Sci. Nutr. 2018, 6, 2239–2251. [Google Scholar] [CrossRef] [PubMed]
- Kraft, K.H.; Brown, C.H.; Nabhan, G.P.; Luedeling, E.; Luna Ruiz, J.d.J.; Coppens d’Eeckenbrugge, G.; Hijmans, R.J.; Gepts, P. Multiple lines of evidence for the origin of domesticated chili pepper, Capsicum annuum, in Mexico. Proc. Natl. Acad. Sci. USA 2014, 111, 6165–6170. [Google Scholar] [CrossRef] [PubMed]
- Kwon, D.Y.; Jang, D.-J.; Yang, H.J.; Chung, K.R. History of Korean gochu, gochujang, and kimchi. J. Ethn. Foods 2014, 1, 3–7. [Google Scholar] [CrossRef]
- Jo, Y.; Choi, H.; Kim, S.-M.; Kim, S.-L.; Lee, B.C.; Cho, W.K. The pepper virome: Natural co-infection of diverse viruses and their quasispecies. BMC Genom. 2017, 18, 453. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.; Jo, Y.; Cho, W.K.; Yu, J.; Tran, P.-T.; Salaipeth, L.; Kwak, H.-R.; Choi, H.-S.; Kim, K.-H. Identification of viruses and viroids infecting tomato and pepper plants in vietnam by metatranscriptomics. Int. J. Mol. Sci. 2020, 21, 7565. [Google Scholar] [CrossRef] [PubMed]
- Choi, G.-S.; Kim, J.-H.; Lee, D.-H.; Kim, J.-S.; Ryu, K.-H. Occurrence and distribution of viruses infecting pepper in Korea. Plant Pathol. J. 2005, 21, 258–261. [Google Scholar] [CrossRef]
- Jo, Y.; Back, C.-G.; Kim, K.-H.; Chu, H.; Lee, J.H.; Moh, S.H.; Cho, W.K. Comparative Study of Metagenomics and Metatranscriptomics to Reveal Microbiomes in Overwintering Pepper Fruits. Int. J. Mol. Sci. 2021, 22, 6202. [Google Scholar] [CrossRef] [PubMed]
- Green, K.J.; Brown, C.J.; Gray, S.M.; Karasev, A.V. Phylogenetic study of recombinant strains of Potato virus Y. Virology 2017, 507, 40–52. [Google Scholar] [CrossRef]
- Glasa, M.; Hančinský, R.; Šoltys, K.; Predajňa, L.; Tomašechová, J.; Hauptvogel, P.; Mrkvová, M.; Mihálik, D.; Candresse, T. Molecular characterization of Potato Virus Y (PVY) using high-throughput sequencing: Constraints on full genome reconstructions imposed by mixed infection involving recombinant PVY strains. Plants 2021, 10, 753. [Google Scholar] [CrossRef]
- Mrkvová, M.; Hančinský, R.; Predajňa, L.; Alaxin, P.; Achs, A.; Tomašechová, J.; Šoltys, K.; Mihálik, D.; Olmos, A.; Ruiz-García, A.B.; et al. High-Throughput Sequencing Discloses the Cucumber Mosaic Virus (CMV) Diversity in Slovakia and Reveals New Hosts of CMV from the Papaveraceae Family. Plants 2022, 11, 1665. [Google Scholar] [CrossRef]
- Shen, L.; Liu, Z.; Yang, S.; Yang, T.; Liang, J.; Wen, J.; Liu, Y.; Li, J.; Shi, L.; Tang, Q.; et al. Pepper CabZIP63 acts as a positive regulator during Ralstonia solanacearum or high temperature–high humidity challenge in a positive feedback loop with CaWRKY40. J. Exp. Bot. 2016, 67, 2439–2451. [Google Scholar] [CrossRef]
- Kwak, H.-R.; Kim, M.-K.; Nam, M.; Kim, J.-S.; Kim, K.-H.; Cha, B.; Choi, H.-S. Genetic compositions of Broad bean wilt virus 2 infecting red pepper in Korea. Plant Pathol. J. 2013, 29, 274–284. [Google Scholar] [CrossRef]
- Svoboda, J.; Leisova-Svobodova, L. First Report of Broad bean wilt virus-2 in Pepper in the Czech Republic. Plant Dis. 2013, 97, 1261. [Google Scholar] [CrossRef]
- Lim, S.; Kim, K.H.; Zhao, F.; Yoo, R.H.; Igori, D.; Lee, S.-H.; Moon, J.S. Complete genome sequence of a novel endornavirus isolated from hot pepper. Arch. Virol. 2015, 160, 3153–3156. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, S.; Tan, R.; Zhang, D.; Zhang, S.; Liu, Y. First report of hot pepper endornavirus infecting pepper in China. J. Plant Pathol. 2020, 102, 229. [Google Scholar] [CrossRef]
- Jo, Y.; Choi, H.; Yoon, J.-Y.; Choi, S.-K.; Cho, W.K. In silico identification of Bell pepper endornavirus from pepper transcriptomes and their phylogenetic and recombination analyses. Gene 2016, 575, 712–717. [Google Scholar] [CrossRef]
- Valverde, R.A.; Khalifa, M.E.; Okada, R.; Fukuhara, T.; Sabanadzovic, S. ICTV virus taxonomy profile: Endornaviridae. J. Gen. Virol. 2019, 100, 1204–1205. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.-S.; Ko, S.-J.; Kim, M.-K.; Park, J.-W.; Lee, S.-H.; Kim, K.-H.; Were, H.-K.; Chois, J.-K.; Takanami, Y. Characteristics of Potato virus Y isolated from paprika in Korea. Plant Pathol. J. 2005, 21, 349–354. [Google Scholar] [CrossRef]
- Ali, A.; Kobayashi, M. Seed transmission of Cucumber mosaic virus in pepper. J. Virol. Methods 2010, 163, 234–237. [Google Scholar] [CrossRef]
- Andino, R.; Domingo, E. Viral quasispecies. Virology 2015, 479–480, 46–51. [Google Scholar] [CrossRef] [Green Version]
- Park, S.-K.; Park, S.-H.; Yoon, J.-Y.; Park, J.-K.; Ryu, K.-H. A Subpopulation of RNA3 of Cucumber mosaic virus Quasispecies. Plant Pathol. J. 2003, 19, 210–216. [Google Scholar] [CrossRef]
- Schneider, W.L.; Roossinck, M.J. Genetic diversity in RNA virus quasispecies is controlled by host-virus interactions. J. Virol. 2001, 75, 6566–6571. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Yu, C.; Yin, Y.; Gao, S.; Wang, F.; Jiao, C.; Yao, M. Pepper crop improvement against cucumber mosaic virus (CMV): A review. Front. Plant Sci. 2020, 11, 598798. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and sensitive protein alignment using DIAMOND. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef]
- Huson, D.H.; Beier, S.; Flade, I.; Górska, A.; El-Hadidi, M.; Mitra, S.; Ruscheweyh, H.-J.; Tappu, R. MEGAN community edition-interactive exploration and analysis of large-scale microbiome sequencing data. PLoS Comput. Biol. 2016, 12, e1004957. [Google Scholar] [CrossRef]
- Wheeler, D.L.; Chappey, C.; Lash, A.E.; Leipe, D.D.; Madden, T.L.; Schuler, G.D.; Tatusova, T.A.; Rapp, B.A. Database resources of the national center for biotechnology information. Nucleic Acids Res. 2000, 28, 10–14. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. trimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Higgins, M.; Ravenhall, M.; Ward, D.; Phelan, J.; Ibrahim, A.; Forrest, M.S.; Clark, T.G.; Campino, S. PrimedRPA: Primer design for recombinase polymerase amplification assays. Bioinformatics 2019, 35, 682–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index | Name | Category | Spicy | Region |
|---|---|---|---|---|
| P01 | PR-9988 | Red hot pepper | Hot | Hampyeong |
| P02 | Mini paprika | Sweet paprika | Sweet | Ilsan |
| P03 | TS-Monster | Red hot pepper | Hot | Hampyeong |
| P04 | Boradori | Sweet pepper | Sweet | Ilsan |
| P05 | Miin | Sweet pepper | Sweet | Ilsan |
| P06 | Chegangtan | Red hot pepper | Hot | Pusan |
| P07 | Dangjo | Sweet pepper | Sweet | Ilsan |
| P08 | Ground cherry (Wrinkly) | Red hot pepper | Hot | Uljin |
| P09 | Yellow paprika | Sweet paprika | Sweet | Anyang |
| P10 | Vitamin | Sweet pepper | Sweet | Hampyeong |
| P11 | Oi (Cucumber) | Sweet pepper | Sweet | Hampyeong |
| P12 | Caltan | Red hot pepper | Hot | Anyang |
| P13 | Cheongyang | Red hot pepper | Hot | Anyang |
| P14 | Red paprika | Sweet paprika | Sweet | Anyang |
| P15 | Green paprika | Sweet paprika | Sweet | Hampyeong |
| Viral Reference and Accession No. | Name | Position | Sequences | Size |
|---|---|---|---|---|
| 18S rRNA of pepper | Ca18SrRNA-F1 | CCGGTCCGCCTATGGTGTGCACCGGTCGTC | 285 | |
| EF564281 | Ca18SrRNA-R1 | GCAGTTGTTCGTCTTTCATAAATCCAAGAA | ||
| Cucumber mosaic virus RNA3 | CMVs3-1507F1 | 1507–1536 | TATTATGGTAAAAGGTTGTTGCTACCTGAT | 155 |
| D28780.1 | CMVs3-1661R1 | 1661–1632 | ACGGATAAGTCCGAGGAGGCAGAAACTTTA | |
| Pepper mild mottle virus | PMMoV-5881F1 | 5881–5910 | CTGGTTTCAAAGTTTTCCGATATAATGCCG | 179 |
| MN496154.1 | PMMoV-6059R1 | 6059–6030 | ACTGGCCCTAATGGCCACCGTCGCATCATC | |
| Broad bean wilt virus 2 RNA2 | BBWVs2-2463F1 | 2463–2492 | TGCAAATTCTTGACTCGCTGCCCCATATCG | 151 |
| JX183230.1 | BBWVs2-2613R1 | 2613–2584 | GAAACATCATCCCCTCTTGCCGAGTCCAAA | |
| Tomato spotted wilt virus S segment | TSWV-S-2004F1 | 2004-2033 | TAAGCAAGTTCTGCGAGTTTTGCCTGTTTT | 150 |
| KU179577.1 | TSWV-S-2153R1 | 2153–5124 | GAAGAAGGGGAAAGAGTATGCTGCTATACT | |
| Beet western yellows virus | BWYV-3892F1 | 3892–3921 | CAGAACTCCGGCTCCATCGCTTACGAGCTG | 169 |
| MK307779.1 | BWYV-4060R1 | 4060–4031 | GGATCCTGAATTGGTCCTCGGCGACGTCGT | |
| Pepper mottle virus | PepMoV-8916F1 | 8916–8945 | ATGGTTTGGTGCATTGAAAATGGCACGTCC | 174 |
| EU586122.1 | PepMoV-9089R1 | 9089–9060 | CATTTCTATATATGCCTCAGCCACATCAGA | |
| Hot pepper endornavirus | HPEV-13489F1 | 13489–13518 | CTTTGATGCAAGTAAAGCAGACATATTGGC | 183 |
| NC_027920.1 | HPEV-13671R1 | 13671–13642 | AGGTGAACATTTAATCTGTTAATAGCATGC | |
| Bell pepper endornavirus | BPEV-13478F1 | 13478–13507 | GTGGCAACTTATTTTGATGCAGACAAAGCA | 188 |
| NC_039216.1 | BPEV-13665R1 | 13665–13636 | ACATTTAGCCTGTTAATGGCATGTAGTTGC | |
| Pepper cryptic virus 1 RNA2 | PCV1s2-1081F1 | 1081–1110 | CCGCAACAGGTACAACACAACGAAGGAAGA | 163 |
| NC_037096.1 | PCV1s2-1243R1 | 1243–1214 | TTAGTCCTGATGACTGGAGGGAAGGTAACT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jo, Y.; Choi, H.; Lee, J.H.; Moh, S.H.; Cho, W.K. Viromes of 15 Pepper (Capsicum annuum L.) Cultivars. Int. J. Mol. Sci. 2022, 23, 10507. https://doi.org/10.3390/ijms231810507
Jo Y, Choi H, Lee JH, Moh SH, Cho WK. Viromes of 15 Pepper (Capsicum annuum L.) Cultivars. International Journal of Molecular Sciences. 2022; 23(18):10507. https://doi.org/10.3390/ijms231810507
Chicago/Turabian StyleJo, Yeonhwa, Hoseong Choi, Jeong Hun Lee, Sang Hyun Moh, and Won Kyong Cho. 2022. "Viromes of 15 Pepper (Capsicum annuum L.) Cultivars" International Journal of Molecular Sciences 23, no. 18: 10507. https://doi.org/10.3390/ijms231810507
APA StyleJo, Y., Choi, H., Lee, J. H., Moh, S. H., & Cho, W. K. (2022). Viromes of 15 Pepper (Capsicum annuum L.) Cultivars. International Journal of Molecular Sciences, 23(18), 10507. https://doi.org/10.3390/ijms231810507