

FGFR1–4 RNA-Based Gene Alteration and Expression Analysis in Squamous Non-Small Cell Lung Cancer

 , ,
, , 
Abstract
1. Introduction
2. Results
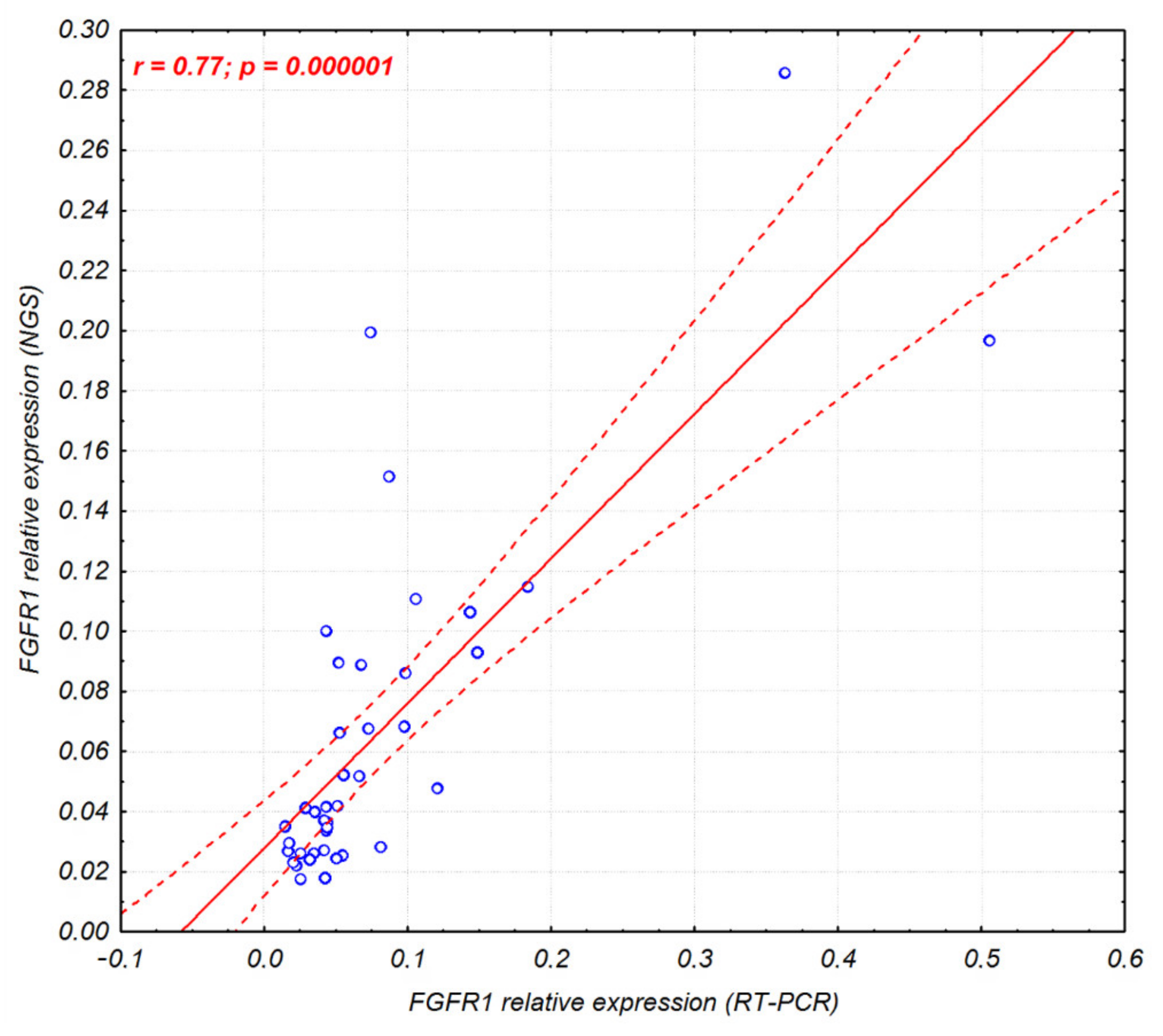
2.1. FGFR1–4 mRNA Expression in Sq-NSCLC Tumors and Tumor-Adjacent Normal Tissue
2.2. Clinical Significance of FGFR1–4 mRNA Expression
2.3. Association between the FGFR1–4 Expression and Clinicopathological Features
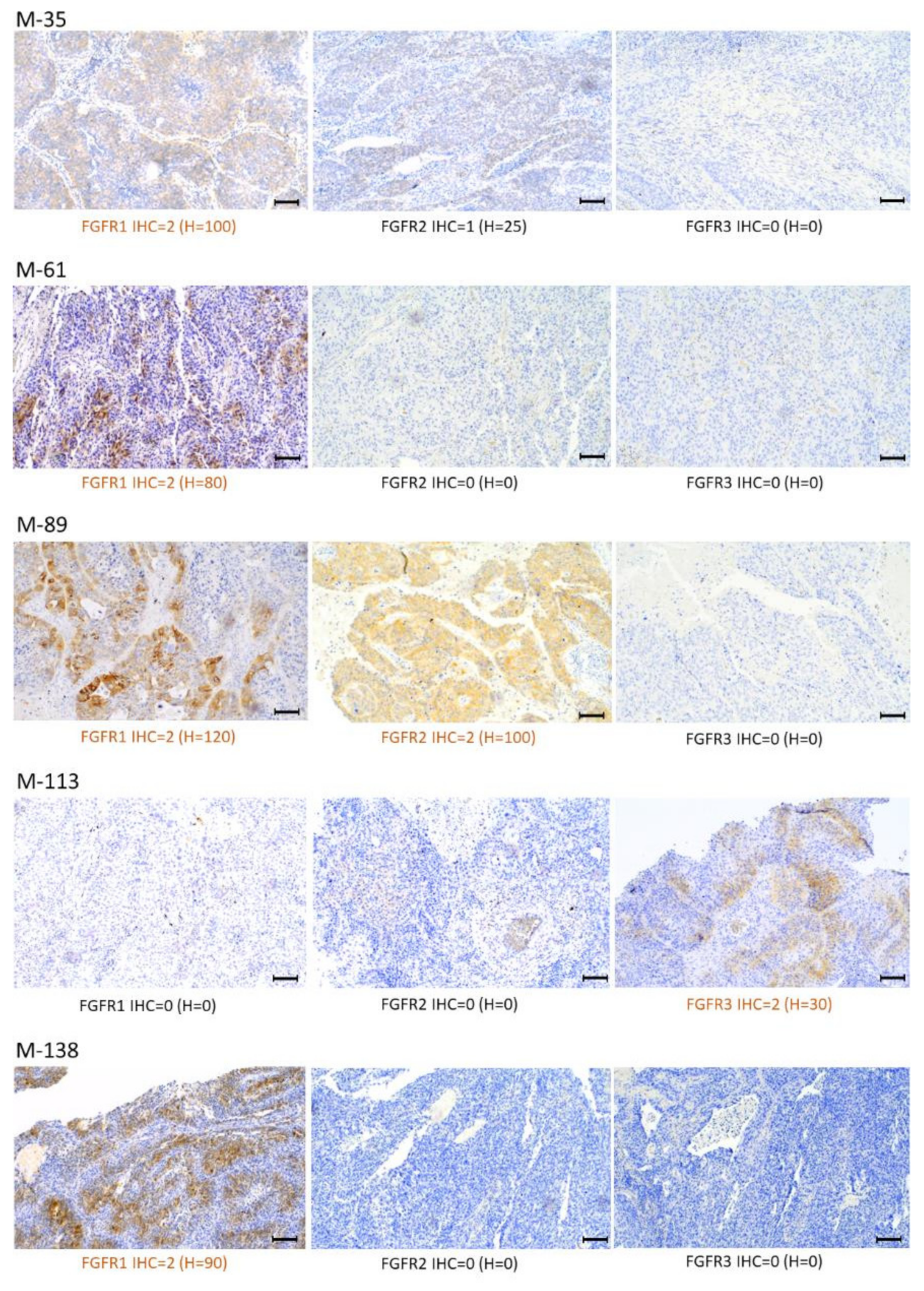
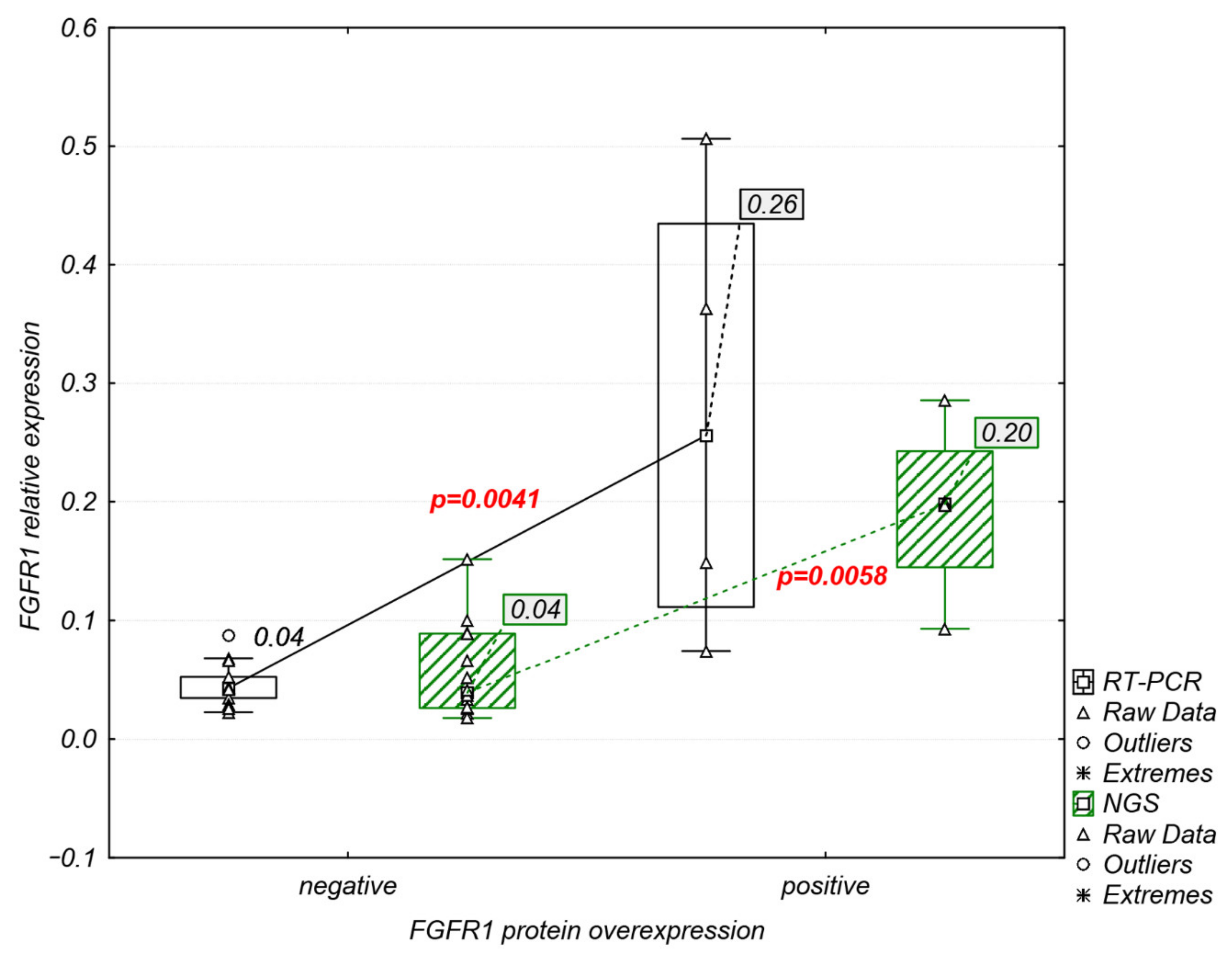
2.4. Analysis of FGFR1–3 Protein Overexpression and FGFR1 Amplification
2.5. Detection of FGFR Oncogenic Gene Fusions and Genetic Variants
2.5.1. Squamous Non-Small Lung Tumors
2.5.2. Commercial Controls
3. Discussion
4. Materials and Methods
4.1. Patient and Tumor Selection
4.2. RNA Control Material for NGS
4.3. RNA Extraction and cDNA Synthesis
4.4. Real-Time Polymerase Chain Reactions (RT-PCR)
4.5. Next-Generation Sequencing (NGS)
4.6. Immunohistochemistry (IHC)
4.7. Fluorescence In Situ Hybridization (FISH)
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Parkin, D.M.; Piñeros, M.; Znaor, A.; Bray, F. Cancer Statistics for the Year 2020: An Overview. Int. J. Cancer 2021, 149, 778–789. [Google Scholar] [CrossRef] [PubMed]
- Reck, M.; Remon, J.; Hellmann, M.D. First-Line Immunotherapy for Non-Small-Cell Lung Cancer. J. Clin. Oncol. 2022, 40, 586–597. [Google Scholar] [CrossRef] [PubMed]
- Reck, M.; Rodríguez-Abreu, D.; Robinson, A.G.; Hui, R.; Csőszi, T.; Fülöp, A.; Gottfried, M.; Peled, N.; Tafreshi, A.; Cuffe, S.; et al. Pembrolizumab versus Chemotherapy for PD-L1–Positive Non–Small-Cell Lung Cancer. N. Engl. J. Med. 2016, 375, 1823–1833. [Google Scholar] [CrossRef]
- Cancer Genome Atlas Research Network Comprehensive Genomic Characterization of Squamous Cell Lung Cancers. Nature 2012, 489, 519–525. [CrossRef]
- Babina, I.S.; Turner, N.C. Advances and Challenges in Targeting FGFR Signalling in Cancer. Nat. Rev. Cancer 2017, 17, 318–332. [Google Scholar] [CrossRef]
- Hashemi-Sadraei, N.; Hanna, N. Targeting FGFR in Squamous Cell Carcinoma of the Lung. Target. Oncol. 2017, 12, 741–755. [Google Scholar] [CrossRef]
- Moes-Sosnowska, J.; Chorostowska-Wynimko, J. Fibroblast Growth Factor Receptor 1-4 Genetic Aberrations as Clinically Relevant Biomarkers in Squamous Cell Lung Cancer. Front. Oncol. 2022, 12, 780650. [Google Scholar] [CrossRef] [PubMed]
- Nogova, L.; Sequist, L.V.; Perez Garcia, J.M.; Andre, F.; Delord, J.-P.; Hidalgo, M.; Schellens, J.H.M.; Cassier, P.A.; Camidge, D.R.; Schuler, M.; et al. Evaluation of BGJ398, a Fibroblast Growth Factor Receptor 1-3 Kinase Inhibitor, in Patients with Advanced Solid Tumors Harboring Genetic Alterations in Fibroblast Growth Factor Receptors: Results of a Global Phase I, Dose-Escalation and Dose-Expansion Study. J. Clin. Oncol. 2017, 35, 157–165. [Google Scholar] [CrossRef]
- Aggarwal, C.; Redman, M.W.; Lara, P.N., Jr.; Borghaei, H.; Hoffman, P.; Bradley, J.D.; Newman, A.J., III; Feldman, M.J.; Minichiello, K.; Miao, J.; et al. SWOG S1400D (NCT02965378), a Phase II Study of the Fibroblast Growth Factor Receptor Inhibitor AZD4547 in Previously Treated Patients with Fibroblast Growth Factor Pathway-Activated Stage IV Squamous Cell Lung Cancer (Lung-MAP Substudy). J. Thorac. Oncol. 2019, 14, 1847–1852. [Google Scholar] [CrossRef]
- Paik, P.K.; Shen, R.; Berger, M.F.; Ferry, D.; Soria, J.-C.; Mathewson, A.; Rooney, C.; Smith, N.R.; Cullberg, M.; Kilgour, E.; et al. A Phase Ib Open-Label Multicenter Study of AZD4547 in Patients with Advanced Squamous Cell Lung Cancers. Clin. Cancer Res. 2017, 23, 5366–5373. [Google Scholar] [CrossRef] [PubMed]
- Andre, F.; Ranson, M.; Dean, E.; Varga, A.; van der Noll, R.; Stockman, P.K.; Ghiorghiu, D.; Kilgour, E.; Smith, P.D.; Macpherson, M.; et al. Abstract LB-145: Results of a Phase I Study of AZD4547, an Inhibitor of Fibroblast Growth Factor Receptor (FGFR), in Patients with Advanced Solid Tumors. Cancer Res. 2013, 73, LB-145. [Google Scholar] [CrossRef]
- Rooney, C.; Geh, C.; Williams, V.; Heuckmann, J.M.; Menon, R.; Schneider, P.; Al-Kadhimi, K.; Dymond, M.; Smith, N.R.; Baker, D.; et al. Characterization of FGFR1 Locus in SqNSCLC Reveals a Broad and Heterogeneous Amplicon. PLoS ONE 2016, 11, e0149628. [Google Scholar] [CrossRef]
- Ren, M.; Hong, M.; Liu, G.; Wang, H.; Patel, V.; Biddinger, P.; Silva, J.; Cowell, J.; Hao, Z. Novel FGFR Inhibitor Ponatinib Suppresses the Growth of Non-Small Cell Lung Cancer Cells Overexpressing FGFR1. Oncol. Rep. 2013, 29, 2181–2190. [Google Scholar] [CrossRef] [PubMed]
- Wynes, M.W.; Hinz, T.K.; Gao, D.; Martini, M.; Marek, L.A.; Ware, K.E.; Edwards, M.G.; Böhm, D.; Perner, S.; Helfrich, B.A.; et al. FGFR1 MRNA and Protein Expression, Not Gene Copy Number, Predict FGFR TKI Sensitivity across All Lung Cancer Histologies. Clin. Cancer Res. 2014, 20, 3299–3309. [Google Scholar] [CrossRef] [PubMed]
- Grünewald, S.; Politz, O.; Bender, S.; Héroult, M.; Lustig, K.; Thuss, U.; Kneip, C.; Kopitz, C.; Zopf, D.; Collin, M.-P.; et al. Rogaratinib: A Potent and Selective Pan-FGFR Inhibitor with Broad Antitumor Activity in FGFR-Overexpressing Preclinical Cancer Models. Int. J. Cancer 2019, 145, 1346–1357. [Google Scholar] [CrossRef]
- Addeo, A.; Joerger, M.; Rothschild, S.; Eboulet, E.I.; Godar, G.; Waibel-Pachinger, C.; Haefliger, S.; Mark, M.T.; Fernandez, E.; Mach, N.; et al. Fibroblast Growth Factor Receptor (FGFR) Inhibitor Rogaratinib in Patients with Advanced Pretreated Squamous-Cell Non-Small Cell Lung Cancer over-Expressing FGFR MRNA: The SAKK 19/18 Phase II Study. J. Clin. Oncol. 2021, 39, e21119. [Google Scholar] [CrossRef]
- Ng, T.L.; Yu, H.; Smith, D.E.; Boyle, T.A.; York, E.R.; Leedy, S.; Gao, D.; Aisner, D.L.; Van Bokhoven, A.; Heasley, L.E.; et al. Preselection of Lung Cancer Cases Using FGFR1 MRNA and Gene Copy Number for Treatment with Ponatinib. Clin. Lung Cancer 2019, 20, e39–e51. [Google Scholar] [CrossRef]
- Ornitz, D.M.; Itoh, N. The Fibroblast Growth Factor Signaling Pathway. Wiley Interdiscip. Rev. Dev. Biol. 2015, 4, 215–266. [Google Scholar] [CrossRef]
- Bogatyrova, O.; Mattsson, J.S.M.; Ross, E.M.; Sanderson, M.P.; Backman, M.; Botling, J.; Brunnström, H.; Kurppa, P.; La Fleur, L.; Strell, C.; et al. FGFR1 Overexpression in Non-Small Cell Lung Cancer Is Mediated by Genetic and Epigenetic Mechanisms and Is a Determinant of FGFR1 Inhibitor Response. Eur. J. Cancer 2021, 151, 136–149. [Google Scholar] [CrossRef]
- Gresner, P.; Gromadzinska, J.; Wasowicz, W. Reference Genes for Gene Expression Studies on Non-Small Cell Lung Cancer. Acta Biochim. Pol. 2009, 56, 307–316. [Google Scholar] [CrossRef]
- Smith, T.A.D.; AbdelKarem, O.A.; Irlam-Jones, J.J.; Lane, B.; Valentine, H.; Bibby, B.A.S.; Denley, H.; Choudhury, A.; West, C.M.L. Selection of Endogenous Control Genes for Normalising Gene Expression Data Derived from Formalin-Fixed Paraffin-Embedded Tumour Tissue. Sci. Rep. 2020, 10, 17258. [Google Scholar] [CrossRef] [PubMed]
- Moes-Sosnowska, J.; Szczepulska-Wojcik, E.; Rozy, A.; Rudzinski, S.; Langfort, R.; Rudzinski, P.; Orlowski, T.; Chorostowska-Wynimko, J. FGFR1–4 and MET Expression Analysis and Evaluation of Reliable Reference Genes in Sq-NSCLC. Eur. Respir. J. 2019, 54, PA3665. [Google Scholar] [CrossRef]
- Huang, H.-P.; Feng, H.; Qiao, H.-B.; Ren, Z.-X.; Zhu, G.-D. The Prognostic Significance of Fibroblast Growth Factor Receptor 4 in Non-Small-Cell Lung Cancer. Onco. Targets. Ther. 2015, 8, 1157–1164. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Justet, A.; Ghanem, M.; Jaillet, M.; Hachem, M.; Boghanim, T.; Vadel, A.; Mailleux, A.; Crestani, B. FGFR4 Has pro Fibrotic Properties in Idiopathic Pulmonary Fibrosis. Eur. Respir. J. 2020, 56, 3359. [Google Scholar] [CrossRef]
- Quintanal-Villalonga, Á.; Ferrer, I.; Guruceaga, E.; Cirauqui, C.; Marrugal, Á.; Ojeda, L.; García, S.; Zugazagoitia, J.; Muñoz-Galván, S.; Lopez-Rios, F.; et al. FGFR1 and FGFR4 Oncogenicity Depends on N-Cadherin and Their Co-Expression May Predict FGFR-Targeted Therapy Efficacy. EBioMedicine 2020, 53, 102683. [Google Scholar] [CrossRef]
- Wang, L.; Ren, Z.; Yu, B.; Tang, J. Development of Nomogram Based on Immune-Related Gene FGFR4 for Advanced Non-Small Cell Lung Cancer Patients with Sensitivity to Immune Checkpoint Inhibitors. J. Transl. Med. 2021, 19, 22. [Google Scholar] [CrossRef]
- Wei, W.; Cao, S.; Liu, J.; Wang, Y.; Song, Q.; Leha, A.; Sun, S.; Zhang, X.; Liang, X.; Jiang, Y. Fibroblast Growth Factor Receptor 4 as a Prognostic Indicator in Triple-Negative Breast Cancer. Transl. Cancer Res. 2020, 9, 6881–6888. [Google Scholar] [CrossRef]
- Inokuchi, M.; Murase, H.; Otsuki, S.; Kawano, T.; Kojima, K. Different Clinical Significance of FGFR1–4 Expression between Diffuse-Type and Intestinal-Type Gastric Cancer. World J. Surg. Oncol. 2017, 15, 2. [Google Scholar] [CrossRef]
- Murase, H.; Inokuchi, M.; Takagi, Y.; Kato, K.; Kojima, K.; Sugihara, K. Prognostic Significance of the Co-Overexpression of Fibroblast Growth Factor Receptors 1, 2 and 4 in Gastric Cancer. Mol. Clin. Oncol. 2014, 2, 509–517. [Google Scholar] [CrossRef]
- Jaakkola, S.; Salmikangas, P.; Nylund, S.; Partanen, J.; Armstrong, E.; Pyrhönen, S.; Lehtovirta, P.; Nevanlinna, H. Amplification of Fgfr4 Gene in Human Breast and Gynecological Cancers. Int. J. Cancer 1993, 54, 378–382. [Google Scholar] [CrossRef]
- Quintanal-Villalonga, Á.; Carranza-Carranza, A.; Meléndez, R.; Ferrer, I.; Molina-Pinelo, S.; Paz-Ares, L. Prognostic Role of the FGFR4-388Arg Variant in Lung Squamous-Cell Carcinoma Patients with Lymph Node Involvement. Clin. Lung Cancer 2017, 18, 667–674.e1. [Google Scholar] [CrossRef]
- Wang, J.; Yu, W.; Cai, Y.; Ren, C.; Ittmann, M.M. Altered Fibroblast Growth Factor Receptor 4 Stability Promotes Prostate Cancer Progression. Neoplasia 2008, 10, 847–856. [Google Scholar] [CrossRef] [PubMed]
- Nannapaneni, S.; Griffith, C.C.; Magliocca, K.R.; Chen, W.; Lyu, X.; Chen, Z.; Wang, D.; Wang, X.; Shin, D.M.; Chen, Z.G.; et al. Co-Expression of Fibroblast Growth Factor Receptor 3 with Mutant P53, and Its Association with Worse Outcome in Oropharyngeal Squamous Cell Carcinoma. PLoS ONE 2021, 16, e0247498. [Google Scholar] [CrossRef]
- Sikic, D.; Taubert, H.; Breyer, J.; Eckstein, M.; Weyerer, V.; Keck, B.; Kubon, J.; Otto, W.; Worst, T.S.; Kriegmair, M.C.; et al. The Prognostic Value of FGFR3 Expression in Patients with T1 Non-Muscle Invasive Bladder Cancer. Cancer Manag. Res. 2021, 13, 6567–6578. [Google Scholar] [CrossRef] [PubMed]
- Jing, P.; Zhao, N.; Xie, N.; Ye, M.; Zhang, Y.; Zhang, Z.; Li, M.; Lai, X.; Zhang, J.; Gu, Z. MiR-24-3p/FGFR3 Signaling as a Novel Axis Is Involved in Epithelial-Mesenchymal Transition and Regulates Lung Adenocarcinoma Progression. J. Immunol. Res. 2018, 2018, 2834109. [Google Scholar] [CrossRef]
- Starska, K.; Forma, E.; Lewy-Trenda, I.; Stasikowska-Kanicka, O.; Skóra, M.; Bryś, M. Fibroblast Growth Factor Receptor 1 and 3 Expression Is Associated with Regulatory PI3K/AKT Kinase Activity, as Well as Invasion and Prognosis, in Human Laryngeal Cancer. Cell. Oncol. 2018, 41, 253–268. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.W.; Kim, Y.-H.; Jeong, P.; Park, C.; Kim, W.T.; Ryu, D.H.; Cha, E.-J.; Ha, Y.-S.; Kim, T.-H.; Kwon, T.G.; et al. Expression Levels of FGFR3 as a Prognostic Marker for the Progression of Primary PT1 Bladder Cancer and Its Association with Mutation Status. Oncol. Lett. 2017, 14, 3817–3824. [Google Scholar] [CrossRef] [PubMed]
- Weiss, J.; Sos, M.L.; Seidel, D.; Peifer, M.; Zander, T.; Heuckmann, J.M.; Ullrich, R.T.; Menon, R.; Maier, S.; Soltermann, A.; et al. Frequent and Focal FGFR1 Amplification Associates with Therapeutically Tractable FGFR1 Dependency in Squamous Cell Lung Cancer. Sci. Transl. Med. 2010, 2, 62ra93. [Google Scholar] [CrossRef] [PubMed]
- Gil, F.; Miranda-Filho, A.; Uribe-Perez, C.; Arias-Ortiz, N.E.; Yépez-Chamorro, M.C.; Bravo, L.M.; de Vries, E. Impact of the Management and Proportion of Lost to Follow-up Cases on Cancer Survival Estimates for Small Population-Based Cancer Registries. J. Cancer Epidemiol. 2022, 2022, 9068214. [Google Scholar] [CrossRef]
- Kang, D.; Jung, J.; Park, S.; Cho, B.-S.; Kim, H.-J.; Kim, Y.; Lee, J.-M.; Kim, H.S.; Ahn, A.; Kim, M.; et al. Genetic Characteristics According to Subgroup of Acute Myeloid Leukemia with Myelodysplasia-Related Changes. J. Clin. Med. 2022, 11, 2378. [Google Scholar] [CrossRef]
- Desmeules, P.; Boudreau, D.K.; Bastien, N.; Boulanger, M.-C.; Bossé, Y.; Joubert, P.; Couture, C. Performance of an RNA-Based next-Generation Sequencing Assay for Combined Detection of Clinically Actionable Fusions and Hotspot Mutations in NSCLC. JTO Clin. Res. Rep. 2022, 3, 100276. [Google Scholar] [CrossRef] [PubMed]
- Guseva, N.V.; Jaber, O.; Tanas, M.R.; Stence, A.A.; Sompallae, R.; Schade, J.; Fillman, A.N.; Miller, B.J.; Bossler, A.D.; Ma, D. Anchored Multiplex PCR for Targeted Next-Generation Sequencing Reveals Recurrent and Novel USP6 Fusions and Upregulation of USP6 Expression in Aneurysmal Bone Cyst. Genes Chromosomes Cancer 2017, 56, 266–277. [Google Scholar] [CrossRef] [PubMed]
- Sands, J.M.; Nguyen, T.; Shivdasani, P.; Sacher, A.G.; Cheng, M.L.; Alden, R.S.; Jänne, P.A.; Kuo, F.C.; Oxnard, G.R.; Sholl, L.M. Next-Generation Sequencing Informs Diagnosis and Identifies Unexpected Therapeutic Targets in Lung Squamous Cell Carcinomas. Lung Cancer 2020, 140, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Theelen, W.S.; Mittempergher, L.; Willems, S.M.; Bosma, A.J.; Peters, D.D.; van der Noort, V.; Japenga, E.J.; Peeters, T.; Koole, K.; Šuštić, T.; et al. FGFR1, 2 and 3 Protein Overexpression and Molecular Aberrations of FGFR3 in Early Stage Non-Small Cell Lung Cancer: FGFR Expression in Early Stage Non-Small Lung Cancer. J. Pathol. Clin. Res. 2016, 2, 223–233. [Google Scholar] [CrossRef]
- Hibi, M.; Kaneda, H.; Tanizaki, J.; Sakai, K.; Togashi, Y.; Terashima, M.; De Velasco, M.A.; Fujita, Y.; Banno, E.; Nakamura, Y.; et al. FGFR Gene Alterations in Lung Squamous Cell Carcinoma Are Potential Targets for the Multikinase Inhibitor Nintedanib. Cancer Sci. 2016, 107, 1667–1676. [Google Scholar] [CrossRef]
- Lim, S.H.; Sun, J.-M.; Choi, Y.-L.; Kim, H.R.; Ahn, S.; Lee, J.Y.; Lee, S.-H.; Ahn, J.S.; Park, K.; Kim, J.H.; et al. Efficacy and Safety of Dovitinib in Pretreated Patients with Advanced Squamous Non-Small Cell Lung Cancer with FGFR1 Amplification: A Single-Arm, Phase 2 Study: Dovitinib InFGFR1-Amplified SCC. Cancer 2016, 122, 3024–3031. [Google Scholar] [CrossRef]
- Wu, Y.-M.; Su, F.; Kalyana-Sundaram, S.; Khazanov, N.; Ateeq, B.; Cao, X.; Lonigro, R.J.; Vats, P.; Wang, R.; Lin, S.-F.; et al. Identification of Targetable FGFR Gene Fusions in Diverse Cancers. Cancer Discov. 2013, 3, 636–647. [Google Scholar] [CrossRef]
- Stransky, N.; Cerami, E.; Schalm, S.; Kim, J.L.; Lengauer, C. The Landscape of Kinase Fusions in Cancer. Nat. Commun. 2014, 5, 4846. [Google Scholar] [CrossRef]
- Wang, R.; Wang, L.; Li, Y.; Hu, H.; Shen, L.; Shen, X.; Pan, Y.; Ye, T.; Zhang, Y.; Luo, X.; et al. FGFR1/3 Tyrosine Kinase Fusions Define a Unique Molecular Subtype of Non-Small Cell Lung Cancer. Clin. Cancer Res. 2014, 20, 4107–4114. [Google Scholar] [CrossRef]
- Qin, A.; Johnson, A.; Ross, J.S.; Miller, V.A.; Ali, S.M.; Schrock, A.B.; Gadgeel, S.M. Detection of Known and Novel FGFR Fusions in Non-Small Cell Lung Cancer by Comprehensive Genomic Profiling. J. Thorac. Oncol. 2019, 14, 54–62. [Google Scholar] [CrossRef]
- Kim, Y.; Hammerman, P.S.; Kim, J.; Yoon, J.-A.; Lee, Y.; Sun, J.-M.; Wilkerson, M.D.; Pedamallu, C.S.; Cibulskis, K.; Yoo, Y.K.; et al. Integrative and Comparative Genomic Analysis of Lung Squamous Cell Carcinomas in East Asian Patients. J. Clin. Oncol. 2014, 32, 121–128. [Google Scholar] [CrossRef]
- Zhou, Z.; Liu, Z.; Ou, Q.; Wu, X.; Wang, X.; Shao, Y.; Liu, H.; Yang, Y. Targeting FGFR in Non-Small Cell Lung Cancer: Implications from the Landscape of Clinically Actionable Aberrations of FGFR Kinases. Cancer Biol. Med. 2021, 18, 490–501. [Google Scholar] [CrossRef] [PubMed]
- Gozgit, J.M.; Wong, M.J.; Moran, L.; Wardwell, S.; Mohemmad, Q.K.; Narasimhan, N.I.; Shakespeare, W.C.; Wang, F.; Clackson, T.; Rivera, V.M. Ponatinib (AP24534), a Multitargeted Pan-FGFR Inhibitor with Activity in Multiple FGFR-Amplified or Mutated Cancer Models. Mol. Cancer Ther. 2012, 11, 690–699. [Google Scholar] [CrossRef] [PubMed]
- Schittenhelm, J.; Ziegler, L.; Sperveslage, J.; Mittelbronn, M.; Capper, D.; Burghardt, I.; Poso, A.; Biskup, S.; Skardelly, M.; Tabatabai, G. FGFR3 Overexpression Is a Useful Detection Tool for FGFR3 Fusions and Sequence Variations in Glioma. Neurooncol. Pract. 2021, 8, 209–221. [Google Scholar] [CrossRef] [PubMed]
- Parker, B.C.; Annala, M.J.; Cogdell, D.E.; Granberg, K.J.; Sun, Y.; Ji, P.; Li, X.; Gumin, J.; Zheng, H.; Hu, L.; et al. The Tumorigenic FGFR3-TACC3 Gene Fusion Escapes MiR-99a Regulation in Glioblastoma. J. Clin. Investig. 2013, 123, 855–865. [Google Scholar] [CrossRef] [PubMed]
- Helsten, T.; Elkin, S.; Arthur, E.; Tomson, B.N.; Carter, J.; Kurzrock, R. The FGFR Landscape in Cancer: Analysis of 4,853 Tumors by next-Generation Sequencing. Clin. Cancer Res. 2016, 22, 259–267. [Google Scholar] [CrossRef]
- Liao, R.G.; Jung, J.; Tchaicha, J.; Wilkerson, M.D.; Sivachenko, A.; Beauchamp, E.M.; Liu, Q.; Pugh, T.J.; Pedamallu, C.S.; Hayes, D.N.; et al. Inhibitor-Sensitive FGFR2 and FGFR3 Mutations in Lung Squamous Cell Carcinoma. Cancer Res. 2013, 73, 5195–5205. [Google Scholar] [CrossRef]
- Dutt, A.; Salvesen, H.B.; Chen, T.-H.; Ramos, A.H.; Onofrio, R.C.; Hatton, C.; Nicoletti, R.; Winckler, W.; Grewal, R.; Hanna, M.; et al. Drug-Sensitive FGFR2 Mutations in Endometrial Carcinoma. Proc. Natl. Acad. Sci. USA 2008, 105, 8713–8717. [Google Scholar] [CrossRef]
- Majewski, I.J.; Mittempergher, L.; Davidson, N.M.; Bosma, A.; Willems, S.M.; Horlings, H.M.; de Rink, I.; Greger, L.; Hooijer, G.K.J.; Peters, D.; et al. Identification of Recurrent FGFR3 Fusion Genes in Lung Cancer through Kinome-Centred RNA Sequencing: Kinase Fusion Genes in NSCLC. J. Pathol. 2013, 230, 270–276. [Google Scholar] [CrossRef]
- Flockerzi, F.A.; Roggia, C.; Langer, F.; Holleczek, B.; Bohle, R.M. FGFR1 Gene Amplification in Squamous Cell Carcinomas of the Lung: A Potential Favorable Prognostic Marker for Women and for Patients with Advanced Cancer. Virchows Arch. 2018, 472, 759–769. [Google Scholar] [CrossRef]
- Nakamura, I.T.; Kohsaka, S.; Ikegami, M.; Ikeuchi, H.; Ueno, T.; Li, K.; Beyett, T.S.; Koyama, T.; Shimizu, T.; Yamamoto, N.; et al. Comprehensive Functional Evaluation of Variants of Fibroblast Growth Factor Receptor Genes in Cancer. NPJ Precis. Oncol. 2021, 5, 66. [Google Scholar] [CrossRef] [PubMed]
- Wilkie, A.O.M.; Bochukova, E.G.; Hansen, R.M.S.; Taylor, I.B.; Rannan-Eliya, S.V.; Byren, J.C.; Wall, S.A.; Ramos, L.; Venâncio, M.; Hurst, J.A.; et al. Clinical Dividends from the Molecular Genetic Diagnosis of Craniosynostosis. Am. J. Med. Genet. A 2006, 140, 2631–2639. [Google Scholar] [CrossRef] [PubMed]
- Kress, W.; Petersen, B.; Collmann, H.; Grimm, T. An Unusual FGFR1 Mutation (Fibroblast Growth Factor Receptor 1 Mutation) in a Girl with Non-Syndromic Trigonocephaly. Cytogenet. Cell Genet. 2000, 91, 138–140. [Google Scholar] [CrossRef] [PubMed]
- Richards, S.; Aziz, N.; Bale, S.; Bick, D.; Das, S.; Gastier-Foster, J.; Grody, W.W.; Hegde, M.; Lyon, E.; Spector, E.; et al. Standards and Guidelines for the Interpretation of Sequence Variants: A Joint Consensus Recommendation of the American College of Medical Genetics and Genomics and the Association for Molecular Pathology. Genet. Med. 2015, 17, 405–424. [Google Scholar] [CrossRef] [PubMed]
- Joerger, M.; Cho, B.C.; Mach, N.; Caballero, C.; Soo, R.A.; Wirth, M.; Sayehli, C.; Navarro, A.; Keam, B.; Piciu, A.-M.; et al. Early Clinical Experience with the Pan-FGFR Inhibitor Rogaratinib in Patients with Non-Small Cell Lung Cancer Selected Based on FGFR MRNA Expression Levels. J. Clin. Oncol. 2019, 37, e20661. [Google Scholar] [CrossRef]
- De Luca, A.; Esposito Abate, R.; Rachiglio, A.M.; Maiello, M.R.; Esposito, C.; Schettino, C.; Izzo, F.; Nasti, G.; Normanno, N. FGFR Fusions in Cancer: From Diagnostic Approaches to Therapeutic Intervention. Int. J. Mol. Sci. 2020, 21, 6856. [Google Scholar] [CrossRef]
- Bahleda, R.; Italiano, A.; Hierro, C.; Mita, A.; Cervantes, A.; Chan, N.; Awad, M.; Calvo, E.; Moreno, V.; Govindan, R.; et al. Multicenter Phase I Study of Erdafitinib (JNJ-42756493), Oral Pan-Fibroblast Growth Factor Receptor Inhibitor, in Patients with Advanced or Refractory Solid Tumors. Clin. Cancer Res. 2019, 25, 4888–4897. [Google Scholar] [CrossRef]
- Travis, W.D.; Brambilla, E.; Burke, A.P.; Marx, A.; Nicholson, A.G. Introduction to the 2015 World Health Organization Classification of Tumors of the Lung, Pleura, Thymus, and Heart. J. Thorac. Oncol. 2015, 10, 1240–1242. [Google Scholar] [CrossRef]
- Brierley, J.D.; Gospodarowicz, M.K.; Wittekind, C. TNM Classification of Malignant Tumours; John Wiley & Sons: Nashville, TN, USA, 2016; ISBN 9781119263579. [Google Scholar]
- Skupinska, M.M.; Obtulowicz, T.; Moes-Sosnowska, J.; Rozy, A.; Szczepulska, E.; Langfort, R.; Wynimko, J.C.; Stanczak, A.; Pieczykolan, J.; Wieczorek, M.; et al. 1409P Comparing Different Methods of FGFR1 Aberrations Analysis in Squamous Cell Lung Cancer (SqCLC) Targeted Therapy. Ann. Oncol. 2020, 31, S891. [Google Scholar] [CrossRef]
- Skupinska, M.M.; Jesiotr, M.; Chrom, P.; Mroz, A.; Cierniak, S.; Winiarek, M.; Wyrwicz, L.S.; Pieczykolan, J.; Wieczorek, M.; Stanczak, A.; et al. The Role of FGFR2 Amplification and Expression in Patients with Advanced or Metastatic Gastric Cancer Receiving Fluoropyrimidine-Based Chemotherapy. Ann. Oncol. 2018, 29 (Suppl. 8), viii218. [Google Scholar] [CrossRef]
- Sosnowski, R.; Popiel, D.; Gapska, P.; Skupińska, M.; Stajno, P.; Sobieszek-Prochorec, M.; Ligaj, M.; Demkow, T.; Stańczak, A.; Wieczorek, M.; et al. 712P Analysis of Fibroblast Growth Factor Receptor 3 Aberrations in Bladder Cancer, for Enabling Personalized and Effective Therapy Based on FGFR Inhibitor. Ann. Oncol. 2021, 32, S719. [Google Scholar] [CrossRef]
- Dowsett, M.; Bartlett, J.; Ellis, I.O.; Salter, J.; Hills, M.; Mallon, E.; Watters, A.D.; Cooke, T.; Paish, C.; Wencyk, P.M.; et al. Correlation between Immunohistochemistry (HercepTest) and Fluorescence in Situ Hybridization (FISH) for HER-2 in 426 Breast Carcinomas from 37 Centres. J. Pathol. 2003, 199, 418–423. [Google Scholar] [CrossRef] [PubMed]
- Tomlinson, D.C.; Baldo, O.; Harnden, P.; Knowles, M.A. FGFR3 Protein Expression and Its Relationship to Mutation Status and Prognostic Variables in Bladder Cancer. J. Pathol. 2007, 213, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Schultheis, A.M.; Bos, M.; Schmitz, K.; Wilsberg, L.; Binot, E.; Wolf, J.; Büttner, R.; Schildhaus, H.-U. Fibroblast Growth Factor Receptor 1 (FGFR1) Amplification Is a Potential Therapeutic Target in Small-Cell Lung Cancer. Mod. Pathol. 2014, 27, 214–221. [Google Scholar] [CrossRef] [PubMed][Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patients n = 40 | Frequency | % |
|---|---|---|
| Age (Range 47–80 Years) Median (67 Years) | ||
| <67 (0) | 18 | 45.00 |
| ≥67 (1) | 22 | 55.00 |
| Gender | ||
| Male (0) | 30 | 75.00 |
| Female (1) | 10 | 25.00 |
| Histological type | ||
| Squamous cell carcinoma | 34 | 85.0 |
| Carcinoma adenosquamous | 6 | 15.0 |
| Histopathological Grade | ||
| G1.G2 (0) | 25 | 62.5 |
| G3.G4 (1) | 15 | 37.5 |
| Clinical stage | ||
| IA. IB. IIA (0) | 21 | 52.5 |
| IIB. IIIA. IV (1) | 18 | 45.0 |
| Cannot be assessed | 1 | 2.50 |
| pT-Primary Tumor size | ||
| ≤4 cm (0) | 18 | 45.0 |
| >4 cm (1) | 22 | 55.0 |
| pN-Regional Lymph Nodes invasion | ||
| Negative (0) | 28 | 70.0 |
| Positive (1) | 11 | 27.5 |
| Cannot be assessed | 1 | 2.50 |
| pV-Venous Invasion | ||
| Negative (0) | 24 | 60.0 |
| Positive (microscopic) (1) | 16 | 40.0 |
| pL-Lymphatic Invasion | ||
| Negative (0) | 21 | 52.5 |
| Positive (1) | 19 | 47.5 |
| pM-Distant metastasis | ||
| Negative (0) | 19 | 47.5 |
| Positive (1) | 1 | 2.50 |
| Not available | 20 | 50.0 |
| pR-Residual tumor | ||
| Negative (0) | 35 | 87.5 |
| Positive (1) | 4 | 10.0 |
| Cannot be assessed | 1 | 2.50 |
| Actinic keratosis | ||
| Negative (0) | 13 | 32.5 |
| Positive (1) | 25 | 62.5 |
| Cannot be assessed | 2 | 5.0 |
| FGFR1 protein expression | ||
| Negative (0) | 14 | 35.0 |
| Positive (1) | 4 | 10.0 |
| Not assessed | 22 | 55.0 |
| FGFR2 protein expression | ||
| Negative (0) | 17 | 42.5 |
| Positive (1) | 1 | 2.5 |
| Not assessed | 22 | 55.0 |
| FGFR3 protein expression | ||
| Negative (0) | 17 | 42.5 |
| Positive (1) | 1 | 2.5 |
| Not assessed | 22 | 55.0 |
| FGFR1 amplification | ||
| Negative (0) | 8 | 20.0 |
| Positive (1) | 7 | 17.5 |
| Not assessed | 25 | 62.5 |
| Number of patients at risk of recurrence (DFS) | ||
| 1 year | 3 | 7.5 |
| 2 year | 4 | 10.0 |
| 3 year | 9 | 12.5 |
| >3 year | 5 | 47.5 |
| No follow up data | 19 | 47.5 |
| Outcome | ||
| NED | 15 | 37.5 |
| AWD | 6 | 15.0 |
| DOD | - | - |
| DOC | - | - |
| No follow up data | 19 | 47.5 |
| FGFR RNA Variants | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Patient No | Sample Name | Symbol | HGVSc | HGVSp | Consequence | Clinical Significance | Depth | AO | AF |
| 1 | M-1 | FGFR2 | c.2419G>A | p.(Glu807Lys) | Missense | Uncertain significance/Benign | 796 | 314 | 0.39 |
| 2 | M-6 | FGFR2 | c.2211G>T | p.(Met737Ile) | Missense | No information/Pathogenic | 700 | 427 | 0.61 |
| 3 | M-11 | FGFR2 | c.2398dup | p.(Ser800PhefsTer22) | Frameshift | Uncertain significance/Pathogenic | 3136 | 62 | 0.01 |
| 4 | M-17 | FGFR2 | c.2398dup | p.(Ser800PhefsTer22) | frameshift | Uncertain significance/Pathogenic | 2624 | 63 | 0.02 |
| 5 | M-77 | FGFR2 | c.2398dup | p.(Ser800PhefsTer22) | frameshift | Uncertain significance/Pathogenic | 2898 | 81 | 0.03 |
| 6 | M-88 | FGFR1 | c.899T>C | p.(Ile300Thr) | missense | Uncertain significance/Uncertain significance | 278 | 184 | 0.66 |
| 7 | M-115 | FGFR3 | c.746C>G | p.(Ser249Cys) (S249C) | missense | Pathogenic/Pathogenic | 251 | 124 | 0.49 |
| 8 | M-119 | FGFR2 | c.870G>T | p.(Trp290Cys) (W290C) | missense | Pathogenic/Pathogenic | 304 | 24 | 0.08 |
| 9 | M-123 | FGFR2 | c.2398dup | p.(Ser800PhefsTer22) | frameshift | Uncertain significance/Pathogenic | 3016 | 70 | 0.02 |
| 10 | M-135 | FGFR2 | c.2398dup | p.(Ser800PhefsTer22) | frameshift | Uncertain significance/Pathogenic | 1189 | 30 | 0.02 |
| M-135P | FGFR2 | c.2398dup | p.(Ser800PhefsTer22) | frameshift | Uncertain significance/Pathogenic | 643 | 15 | 0.02 | |
| 11 | M-138 | FGFR2 | c.2398dup | p.(Ser800PhefsTer22) | frameshift | Uncertain significance/Pathogenic | 1633 | 40 | 0.02 |
| M-138P | nd | ||||||||
| FGFR RNA Fusions | |||||||||
| Patient no | Sample name | Symbol | SS | Reads | %Reads | Breakpoint | |||
| 12 | M-20 | FGFR3::TACC3 | 17 | 35 | 5.49 | chr4:1808661.chr4:1741429 | |||
| M-20P | nd | ||||||||
| 13 | M-61 | TACC1::FGFR1 | 17 | 25 | 4.66 | chr8:38646337.chr8:38315052 | |||
| M-61P | nd | ||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moes-Sosnowska, J.; Skupinska, M.; Lechowicz, U.; Szczepulska-Wojcik, E.; Skronska, P.; Rozy, A.; Stepniewska, A.; Langfort, R.; Rudzinski, P.; Orlowski, T.; et al. FGFR1–4 RNA-Based Gene Alteration and Expression Analysis in Squamous Non-Small Cell Lung Cancer. Int. J. Mol. Sci. 2022, 23, 10506. https://doi.org/10.3390/ijms231810506
Moes-Sosnowska J, Skupinska M, Lechowicz U, Szczepulska-Wojcik E, Skronska P, Rozy A, Stepniewska A, Langfort R, Rudzinski P, Orlowski T, et al. FGFR1–4 RNA-Based Gene Alteration and Expression Analysis in Squamous Non-Small Cell Lung Cancer. International Journal of Molecular Sciences. 2022; 23(18):10506. https://doi.org/10.3390/ijms231810506
Chicago/Turabian StyleMoes-Sosnowska, Joanna, Monika Skupinska, Urszula Lechowicz, Ewa Szczepulska-Wojcik, Paulina Skronska, Adriana Rozy, Aneta Stepniewska, Renata Langfort, Piotr Rudzinski, Tadeusz Orlowski, and et al. 2022. "FGFR1–4 RNA-Based Gene Alteration and Expression Analysis in Squamous Non-Small Cell Lung Cancer" International Journal of Molecular Sciences 23, no. 18: 10506. https://doi.org/10.3390/ijms231810506
APA StyleMoes-Sosnowska, J., Skupinska, M., Lechowicz, U., Szczepulska-Wojcik, E., Skronska, P., Rozy, A., Stepniewska, A., Langfort, R., Rudzinski, P., Orlowski, T., Popiel, D., Stanczak, A., Wieczorek, M., & Chorostowska-Wynimko, J. (2022). FGFR1–4 RNA-Based Gene Alteration and Expression Analysis in Squamous Non-Small Cell Lung Cancer. International Journal of Molecular Sciences, 23(18), 10506. https://doi.org/10.3390/ijms231810506