
Light Spectral Composition Modifies Polyamine Metabolism in Young Wheat Plants

 ,
,  , , , ,
, , , ,  ,
, 

Abstract
:1. Introduction
2. Results
2.1. The Effect of Different Spectral Compositions in Combination with Polyamine Treatments on Physiological Parameters, Proline Content, and Plant Hormone Composition
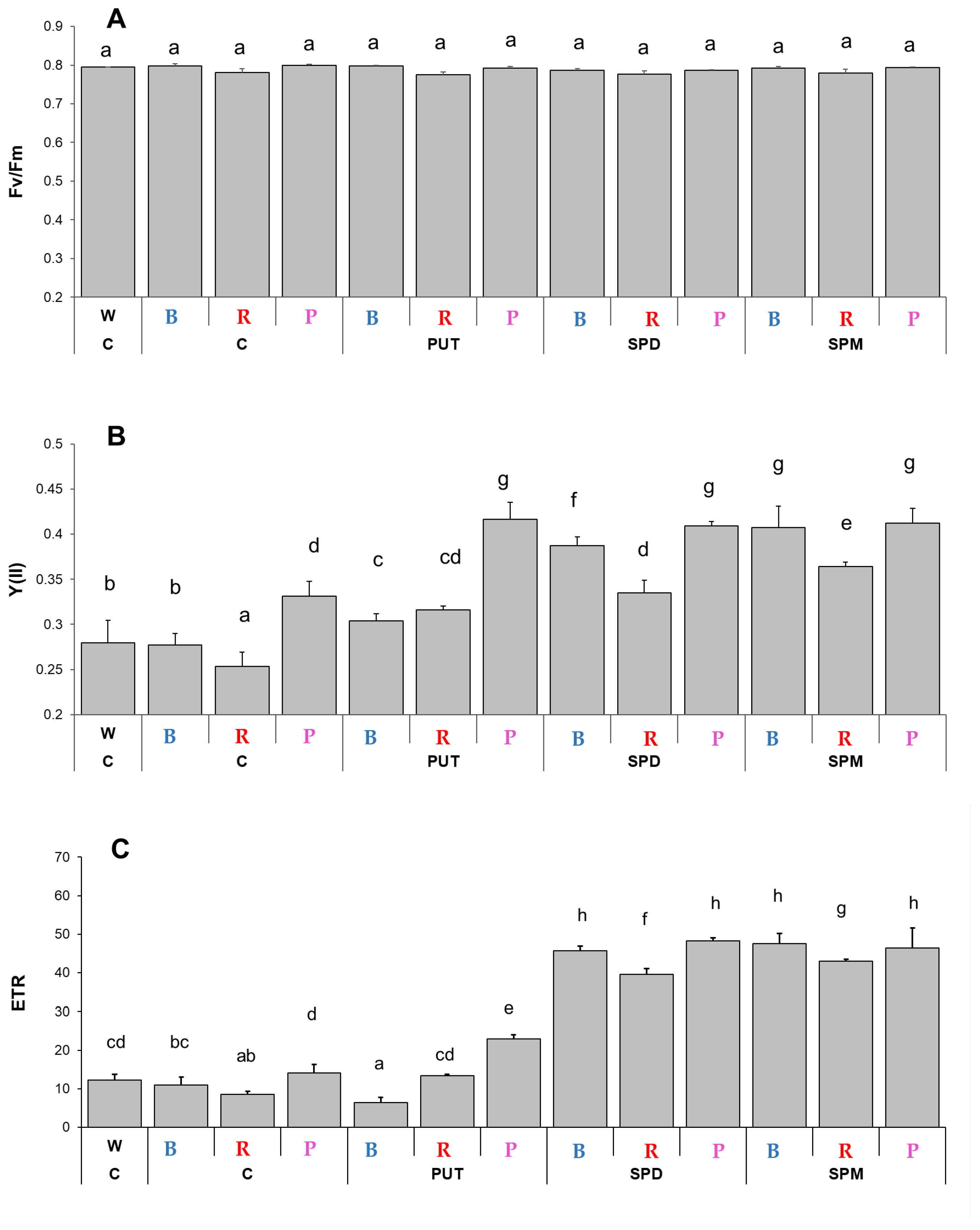
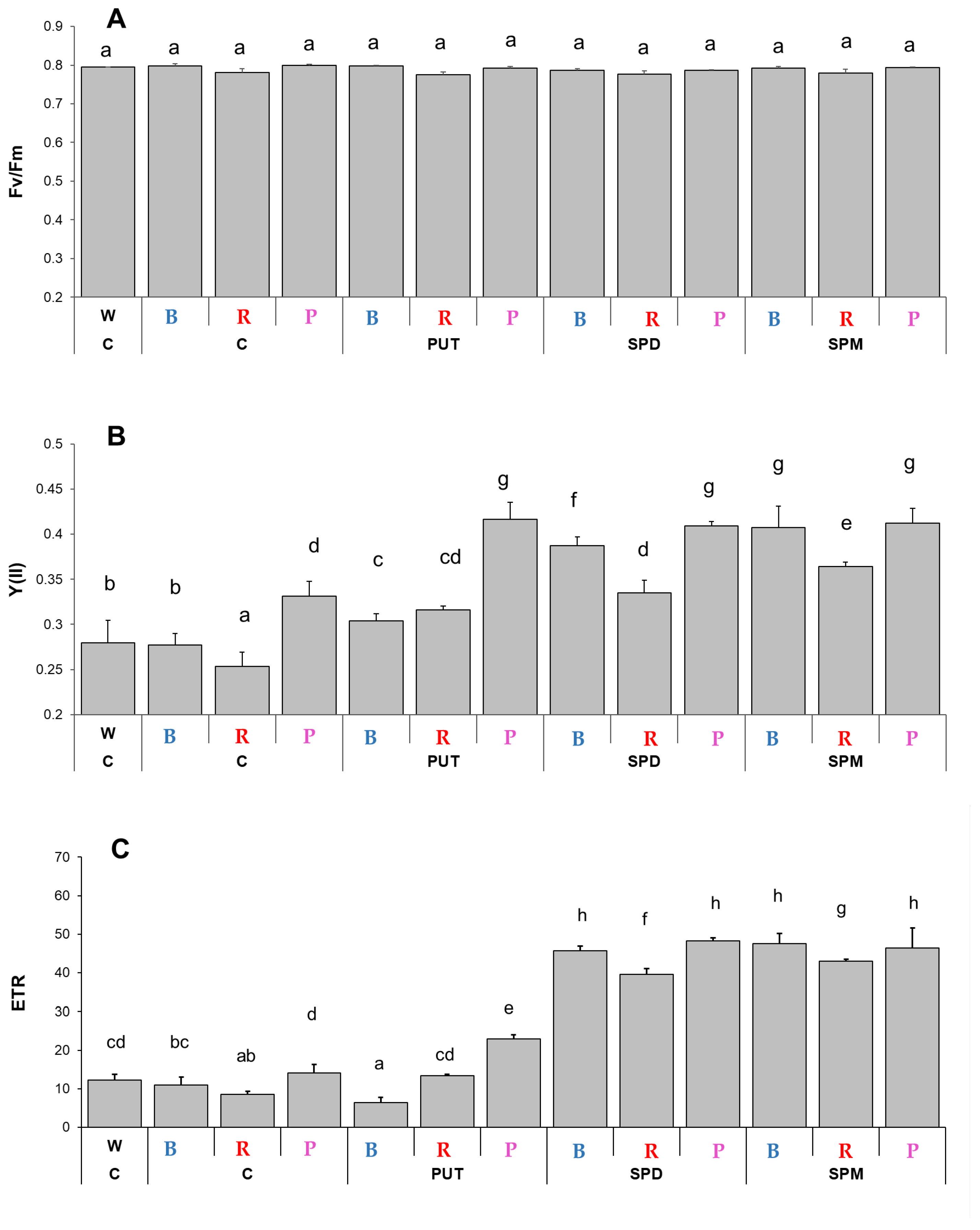
2.1.1. Changes in Chlorophyll-a Fluorescence Induction Parameters and Pigment Contents
2.1.2. Differences in Biomass Parameters
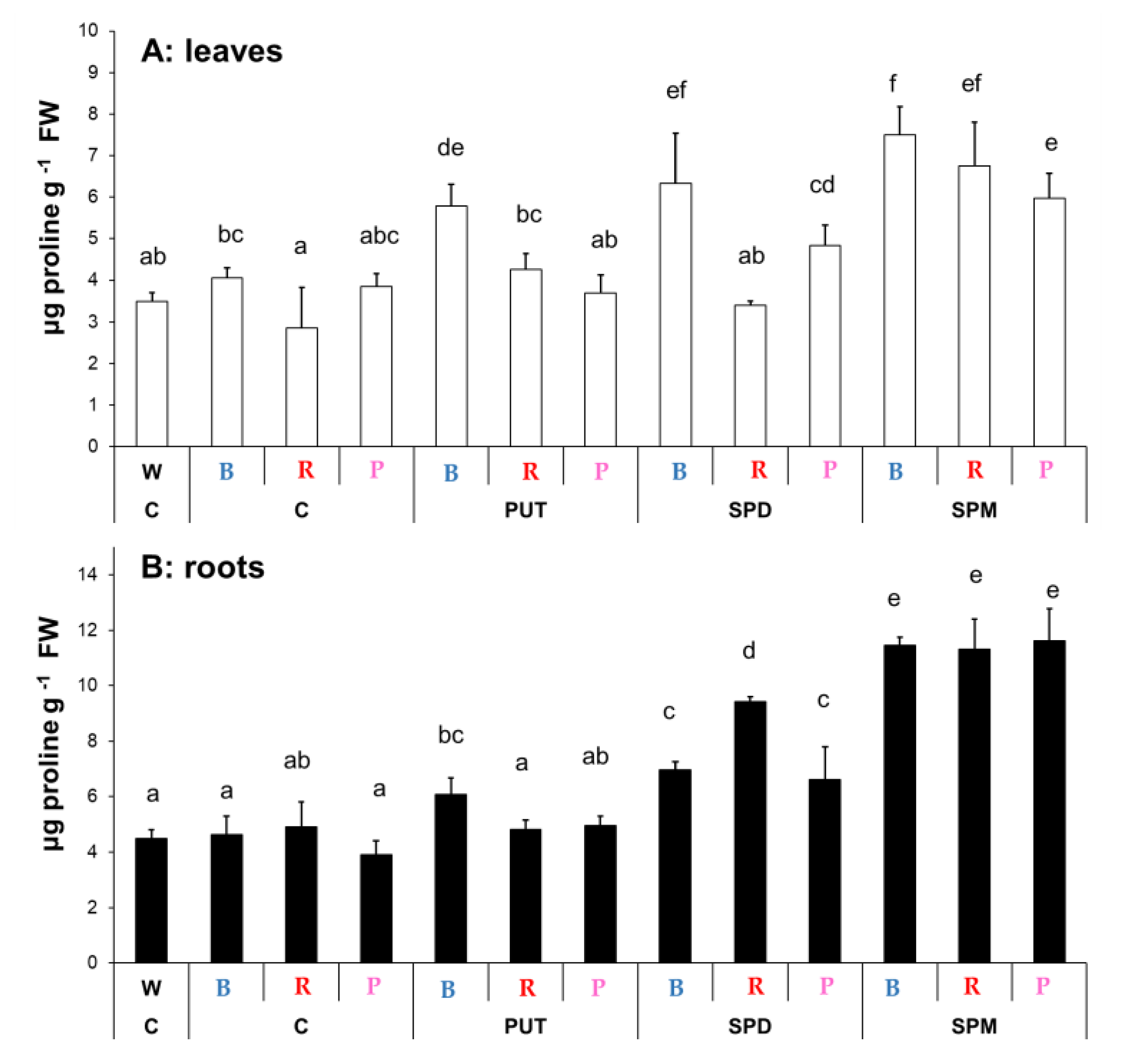
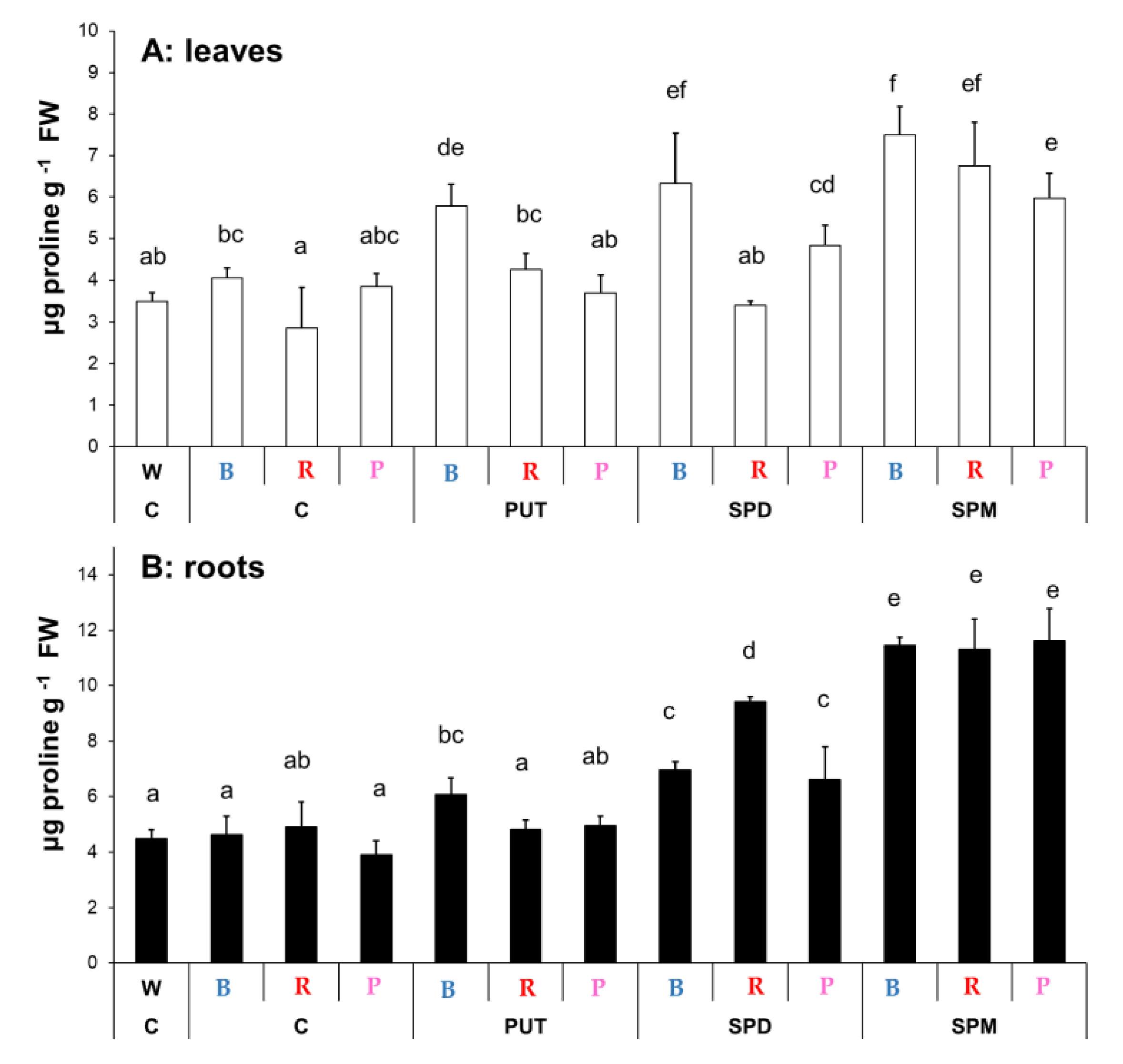
2.1.3. Changes in Proline Content
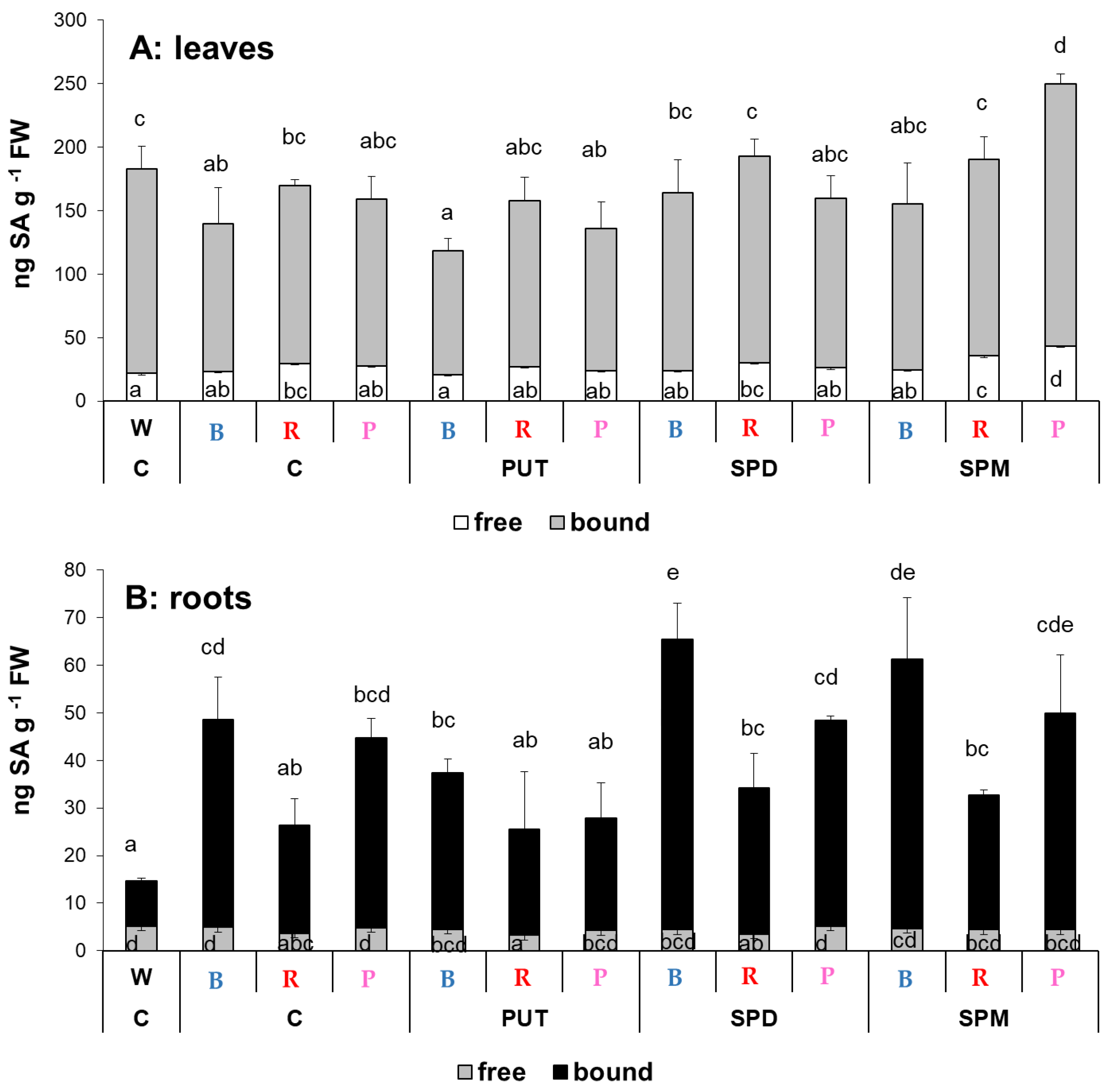
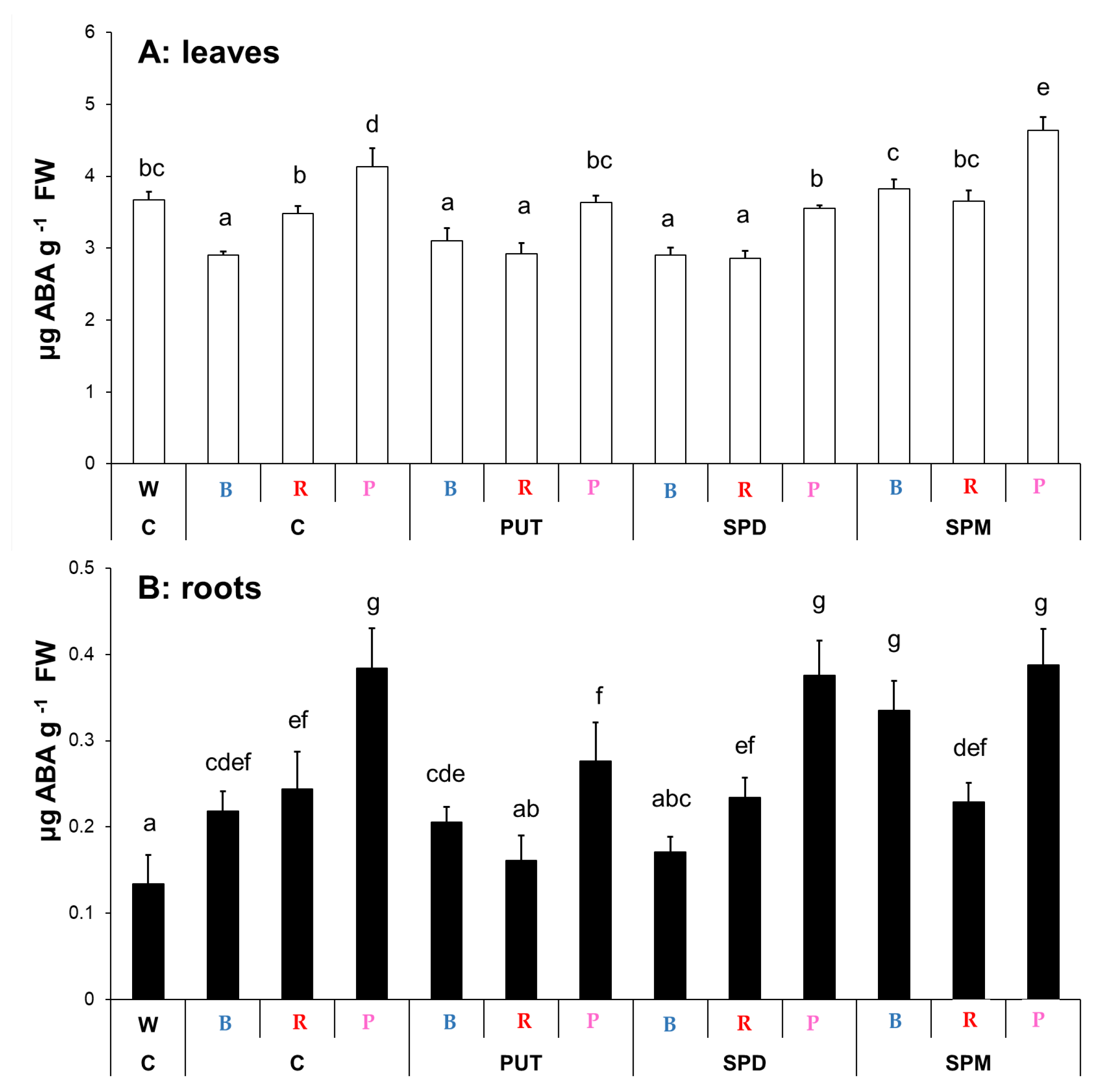
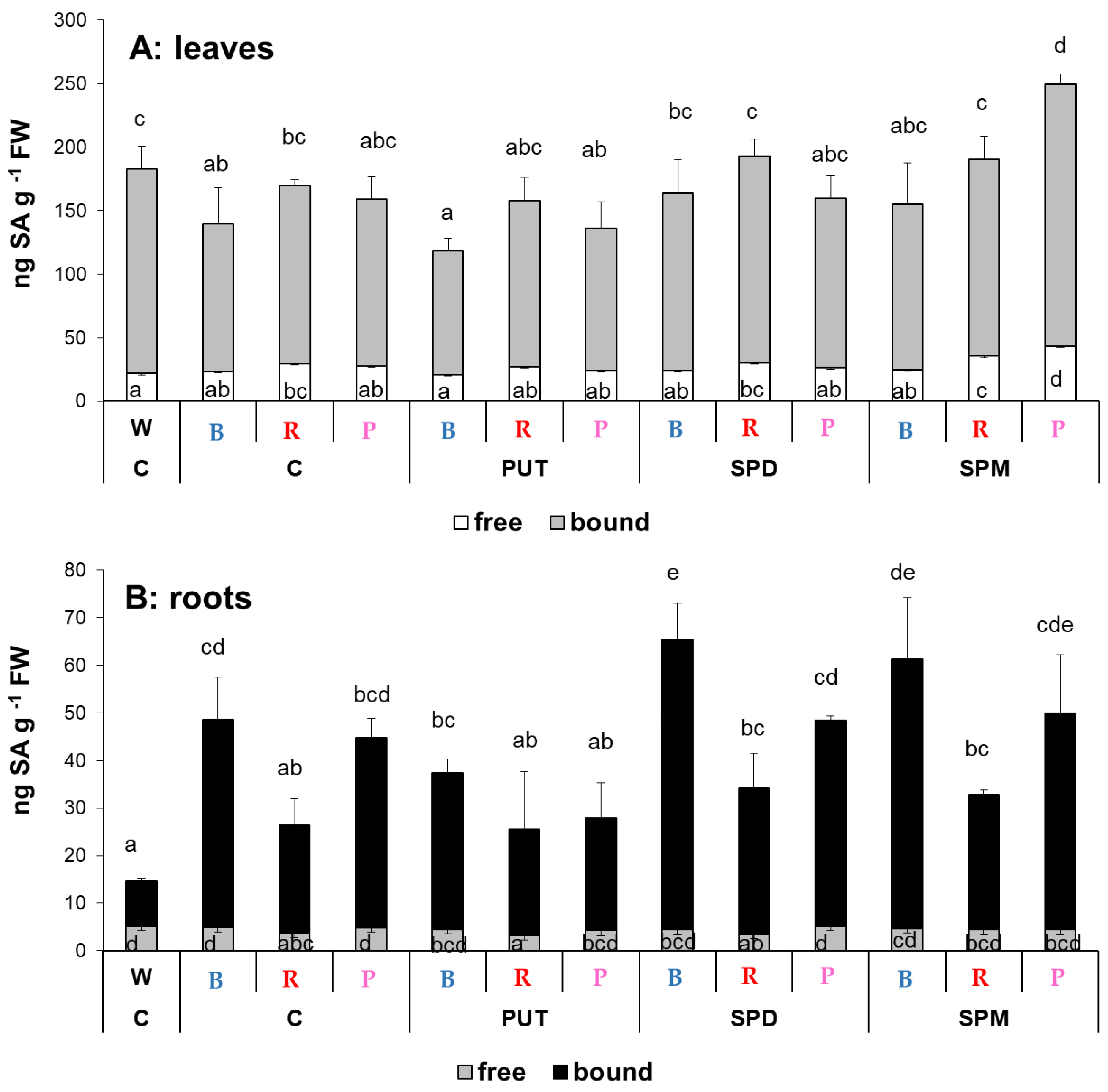
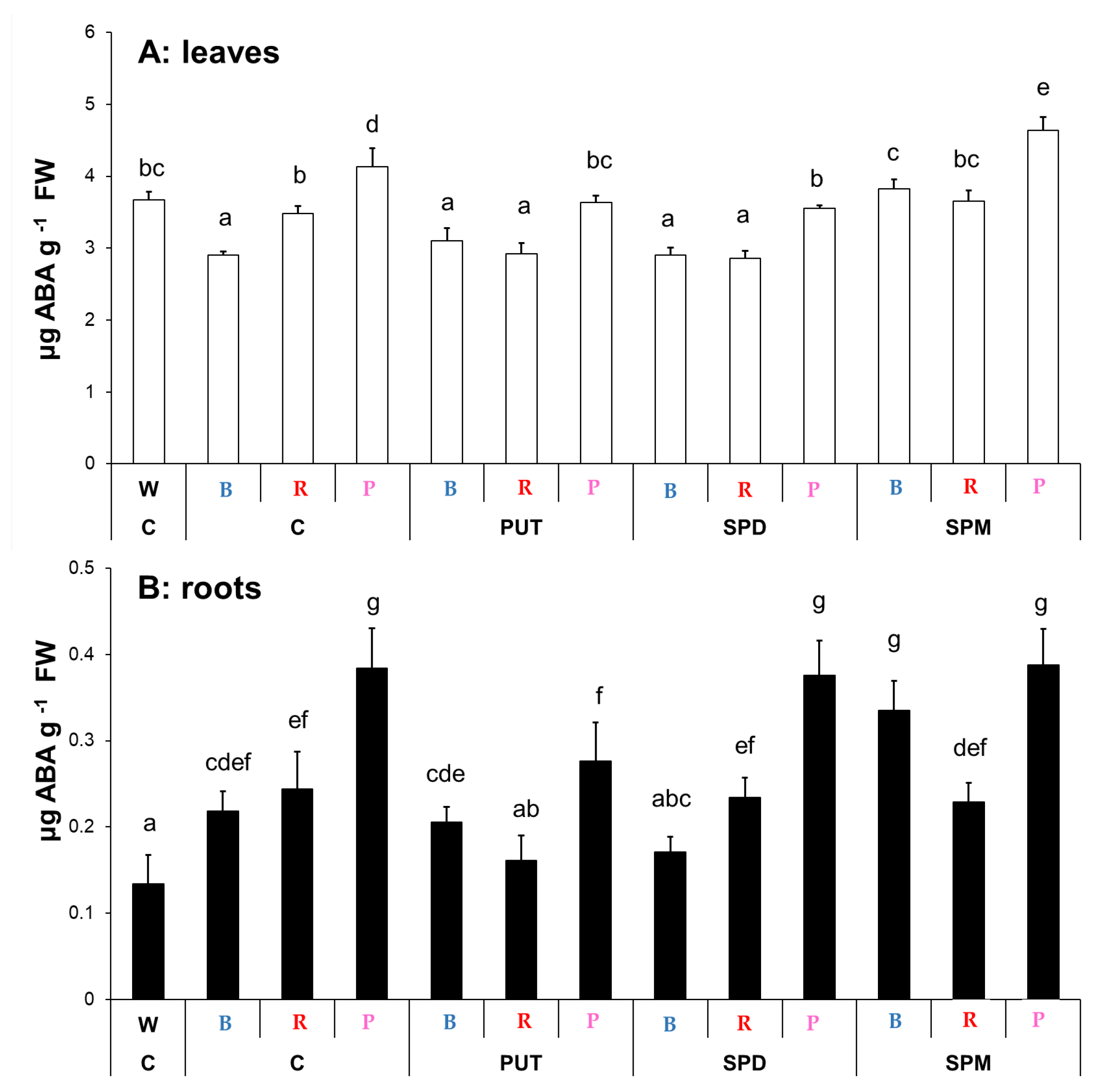
2.1.4. Changes in Salicylic Acid and Abscisic Acid Contents
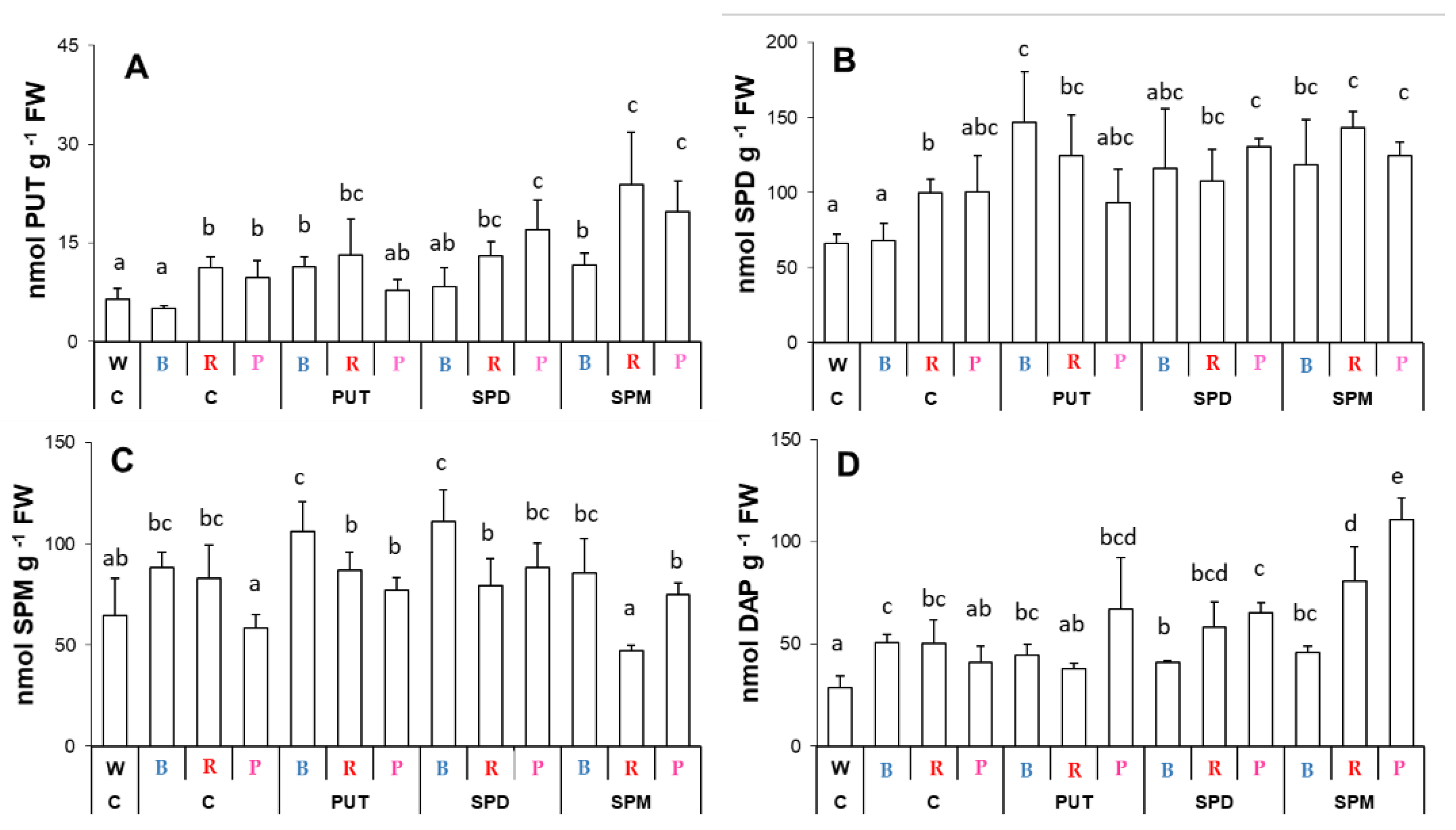
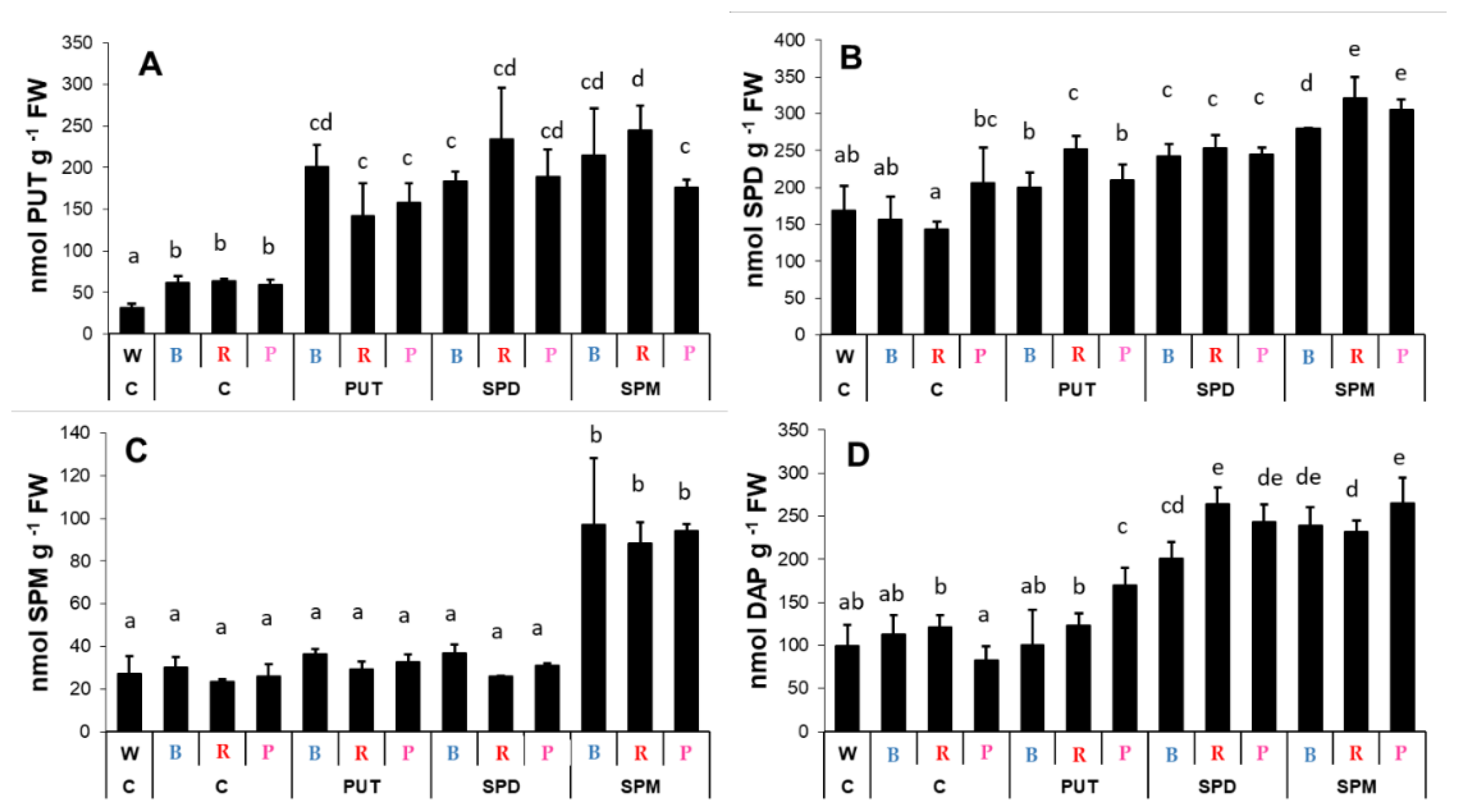
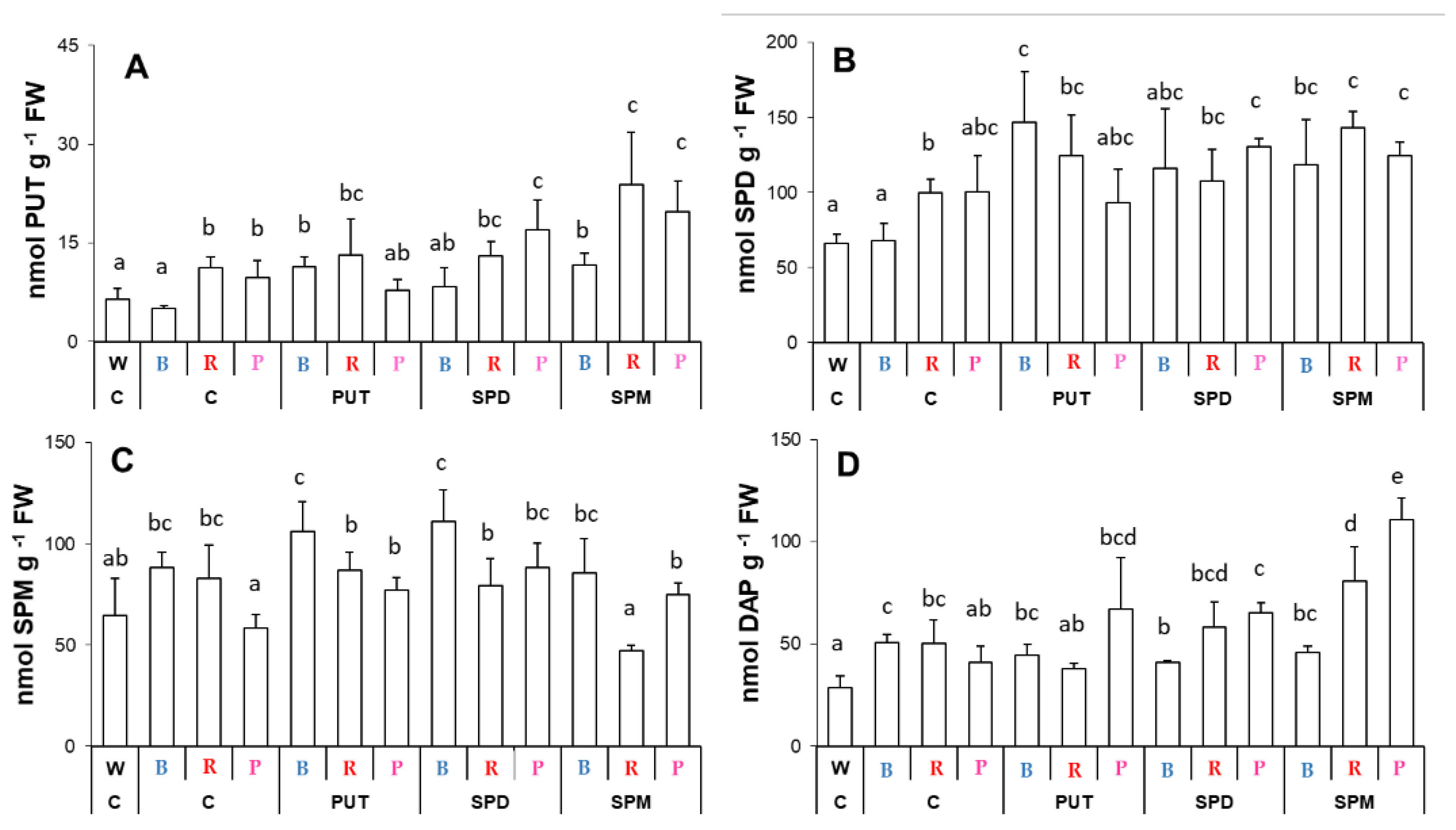
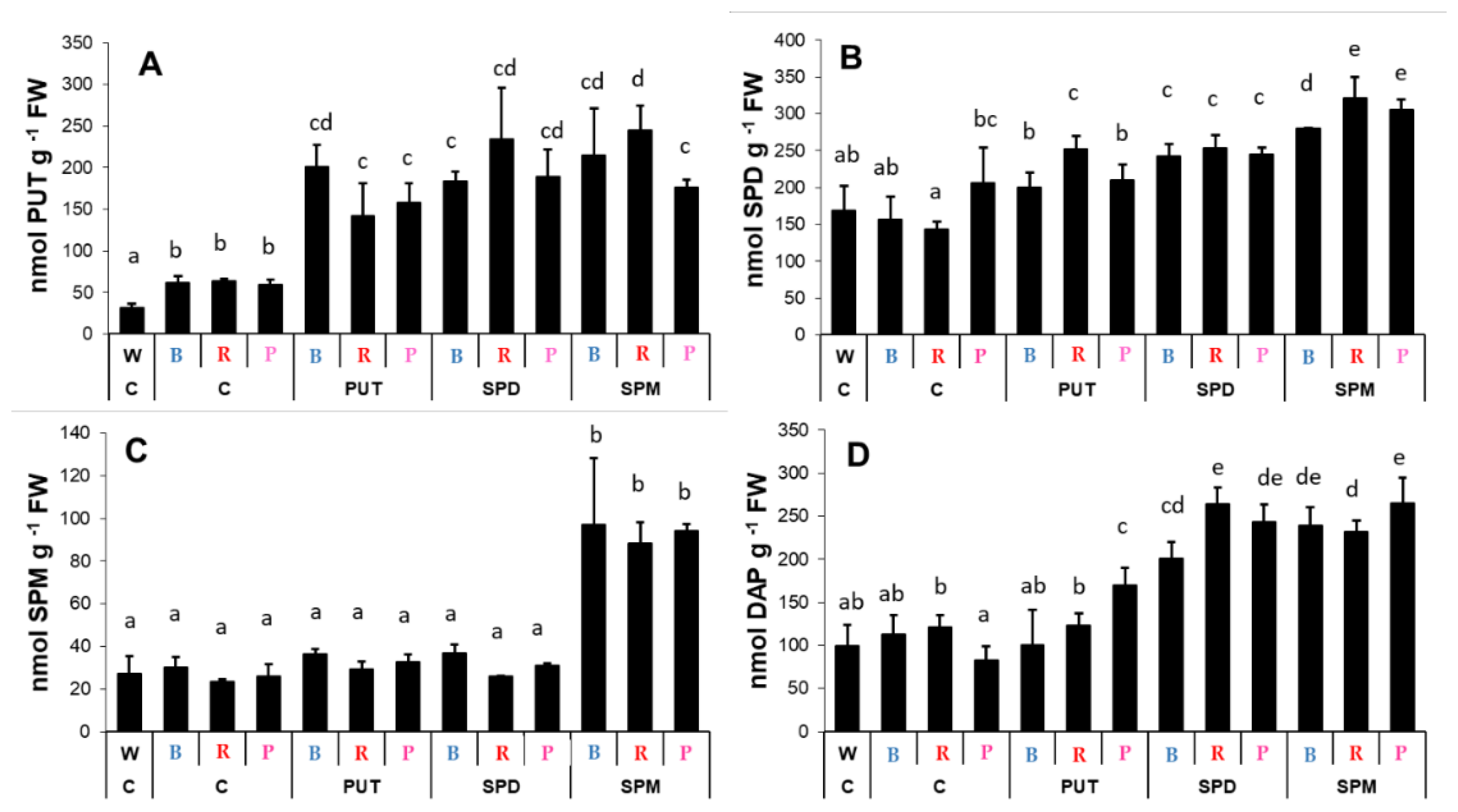
2.2. Effects of Different Spectral Conditions in Combination with Polyamine Treatments on Polyamine Metabolism
2.2.1. Changes in Polyamine Contents
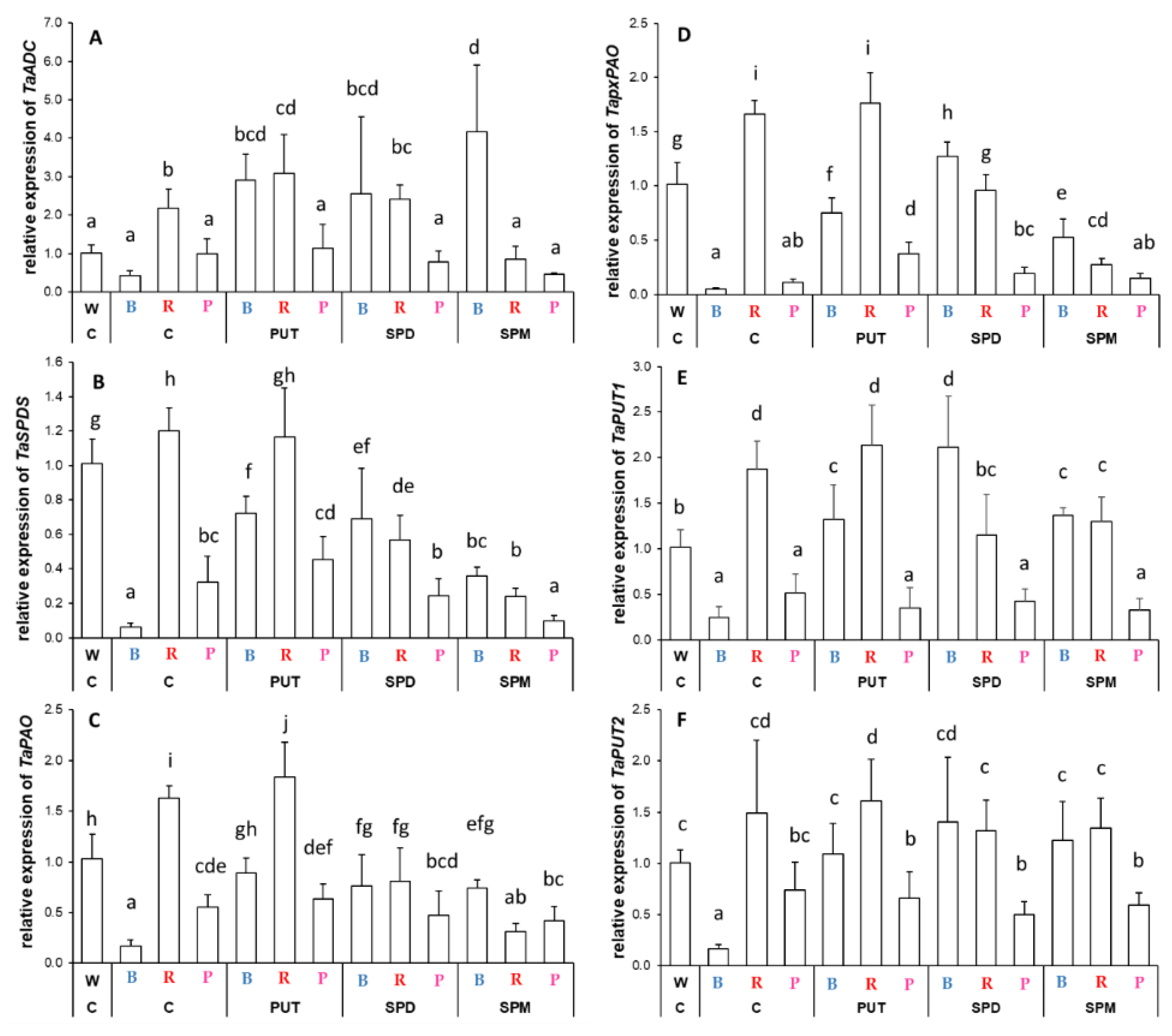
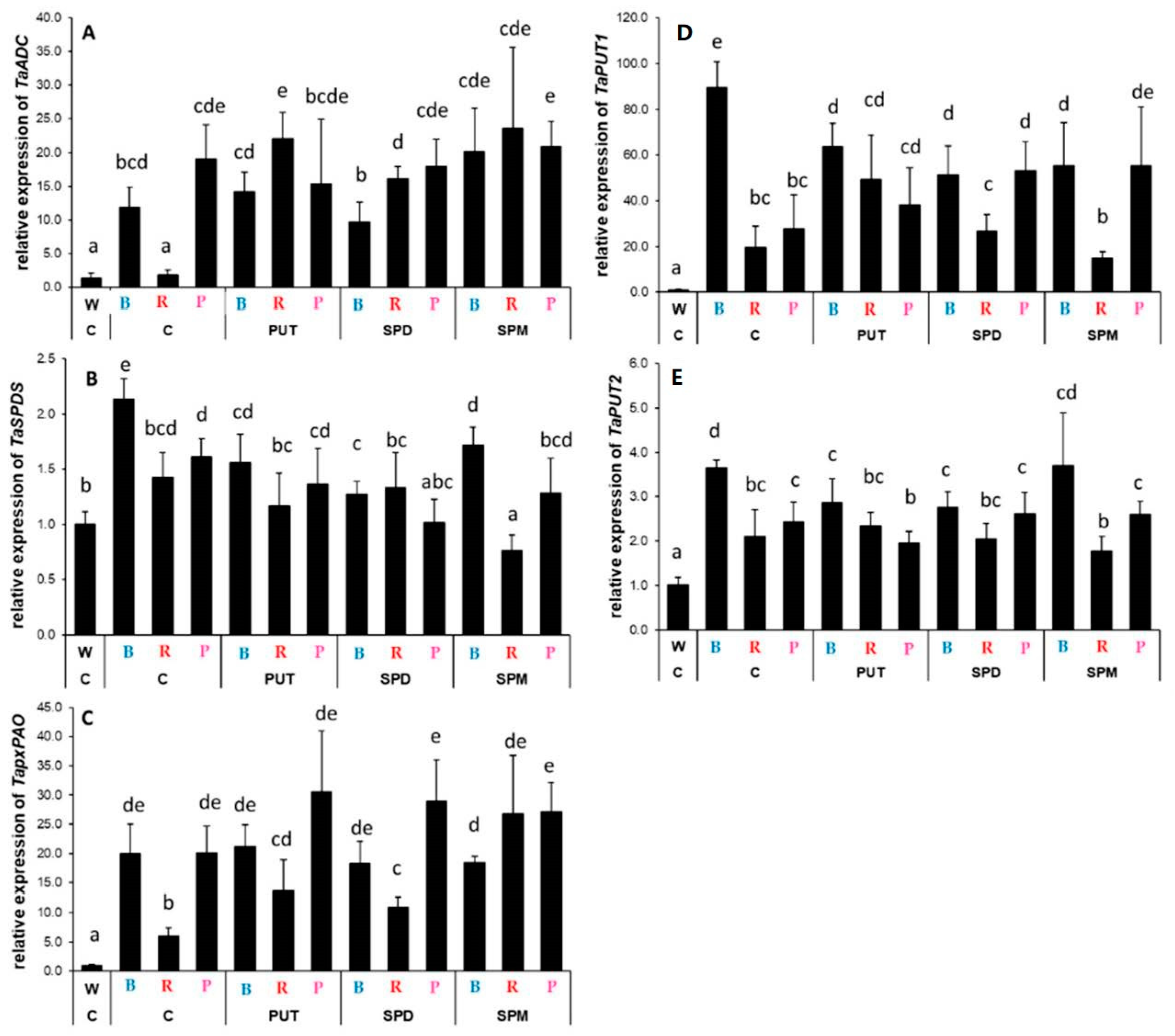
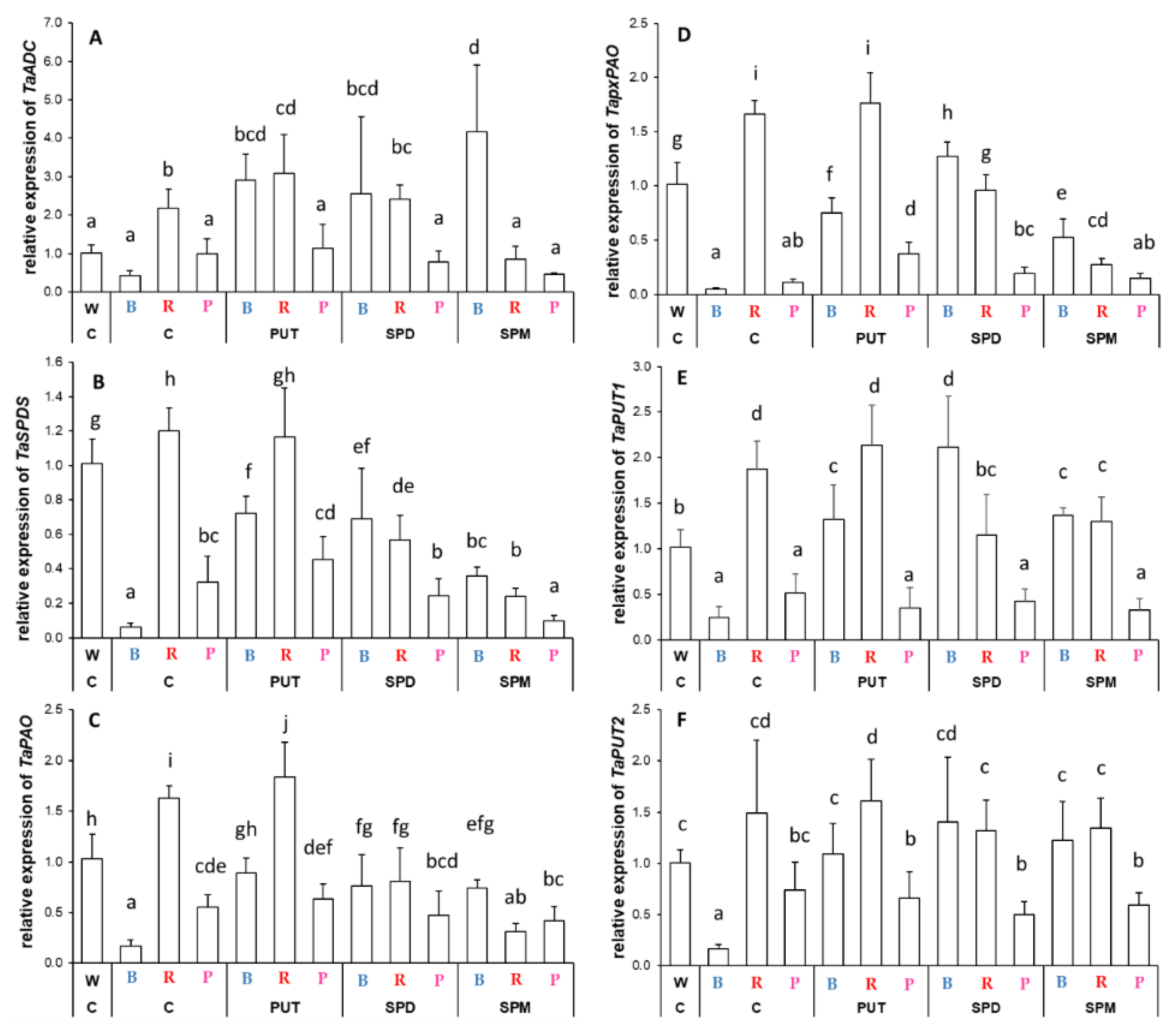
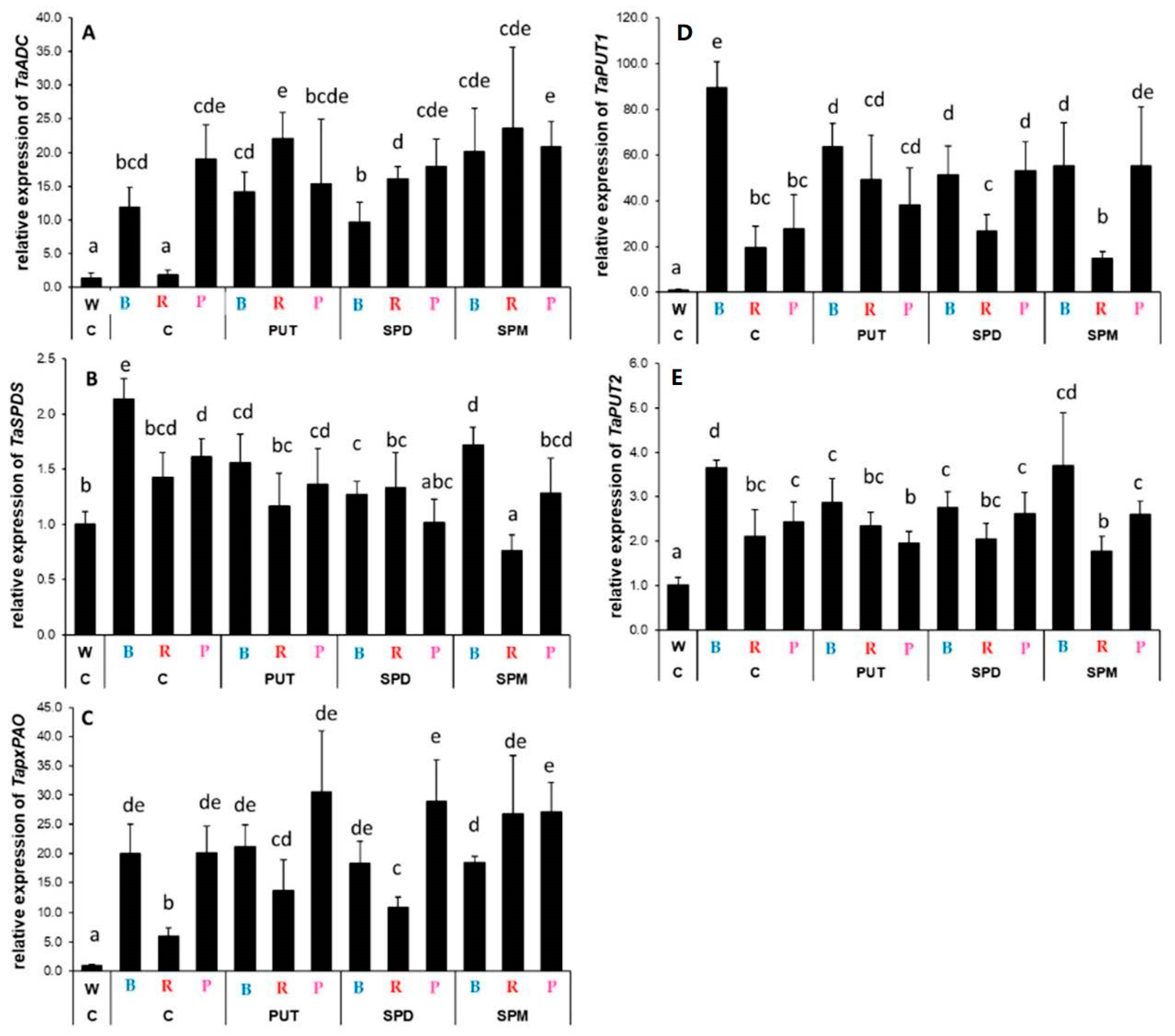
2.2.2. Differences in the Expression Levels of Genes Involved
3. Discussion
3.1. Light Quality Modifies Certain Photosynthesis- and Biomass-Related Parameters and Plant Hormone Levels
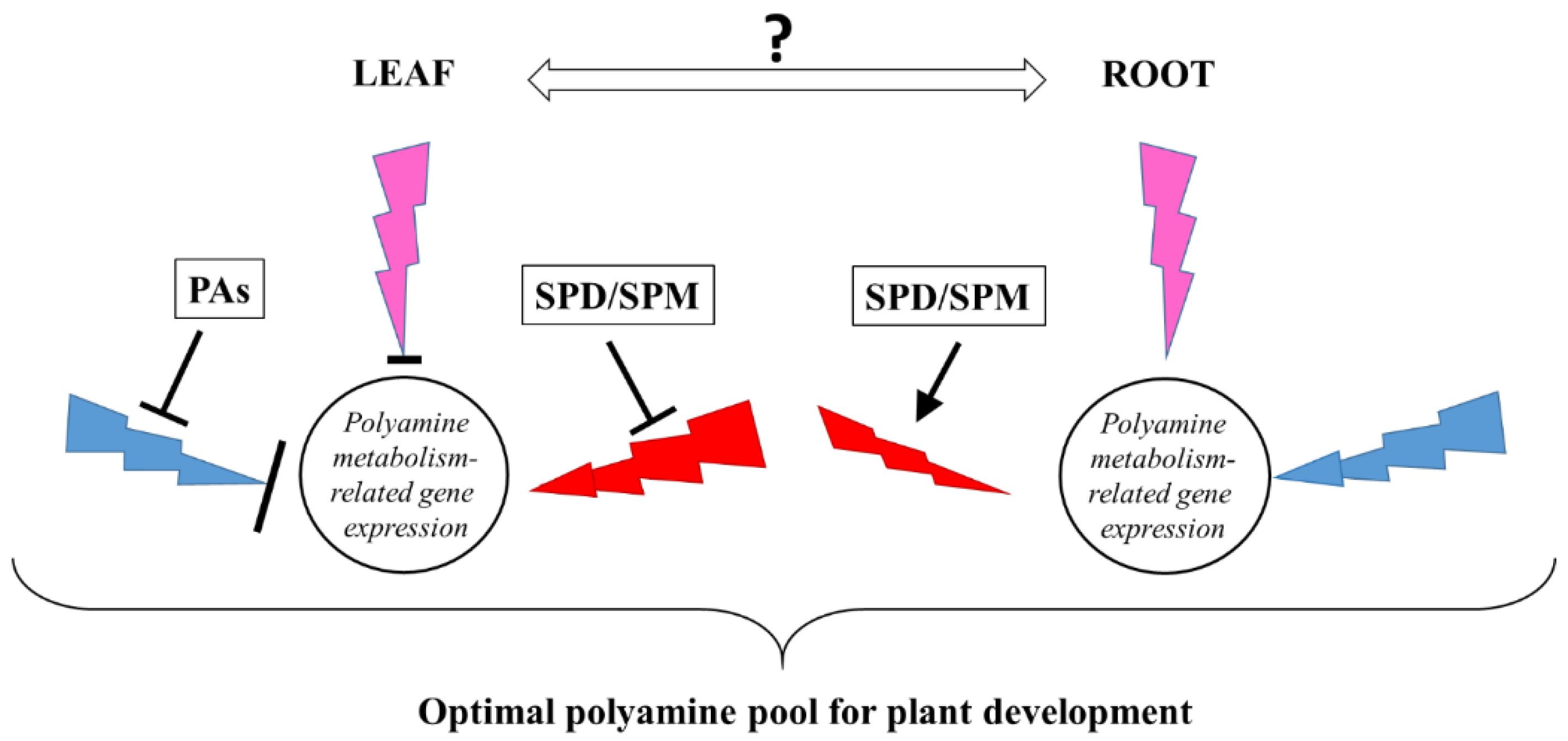
3.2. Light Quality May Influence the Expression of Genes Related to Polyamine Metabolism without Affecting the Polyamine Levels
3.3. PA Treatments Have Roborative Effects under Different Spectral Compositions and Their Effects on PA Metabolism Depend on the Light Quality
4. Materials and Methods
4.1. Plant Materials, Growth Conditions, and Treatments
4.2. Chlorophyll a Fluorescence Induction (FI) Analysis
4.3. Polyamine Analysis
4.4. Extraction of Plant Hormones and Analytical Procedure
4.5. Pigment Extraction and Analyses
4.6. Proline Measurement
4.7. Gene Expression Analysis
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Poorter, H.; Niinemets, Ü.; Ntagkas, N.; Siebenkäs, A.; Mäenpää, M.; Matsubara, S.; Pons, T. A meta-analysis of plant responses to light intensity for 70 traits ranging from molecules to whole plant performance. New Phytol. 2019, 223, 1073–1105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Z.; Li, S.; Zhang, M.; Jiang, S.; Xiao, Y. Light intensity affects growth, photosynthetic capability, and total flavonoid accumulation of Anoectochilus plants. HortScience 2010, 45, 863–867. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.Y.; Wu, K.X.; Liu, Y.; Liu, J.; Tang, Z.H. Effects of light intensity on the growth, photosynthetic characteristics, and secondary metabolites of Eleutherococcus senticosus Harms. Photosynthetica 2020, 58, 881–889. [Google Scholar] [CrossRef]
- Kitazaki, K.; Fukushima, A.; Nakabayashi, R.; Okazaki, Y.; Kobayashi, M.; Mori, T.; Nishizawa, T.; Reyes-Chin-Wo, S.; Michelmore, R.W.; Saito, K.; et al. Metabolic reprogramming in leaf lettuce grown under different light quality and intensity conditions using narrow-band LEDs. Sci. Rep. 2018, 8, 7914. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Hu, T.; Poethig, R.S. Low light intensity delays vegetative phase change. Plant Physiol. 2021, 187, 1177–1188. [Google Scholar] [CrossRef]
- Vialet-Chabrand, S.; Matthews, J.S.A.; Simkin, A.J.; Raines, C.A.; Lawson, T. Importance of fluctuations in light on plant photosynthetic acclimation. Plant Physiol. 2017, 173, 2163–2179. [Google Scholar] [CrossRef] [Green Version]
- Kurepin, L.V.; Walton, L.J.; Reid, D.M. Interaction of red to far red light ratio and ethylene in regulating stem elongation of Helianthus annuus. Plant Growth Regul. 2007, 51, 53–61. [Google Scholar] [CrossRef]
- Li, Q.; Kubota, C. Effects of supplemental light quality on growth and phytochemicals of baby leaf lettuce. Environ. Exp. Bot. 2009, 67, 59–64. [Google Scholar] [CrossRef]
- Li, H.; Tang, C.; Xu, Z.; Liu, X.; Han, X. Effects of different light sources on the growth of non-heading Chinese cabbage (Brassica campestris L.). J. Agric. Sci. 2012, 4, 262–273. [Google Scholar] [CrossRef] [Green Version]
- Dong, C.; Fu, Y.; Liu, G.; Liu, H. Growth, photosynthetic characteristics, antioxidant capacity and biomass yield and quality of wheat (Triticum aestivum L.) exposed to LED light sources with different spectra combinations. J. Agron. Crop Sci. 2014, 200, 219–230. [Google Scholar] [CrossRef]
- Gautam, P.; Terfa, M.T.; Olsen, J.E.; Torre, S. Red and blue light effects on morphology and flowering of Petunia × hybrida. Sci. Hortic.-Amst. 2015, 184, 171–178. [Google Scholar] [CrossRef]
- Rehman, M.; Ullah, S.; Bao, Y.; Wang, B.; Peng, D.; Liu, L. Light-emitting diodes: Whether an efficient source of light for indoor plants? Environ. Sci. Pollut. Res. 2017, 24, 24743–24752. [Google Scholar] [CrossRef] [PubMed]
- Kalaitzoglou, P.; Taylor, C.; Calders, K.; Hogervorst, M.; van Ieperen, W.; Harbinson, J.; de Visser, P.; Nicole, C.C.S.; Marcelis, L.F.M. Unraveling the effects of blue light in an artificial solar background light on growth of tomato plants. Environ. Exp. Bot. 2021, 184, 104377. [Google Scholar] [CrossRef]
- Johkan, M.; Shoji, K.; Goto, F.; Hashida, S.; Yoshihara, T. Blue light-emitting diode light irradiation of seedlings improves seedling quality and growth after transplanting in red leaf lettuce. HortScience 2010, 45, 1809–1814. [Google Scholar] [CrossRef] [Green Version]
- Ramalho, J.C.; Marques, N.C.; Semedo, J.N.; Matos, M.C.; Quartin, V.L. Photosynthetic performance and pigment composition of leaves from two tropical species is determined by light quality. Plant Biol. 2002, 4, 112–120. [Google Scholar] [CrossRef]
- Inoue, S.I.; Kinoshita, T. Blue light regulation of stomatal opening and the plasma membrane H+-ATPase. Plant Physiol. 2017, 174, 531–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savvides, A.; Fanourakis, D.; van Ieperen, W. Co-ordination of hydraulic and stomatal conductances across light qualities in cucumber leaves. J. Exp. Bot. 2012, 63, 1135–1143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muneer, S.; Kim, E.J.; Park, J.S.; Lee, J.H. Influence of green, red and blue light emitting diodes on multiprotein complex proteins and photosynthetic activity under different light intensities in lettuce leaves (Lactuca sativa L.). Int. J. Mol. Sci. 2014, 15, 4657–4670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Xu, H.; Shao, L.; Wang, R.; Li, T.; Wang, Y. Response of photosynthetic capacity of tomato leaves to different LED light wavelength. Environ. Exp. Bot. 2018, 150, 161–171. [Google Scholar] [CrossRef]
- Sellaro, R.; Hoecker, U.; Yanovsky, M.; Chory, J.; Casal, J.J. Synergism of red and blue light in the control of Arabidopsis gene expression and development. Curr. Biol. 2009, 19, 1216–1220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hosotani, S.; Yamauchi, S.; Kobayashi, H.; Fuji, S.A.; Koya, S.; Shimazaki, K.; Takemiya, A. A BLUS1 kinase signal and a decrease in intercellular CO2 concentration are necessary for stomatal opening in response to blue light. Plant Cell 2021, 33, 1813–1827. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Xin, G.; Liu, C.; Shi, Q.; Yang, F.; Wei, M. Effects of red and blue light on leaf anatomy, CO2 assimilation and the photosynthetic electron transport capacity of sweet pepper (Capsicum annuum L.) seedlings. BMC Plant Biol. 2020, 20, 318. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, E.; Ouzounis, T.; Giday, H.; Schipper, R.; Heuvelink, E.; Marcelis, L.F.M. Adding blue to red supplemental light increases biomass and yield of greenhouse-grown tomatoes, but only to an optimum. Front. Plant Sci. 2019, 9, 2002. [Google Scholar] [CrossRef] [Green Version]
- Naznin, M.T.; Lefsrud, M.; Azad, M.O.K.; Park, C.H. Effect of different combinations of red and blue LED light on growth characteristics and pigment content of in vitro tomato plantlets. Agriculture 2019, 9, 196. [Google Scholar] [CrossRef] [Green Version]
- Thoma, F.; Somborn-Schulz, A.; Schlehuber, D.; Keuter, V.; Deerberg, G. Effects of light on secondary metabolites in selected leafy greens: A review. Front. Plant Sci. 2020, 11, 497. [Google Scholar] [PubMed]
- Yavari, N.; Tripathi, R.; Wu, B.S.; MacPherson, S.; Singh, J.; Lefsrud, M. The effect of light quality on plant physiology, photosynthetic, and stress response in Arabidopsis thaliana leaves. PLoS ONE 2021, 16, e0247380. [Google Scholar] [CrossRef]
- Bayat, L.; Arab, M.; Aliniaeifard, S.; Seif, M.; Lastochkina, O.; Li, T. Effects of growth under different light spectra on the subsequent high light tolerance in rose plants. AoB Plants 2018, 10, ply052. [Google Scholar] [CrossRef] [PubMed]
- Landi, M.; Zivcak, M.; Sytar, O.; Brestic, M.; Allakhverdiev, S.I. Plasticity of photosynthetic processes and the accumulation of secondary metabolites in plants in response to monochromatic light environments: A review. Biochim. Biophys. Acta Bioenerg. 2020, 1861, 148131. [Google Scholar] [CrossRef]
- Samuoliene, G.; Brazaityte, A.; Sirtautas, R.; Novickovas, A.; Duchovskis, P. Supplementary red-LED lighting affects phytochemicals and nitrate of baby leaf lettuce. J. Food Agric. Environ. 2011, 9, 271–274. [Google Scholar]
- Samuoliene, G.; Brazaityte, A.; Sirtautas, R.; Novièkovas, A.; Duchovskis, P. The effect of supplementary LED lighting on the antioxidant and nutritional properties of lettuce. Acta Hortic. 2012, 952, 835–842. [Google Scholar] [CrossRef]
- Kim, K.; Kook, H.S.; Jang, Y.J.; Lee, W.H.; Seralathan, K.K.; Chae, J.C.; Lee, K.J. The effect of blue-light-emitting diodes on antioxidant properties and resistance to Botrytis cinerea in tomato. J. Plant Pathol. Microbiol. 2013, 4, 203. [Google Scholar]
- Monostori, I.; Heilmann, M.; Kocsy, G.; Rakszegi, M.; Ahres, M.; Altenbach, S.B.; Szalai, G.; Pál, M.; Toldi, D.; Simon-Sarkadi, L.; et al. LED Lighting—Modification of growth, metabolism, yield and flour composition in wheat by spectral quality and intensity. Front. Plant Sci. 2018, 9, 605. [Google Scholar] [CrossRef] [PubMed]
- Toldi, D.; Gyugos, M.; Darkó, É.; Szalai, G.; Gulyás, Z.; Gierczik, K.; Székely, A.; Boldizsár, Á.; Galiba, G.; Müller, M.; et al. Light intensity and spectrum affect metabolism of glutathione and amino acids at transcriptional level. PLoS ONE 2019, 14, e0227271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gyugos, M.; Ahres, M.; Gulyás, Z.; Szalai, G.; Darkó, É.; Mednyánszky, Z.; Dey, N.; Kar, R.K.; Simon-Sarkadi, L.; Kocsy, G. Light spectrum modifies the drought-induced changes of glutathione and free amino acid levels in wheat. Acta Physiol. Plant. 2021, 43, 90. [Google Scholar] [CrossRef]
- González-Hernández, A.I.; Scalschi, L.; Vicedo, B.; Marcos-Barbero, E.L.; Morcuende, R.; Camañes, G. Putrescine: A key metabolite involved in plant development, tolerance and resistance responses to stress. Int. J. Mol. Sci. 2022, 23, 2971. [Google Scholar] [CrossRef] [PubMed]
- Kusano, T.; Berberich, T.; Tateda, C.; Takahashi, Y. Polyamines: Essential factors for growth and survival. Planta 2008, 228, 367–381. [Google Scholar] [CrossRef] [PubMed]
- Kotzabasis, K.; Fotinou, C.; Roubelakis-Angelakis, K.A.; Ghanotakis, D. Polyamines in the photosynthetic apparatus—Photosystem II highly resolved subcomplexes are enriched in spermine. Photosynth. Res. 1993, 38, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Demetriou, G.; Neonaki, C.; Navakoudis, E.; Kotzabasis, K. Salt stress impact on the molecular structure and function of the photosynthetic apparatus—The protective role of polyamines. BBA Bioenerg. 2007, 1767, 272–280. [Google Scholar]
- Del Duca, S.; Serafini-Fracassini, D.; Cai, G. Senescence and programmed cell death in plants: Polyamine action mediated by transglutaminase. Front. Plant Sci. 2014, 5, 120. [Google Scholar] [CrossRef] [Green Version]
- Zhong, M.; Wang, Y.; Zhang, Y.; Shu, S.; Sun, J.; Guo, S. Overexpression of transglutaminase from cucumber in tobacco increases salt tolerance through regulation of photosynthesis. Int. J. Mol. Sci. 2019, 20, 894. [Google Scholar] [CrossRef] [Green Version]
- Ioannidis, N.E.; Kotzabasis, K. Effects of polyamines on the functionality of photosynthetic membrane in vivo and in vitro. BBA Bioenerg. 2007, 1767, 1372–1382. [Google Scholar] [CrossRef] [Green Version]
- Voigt, J.; Deinert, B.; Bohley, P. Subcellular localization and light-dark control of ornithine decarboxylase in the unicellular green alga Chlamydomonas reinhardtii. Physiol. Plant 2000, 108, 353–360. [Google Scholar] [CrossRef]
- Takács, Z.; Poór, P.; Tari, I. Comparison of polyamine metabolism in tomato plants exposed to different concentrations of salicylic acid under light or dark conditions. Plant Physiol. Biochem. 2016, 108, 266–278. [Google Scholar] [CrossRef] [PubMed]
- Laurenzi, M.; Rea, G.; Federico, R.; Tavladoraki, P.; Angelini, R. De-etiolation causes a phytochrome-mediated increase of polyamine oxidase expression in outer tissues of the maize mesocotyl: A role in the photomodulation of growth and cell wall differentiation. Planta 1999, 208, 146–154. [Google Scholar] [CrossRef]
- Lv, Y.; Shao, G.; Jiao, G.; Sheng, Z.; Xie, L.; Hu, S.; Tang, S.; Wei, X.; Hu, P. Targeted mutagenesis of POLYAMINE OXIDASE 5 that negatively regulates mesocotyl elongation enables the generation of direct-seeding rice with improved grain yield. Mol. Plant 2021, 14, 344–351. [Google Scholar] [CrossRef] [PubMed]
- Shu, S.; Guo, S.R.; Yuan, L. A review: Polyamines and photosynthesis. In Advances in Photosynthesis—Fundamental Aspects; Najafpour, M., Ed.; InTech: Rijeka, Croatia, 2012; pp. 439–464. [Google Scholar]
- Fan, X.X.; Xue, F.; Song, B.; Chen, L.Z.; Xu, G.; Xu, H. Effects of blue and red light on growth and nitrate metabolism in Pakchoi. Open Chem. J. 2019, 17, 456–464. [Google Scholar] [CrossRef]
- Oliveira, T.D.R.; Aragão, V.P.M.; Moharana, K.C.; Fedosejevs, E.; do Amaral, F.P.; Sousa, K.R.; Thelen, J.J.; Venâncio, T.M.; Silveira, V.; Santa-Catarina, C. Light spectra affect the in vitro shoot development of Cedrela fissilis Vell. (Meliaceae) by changing the protein profile and polyamine contents. Biochim. Biophys. Acta Proteins Proteom. 2020, 1868, 140529. [Google Scholar] [CrossRef] [PubMed]
- Lerin, J.; Aragão, V.P.M.; Reis, R.S.; Silveira, V.; Santa-Catarina, C. Proteomic profile and polyamine contents are modulated by light source to promote in vitro shoot development in Cariniana legalis (Martius) O. Kuntze (Lecythidaceae). Plant Cell Tiss. Organ Cult. 2019, 137, 329–342. [Google Scholar] [CrossRef]
- Hunter, D.C.; Burritt, D.J. Light quality influences the polyamine content of lettuce (Lactuca sativa L.) cotyledon explants during shoot production in vitro. Plant Growth Regul. 2005, 45, 53–61. [Google Scholar] [CrossRef]
- Yu, Y.; Qin, W.; Li, Y.; Zhang, C.; Wang, Y.; Yang, Z.; Ge, X.; Li, F. Red light promotes cotton embryogenic callus formation by influencing endogenous hormones, polyamines and antioxidative enzyme activities. Plant Growth Regul. 2019, 87, 187–199. [Google Scholar] [CrossRef]
- Chen, D.; Shao, Q.; Yin, L.; Younis, A.; Zheng, B. Polyamine function in plants: Metabolism, regulation on development, and roles in abiotic stress responses. Front. Plant Sci. 2019, 9, 1945. [Google Scholar] [CrossRef]
- Pál, M.; Szalai, G.; Gondor, O.K.; Janda, T. Unfinished story of polyamines: Role of conjugation, transport and light-related regulation in the polyamine metabolism in plants. Plant Sci. 2021, 308, 110923. [Google Scholar] [CrossRef] [PubMed]
- Kramer, G.F.; Krizek, D.T.; Roman, M.; Mirecki, R.M. Influence of photosynthetically active radiation and spectral quality on UV-B-induced polyamine accumulation in soybean. Phytochemistry 1992, 31, 1119–1125. [Google Scholar] [CrossRef]
- Yoshida, I.; Yamagata, H.; Hirasawa, E. Blue- and red-light regulation and circadian control of gene expression of S-adenosylmethionine decarboxylase in Pharbitis nil. J. Exp. Bot. 1999, 50, 319–326. [Google Scholar]
- Kim, W.; Zeljković, S.Ć.; Piskurewicz, U.; Megies, C.; Tarkowski, P.; Lopez-Molina, L. Polyamine uptake transporter 2 (put2) and decaying seeds enhance phyA-mediated germination by overcoming PIF1 repression of germination. PLoS Genet. 2019, 15, e1008292. [Google Scholar] [CrossRef] [PubMed]
- Gondor, O.K.; Tajti, J.; Hamow, K.Á.; Majláth, I.; Szalai, G.; Janda, T.; Pál, M. Polyamine metabolism under different light regimes in wheat. Int. J. Mol. Sci. 2021, 22, 11717. [Google Scholar] [CrossRef]
- Klem, K.; Gargallo-Garriga, A.; Rattanapichai, W.; Oravec, M.; Holub, P.; Veselá, B.; Sardans, J.; Peñuelas, J.; Urban, O. Distinct morphological, physiological, and biochemical responses to light quality in barley leaves and roots. Front. Plant Sci. 2019, 10, 1026. [Google Scholar] [CrossRef] [Green Version]
- Tan, T.; Li, S.; Fan, Y.; Wang, Z.; Raza, M.A.; Shafiq, I.; Wang, B.; Wu, X.; Yong, T.; Wang, X.; et al. Far-red light: A regulator of plant morphology and photosynthetic capacity. Crop J. 2022, 10, 300–309. [Google Scholar] [CrossRef]
- Tajti, J.; Janda, T.; Majláth, I.; Szalai, G.; Pál, M. Comparative study on the effects of putrescine and spermidine pre-treatment on cadmium stress in wheat. Ecotoxicol. Environ. Saf. 2018, 148, 546–554. [Google Scholar] [CrossRef] [Green Version]
- Szalai, G.; Janda, K.; Darkó, É.; Janda, T.; Peeva, V.; Pál, M. Comparative analysis of polyamine metabolism in wheat and maize plants. Plant Physiol. Biochem. 2017, 112, 239–250. [Google Scholar] [CrossRef] [Green Version]
- Pál, M.; Csávás, G.; Szalai, G.; Oláh, T.; Khalil, R.; Yordanova, R.; Gell, G.; Birinyi, Z.; Németh, E.; Janda, T. Polyamines may influence phytochelatin synthesis during Cd stress in rice. J. Hazard. Mater. 2017, 340, 272–280. [Google Scholar] [CrossRef] [Green Version]
- Pál, M.; Ivanovska, B.; Oláh, T.; Tajti, J.; Hamow, K.Á.; Szalai, G.; Khalil, R.; Vanková, R.; Dobrev, P.; Misheva, S.P.; et al. Role of polyamines in plant growth regulation of Rht wheat mutants. Plant Physiol. Biochem. 2019, 137, 189–202. [Google Scholar] [CrossRef] [PubMed]
- Pál, M.; Tajti, J.; Szalai, G.; Peeva, V.; Végh, B.; Janda, T. Interaction of polyamines, abscisic acid and proline under osmotic stress in the leaves of wheat plants. Sci. Rep. 2018, 8, 12839. [Google Scholar] [CrossRef] [PubMed]
- Szalai, G.; Tajti, J.; Hamow, K.Á.; Ildikó, D.; Khalil, R.; Vanková, R.; Dobrev, P.; Misheva, S.P.; Janda, T.; Pál, M. Molecular background of cadmium tolerance in Rht dwarf wheat mutant is related to a metabolic shift from proline and polyamine to phytochelatin synthesis. Environ. Sci. Pollut. Res. 2020, 27, 23664–23676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bukhov, N.G.; Drozdova, I.S.; Bondar, V.V.; Mokronosov, A.T. Blue, red and blue plus red light control of chlorophyll content and CO2 gas exchange in barley leaves: Quantitative description of the effects of light quality and fluence rate. Physiol. Plant. 1992, 85, 632–638. [Google Scholar] [CrossRef]
- Makowski, W.; Tokarz, B.; Banasiuk, R.; Królicka, A.; Dziurka, M.; Wojciechowska, R.; Tokarz, K.M. Is a blue-red light a good elicitor of phenolic compounds in the family Droseraceae? A comparative study. J. Photochem. Photobiol. B 2019, 201, 111679. [Google Scholar] [CrossRef] [PubMed]
- Bantis, F.; Panteris, E.; Dangitsis, C.; Carrera, E.; Koukounaras, A. Blue light promotes vascular reconnection, while red light boosts the physiological response and quality of grafted watermelon seedlings. Sci. Rep. 2021, 11, 21754. [Google Scholar] [CrossRef]
- Zhang, S.; Ma, J.; Zou, H.; Zhang, L.; Li, S.; Wang, Y. The combination of blue and red LED light improves growth and phenolic acid contents in Salvia miltiorrhiza Bunge. Ind. Crops Prod. 2020, 158, 112959. [Google Scholar] [CrossRef]
- Terfa, M.T.; Olsen, J.E.; Torre, S. Blue light improves stomatal function and dark-induced closure of rose leaves (Rosa x hybrida) developed at high air humidity. Front. Plant Sci. 2020, 11, 1036. [Google Scholar]
- Ahres, M.; Pálmai, T.; Gierczik, K.; Dobrev, P.; Vanková, R.; Galiba, G. The impact of far-red light supplementation on hormonal responses to cold acclimation in barley. Biomolecules 2021, 11, 450. [Google Scholar] [CrossRef]
- Seiler, F.; Soll, J.; Bölter, B. Comparative phenotypical and molecular analyses of Arabidopsis grown under fluorescent and LED light. Plants 2017, 6, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunter, D.C.; Burritt, D.J. Improved adventitious shoot production from cotyledon explants of lettuce (Lactuca sativa L.). Sci. Horticult. 2002, 95, 269–276. [Google Scholar] [CrossRef]
- Seung, D.; Risopatron, J.; Jones, B.; Marc, J. Circadian clock-dependent gating in ABA signalling networks. Protoplasma 2012, 249, 445–457. [Google Scholar] [CrossRef] [PubMed]
- Kurepin, L.V.; Walton, L.J.; Reid, D.M.; Chinnappa, C.C. Light regulation of endogenous salicylic acid levels in hypocotyls of Helianthus annuus seedlings. Botany 2010, 88, 668–674. [Google Scholar] [CrossRef]
- Pál, M.; Horváth, E.; Janda, T.; Páldi, E.; Szalai, G. Cadmium stimulates the accumulation of salicylic acid and its putative precursors in maize (Zea mays L.) plants. Physiol. Plant. 2005, 125, 356–364. [Google Scholar] [CrossRef]
- Genty, B.; Briantais, J.M.; Baker, N.R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim. Biophys. Acta 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Németh, M.; Janda, T.; Horváth, E.; Páldi, E.; Szalai, G. Exogenous salicylic acid increases polyamine content but may decrease drought tolerance in maize. Plant Sci. 2002, 162, 569–574. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Tajti, J.; Pál, M.; Janda, T. Validation of reference genes for studying different abiotic stresses in oat (Avena sativa L.) by RT-qPCR. Plants 2021, 10, 1272. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−∆∆Ct method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Paolacci, A.R.; Tanzarella, O.A.; Porceddu, E.; Ciaffi, M. Identification and validation of reference genes for quantitative RT-PCR normalization in wheat. BMC Mol. Biol. 2009, 10, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gardiner, D.M.; Kazan, K.; Praud, S.; Torney, F.J.; Rusu, A.; Manners, J.M. Early activation of wheat polyamine biosynthesis during Fusarium head blight implicates putrescine as an inducer of trichothecene mycotoxin production. BMC Plant Biol. 2010, 10, 289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, H.; Guo, H.; Xie, Y.; Zhao, L.; Gu, J.; Zhao, S.; Li, J.; Liu, L. RNAseq analysis reveals pathways and candidate genes associated with salinity tolerance in a spaceflight-induced wheat mutant. Sci. Rep. 2017, 7, 2731. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| μg g −1 FW | Trans-Violaxanthin | Trans-Neoxanthin | Trans-Lutein | Chlorophyll a | Chlorophyll b | Trans-β-Carotene | 9-Cis-β-Carotene | |
|---|---|---|---|---|---|---|---|---|
| C | W | 57.7 ± 0.4 a | 52.4 ± 0.5 g | 189 ± 1 d | 639. 7 ± 3 bc | 280.7 ± 1.2 a | 59.3 ± 0.7 c | 6.4 ± 0.1 ab |
| C | B | 77.8 ± 1.8 | 48.5 ± 1.7 ef | 193 ± 3.6 d | 714. 3 ± 7.5 e | 309. 7 ± 2.5 fg | 72.2 ± 1.6 e | 8.8 ± 0.1 e |
| R | 54.9 ± 0.8 a | 33 ± 0.8 a | 157. 7 ± 1.5 a | 622 ± 11.1 ab | 288. 3 ± 5.5 ab | 51.8 ± 0.6 a | 6.7 ± 0.1 bc | |
| P | 67.7 ± 1.4 d | 39.3 ± 0.5 b | 178. 7 ± 2.1 c | 691 ± 8.7 d | 304 ± 2.6 ef | 64.2 ± 1.2 d | 8.3 ± 0.1 d | |
| PUT | B | 74.5 ± 1.8 e | 43.8 ± 1.7 cd | 190 ± 4.6 d | 746. 3 ± 4 fg | 323 ± 1 h | 69.4 ± 2.8 de | 8.8 ± 0.2 e |
| R | 57.5 ± 1.9 a | 38.8 ± 1.3 b | 165. 7 ± 5 ab | 605 ± 12.5 a | 282 ± 5.6 a | 51.4 ± 1 a | 6.2 ± 0.1 a | |
| P | 75.3 ± 0.6 e | 45.9 ± 0.8 de | 192 ± 1.7 d | 725 ± 7 e | 316. 3 ± 3 gh | 70.4 ± 1.7 de | 8.4 ± 0.2 d | |
| SPD | B | 86.8 ± 2 f | 52.5 ± 1.4 g | 204. 7 ± 4.6 ef | 772 ± 15.9 h | 331. 7 ± 6.7 i | 76.4 ± 0.4 f | 9.4 ± 0.1 f |
| R | 62.7 ± 3.3 b | 41.5 ± 2.5 bc | 176. 3 ± 7 c | 651. 3 ± 11 c | 294. 7 ± 4.5 bc | 56.9 ± 2.7 bc | 6.8 ± 0.7 c | |
| P | 74.3 ± 0.6 e | 44.9 ± 1 cd | 190. 7 ± 1.5 d | 731. 3 ± 4 ef | 318 ± 2 h | 70.6 ± 2.4 de | 8.3 ± 0.1 d | |
| SPM | B | 84.2 ± 2.5 f | 51.5 ± 1.7 fg | 209.3 ± 4.9 f | 790. 3 ± 12.9 i | 346. 7 ± 4.7 j | 75.8 ± 1.1 f | 9.3 ± 0.2 f |
| R | 62 ± 0.9 b | 39.9 ± 0.8 b | 171. 3 ± 2.5 bc | 656 ± 11.1 c | 298. 3 ± 4.5 cd | 54.4 ± 0.9 ab | 6.6 ± 0.04 bc | |
| P | 74 ± 6.5 e | 44.1 ± 5.5 cd | 204 ± 12 de | 755. 7 ± 20.5 gh | 330. 7 ± 7.5 i | 68.1 ± 4 d | 8.6 ± 0.4 de | |

| Shoot Length (cm) | Root Length (cm) | Shoot Weight (g −1 plant) | Root Weight (g −1 plant) | ||
|---|---|---|---|---|---|
| C | W | 27.04 ± 1.45 h | 18.84 ± 1.93 e | 0.31 ± 0.02 a | 0.15 ± 0.02 a |
| C | B | 19.4 ± 0.96 a | 16.56 ± 1.44 bc | 0.34 ± 0.02 a | 0.23 ± 0.02 bc |
| R | 25.8 ± 2.18 g | 17.88 ± 4 de | 0.38 ± 0.06 abc | 0.22 ± 0.04 abc | |
| P | 21.56 ± 1.33 c | 15.88 ± 2.11 ab | 0.32 ± 0.02 a | 0.21 ± 0.03 abc | |
| PUT | B | 20.56 ± 1.36 b | 17.84 ± 1.62 dc | 0.35 ± 0.08 ab | 0.22 ± 0.02 abc |
| R | 25.84 ± 1.37 g | 18.24 ± 2.22 de | 0.45 ± 0.04 d | 0.31 ± 0.03 c | |
| P | 22.04 ± 1 cd | 16.16 ± 2.37 ab | 0.35 ± 0.04 a | 0.28 ± 0.04 cd | |
| SPD | B | 20.52 ± 1.85 b | 14.96 ± 3.12 a | 0.33 ± 0.06 a | 0.19 ± 0.04 ab |
| R | 24.52 ± 1.69 f | 15.76 ± 1.83 ab | 0.44 ± 0.03 cd | 0.28 ± 0.03 cd | |
| P | 22.32 ± 1.63 d | 17.6 ± 2.5 cd | 0.37 ± 0.0 abc | 0.23 ± 0.04 abc | |
| SPM | B | 21.43 ± 1.14 c | 15.53 ± 2.26 ab | 0.38 ± 0.03 abc | 0.2 ± 0.02 ab |
| R | 25.27 ± 1.68 g | 14.93 ± 2.36 a | 0.47 ± 0.04 d | 0.22 ± 0.01 abc | |
| P | 23.13 ± 1 e | 15.47 ± 1.5 ab | 0.379 ± 0.02 abc | 0.16 ± 0.01 ab |

| Treatments | Intensity PAR (µmol) | Blue µW/cm2 (400–500 nm) | Green µW/cm2 (500–600 nm) | Red µW/cm2 (600–700 nm) | Far-red µW/cm2 (700–800 nm) | Blue% | Green% | Red% | Far-red% | Blue/Red | Red/Far-red |
|---|---|---|---|---|---|---|---|---|---|---|---|
| White (W) | 250 | 1120 | 1840 | 2240 | 130 | 21 | 34.52 | 42.03 | 2.44 | 0.5 | 17.23 |
| Blue (B) | 250 | 5060 | 50 | 1010 | 10 | 82.55 | 0.82 | 16.48 | 0.16 | 5 | 101 |
| Red + Far-red (R) | 250 | 720 | 680 | 3420 | 340 | 13.95 | 13.18 | 66.28 | 6.59 | 0.21 | 10.06 |
| Pink (P) | 250 | 2720 | 20 | 2690 | 20 | 49.91 | 0.37 | 49.36 | 0.37 | 1.01 | 134.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pál, M.; Hamow, K.Á.; Rahman, A.; Majláth, I.; Tajti, J.; Gondor, O.K.; Ahres, M.; Gholizadeh, F.; Szalai, G.; Janda, T. Light Spectral Composition Modifies Polyamine Metabolism in Young Wheat Plants. Int. J. Mol. Sci. 2022, 23, 8394. https://doi.org/10.3390/ijms23158394
Pál M, Hamow KÁ, Rahman A, Majláth I, Tajti J, Gondor OK, Ahres M, Gholizadeh F, Szalai G, Janda T. Light Spectral Composition Modifies Polyamine Metabolism in Young Wheat Plants. International Journal of Molecular Sciences. 2022; 23(15):8394. https://doi.org/10.3390/ijms23158394
Chicago/Turabian StylePál, Magda, Kamirán Áron Hamow, Altafur Rahman, Imre Majláth, Judit Tajti, Orsolya Kinga Gondor, Mohamed Ahres, Fatemeh Gholizadeh, Gabriella Szalai, and Tibor Janda. 2022. "Light Spectral Composition Modifies Polyamine Metabolism in Young Wheat Plants" International Journal of Molecular Sciences 23, no. 15: 8394. https://doi.org/10.3390/ijms23158394
APA StylePál, M., Hamow, K. Á., Rahman, A., Majláth, I., Tajti, J., Gondor, O. K., Ahres, M., Gholizadeh, F., Szalai, G., & Janda, T. (2022). Light Spectral Composition Modifies Polyamine Metabolism in Young Wheat Plants. International Journal of Molecular Sciences, 23(15), 8394. https://doi.org/10.3390/ijms23158394