
Transcriptional Regulation and Protein Localization of Zip10, Zip13 and Zip14 Transporters of Freshwater Teleost Yellow Catfish Pelteobagrus fulvidraco Following Zn Exposure in a Heterologous HEK293T Model

 ,
, 
Abstract
:1. Introduction
2. Results
2.1. Cloning and Sequence Analysis of zip10, zip13, and zip14 Promoters of Yellow Catfish
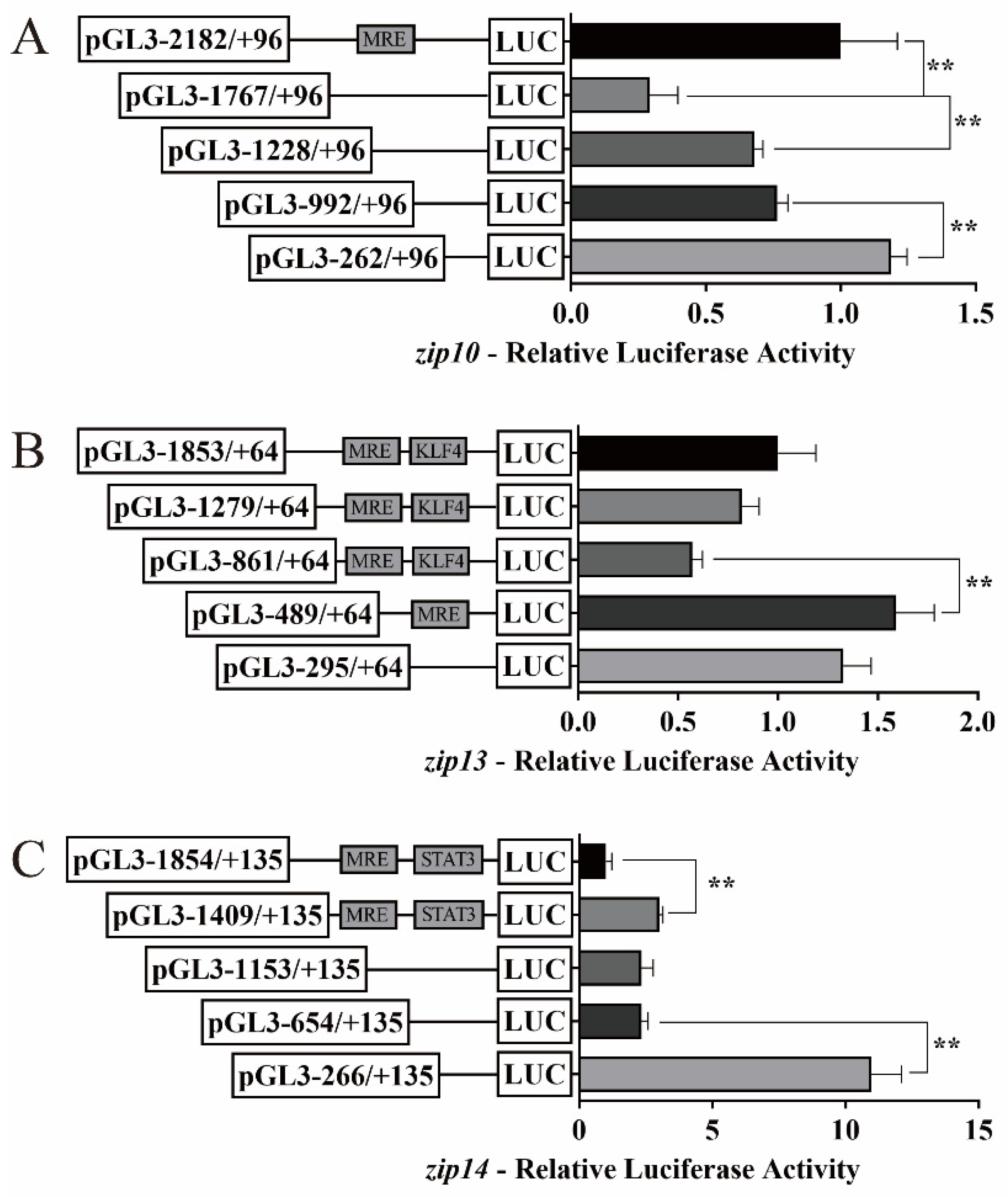
2.2. 5′-Sequence Deletion Mutation Assays of zip10, zip13, and zip14 Promoters of Yellow Catfish
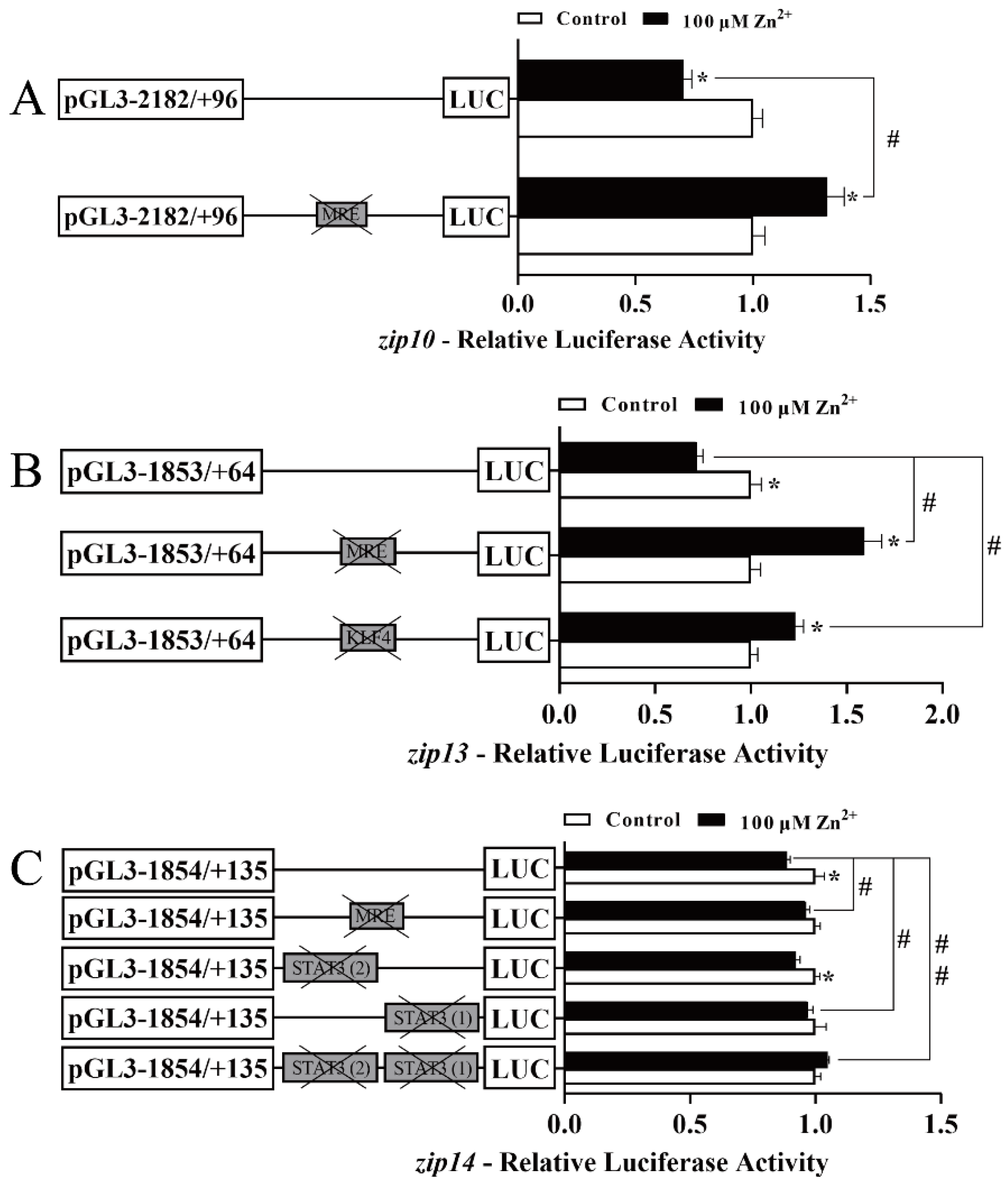
2.3. Site-Mutation Assays of the MTF-1, KLF4, and STAT3 Binding Sites in the zip10, zip13 and zip14 Promoters of Yellow Catfish
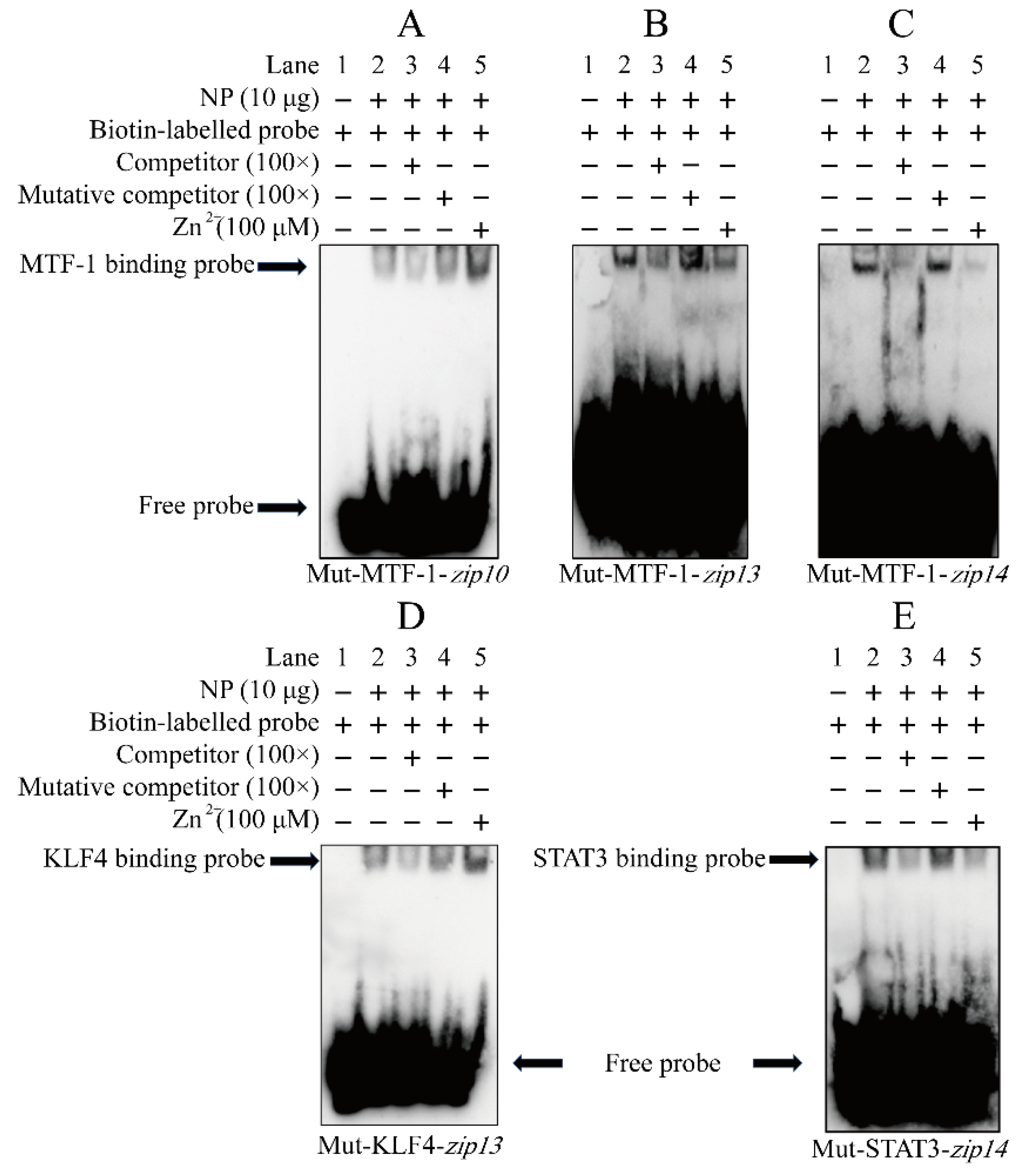
2.4. Electrophoretic Mobility Shift Assay
2.5. Response of pfZip10-EGFP, pfZip13-EGFP, and pfZip14-EGFP to Zn Administration in HEK293T Cells
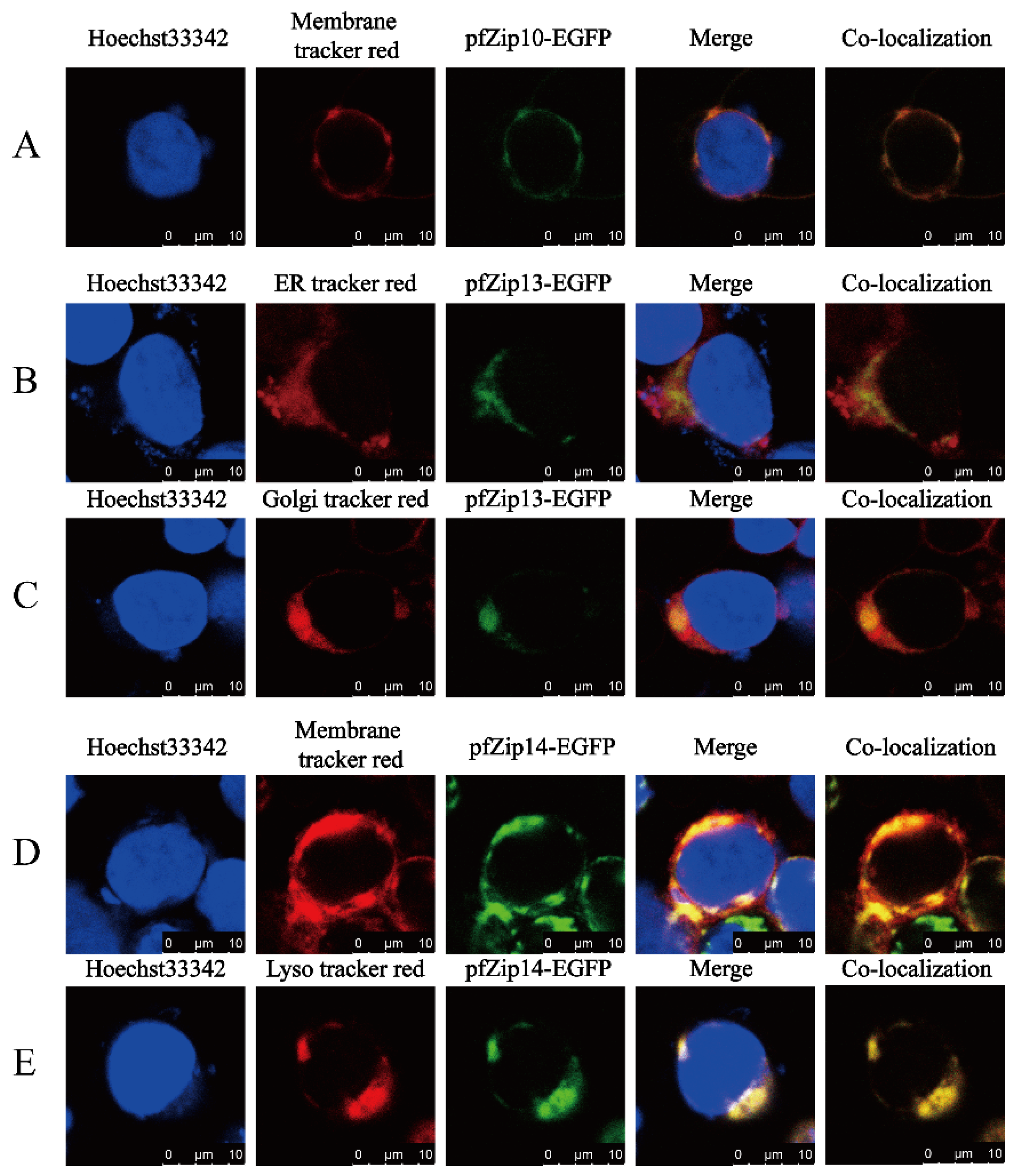
2.6. The Subcellular Localization of the Zip10, Zip13, and Zip14 Proteins of Yellow Catfish in HEK293T Cells
3. Discussion
4. Materials and Methods
4.1. Animals, Cells, and Reagents
4.2. Promoters Cloning and Plasmids Construction
4.3. Promoter Sequence Analysis
4.4. Dual Luciferase Activities Assays
4.5. Site Mutation Analysis of TFBS in the zip10, zip13, and zip14 Promoters
4.6. Electrophoretic Mobility-Shift Assay
4.7. Fluorescence Intensity Analysis
4.8. Subcellular Localization Investigation of Zip10, Zip13, and Zip14
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Roh, H.C.; Collier, S.; Guthrie, J.; Robertson, J.D.; Kornfeld, K. Lysosome-related organelles in intestinal cells are a zinc storage site in C. elegans. Cell Metab. 2012, 15, 88–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, T.; Kiron, V.; Satoh, S. Trace minerals in fish nutrition. Aquaculture 1997, 151, 185–207. [Google Scholar] [CrossRef]
- Lall, S.P.; Kaushik, S.J. Nutrition and Metabolism of Minerals in Fish. Animals 2021, 11, 2711. [Google Scholar] [CrossRef] [PubMed]
- De Schamphelaere, K.A.C.; Janssen, C.R. Bioavailability and Chronic Toxicity of Zinc to Juvenile Rainbow Trout (Oncorhynchus mykiss): Comparison with Other Fish Species and Development of a Biotic Ligand Model. Environ. Sci. Technol. 2004, 38, 6201–6209. [Google Scholar] [CrossRef] [PubMed]
- Kambe, T.; Tsuji, T.; Hashimoto, A.; Itsumura, N. The Physiological, Biochemical, and Molecular Roles of Zinc Transporters in Zinc Homeostasis and Metabolism. Physiol. Rev. 2015, 95, 3. [Google Scholar] [CrossRef]
- Zhao, L.; Tan, J.Q.; Li, D.; Jiang, L.J.; Li, T.J.; Yang, Y.; Wang, G.X.; Shang, Z.; Wang, J.; Zhou, J.F. SLC39A6/ZIP6 is essential for zinc homeostasis and T-cell development in zebrafish. Biochem. Biophys. Res. Commun. 2019, 511, 896–902. [Google Scholar] [CrossRef]
- Mackay, E.W.; Molero, S.I.; Tirathdas, L.H.; Peterson-Maduro, J.; Zang, J.J.; Neuhauss, S.C.F.; Schulte-Merker, S.; Wilson, S.W. Zinc dysregulation in slc30a8 (znt8) mutant zebrafish leads to blindness and disrupts bone mineralisation. bioRxiv 2020. [Google Scholar] [CrossRef]
- Muylle, F.A.R.; Adriaensen, D.; Coen, W.D.; Jean-Pierre Timmermans, J.P.; Blust, R. Tracing of labile zinc in live fish hepatocytes using FluoZin-3. BioMetals 2006, 19, 437–450. [Google Scholar] [CrossRef]
- Nimmanon, T.; Ziliotto, S.; Ogle, O.; Burt, A.; Gee, J.M.W.; Andrews, G.K. The ZIP6/ZIP10 heteromer is essential for the zinc-mediated trigger of mitosis. Cell Mol. Life Sci. 2021, 78, 1781–1798. [Google Scholar] [CrossRef]
- Bin, B.H.; Fukada, T.; Hosaka, T.; Nishida, K.; Yokoyama, S.; Hirano, T. Biochemical Characterization of Human ZIP13 Protein: A homo-dimerized zinc transporter involved in the spondylocheiro dysplastic ehlers-danlos syndrome. J. Biol. Chem. 2011, 286, 40255–40265. [Google Scholar] [CrossRef] [Green Version]
- Aydemir, T.B.; Troche, C.; Kim, M.H.; Cousins, R.J. Hepatic ZIP14-mediated Zinc Transport Contributes to Endosomal Insulin Receptor Trafficking and Glucose. J. Biol. Chem. 2016, 291, 23939–23951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Croxford, T.P.; McCormick, N.H.; Kelleher, S.L. Moderate Zinc Deficiency Reduces Testicular Zip6 and Zip10 Abundance and Impairs Spermatogenesis in Mice. J. Nutr. 2011, 41, 359–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, D.; Feeney, G.P.; Kille, P.; Hogstrand, C. Reglation of ZIP and ZnT zinc transporters in zebrafish gill: Zinc repression of ZIP10 transcription by an intronic MRE cluster. Physiol. Genom. 2008, 34, 205–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lichten, L.A.; Ryu, M.S.; Guo, L.; Embury, J.; Cousins, R.J. MTF-1-mediated repression of the zinc transporter Zip10 is alleviated by zinc restriction. PLoS ONE 2011, 6, e21526. [Google Scholar] [CrossRef]
- Lee, M.G.; Bin, B.H. Different actions of intracellular zinc transporters ZIP7 and ZIP13 are essential for dermal development. Int. J. Mol. Sci. 2019, 20, 3941. [Google Scholar] [CrossRef] [Green Version]
- Fukada, T.; Asada, Y.; Mishima, K.; Shimoda, S.; Saito, I. Slc39a13/Zip13, a crucial zinc transporter involved in tooth development and inherited disorders. J. Oral Biosci. 2011, 531, 1–12. [Google Scholar] [CrossRef]
- Jeong, J.; Walker, J.M.; Wang, F.; Park, J.G.; Palmer, A.E.; Giunta, C.; Rohrbach, M.; Steinmann, B.; Eidea, D.J. Promotion of vesicular zinc efflux by ZIP13 and its implications for spondylocheiro dysplastic Ehlers–Danlos syndrome. Proc. Natl. Acad. Sci. USA 2012, 109, E3530–E3538. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.X.; Wan, Z.H.; Zhou, B. Drosophila ZIP13 is post-translationally regulated by iron-mediated stabilization. BBA–Mol. Cell Res. 2019, 1866, 1487–1497. [Google Scholar] [CrossRef]
- Liuzzi, J.P.; Lichten, L.A.; Rivera, S.; Blanchard, R.K.; Aydemir, T.B.; Knutson, M.D.; Ganz, T.; Cousins, R.J. Interleukin-6 regulates the zinc transporter Zip14 in liver and contributes to the hypozincemia of the acute-phase response. Proc. Natl. Acad. Sci. USA 2005, 102, 6843–6848. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.H.; Aydemir, T.B.; Kim, J.; Cousins, R.J. Hepatic ZIP14-mediated zinc transport is required for adaptation to endoplasmic reticulum stress. Proc. Natl. Acad. Sci. USA 2017, 114, E5805–E5814. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.W.; Wu, K.; Lv, W.H.; Song, C.C.; Luo, Z. Molecular characterization of ten zinc Zn transporter genes and their regulation to Zn metabolism in freshwater teleost yellow catfish Pelteobagrus fulvidraco. J. Trace Elem. Med. Bio. 2020, 59, 126433. [Google Scholar] [CrossRef] [PubMed]
- Khan, R.; Abbas Raza, S.H.; Schreurs, N.; Wang, X.Y.; Wang, H.B.; Ullah, I.; AbdurRahman, A.; Suhail, S.M.; Khan, S.; Linsen, Z. Bioinformatics analysis and transcriptional regulation of TORC1 gene through transcription factors NRF1 and Smad3 in bovine preadipocytes. Genomics 2020, 112, 1575–1587. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.; Li, A.; Raza, S.H.A.; Khan, R.; Wang, X.; Wang, S.; Wang, G.; Zhang, Y.; Zan, L. The Molecular Characteristics of the FAM13A Gene and the Role of Transcription Factors ACSL1 and ASCL2 in Its Core Promoter Region. Genes 2019, 10, 981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laity, J.H. and Andrews, G.K. Understanding the mechanisms of zinc-sensing by metal response element binding transcription factor-1 (MTF-1). Arch. Biochem. Biophys. 2007, 463, 201–210. [Google Scholar] [CrossRef]
- Kimura, T.; Itoh, N.; Andrews, G.K. Mechanisms of heavy metal sensing by metal response element-binding transcription factor-1. J. Health Sci. 2009, 55, 484–494. [Google Scholar] [CrossRef] [Green Version]
- Giedroc, D.P.; Chen, X.H.; Apuy, J.L. Metal response element (MRE)-binding transcription factor-1 (MTF-1), structure, function, and regulation. Antioxid. Redox. Sign. 2001, 3, 577–596. [Google Scholar] [CrossRef]
- Song, C.C.; Chen, G.H.; Zhong, C.C.; Chen, F.; Chen, S.W.; Luo, Z. Transcriptional responses of four slc30a/znt family members and their roles in Zn homeostatic modulation in yellow catfish Pelteobagrus fulvidraco. BBA-Gene Regul. Mech. 2021, 1864, 194723. [Google Scholar] [CrossRef]
- Chen, S.W.; Wu, K.; Lv, W.H.; Chen, F.; Song, C.C.; Luo, Z. Functional analysis of two zinc Zn, ransporters ZIP3 and ZIP8, promoters and their distinct response to MTF1 and RREB1 in the regulation of Zn metabolism. Int. J. Mol. Sci. 2020, 21, 6135. [Google Scholar] [CrossRef]
- Hardyman, J.E.; Tyson, J.; Jackson, K.A.; Aldridge, C.; Cockell, S.J.; Wakeling, L.A.; Valentine, R.A.; Ford, D. Zinc sensing by metal-responsive transcription factor 1 (MTF1) controls metallothionein and ZnT1 expression to buffer the sensitivity of the transcriptome response to zinc. Metallomics 2016, 8, 337–343. [Google Scholar] [CrossRef] [Green Version]
- Jackson, K.A.; Helston, R.M.; McKay, J.A.; O’Neill, E.D.; Mathers, J.C.; Ford, D. Splice variants of the human zinc transporter ZnT5 SLC30A5 are differentially localized and regulated by zinc through transcription and mRNA stability. J. Biol. Chem. 2007, 282, 10423–10431. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.J.; Liu, Y.C.; Lin, M.C.; Chen, Y.T.; Lin, L.Y. Coordinative modulation of human zinc transporter 2 gene expression through active and suppressive regulators. J. Nutr. Biochem. 2015, 26, 351–359. [Google Scholar] [CrossRef]
- Ghaleb, A.M. and Yang, V.W. Krüppel-like factor 4 (KLF4): What we currently know. Gene 2017, 611, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Liuzzi, J.P.; Guo, L.; Chang, S.M.; Cousins, R.J. Krüppel-like factor 4 regulates adaptive expression of the zinc transporter Zip4 in mouse small intestine. Am. J. Physiol.-Gastr. Liver Physiol. 2009, 296, G517–G523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segre, J.A.; Bauer, C.; Fuchs, E. KLF4 is a transcription factor required for establishing the barrier function of the skin. Nat. Genet. 1999, 22, 356–360. [Google Scholar] [CrossRef] [PubMed]
- Levy, D.E.; Lee, C.K. What does Stat3 do? J. Clin. Investig. 2002, 109, 1143–1148. [Google Scholar] [CrossRef]
- Liu, J.; Zhou, Z.X.; Zhang, W.; Bell, M.W.; Waalkes, M.P. Changes in hepatic gene expression in response to hepatoprotective levels of zinc. Liver Int. 2009, 29, 1222–1229. [Google Scholar] [CrossRef] [Green Version]
- Kitabayashi, C.; Fukada, T.; Kanamoto, M.; Ohashi, W.; Hojyo, S.; Atsumi, T.; Ueda, N.; Azuma, I.; Hirota, H.; Murakami, M.; et al. Zinc suppresses Th17 development via inhibition of STAT3 activation. Int. Immunol. 2010, 22, 375–386. [Google Scholar] [CrossRef] [Green Version]
- Cousins, R.J.; Liuzzi, J.P.; Lichten, L.A. Mammalian zinc transport, trafficking, and signals. J. Biol. Chem. 2006, 281, 24085–24089. [Google Scholar] [CrossRef] [Green Version]
- Bury, N.R.; Walker, P.A.; Glover, G.N. Nutritive metal uptake in teleost fish. J. Exp. Biol. 2003, 206, 11–23. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.H.; Lv, W.H.; Xu, Y.H.; Wei, X.L.; Xu, Y.C.; Luo, Z. Functional analysis of MTF-1 and MT promoters and their transcriptional response to zinc Zn and copper Cu in yellow catfish Pelteobagrus fulvidraco. Chemosphere 2020, 246, 125792. [Google Scholar] [CrossRef]
- Song, C.C.; Wu, L.X.; Chen, G.H.; Lv, W.H.; Chen, S.W.; Luo, Z. Six members of SLC30A/ZnTs family related with the control of zinc homeostasis, characterization, mRNA expression and their responses to dietary ZnO nanoparticles in yellow catfish. Aquaculture 2020, 528, 735570. [Google Scholar] [CrossRef]
- Chen, G.H.; Zhao, T.; Wei, X.L.; Zhang, D.G.; Zhuo, M.Q.; Luo, Z. miR-101b Regulates Lipid Deposition and Metabolism of Primary Hepatocytes in Teleost Yellow Catfish Pelteobagrus fulvidraco. Genes 2020, 11, 861. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.G.; Zhao, T.; Xu, X.J.; Lv, W.H.; Luo, Z. Dietary marginal and excess selenium increased triglycerides deposition, induced endoplasmic reticulum stress and differentially influenced selenoproteins expression in the anterior and middle intestines of yellow catfish Pelteobagrus fulvidraco. Antioxidants 2021, 10, 535. [Google Scholar] [CrossRef]
- Wu, L.X.; Xu, Y.C.; Hogstrand, C.; Zhao, T.; Wu, K.; Xu, Y.H.; Liu, W.; Luo, Z. Lipophagy mediated glucose-induced changes of lipid deposition and metabolism via ROS dependent AKT-Beclin1 activation. J. Nutr. Biochem. 2022, 100, 108882. [Google Scholar] [CrossRef] [PubMed]
- Miller, D.A. ‘Significant’ and ‘Highly Significant’. Nature 1966, 210, 1190. [Google Scholar] [CrossRef] [PubMed]
- Shihan, M.H.; Novo, S.G.; Le Marchand, S.J.; Wang, Y.; Duncan, M.K. A simple method for quantitating confocal flu-orescent images. Biochem. Biophys. Rep. 2021, 25, 100916. [Google Scholar] [CrossRef] [PubMed]
- Ruggiero, I.; Terracciano, M.; Martucci, N.M.; Stefano, L.D.; Migliaccio, N.; Tatè, R.; Rendina, I.; Arcari, P.; Lamberti, A.; Rea, I. Diatomite silica nanoparticles for drug delivery. Nanoscale Res. Lett. 2014, 9, 329. [Google Scholar] [CrossRef] [Green Version]
- Shihan, M.H.; Kanwar, M.; Wang, Y.; Jackson, E.E.; Faranda, A.P.; Duncan, M.K. Fibronectin has multifunctional roles in posterior capsular opacification (PCO). Matrix Biol. 2020, 90, 79–108. [Google Scholar] [CrossRef]
- Khanh Le, N.Q.; Yee Yapp, E.K.; Nagasundaram, N.; Yeh, H.Y. Classifying Promoters by Interpreting the Hidden In-formation of DNA Sequences via Deep Learning and Combination of Continuous FastText N-Grams. Front. Bioeng. Biotechnol. 2019, 7, 305. [Google Scholar] [CrossRef] [Green Version]
- Yan, C.H.M.; Chan, K.M. Cloning of zebrafish metallothionein gene and characterization of its gene promoter region in HepG2 cell line. Biochim. Biophys. Acta (BBA)—Gene Struct. Expr. 2004, 1679, 47–58. [Google Scholar] [CrossRef]
- Sloutskin, A.; Shir-Shapira, H.; Freiman, R.N.; Gershon, T.J. The core promoter is a regulatory hub for developmental gene expression. Front. Cell. Dev. Biol. 2021, 9, 666508. [Google Scholar] [CrossRef] [PubMed]
- Goodrich, J.A.; Tjian, R. Unexpected roles for core promoter recognition factors in cell-type-specific transcription and gene regulation. Nat. Rev. Genet. 2010, 11, 549–558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zang, Z.S.; Xu, Y.M.; Lau Andy, T.Y. Molecular and pathophysiological aspects of metal ion uptake by the zinc transporter ZIP8 (SLC39A8). Toxicol. Res. 2016, 5, 987–1002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langmade, S.J.; Ravindra, R.; Daniels, P.J.; Andrews, G.K. The transcription factor MTF-1 mediates metal regulation of the mouse ZnT1 gene. J. Biol. Chem. 2000, 275, 34803–34809. [Google Scholar] [CrossRef] [Green Version]
- Bird, J.A.; Blankman, E.; Stillman, D.J.; Eide, D.J.; Winge, D.R. The Zap1 transcriptional activator also acts as a repressor by binding downstream of the TATA box in ZRT2. EMBO J. 2004, 23, 1123–1132. [Google Scholar] [CrossRef] [Green Version]
- Dolfini, D.; Zambelli, F.; Pavesi, G.; Mantovani, R.A. perspective of promoter architecture from the CCAAT-box. Cell Cycle 2009, 8, 4127–4137. [Google Scholar] [CrossRef] [Green Version]
- Nishito, Y.; Kambe, T. Zinc transporter 1 ZNT1 expression on the cell surface is elaborately controlled by cellular zinc levels. J. Biol. Chem. 2019, 294, 15686–15697. [Google Scholar] [CrossRef]
- Guo, L.; Lichten, L.A.; Ryu, M.S.; Liuzzi, J.P.; Wang, F.; Cousins, R.J. STAT5-glucocorticoid receptor interaction and MTF-1 regulate the expression of Znt2 (SLC30A2) in pancreatic acinar cells. Proc. Natl. Acad. Sci. USA 2010, 107, 2818–2823. [Google Scholar] [CrossRef] [Green Version]
- Du, L.; Zhang, H.; Zhao, H.; Cheng, X.; Qin, J.; Teng, T.; Yang, Q.; Xu, Z. The critical role of the zinc transporter Zip2 SLC39A2 in ischemia/reperfusion injury in mouse hearts. J. Mol. Cell Cardiol. 2019, 132, 136–145. [Google Scholar] [CrossRef]
- Hogstrand, C.; Kille, P.; Ackland, M.L.; Hiscox, S.; Taylor, K.M. A mechanism for epithelial-mesenchymal transition and anoikis resistance in breast cancer triggered by zinc channel ZIP6 and STAT3 (signal transducer and activator of transcription 3). Biochem. J. 2013, 455, 229–237. [Google Scholar] [CrossRef]
- Yamashita, S.; Miyagi, C.; Fukada, T.; Kagara, N.; Che, Y.S.; Hirano, T. Zinc transporter LIVI controls epithelial mesenchymal transition in zebrafish gastrula organizer. Nature 2004, 429, 298–302. [Google Scholar] [CrossRef] [PubMed]
- Tan, G.; Lenhard, B. TFBSTools: An R/Bioconductor package for transcription factor binding site analysis. Bioinformatics 2016, 32, 1555–1556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, R.R.; Song, Z.X.; Lin, J.M.; Li, Z.G.; Shan, G.; Huang, C. Gawky modulates MTF-1-mediated transcription activation and metal discrimination. Nucleic Acids Res. 2021, 49, 6296–6314. [Google Scholar] [CrossRef] [PubMed]
- Wingelhofer, B.; Neubauer, H.A.; Valent, P. Implications of STAT3 and STAT5 signaling on gene regulation and chromatin remodeling in hematopoietic cancer. Leukemia 2018, 32, 1713–1726. [Google Scholar] [CrossRef] [Green Version]
- Ilsley, M.D.; Gillinder, K.R.; Magor, G.W.; Huang, S.; Bailey, T.L.; Crossley, M.; Perkins, A.C. Krüppel-like factors compete for promoters and enhancers to fine-tune transcription. Nucleic Acids Res. 2017, 45, 6572–6588. [Google Scholar] [CrossRef] [Green Version]
- Kong, B.Y.; Duncan, F.E.; Que, E.L.; Kim, A.M.; O’Halloran, T.V. Maternally-derived zinc transporters ZIP6 and ZIP10 drive the mammalian oocyte-to-egg transition. Mol. Hum. Reprod. 2014, 20, 1077–1089. [Google Scholar] [CrossRef]
- Fukunaka, A.; Fukada, T.; Bhin, J.; Suzuki, L.; Tsuzuki, T.; Takamine, Y.; Bin, B.H.; Yoshihara, T.; Sekine, N.I.; Naito, H.; et al. Zinc transporter ZIP13 suppresses beige adipocyte biogenesis and energy expenditure by regulating C/EBP-β expression. PLoS Genet. 2017, 138, e1006950. [Google Scholar] [CrossRef] [Green Version]
- Landry, G.M.; Furrow, E.; Holmes, H.L.; Hirata, T.; Kato, A.; Williams, P.; Strohmaier, K.; Gallo, C.J.R.; Chang, M.; Pandey, M.K.; et al. Cloning, function, and localization of human, canine and Drosophila ZIP10 (SLC39A10), a Zn2+ transporter. Am. J. Physiol.-Renal. 2019, 316, F263–F273. [Google Scholar] [CrossRef]
- Xiao, G.R.; Wan, Z.H.; Fan, Q.W.; Tang, X.N.; Zhou, B. The metal transporter ZIP13 supplies iron into the secretory pathway in Drosophila melanogaster. eLife 2014, 3, e03191. [Google Scholar] [CrossRef]
- Zhao, N.N.; Gao, J.W.; Enns, C.A.; Knutson, M.D. ZRT/IRT-like Protein 14 (ZIP14) Promotes the cellular assimilation of iron from transferrin. J. Biol. Chem. 2010, 285, 32141–32150. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Beattie, J.D.; Kim, B.F.; Petris, M.J.; Andrews, G.; Eide, D.J. Zinc-stimulated endocytosis controls activity of the mouse ZIP1 and ZIP3 zinc uptake transporters. J. Biol. Chem. 2004, 279, 24631–24639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, B.E.; Wang, F.; Dufner-Beattie, J.; Andrews, G.K.; Eide, D.J.; Petris, M.J. Zn2+-stimulated endocytosis of the mZIP4 zinc transporter regulates its location at the plasma membrane. J. Biol. Chem. 2004, 279, 4523–4530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.H.; Luo, Z.; Wu, K.; Song, Y.F.; You, W.J.; Zhang, L.H. Structure and functional analysis of promoters from two liver isoforms of CPT I in grass carp Ctenopharyngodon idella. Int. J. Mol. Sci. 2017, 18, 2405. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Location in zip10 Genomic Locus | Location in zip13 Genomic Locus | Location in zip14 Genomic Locus |
|---|---|---|---|
| MRE | −2017 bp/−2004 bp | −360 bp/−345 bp | −1457 bp/−1442 bp |
| PPARα:RXR | −1997 bp/−1980 bp | −543 bp/−529 bp | None |
| FOXO4 | −1976 bp/−1970 bp | −981 bp/−975 bp | −1687 bp/−1681 bp |
| FOXO6 | −1043 bp/−1037 bp | −248 bp/−242 bp | −681 bp /−675 bp and −435 bp /−429 bp |
| SREBP | −1760 bp/−1751 bp | None | −1785 bp/−1776 bp |
| CREB1 | −1729 bp/−1718 bp and −1330 bp/−1319 bp | None | −1293 bp/−1286 bp and −119 bp/−112 bp |
| AP1 | None | −1393 bp/−1383 bp | None |
| ZEB1 | −1601 bp/−1596 bp | −651/−640 bp | −1306 bp /−1301 bp |
| RREB1 | −353 bp/−334 bp | −731 bp/−724 bp | None |
| SP1 | −1423 bp/−1414 bp | None | −1253 bp/−1243 bp |
| KLF4 | −1124 bp/−1114 bp | −606 bp/−594 bp | None |
| STAT3 | None | None | (−1383 bp/−1375 bp and −1358 bp/−1348 bp) |
| STAT5a:STAT5b | −456 bp/−445 bp | None | None |
| STAT6 | −960 bp/−946 bp | −1126 bp/−1212 bp | None |
| CEBPα | −890 bp/−881 bp and −907 bp/−899 bp | −1300 bp/−1292 bp and −1123 bp/−1115 bp | −206 bp/−196 bp |
| ATF4 | None | −1027 bp/−1014 bp | None |
| CCAAT-box | −687 bp/−676 bp | −106/−95 bp | None |
| TATA-box | −319 bp/−305 bp | −92 bp/−78 bp | None |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, S.-Z.; Xu, Y.-C.; Tan, X.-Y.; Zhao, T.; Zhang, D.-G.; Yang, H.; Luo, Z. Transcriptional Regulation and Protein Localization of Zip10, Zip13 and Zip14 Transporters of Freshwater Teleost Yellow Catfish Pelteobagrus fulvidraco Following Zn Exposure in a Heterologous HEK293T Model. Int. J. Mol. Sci. 2022, 23, 8034. https://doi.org/10.3390/ijms23148034
Liu S-Z, Xu Y-C, Tan X-Y, Zhao T, Zhang D-G, Yang H, Luo Z. Transcriptional Regulation and Protein Localization of Zip10, Zip13 and Zip14 Transporters of Freshwater Teleost Yellow Catfish Pelteobagrus fulvidraco Following Zn Exposure in a Heterologous HEK293T Model. International Journal of Molecular Sciences. 2022; 23(14):8034. https://doi.org/10.3390/ijms23148034
Chicago/Turabian StyleLiu, Sheng-Zan, Yi-Chuang Xu, Xiao-Ying Tan, Tao Zhao, Dian-Guang Zhang, Hong Yang, and Zhi Luo. 2022. "Transcriptional Regulation and Protein Localization of Zip10, Zip13 and Zip14 Transporters of Freshwater Teleost Yellow Catfish Pelteobagrus fulvidraco Following Zn Exposure in a Heterologous HEK293T Model" International Journal of Molecular Sciences 23, no. 14: 8034. https://doi.org/10.3390/ijms23148034
APA StyleLiu, S.-Z., Xu, Y.-C., Tan, X.-Y., Zhao, T., Zhang, D.-G., Yang, H., & Luo, Z. (2022). Transcriptional Regulation and Protein Localization of Zip10, Zip13 and Zip14 Transporters of Freshwater Teleost Yellow Catfish Pelteobagrus fulvidraco Following Zn Exposure in a Heterologous HEK293T Model. International Journal of Molecular Sciences, 23(14), 8034. https://doi.org/10.3390/ijms23148034