
Molecular Mechanism of Equine Endometrosis: The NF-κB-Dependent Pathway Underlies the Ovarian Steroid Receptors’ Dysfunction

 , ,
, ,  ,
,  and
and
Abstract
1. Introduction
2. Results
2.1. The Categories and Histological Types of Equine Endometrosis and the Ovarian Steroid Receptors
2.2. Phases of the Estrous Cycle and Ovarian Steroid Receptors Regarding Equine Endometrosis Categories
2.3. Phases of the Estrous Cycle and Ovarian Steroid Receptors Regarding Histopathological Types of Equine Endometrosis
2.4. The Nf-Κb Signaling Pathway and Ovarian Steroid Receptors in Endometrium Categories and Endometrosis Types
3. Discussion
4. Materials and Methods
4.1. Biological Material Collection
4.2. Phases of Estrous Cycle Determination
4.3. Histopathological Examination of Mares’ Endometria
4.4. Gene Transcription Evaluation in Mares’ Endometria
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Aupperle, H.; Schoon, D.; Schoon, H.-A. Physiological and pathological expression of intermediate filaments in the equine endometrium. Res. Vet. Sci. 2004, 76, 249–255. [Google Scholar] [CrossRef] [PubMed]
- Da Costa, R.R.; Ferreira-Dias, G.; Mateus, L.; Korzekwa, A.; Andronowska, A.; Platek, R.; Skarzynski, D.J. Endometrial nitric oxide production and nitric oxide synthases in the equine endometrium: Relationship with microvascular density during the estrous cycle. Domest. Anim. Endocrinol. 2007, 32, 287–302. [Google Scholar] [CrossRef] [PubMed]
- Schöniger, S.; Schoon, H.A. The healthy and diseased equine endometrium: A review of morphological features and molecular analyses. Animals 2020, 10, 625. [Google Scholar] [CrossRef] [PubMed]
- Skarzynski, D.J.; Szóstek-Mioduchowska, A.Z.; Rebordão, M.R.; Jalali, B.M.; Piotrowska-Tomala, K.K.; Leciejewska, N.; Łazarczyk, M.; Ferreira-Dias, G.M. Neutrophils, monocytes and other immune components in the equine endometrium: Friends or foes? Theriogenology 2020, 150, 150–157. [Google Scholar] [CrossRef]
- Hoffmann, C.; Bazer, F.W.; Klug, J.; Aupperle. H.; Ellenberger, C.; Schoon, H.A. Immunohistochemical and histochemical identification of proteins and carbohydrates in the equine endometrium Expression patterns for mares suffering from endometrosis. Theriogenology 2009, 71, 264–274. [Google Scholar] [CrossRef]
- Rebordão, M.R.; Galvão, A.; Szóstek, A.; Amaral, A.; Mateus, L.; Skarzynski, D.J.; Ferreira-Dias, G. Physiopathologic mechanisms involved in mare endometrosis. Reprod. Dom. Anim. 2014, 49, 82–87. [Google Scholar] [CrossRef]
- Szóstek-Mioduchowska, A.; Leciejewska, N.; Zelmańska, B.; Staszkiewicz-Chodor, J.; Ferreira-Dias, G.; Skarzynski, D. Lysophosphatidic acid as a regulator of endometrial connective tissue growth factor and prostaglandin secretion during estrous cycle and endometrosis in the mare. BMC Vet. Res. 2020, 16, 343. [Google Scholar] [CrossRef]
- Minkwitz, C.; Schoon, H.-A.; Zhang, Q.; Schöniger, S. Plasticity of endometrial epithelial and stromal cells—A new approach towards the pathogenesis of equine endometrosis. Reprod. Domest. Anim. 2019, 54, 835–845. [Google Scholar] [CrossRef]
- Szóstek-Mioduchowska, A.Z.; Lukasik, K.; Skarzynski, D.J.; Okuda, K. Effect of transforming growth factor-β1 on α-smooth muscle actin and collagen expression in equine endometrial fibroblasts. Theriogenology 2019, 124, 9–17. [Google Scholar] [CrossRef]
- Mambelli, L.I.; Mattos, R.C.; Winter, G.H.Z.; Madeiro, D.S.; Morais, B.P.; Malschitzky, E.; Miglino, M.A.; Kerkis, A.; Kerkis, I. Changes in expression pattern of selected endometrial proteins following mesenchymal stem cells infusion in mares with endometrosis. PLoS ONE 2014, 9, e97889. [Google Scholar] [CrossRef]
- Amaral, A.; Fernandes, C.; Morazzo, S.; Rebordão, M.R.; Szóstek-Mioduchowska, A.; Lukasik, K.; Gawronska-Kozak, B.; da Gama, L.T.; Skarzynski, D.J.; Ferreira-Dias, G. The inhibition of cathepsin G on endometrial explants with endometrosis in the mare. Front. Vet. Sci. 2020, 7, 582211. [Google Scholar] [CrossRef]
- Katila, T.; Ferreira-Dias, G. Evolution of the Concepts of Endometrosis, Post Breeding Endometritis, and Susceptibility of Mares. Animals 2022, 12, 779. [Google Scholar] [CrossRef]
- Hoffman, C.; Ellenberger, C.; Mattos, R.C.; Aupperle, H.; Dhein, S.; Stief, B.; Schoon, H.-A. The equine endometrosis: New insights into the pathogenesis. Anim. Reprod. Sci. 2009, 111, 261–278. [Google Scholar] [CrossRef]
- Lehmann, J.; Ellenberger, C.; Hoffmann, C.; Bazer, F.W.; Klug, J.; Allen, W.R.; Sieme, H.; Schoon, H.A. Morpho-functional studies regarding the fertility prognosis of mares suffering from equine endometrosis. Theriogenology 2011, 76, 1326–1336. [Google Scholar] [CrossRef]
- Kenney, R.M.; Doig, P.A. Equine endometrial biopsy. In Current Therapy in Theriogenology; Morrow, D.A., Ed.; W.B. Saunders: Philadelphia, PA, USA, 1986; pp. 723–729. [Google Scholar]
- Schoon, H.-A.; Wiegandt, I.; Schoon, D.; Aupperle, H.; Bartmann, C.-P. Functional disturbances in the endometrium of barren mares: A histological and immunohistological study. J. Reprod. Fertil. Suppl. 2000, 56, 381–391. [Google Scholar]
- Brunckhorst, D.; Schoon, H.A.; Bader, H.; Sieme, H. Morphologische, enzym- und immunhistochemische Charakteristika des endometrialen Zyklus bei der Stute. Fertilitat 1991, 7, 44–51. [Google Scholar]
- Walter, I.; Klein, M.; Handler, J.; Aurich, J.E.; Reifinger, M.; Aurich, C. Lectin binding patterns of uterine glands in mares with chronic endometrial degeneration. Am. J. Vet. Res. 2001, 62, 840–845. [Google Scholar] [CrossRef]
- Wilkenfeld, S.R.; Lin, C.; Frigo, D.E. Communication between genomic and non-genomic signaling events coordinate steroid hormone actions. Steroids 2018, 133, 2–7. [Google Scholar] [CrossRef]
- Enmark, E.; Pelto-Huikko, M.; Grandien, K.; Lagercrantz, S.; Lagercrantz, J.; Fried, G.; Nordenskjold, M.; Gustafsson, J.A. Human estrogen receptor b-gene structure, chromosomal localization, and expression pattern. J. Clin. Endocr. Metab. 1997, 82, 4258–4265. [Google Scholar] [CrossRef]
- Silva, E.S.M.; Scoggin, K.E.; Canisso, I.F.; Troedsson, M.H.T.; Squires, E.L.; Ball, B.A. Expression of receptors for ovarian steroids and prostaglandin E2 in the endometrium and myometrium of mares during estrus, diestrus and early pregnancy. Anim. Reprod. Sci. 2014, 151, 169–181. [Google Scholar] [CrossRef]
- Mote, P.A.; Arnett-Mansfield, R.L.; Gava, N.; Defazio, A.; Mulac-Jericevic, B.; Conneely, O.M.; Clarke, C.L. Overlapping and distinct expression of progesterone receptors A and B in mouse uterus and mammary gland during the estrus cycle. Endocrinology 2006, 147, 5503–5512. [Google Scholar] [CrossRef]
- Watson, E.D.; Skolnik, S.B.; Zanecosky, H.G. Progesterone and estrogen receptor distribution in the endometrium of the mare. Theriogenology 1992, 38, 575–580. [Google Scholar] [CrossRef]
- Aupperle, H.; Özgen, S.; Schoon, H.A.; Schoon, D.; Hoppen, H.O.; Sieme, H.; Tannapfel, A. Cyclical endometrial steroid hormone receptor expression and proliferation intensity in the mare. Equine Vet. J. 2000, 32, 228–232. [Google Scholar] [CrossRef]
- DeFranco, D.B. Navigating steroid hormone receptors through the nuclear compartment. Mol. Endocrinol. 2002, 16, 1449–1455. [Google Scholar] [CrossRef]
- Weihua, Z.; Saji, S.; Makine, S.; Cheng, G.; Jensen, E.V.; Warner, M.; Gustafsson, J.A. Estrogen receptor (ER) ß, a modulator of ERα in the uterus. Proc. Natl. Acad. Sci. USA 2000, 97, 5936–5941. [Google Scholar] [CrossRef]
- Cunha, G.R.; Cooke, P.S.; Kurita, T. Role of stromal-epithelial interactions in hormonal responses. Arch. Histol. Cytol. 2004, 67, 417–434. [Google Scholar] [CrossRef]
- Mulac-Jericevic, B.; Lydon, J.P.; Demayo, F.J.; Conneely, O.M. Defective mammary gland morphogenesis in mice lacking the progesterone receptor B isoform. Proc. Natl. Acad. Sci. USA 2003, 100, 9744–9749. [Google Scholar] [CrossRef]
- Hartt, L.S.; Carling, S.J.; Joyce, M.M.; Johnson, G.A.; Vanderwall, D.K.; Ott, T.L. Temporal and spatial associations of oestrogen receptor alpha and progesterone receptor in the endometrium of cyclic and early pregnant mares. Reproduction 2005, 130, 241–250. [Google Scholar] [CrossRef]
- Cooke, P.S.; Buchanan, D.L.; Young, P.; Setiawan, T.; Brody, J.; Korach, K.S.; Taylor, J.; Lubahn, D.B.; Cunha, G.R. Stromal estrogen receptors mediate mitogenic effects of estradiol on uterine epithelium. Proc. Natl. Acad. Sci. USA 1997, 94, 6535–6540. [Google Scholar] [CrossRef]
- Domino, M.; Jasinski, T.; Kautz, E.; Juszczuk-Kubiak, E.; Ferreira-Dias, G.; Zabielski, R.; Sady, M.; Gajewski, Z. Expression of genes involved in the NF-κB-dependent pathway of the fibrosis in the mare endometrium. Theriogenology 2020, 147, 18–24. [Google Scholar] [CrossRef]
- Jasiński, T.; Zdrojkowski, Ł.; Kautz, E.; Juszczuk-Kubiak, E.; Ferreira-Dias, G.; Domino, M. Equine Endometrosis Pathological Features: Are They Dependent on NF-κB Signaling Pathway? Animals 2021, 11, 3151. [Google Scholar] [CrossRef] [PubMed]
- Umezawa, K. Possible role of peritoneal NF-kB in peripheral inflammation and cancer: Lessons from the inhibitor DHMEQ. Biomed Pharm. 2011, 65, 252.e9. [Google Scholar] [CrossRef] [PubMed]
- May, M.J.; Ghosh, S. Signal transduction through NF-κB. Trends Immunol. 1998, 19, 80–88. [Google Scholar] [CrossRef]
- Lind, D.S.; Hochwald, S.N.; Malaty, J.; Rekkas, S.; Hebig, P.; Mishra, G.; Moldawer, L.L.; Copeland III, E.M.; MacKay, S. Nuclear factor-κB is upregulated in colorectal cancer. Surgery 2001, 130, 363–369. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, P.; Aggarwal, A. NF-kB transcription factor: A key player in the generation of immune response. Curr. Sci. 2001, 90, 519–531. [Google Scholar]
- Ahn, B.N.; Song, M.H.; Kim, J.H.; Kim, K.H.; Park, K.K.; Choi, Y.S. Intra-peritoneal NF-kappaB decoy oligodeoxynucleotide decreases postoperative intra-abdominal adhesion. Korean J. Obstet. Gynecol. 2012, 55, 244–249. [Google Scholar] [CrossRef][Green Version]
- Sosinska, P.; Baum, E.; Mackowiak, B.; Staniszewski, R.; Jasinski, T.; Umezawa, K.; Breborowicz, A. Inhibition of NF-kappaB with Dehydroxymethylepoxyquinomicin modifies the function of human peritoneal mesothelial cells. Am. J. Transl. Res. 2016, 8, 5756. [Google Scholar]
- Alekseevna, R.V.; Pavlovich, D.A.; Evgenievich, B.Y.; Viktorovich, N.S. Nuclear factor kappa B as a potential target for pharmacological correction endothelium-associated pathology. Res. Res. Pharm. 2017, 3, 114–124. [Google Scholar]
- Arjmand, M.H. The association between visceral adiposity with systemic inflammation, oxidative stress, and risk of post-surgical adhesion. Arch. Physiol. Biochem. 2020, 6, 1–6. [Google Scholar] [CrossRef]
- Dejban, P.; Nikravangolsefid, N.; Chamanara, M.; Dehpour, A.; Rashidian, A. The role of medicinal products in the treatment of inflammatory bowel diseases (IBD) through inhibition of TLR4/NF-kappaB pathway. Phytother. Res. 2021, 35, 835–845. [Google Scholar] [CrossRef]
- De Sousa Abreu, R.; Penalva, L.O.; Marcotte, E.; Vogel, C. Global signatures of protein and mRNA expression levels. Mol. Biosyst. 2009, 5, 1512–1526. [Google Scholar] [CrossRef]
- Vogel, C.; Marcotte, E.M. Insights into the regulation of protein abundance from proteomic and transcriptomic analyses. Nat. Rev. Genet. 2012, 13, 227–232. [Google Scholar] [CrossRef]
- Schoon, H.-A.; Schoon, D.; Klug, E. Uterusbiopsien als Hilfsmittel für Diagnose und Prognose von Fertilitätsstörungen der Stute. Pferdeheilkunde 1992, 8, 355–362. [Google Scholar] [CrossRef]
- Da Costa, R.R.P.; Serrao, P.M.; Monteiro, S.; Pessa, P.; Robalo Silva, J.; Ferreira-Dias, G. Caspase-3 mediated apoptosis and cell proliferation in the equine endometrium during the oestrous cycle. Reprod. Fertil. Dev. 2007, 19, 925–932. [Google Scholar] [CrossRef]
- Gebhardt, S.; Merkl, M.; Herbach, N.; Wanke, R.; Handler, J.; Bauersachs, S. Exploration of global gene expression changes during theestrous cycle in equine endometrium. Biol. Reprod. 2012, 87, 1–13. [Google Scholar] [CrossRef]
- McDowell, K.J.; Adams, M.H.; Adam, C.Y.; Simpson, K.S. Changes in equine endometrial oestrogen receptor and progesterone receptor mRNAs during the oestrus cycle, early pregnancy and after treatment with exogenous steroids. J. Reprod. Fertil. 1999, 117, 135–142. [Google Scholar] [CrossRef]
- Honnens, A.; Weisser, S.; Welter, H.; Einspanier, R.; Bollwein, H. Relationships between uterine blood flow, peripheral sex steroids, expression of endometrial estrogen receptors and nitric oxide synthases during the estrus cycle in mares. J. Reprod. Develop. 2011, 57, 43–48. [Google Scholar] [CrossRef]
- Paech, K.; Webb, P.; Nisson, S.; Gustafsson, J.-Å.; Kushner, P.J.; Scanlan, S. Differential Ligand Activation of Estrogen Receptors ERα and ERβ at AP1 Sites. Science 1997, 177, 1508–1510. [Google Scholar] [CrossRef]
- Hall, J.M.; McDonnell, D.P. The Estrogen Receptor β-Isoform (ERβ) of the Human Estrogen Receptor Modulates ERα Transcriptional Activity and Is a Key Regulator of the Cellular Response to Estrogens and Antiestrogens. Endocrinology 1999, 140, 5566–5578. [Google Scholar] [CrossRef]
- Pettersson, K.; Grandien, K.; Kuiper, G.G.J.M.; Gustafsson, J.-Å. Mouse Estrogen Receptor β Forms Estrogen Response Element-Binding Heterodimers with Estrogen Receptor α. Mol. Endocrinol. 1997, 11, 1486–1496. [Google Scholar]
- Cowley, S.M.; Hoare, S.; Mosselman, S.; Parker, M.G. Estrogen Receptors α and β Form Heterodimers on DNA. J. Biol. Chem. 1997, 272, 19858–19862. [Google Scholar] [CrossRef]
- Lubahn, D.B.; Moyer, J.S.; Golding, T.S.; Couse, J.F.; Korach, K.S.; Smithies, O. Alteration of reproduction function but not prenatal sexual development after insertional disruption of the mouse estrogen receptor gene. Proc. Natl. Acad. Sci. USA 1993, 90, 11162–11166. [Google Scholar] [CrossRef] [PubMed]
- Pierro, E.; Minici, F.; Alesiani, O.; Miceli, F.; Proto, C.; Screpanti, I.; Lanzone, A. Stromal-epithelial interactions modulate estrogen responsiveness in normal human endometrium. Biol. Reprod. 2001, 64, 831–838. [Google Scholar] [CrossRef] [PubMed]
- Arnold, J.T.; Kaufman, D.G.; Seppala, M.; Lessey, B.A. Endometrial stromal cells regulate epithelial cell growth in vitro: A new co-culture model. Hum. Reprod. 2001, 16, 836–845. [Google Scholar] [CrossRef] [PubMed]
- Klymkowsky, M.W.; Bachant, J.B.; Domingo, A. Functions of intermediate filaments. Cell Motil. Cytoskeleton 1989, 14, 309–331. [Google Scholar] [CrossRef] [PubMed]
- Rebordão, M.R.; Amaral, A.; Lukasik, K.; Szóstek-Mioduchowska, A.; Pinto-Bravo, P.; Galvão, A.; Skarzynski, D.J.; Ferreira-Dias, G. Constituents of neutrophil extracellular traps induce in vitro collagen formation in mare endometrium. Theriogenology 2018, 113, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Brasier, A.R. The nuclear factor-B–interleukin-6 signalling pathway mediating vascular inflammation. Cardiovasc. Res. 2010, 86, 211–218. [Google Scholar] [CrossRef]
- Ohkawa, T.; Ueki, N.; Taguchi, T.; Shindo, Y.; Adachi, M.; Amuroa, Y.; Hada, T.; Higashino, K. Stimulation of hyaluronan synthesis by tumor necrosis factor- is mediated by the p50/p65 NF–B complex in MRC-5 myofibroblasts. Biochim. Biophys. Acta Mol. Cell Res. 1999, 1448, 416–424. [Google Scholar] [CrossRef][Green Version]
- Caon, I.; Bartolini, B.; Moretto, P.; Parnigoni, A.; Carava, E.; Vitale, D.L.; Alaniz, L.; Viola, M.; Karousou, E.; De Luca, G.; et al. Sirtuin 1 reduces hyaluronan synthase 2 expression by inhibiting nuclear translocation of NF-B and expression of the long-noncoding RNA HAS2–AS1. J. Biol. Chem. 2017, 295, 3485–3496. [Google Scholar] [CrossRef]
- Tong, W.; Geng, Y.; Huang, Y.; Shi, Y.; Xiang, S.; Zhang, N.; Qin, L.; Shi, Q.; Chen, Q.; Dai, K.; et al. In vivo identification and induction of articular cartilage stem cells by inhibiting NF-B signaling in osteoarthritis. Stem Cells 2015, 33, 3125–3137. [Google Scholar] [CrossRef]
- Chung, S.; Son, M.; Kim, M.; Koh, E.S.; Shin, S.J.; Park, C.W.; Kim, S.; Kim, H.S. Inhibition of p300/CBP-associated factor attenuates renal tubulointerstitial fibrosis through modulation of NF-kB and Nrf2. Int. J. Mol. Sci. 2019, 20, 1554. [Google Scholar] [CrossRef]
- Schoon, H.A.; Schoon, D.; Klug, E. Die Endometriumbiopsie bei der Stute im klinisch-gynäkologischen Kontext. Pferdeheilkunde 1997, 13, 453–464. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
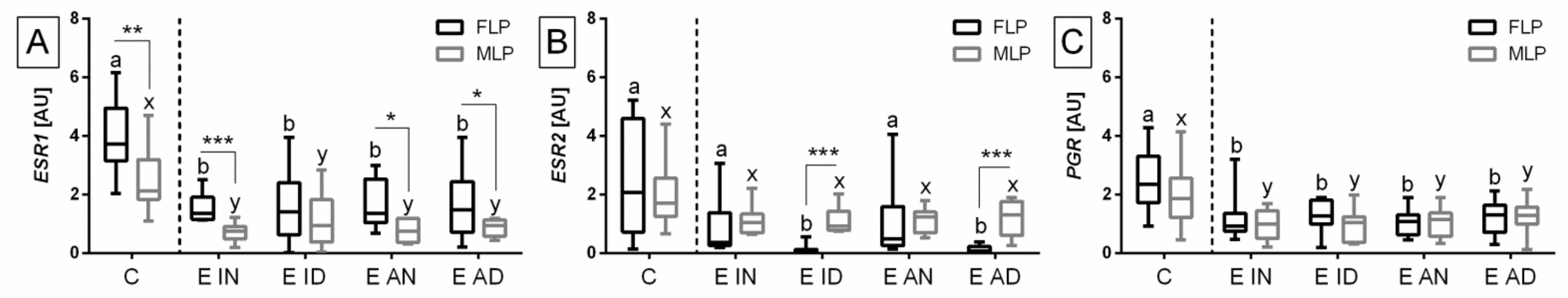{kind=link}
| RelA | NK-κB1 | NK-κB2 | MCP-1 | IL-6 | HAS1 | HAS2 | HAS3 | |
|---|---|---|---|---|---|---|---|---|
| Category I | ||||||||
| ESR1 | 0.14; 0.17 | 0.00; −0.12 | 0.01; −0.31 | −0.15; 0.37 | 0.09; −0.21 | 0.00; −0.47 | −0.05; 0.13 | −0.09; 0.24 |
| ESR2 | 0.05; 0.17 | −0.05; 0.49 | −0.09; −0.14 | −0.10; 0.46 | 0.40; −0.75 * | 0.63; 0.43 | −0.05; −0.32 | −0.40; −0.03 |
| PGR | −0.28; 0.04 | 0.10; 0.49 | 0.06; 0.39 | −0.34; −0.19 | −0.18; 0.04 | −0.09; 0.41 | 0.10; 0.02 | 0.18; −0.28 |
| Category IIa | ||||||||
| ESR1 | 0.36; 0.18 | −0.01; 0.35 | 0.30; −0.28 | −0.22; 0.39 | −0.22; 0.58 * | 0.20; 0.05 | 0.41; −0.06 | 0.11; 0.41 |
| ESR2 | 0.37; 0.12 | 0.12; −0.02 | 0.20; −0.19 | −0.47; 0.19 | −0.47; 0.05 | 0.17; 0.21 | 0.63 *; 0.09 | 0.38; 0.39 |
| PGR | 0.75 *; −0.24 | −0.04; −0.05 | 0.35; −0.52 * | −0.15; 0.41 | −0.15; 0.28 | 0.20; 0.17 | 0.37; 0.61 * | 0.27; 0.16 |
| Category IIb | ||||||||
| ESR1 | 0.48; 0.27 | −0.09; −0.09 | 0.19; −0.16 | −0.07; 0.03 | −0.30; −0.24 | −0.30; 0.03 | 0.03; 0.29 | −0.48; 0.38 |
| ESR2 | 0.28; 0.22 | −0.02; 0.43 | 0.11; 0.32 | −0.05; −0.05 | −0.22; 0.01 | −0.14; −0.02 | −0.26; −0.02 | −0.13; 0.02 |
| PGR | 0.30; −0.34 | 0.04; −0.11 | 0.14; −0.03 | 0.00; −0.34 | −0.20; 0.25 | −0.20; −0.17 | −0.13; −0.03 | −0.16; −0.64 * |
| Category III | ||||||||
| ESR1 | −0.16; 0.00 | −0.09; 0.48 | −0.15; 0.43 | 0.07; 0.16 | 0.02; 0.08 | −0.16; 0.28 | 0.01; −0.10 | −0.14; −0.07 |
| ESR2 | −0.18; −0.39 | −0.04; −0.09 | 0.15; 0.10 | −0.17; 0.10 | −0.01; 0.46 | −0.69 *; 0.21 | −0.44; 0.15 | −0.67 *; −0.16 |
| PGR | −0.40; 0.27 | −0.14; 0.09 | −0.34; 0.22 | −0.45; −0.32 | 0.31; −0.21 | −0.46 *; −0.66 * | −0.69; −0.05 | −0.64 *; 0.45 |
| RelA | NK-κB1 | NK-κB2 | MCP-1 | IL-6 | HAS1 | HAS2 | HAS3 | |
|---|---|---|---|---|---|---|---|---|
| E IN | ||||||||
| ESR1 | 0.12; −0.21 | −0.06; 0.36 | 0.58; 0.18 | −0.12; −0.12 | 0.07; 0.22 | 0.11; 0.45 | −0.50; −0.33 | −0.34; 0.56 |
| ESR2 | 0.14; −0.66 * | −0.43; −0.62 | 0.60; −0.14 | 0.10; −0.65 * | 0.29; 0.02 | 0.35; 0.12 | −0.43; 0.53 | 0.34; −0.05 |
| PGR | 0.09; 0.16 | −0.23; 0.31 | 0.77 *; −0.11 | −0.21; −0.15 | 0.04; −0.66 * | 0.09; 0.05 | 0.37; −0.65 * | 0.00; 0.35 |
| E ID | ||||||||
| ESR1 | −0.55; 0.53 * | −0.27; 0.40 * | −0.18; −0.66 | −0.18; 0.39 * | −0.18; 0.78 * | 0.76 *; −0.78 * | 0.58 *; −0.66 * | 0.58; −0.12 |
| ESR2 | 0.51; 0.19 | 0.38; 0.14 | 0.45; 0.66 | 0.45; −0.36 | 0.45; 0.55 | −0.34; 0.55 | −0.11; 0.66 | 0.11; 0.64 |
| PGR | −0.51; 0.00 | −0.06; 0.54 | −0.13; 0.03 | −0.13; 0.27 | −0.13; 0.32 | 0.27; 0.32 | −0.13; 0.03 | 0.13; −0.27 |
| E AN | ||||||||
| ESR1 | −0.12; −0.30 | 0.31; 0.00 | −0.31; −0.34 | 0.39; 0.33 | −0.31; 0.31 | −0.39; −0.11 | 0.39; −0.12 | −0.03; −0.33 |
| ESR2 | −0.21; 0.17 | 0.27; −0.69 * | −0.27; −0.48 | 0.44; 0.10 | −0.27; 0.22 | −0.44; −0.03 | 0.44; 0.50 | 0.03; −0.10 |
| PGR | 0.53; 0.06 | 0.18; 0.47 | −0.18; −0.21 | −0.37; −0.75 * | −0.18; 0.25 | 0.37; 0.20 | −0.37; 0.31 | −0.45; 0.75 * |
| E AD | ||||||||
| ESR1 | 0.23; −0.23 | −0.41; 0.36 | 0.19; −0.07 | −0.21; 0.03 | −0.37; 0.15 | 0.10; −0.16 | 0.72 *; 0.06 | 0.19; 0.48 |
| ESR2 | −0.77 *; 0.02 | −0.66 *; −0.13 | −0.23; 0.54 | 0.42; −0.09 * | −0.11; 0.72 | −0.31; −0.43 | 0.15; −0.06 | −0.15; −0.10 |
| PGR | −0.20; 0.05 | −0.78 *; 0.15 | 0.36; 0.10 | −0.07; −0.01 | −0.51; 0.04 | 0.10; 0.00 | −0.02; −0.06 | 0.28; 0.42 |
| C/I | IIa | IIb | III | Totals | |
|---|---|---|---|---|---|
| E IN | 5 | 11 | 4 | 20 | |
| E ID | 10 | 5 | 5 | 20 | |
| E AN | 12 | 5 | 3 | 20 | |
| E AD | 3 | 9 | 8 | 20 | |
| Totals | 20 | 30 | 30 | 20 | 100/80 |
| Gene | Primer Sequence |
|---|---|
| ESR1 | Forward: 5′-TCCATGGAGCACCCAGGAAAGC-3′ Reverse: 3′-CGGAGCCGAGATGACGTAGCC-5′ |
| ESR2 | Forward: 5′-TCCTGAATGCTGTGACCGAC-3′ Reverse: 3′-GTGCCTGACGTGAGAAAGGA-5′ |
| PGR | Forward: 5′-CTTCCCCGACTGCGCGTACC-3′ Reverse: 3′-TTGTGTGGCTGGAAGTCGCCG-5′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jasiński, T.; Zdrojkowski, Ł.; Ferreira-Dias, G.; Kautz, E.; Juszczuk-Kubiak, E.; Domino, M. Molecular Mechanism of Equine Endometrosis: The NF-κB-Dependent Pathway Underlies the Ovarian Steroid Receptors’ Dysfunction. Int. J. Mol. Sci. 2022, 23, 7360. https://doi.org/10.3390/ijms23137360
Jasiński T, Zdrojkowski Ł, Ferreira-Dias G, Kautz E, Juszczuk-Kubiak E, Domino M. Molecular Mechanism of Equine Endometrosis: The NF-κB-Dependent Pathway Underlies the Ovarian Steroid Receptors’ Dysfunction. International Journal of Molecular Sciences. 2022; 23(13):7360. https://doi.org/10.3390/ijms23137360
Chicago/Turabian StyleJasiński, Tomasz, Łukasz Zdrojkowski, Graça Ferreira-Dias, Ewa Kautz, Edyta Juszczuk-Kubiak, and Małgorzata Domino. 2022. "Molecular Mechanism of Equine Endometrosis: The NF-κB-Dependent Pathway Underlies the Ovarian Steroid Receptors’ Dysfunction" International Journal of Molecular Sciences 23, no. 13: 7360. https://doi.org/10.3390/ijms23137360
APA StyleJasiński, T., Zdrojkowski, Ł., Ferreira-Dias, G., Kautz, E., Juszczuk-Kubiak, E., & Domino, M. (2022). Molecular Mechanism of Equine Endometrosis: The NF-κB-Dependent Pathway Underlies the Ovarian Steroid Receptors’ Dysfunction. International Journal of Molecular Sciences, 23(13), 7360. https://doi.org/10.3390/ijms23137360