
Establishment of a Landscape of UPL5-Ubiquitinated on Multiple Subcellular Components of Leaf Senescence Cell in Arabidopsis


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
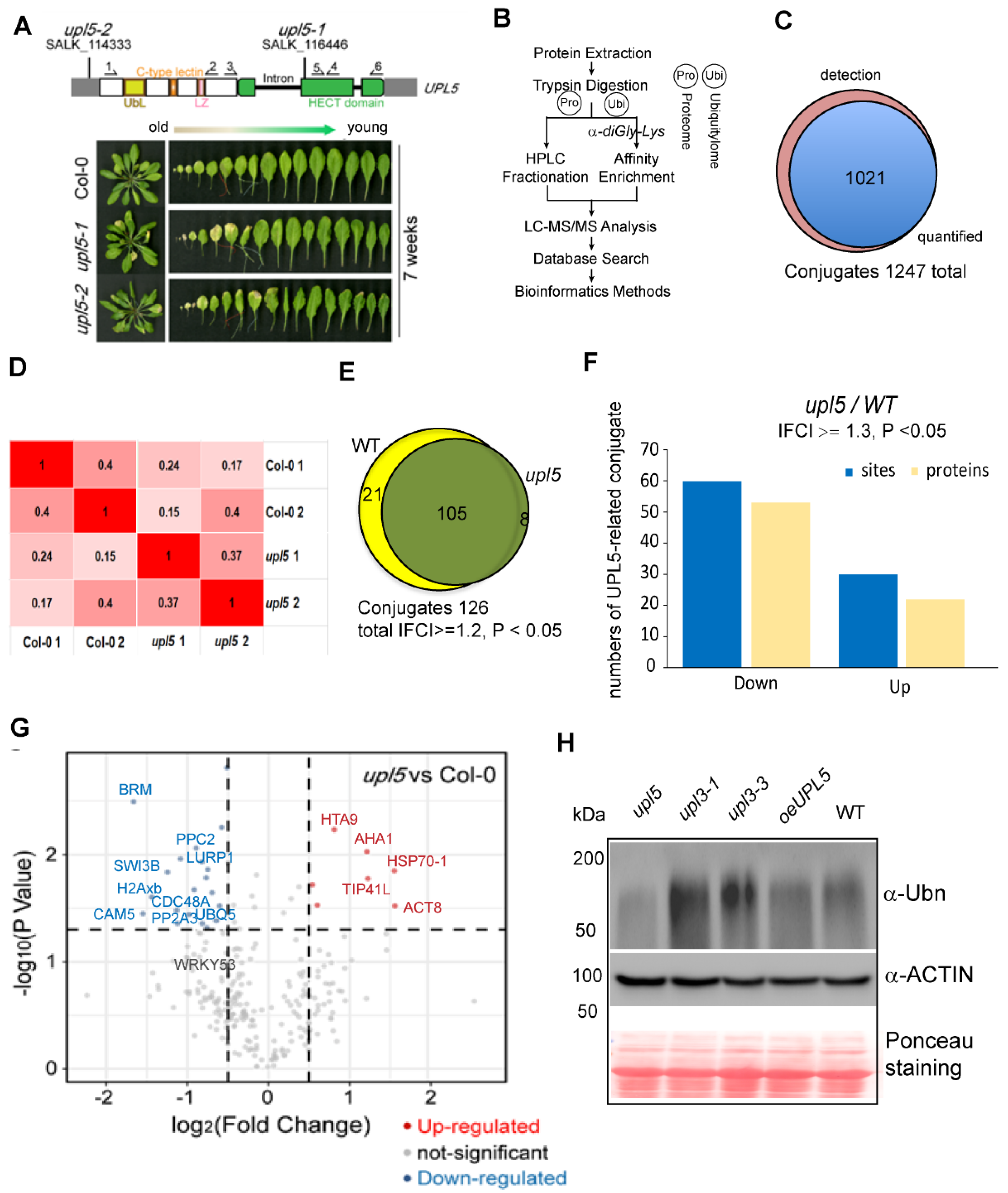
2.1. Loss of UPL5 Declines Globally Ubiquitination of Lysine Sites at Onset of Leaf Senescence
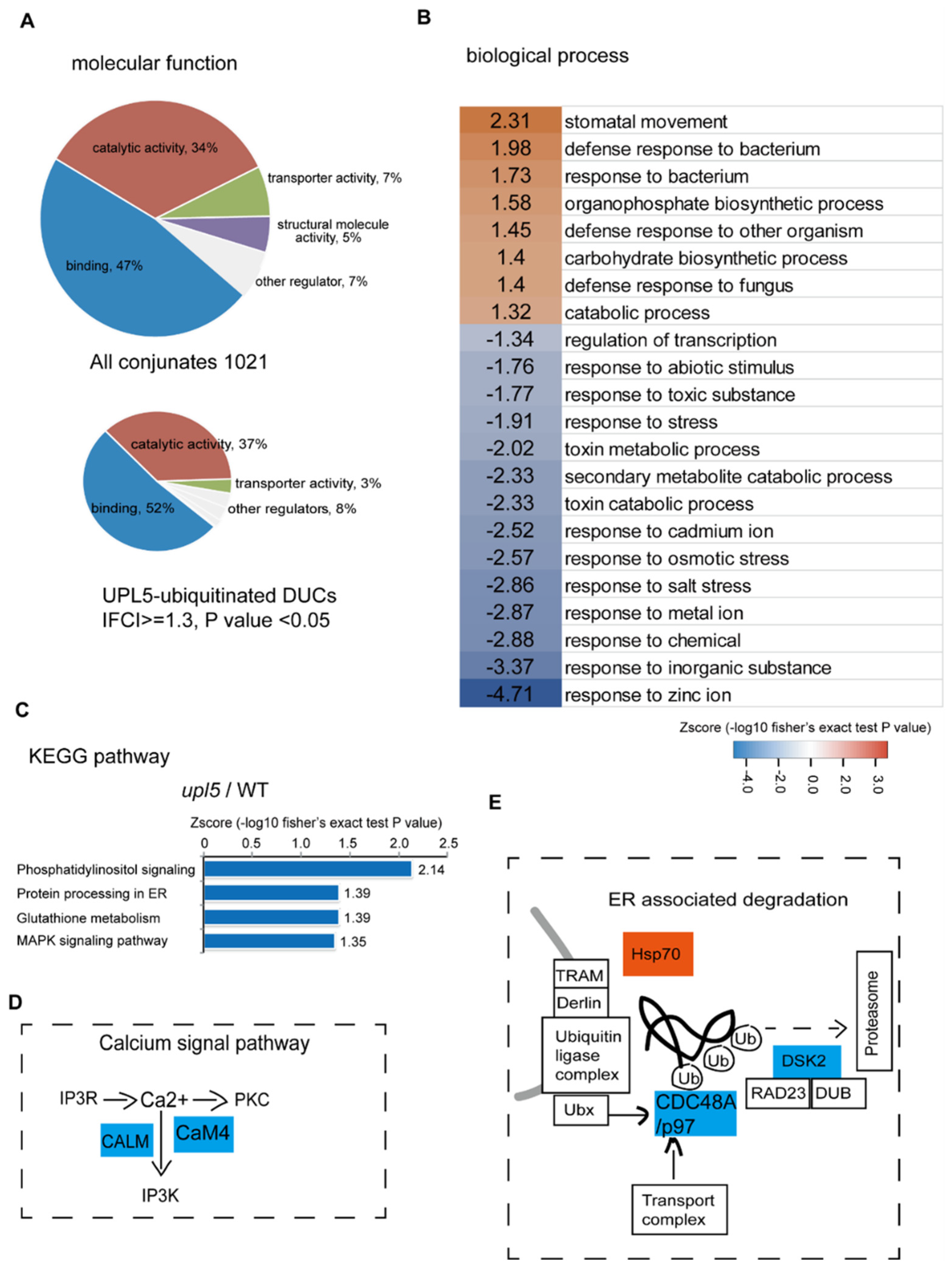
2.2. UPL5 Affects DUCs Enriched in Endoplasmic Reticulum (ER) Associated Degradation and Calcium Signal Pathway
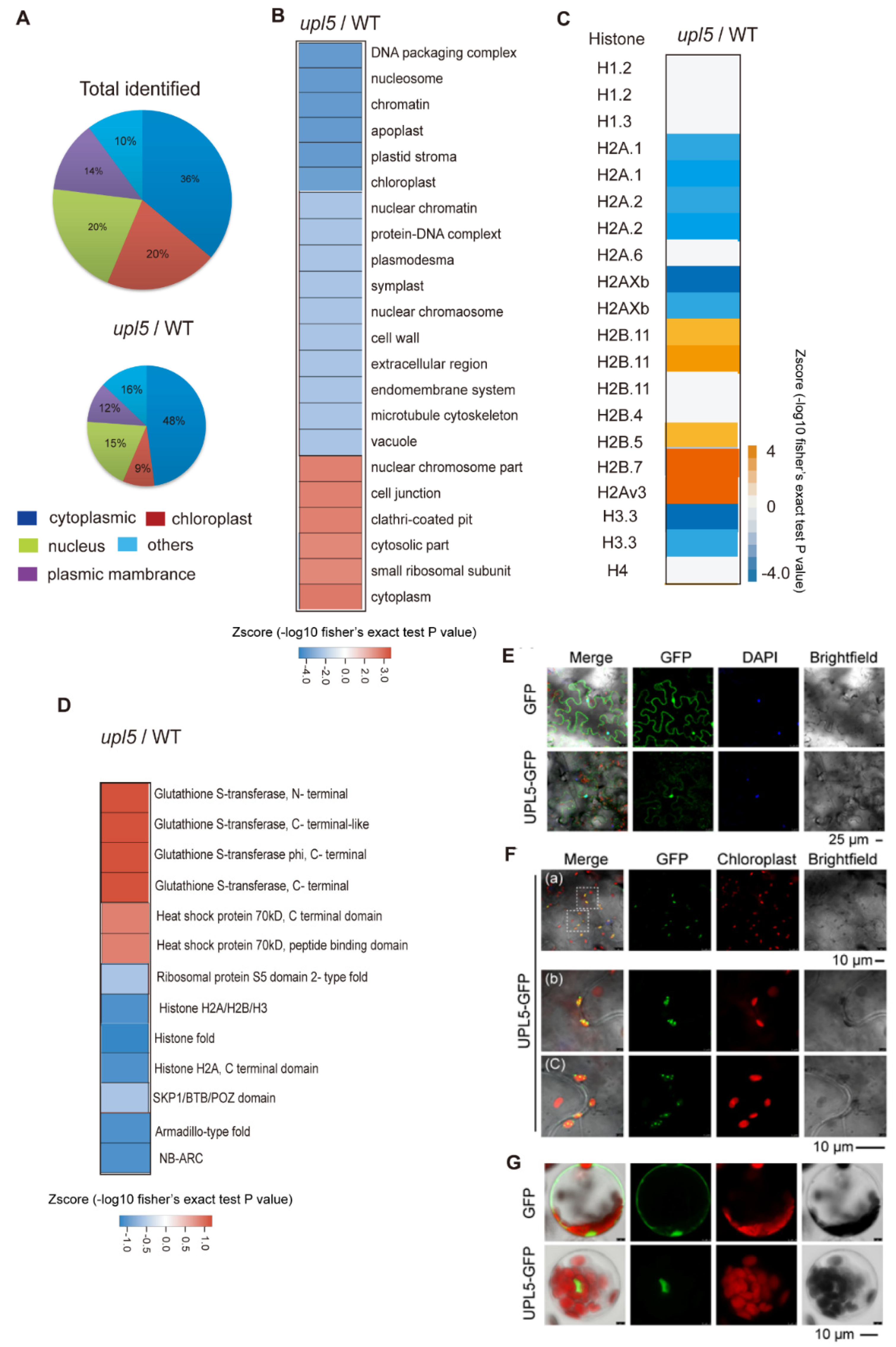
2.3. UPL5 Affects DUCs Enriched in Nucleus, Cytoplasm, and Plastids
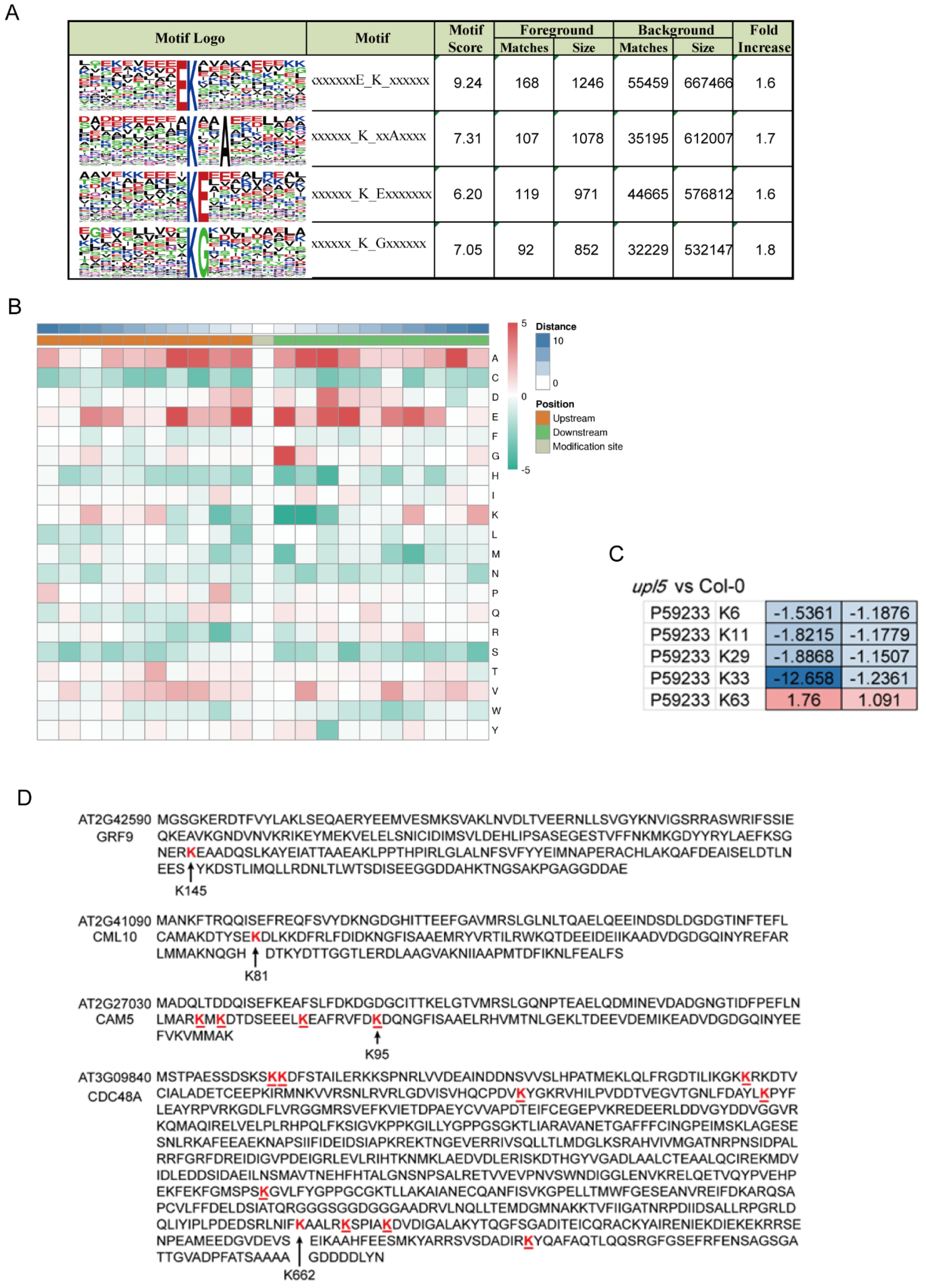
2.4. UPL5 Affects Various Patterns of Lysine Ubiquitination
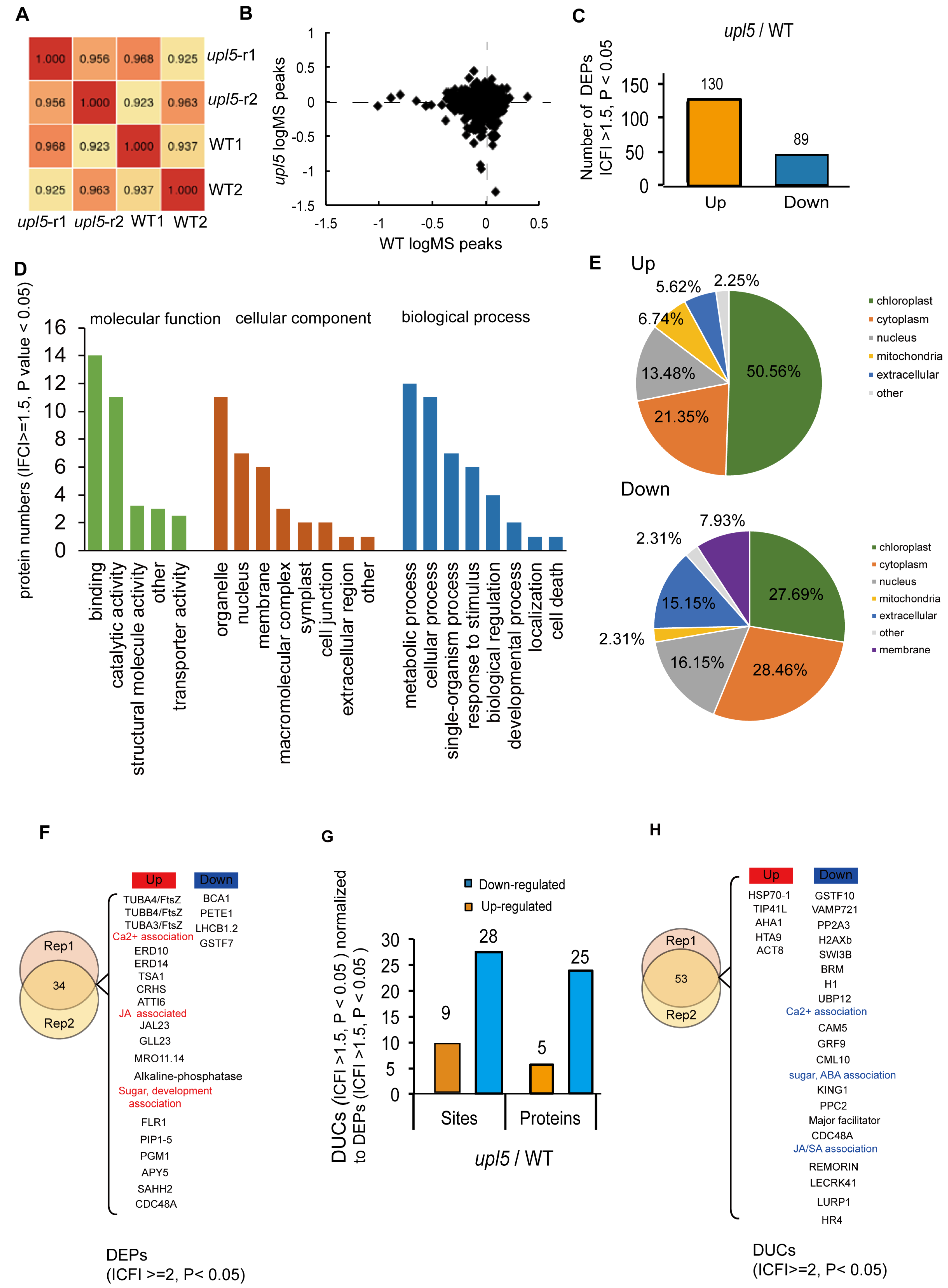
2.5. UPL5 Affects Diferentially Expressed Proteins (DEPs) Enrichment in Calcium and Hormone Signaling Pathway
3. Discussion
4. Materials and Methods
4.1. Comparative Ubiquitination Modification Proteomics
4.2. Dataset Collection
4.3. Dataset Analysis
4.4. Transient Expression in Arabidopsis Protoplast and Tobacco Leaves
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
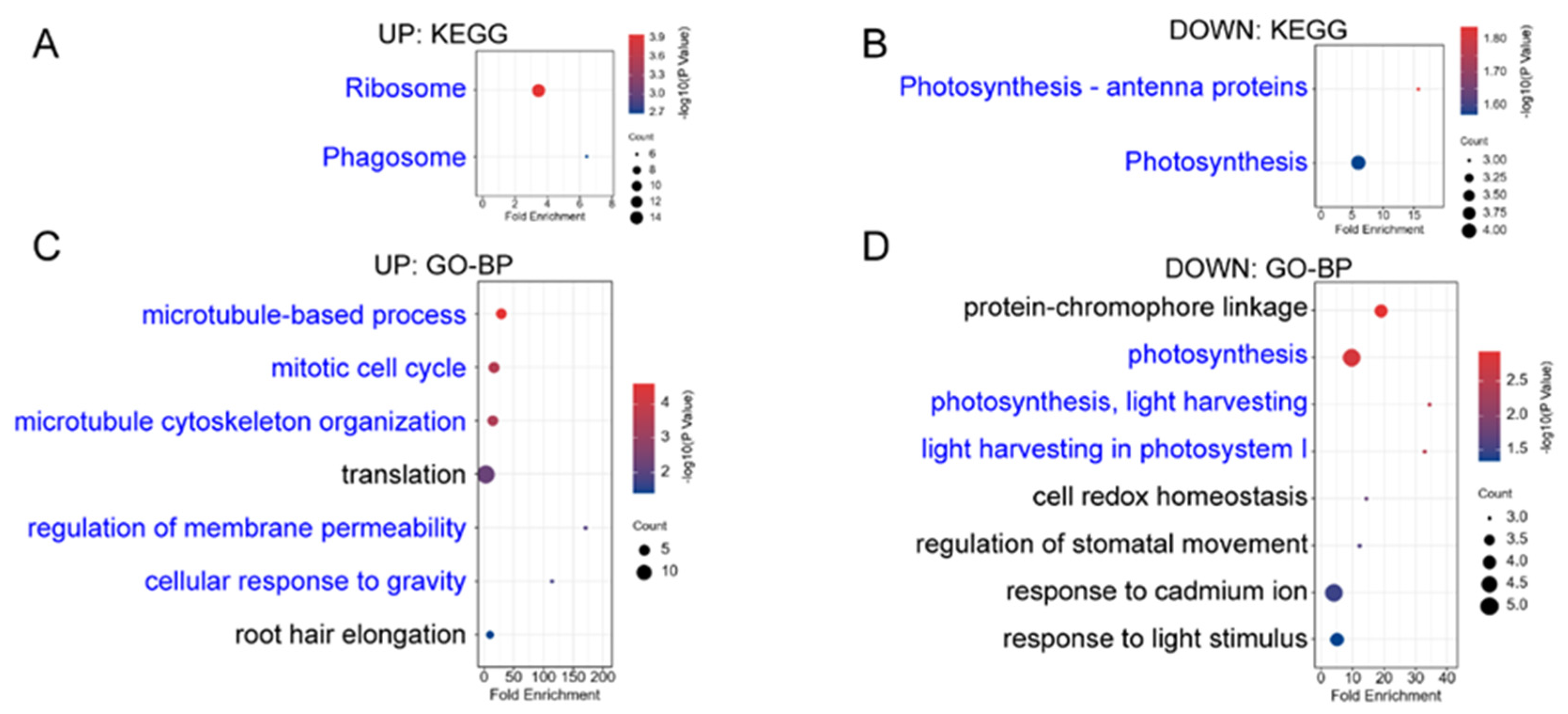
Appendix B
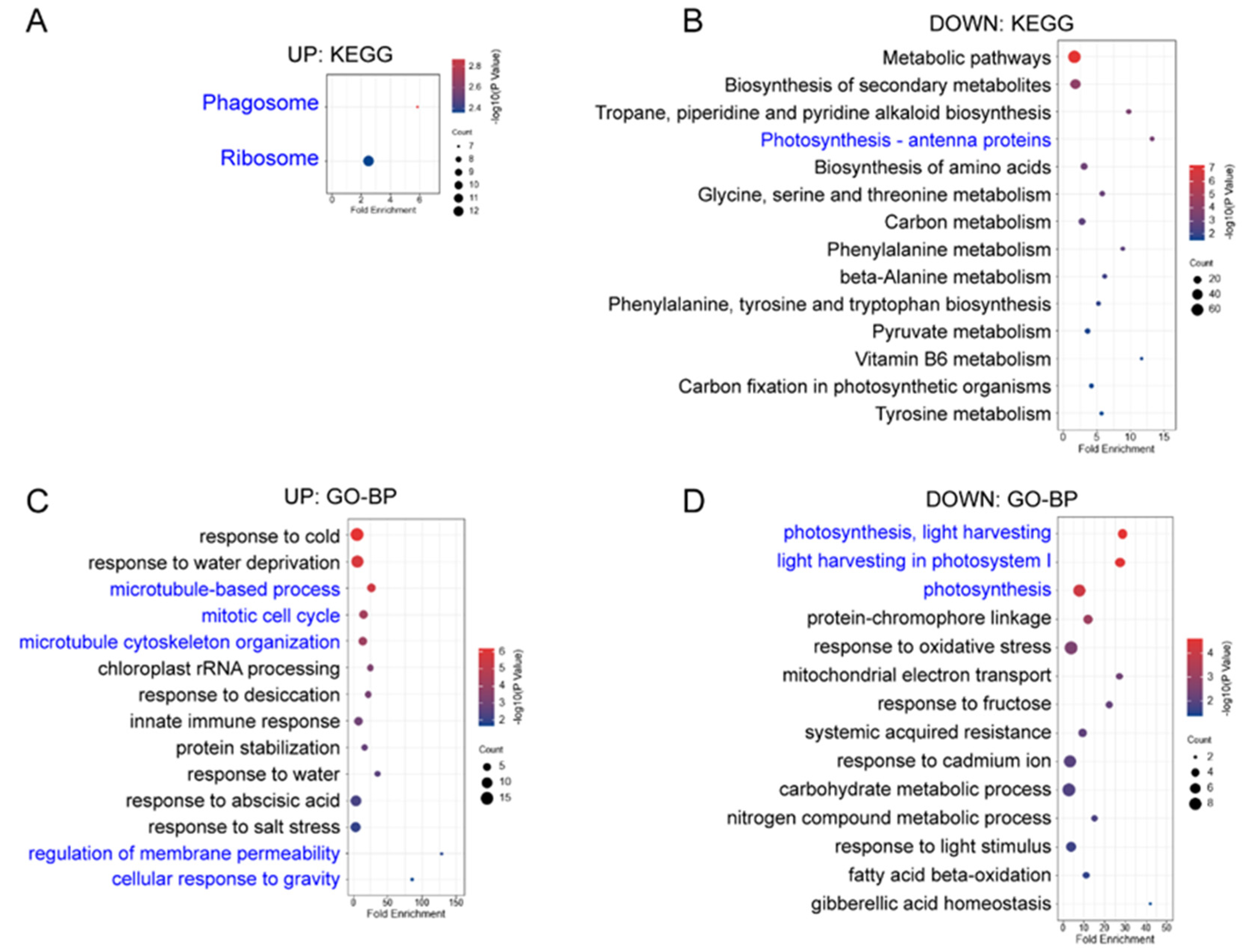
References
- Guo, Y.; Ren, G.; Zhang, K.; Li, Z.; Miao, Y.; Guo, H. Leaf Senescence: Progression, Regulation, and Application. Mol. Hortic. 2021, 1, 5. [Google Scholar] [CrossRef]
- Carrión, C.A.; Costa, M.L.; Martínez, D.E.; Mohr, C.; Humbeck, K.; Guiamet, J.J. In vivo inhibition of cysteine proteases provides evidence for the in-volvement of ‘senescence-associated vacuoles’ in chloroplast protein degradation during dark-induced senescence of to-bacco leaves. J. Exp. Bot. 2013, 64, 4967–4980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurepa, J.; Smalle, J.A. Structure, function and regulation of plant proteasomes. Biochimie 2008, 90, 324–335. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Sun, S.; Zhu, W.; Jia, K.; Yang, H.; Wang, X. Strigolactone/MAX2-Induced Degradation of Brassinosteroid Transcriptional Effector BES1 Regulates Shoot Branching. Dev. Cell 2013, 27, 681–688. [Google Scholar] [CrossRef] [Green Version]
- Chung, Y.; Kwon, S.I.; Choe, A.S. Antagonistic Regulation of Arabidopsis Growth by Brassinosteroids and Abiotic Stresses. Mol. Cells 2014, 37, 795–803. [Google Scholar] [CrossRef] [Green Version]
- Woo, H.R.; Chung, K.M.; Park, J.-H.; Oh, S.A.; Ahn, T.; Hong, S.H.; Jang, S.K.; Gil Nam, H.; Gil Nam, S.K.J. ORE9, an F-Box Protein That Regulates Leaf Senescence in Arabidopsis. Plant. Cell 2001, 13, 1779. [Google Scholar] [CrossRef] [Green Version]
- Bu, Q.; Li, H.; Zhao, Q.; Jiang, H.; Zhai, Q.; Zhang, J.; Wu, X.; Sun, J.; Xie, Q.; Wang, D.; et al. The Arabidopsis RING finger E3 ligase RHA2a is a novel positive regulator of abscisic acid sig-naling during seed germination and early seedling development. Plant Physiol. 2009, 150, 463–818. [Google Scholar] [CrossRef] [Green Version]
- Mendiondo, G.M.; Gibbs, D.J.; Szurman-Zubrzycka, M.; Korn, A.; Marquez, J.; Szarejko, I.; Maluszynski, M.; King, J.; Axcell, B.; Smart, K.; et al. Enhanced waterlogging tolerance in barley by manipulation of expression of the N-end rule pathway E3 ligase PROTEOLYSIS6. Plant Biotechnol. J. 2015, 14, 40–50. [Google Scholar] [CrossRef]
- Lan, W.; Miao, Y. New Aspects of HECT-E3 Ligases in Cell Senescence and Cell Death of Plants. Plants 2019, 8, 483. [Google Scholar] [CrossRef] [Green Version]
- Downes, B.P.; Stupar, R.M.; Gingerich, D.J.; Vierstra, R.D. The HECT ubiquitin-protein ligase (UPL) family in Arabidopsis: UPL3 has a specific role in trichome development. Plant J. 2003, 35, 729–742. [Google Scholar] [CrossRef]
- Marín, I. Evolution of Plant HECT Ubiquitin Ligases. PLoS ONE 2013, 8, e68536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miao, Y.; Zentgraf, U. The antagonist function of Arabidopsis WRKY53 and ESR/ESPin leaf senescence is modulated by the jasmonic and salicylic acid equilibrium. Plant Cell 2007, 19, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miao, Y.; Zentgraf, U. A HECT E3 ubiquitin ligase negatively regulates Arabidopsis leaf senescence through degradation of the transcription factor WRKY53. Plant J. 2010, 63, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Bresson, J.; Doll, J.; Vasseur, F.; Stahl, M.; von Roepenack-Lahaye, E.; Kilian, J.; Stadelhofer, B.; Kremer, J.M.; Kolb, D.; Wenkel, S.; et al. The genetic interaction of REVOLUTA and WRKY53 links plant development, senescence, and immune responses. PLoS ONE 2022, 17, e0254741. [Google Scholar] [CrossRef]
- Furniss, J.J.; Grey, H.; Wang, Z.; Nomoto, M.; Jackson, L.; Tada, Y.; Spoel, S.H. Proteasome-associated HECT-type ubiquitin ligase activity is required for plant im-munity. PLoS Pathog. 2018, 14, 1007447. [Google Scholar] [CrossRef] [PubMed]
- Komander, D.; Rape, M. The Ubiquitin Code. Annu. Rev. Biochem. 2012, 81, 203–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, G.; Paige, J.S.; Jaffrey, S.R. Global analysis of lysine ubiquitination by ubiquitin remnant immunoaffinity profiling. Nat. Biotechnol. 2010, 28, 868–873. [Google Scholar] [CrossRef] [Green Version]
- Han, S.-K.; Wu, M.-F.; Cui, S.; Wagner, D. Roles and activities of chromatin remodeling ATPases in plants. Plant J. 2015, 83, 62–77. [Google Scholar] [CrossRef]
- Archacki, R.; Yatusevich, R.; Buszewicz, D.; Krzyczmonik, K.; Patryn, J.; Iwanicka-Nowicka, R.; Biecek, P.; Wilczynski, B.; Koblowska, M.; Erzmanowski, A.; et al. Arabidopsis SWI/SNF chromatin remodeling complex binds both promoters and ter-minators to regulate gene expression. Nucleic Acids Res. 2017, 45, 63116–63129. [Google Scholar]
- Yu, M.; Meng, B.; Wang, F.; He, Z.; Hu, R.; Du, J.; Lai, J.; Yang, C. A Sumo ligase AtMMS21 regulates activity of the 26S proteasome in root development. Plant Sci. 2018, 280, 314–320. [Google Scholar] [CrossRef]
- Sakamoto, T.; Tsujimoto-Inui, Y.; Sotta, N.; Hirakawa, T.; Matsunaga, T.M.; Fukao, Y.; Matsunaga, S.; Fujiwara, T. Proteasomal degradation of BRAHMA promotes Boron tolerance in Arabidopsis. Nat. Commun. 2018, 9, 5285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buszewicz, D.; Archacki, R.; Palusiński, A.; Kotliński, M.; Fogtman, A.; Iwanicka-Nowicka, R.; Sosnowska, K.; Kuciński, J.; Pupel, P.; Olędzki, J.; et al. HD2C histone deacetylase and a SWI/SNF chromatin remodelling complex interact and both are involved in mediating the heat stress response in Arabidopsis. Plant. Cell Environ. 2016, 39, 2108–2122. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Yuan, L.; Yen, M.R.; Zheng, F.; Ji, R.; Peng, T.; Gu, D.; Yang, S.; Cui, Y.; Chen, P.Y.; et al. SWI3B and HDA6 interact and are required for transposon silencing in Arabidopsis. Plant J. 2020, 102, 809–822. [Google Scholar] [CrossRef] [PubMed]
- Moyet, L.; Salvi, D.; Bouchnak, I.; Miras, S.; Perrot, L.; Seigneurin-Berny, D.; Kuntz, M.; Rolland, N. Calmodulin is involved in the dual subcellular location of two chloroplast proteins. J. Biol. Chem. 2019, 294, 17543–17554. [Google Scholar] [CrossRef] [Green Version]
- Begue, H.; Jeandroz, S.; Blanchard, C.; Wendehenne, D.; Rosnoblet, C. Structure and functions of the chaperone-like p97/CDC48 in plants. Biochim. Bio-Phys. Acta Gen. Subj. 2017, 1861, 3053–3060. [Google Scholar] [CrossRef]
- Ling, Q.; Broad, W.; Trösch, R.; Töpel, M.; Sert, T.D.; Lymperopoulos, P.; Baldwin, A.; Jarvis, R.P. Ubiquitin-dependent chloroplast-associated protein degradation in plants. Science 2019, 363, eaav4467. [Google Scholar] [CrossRef]
- Li, J.; Yuan, J.; Li, Y.; Yang, W.; Zhang, W.; Lin, R. The CDC48 complex mediates ubiquitin-dependent degradation of in-tra-chloroplast proteins in plants. Cell Rep. 2022, 39, 110664. [Google Scholar] [CrossRef]
- Kikuchi, Y.; Nakamura, S.; Woodson, J.D.; Ishida, H.; Ling, Q.; Hidema, J.; Jarvis, R.P.; Hagihara, S.; Izumi, M. Chloroplast Autophagy and Ubiquitination Combine to Manage Oxidative Damage and Starvation Responses. Plant Physiol. 2020, 183, 1531–1544. [Google Scholar] [CrossRef]
- Mayta, M.L.; Hajirezaei, M.-R.; Carrillo, N.; Lodeyro, A.F. Leaf Senescence: The Chloroplast Connection Comes of Age. Plants 2019, 8, 495. [Google Scholar] [CrossRef] [Green Version]
- Domínguez, F.; Cejudo, F.J. Chloroplast dismantling in leaf senescence. J. Exp. Bot. 2021, 72, 5905–5918. [Google Scholar] [CrossRef]
- Ma, W.; Berkowitz, G.A. Cyclic nucleotide gated channel and Ca2+-mediated signal transduction during plant senescence signaling. Plant Signal Behav. 2011, 6, 413–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, C.; Lee, Y.; Lee, I.C.; Nam, H.G.; Kwak, J.M. Calmodulin 1 Regulates Senescence and ABA Response in Arabidopsis. Front. Plant Sci. 2018, 9, 803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Xing, Y.; Chang, D.; Fang, S.; Cui, B.; Li, Q.; Wang, X.; Guo, S.; Yang, X.; Men, S.; et al. CaM/BAG5/Hsc70 signaling complex dynamically regulates leaf senescence. Sci. Rep. 2016, 6, 31889. [Google Scholar] [CrossRef] [PubMed]
- Turi, Z.; Lacey, M.; Mistrik, M.; Moudry, P. Impaired ribosome biogenesis: Mechanisms and relevance to cancer and aging. Aging 2019, 11, 2512–2540. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Leister, D.; Bolle, C. Photosynthetic lesions can trigger accelerated senescence inArabidopsis thaliana. J. Exp. Bot. 2015, 66, 6891–6903. [Google Scholar] [CrossRef] [Green Version]
- Huang, D.; Lin, W.; Deng, B.; Ren, Y.; Miao, Y. Dual-Located WHIRLY1 Interacting with LHCA1 Alters Photochemical Activities of Photosystem I and Is Involved in Light Adaptation in Arabidopsis. Int. J. Mol. Sci. 2017, 18, 2352. [Google Scholar] [CrossRef] [Green Version]
- Choi, H.I.; Park, H.J.; Park, J.H.; Kim, S.; Im, M.Y.; Seo, H.H.; Kim, Y.W.; Hwang, I.; Kim, S.Y. Arabidopsis calcium-dependent protein kinase AtCPK32 interacts with ABF4, a transcrip-tional regulator of abscisic acid-responsive gene expression, and modulates its activity. Plant Physiol. 2005, 139, 1750–1761. [Google Scholar] [CrossRef] [Green Version]
- Breeze, E.; Harrison, E.; McHattie, S.; Hughes, L.; Hickman, R.; Hill, C.; Kiddle, S.; Kim, Y.-S.; Penfold, C.; Jenkins, D.; et al. High-Resolution Temporal Profiling of Transcripts during Arabidopsis Leaf Senescence Reveals a Distinct Chronology of Processes and Regulation. Plant Cell 2011, 23, 873–894. [Google Scholar] [CrossRef] [Green Version]
- Ren, Y.; Li, Y.; Jiang, Y.; Wu, B.; Miao, Y. Phosphorylation of WHIRLY1 by CIPK14 Shifts Its Localization and Dual Functions in Arabidopsis. Mol. Plant 2017, 10, 749–763. [Google Scholar] [CrossRef] [Green Version]
- Durian, G.; Sedaghatmehr, M.; Matallana-Ramirez, L.P.; Schilling, S.M.; Schaepe, S.; Guerra, T.; Herde, M.; Witte, C.P.; Mueller-Roeber, B.; Schulze, W.X.; et al. Calcium-Dependent Protein Kinase CPK1 Controls Cell Death by In Vivo Phos-phorylation of Senescence Master Regulator ORE1. Plant Cell. 2020, 32, 1610–1625. [Google Scholar] [CrossRef] [Green Version]
- Kanehisa, M.; Goto, S. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Horton, P.; Park, K.-J.; Obayashi, T.; Fujita, N.; Harada, H.; Adams-Collier, C.J.; Nakai, K. WoLF PSORT: Protein localization predictor. Nucleic Acids Res. 2007, 35, W585–W587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lan, W.; Zheng, S.; Yang, P.; Qiu, Y.; Xu, Y.; Miao, Y. Establishment of a Landscape of UPL5-Ubiquitinated on Multiple Subcellular Components of Leaf Senescence Cell in Arabidopsis. Int. J. Mol. Sci. 2022, 23, 5754. https://doi.org/10.3390/ijms23105754
Lan W, Zheng S, Yang P, Qiu Y, Xu Y, Miao Y. Establishment of a Landscape of UPL5-Ubiquitinated on Multiple Subcellular Components of Leaf Senescence Cell in Arabidopsis. International Journal of Molecular Sciences. 2022; 23(10):5754. https://doi.org/10.3390/ijms23105754
Chicago/Turabian StyleLan, Wei, Shuai Zheng, Ping Yang, Yuhao Qiu, Yun Xu, and Ying Miao. 2022. "Establishment of a Landscape of UPL5-Ubiquitinated on Multiple Subcellular Components of Leaf Senescence Cell in Arabidopsis" International Journal of Molecular Sciences 23, no. 10: 5754. https://doi.org/10.3390/ijms23105754
APA StyleLan, W., Zheng, S., Yang, P., Qiu, Y., Xu, Y., & Miao, Y. (2022). Establishment of a Landscape of UPL5-Ubiquitinated on Multiple Subcellular Components of Leaf Senescence Cell in Arabidopsis. International Journal of Molecular Sciences, 23(10), 5754. https://doi.org/10.3390/ijms23105754