
A Critical Overview of Targeted Therapies for Vestibular Schwannoma


Abstract
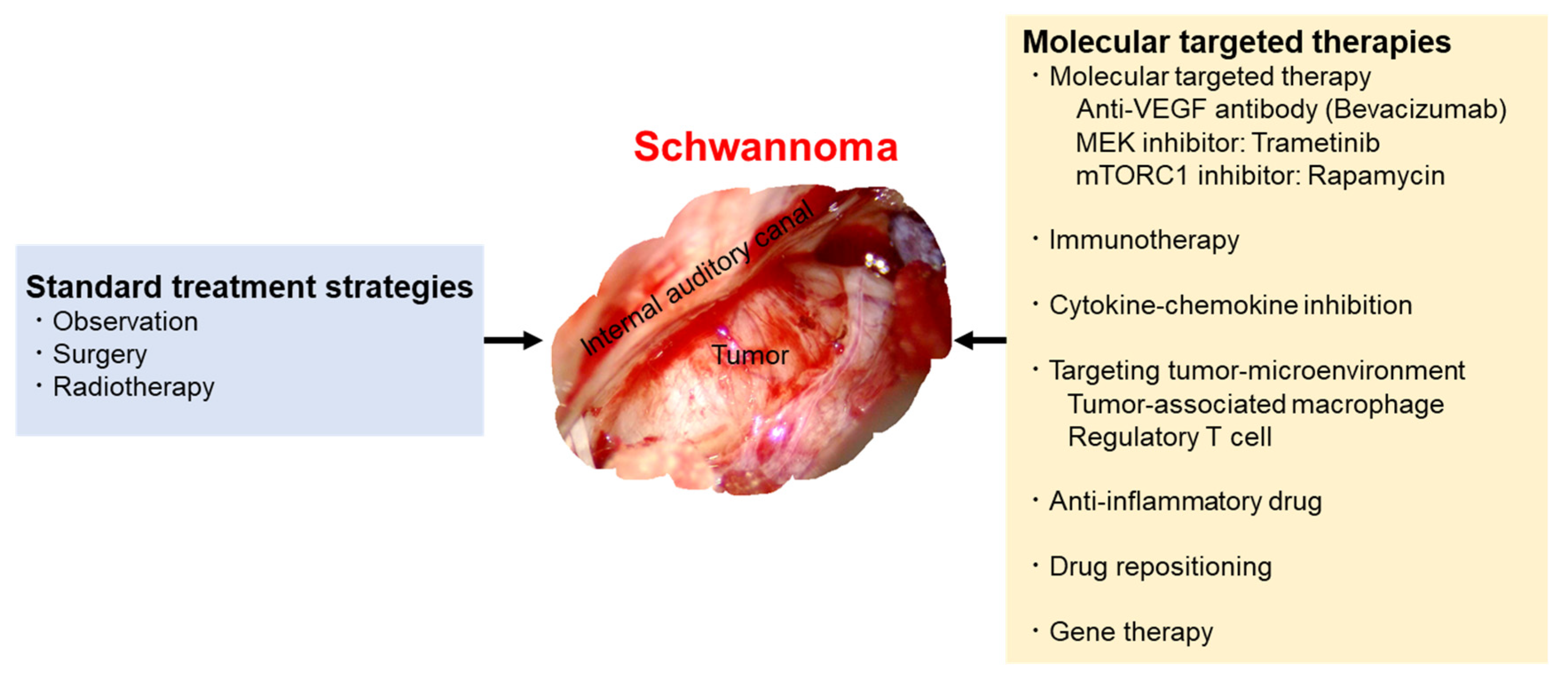
:1. Introduction
2. NF2 Gene
3. SH3PXD2A-HTRA1 Fusion
4. Protein-Kinase-Related Pathway
4.1. VEGF-A/VEGFRs
4.2. ErbB
4.3. PDGFR
4.4. HGFR
4.5. PI3K/Akt/mTOR
5. Cytokines and Chemokines
6. Tumor Microenvironment
7. Inflammation and Stress Reaction
7.1. COX2
7.2. Hsp90
8. Drug Repositioning
9. Gene Therapy
10. Ongoing Clinical Trials
11. Future Direction
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jessen, K.R.; Mirsky, R.; Lloyd, A.C. Schwann Cells: Development and Role in Nerve Repair. Cold Spring Harb. Perspect. Biol. 2015, 7, a020487. [Google Scholar] [CrossRef] [PubMed]
- Fisher, J.L.; Pettersson, D.; Palmisano, S.; Schwartzbaum, J.; Edwards, C.G.; Mathiesen, T.; Prochazka, M.; Bergenheim, T.; Florentzson, R.; Harder, H.; et al. Loud Noise Exposure and Acoustic Neuroma. Am. J. Epidemiol. 2014, 180, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Slattery, W.H. Neurofibromatosis type 2. Otolaryngol. Clin. N. Am. 2015, 48, 443–460. [Google Scholar] [CrossRef] [PubMed]
- Evans, D.G.R.; Ramsden, R.T.; Shenton, A.; Gokhale, C.; Bowers, N.L.; Huson, S.M.; Pichert, G.; Wallace, A. Mosaicism in neurofibromatosis type 2: An update of risk based on uni/bilaterality of vestibular schwannoma at presentation and sensitive mutation analysis including multiple ligation-dependent probe amplification. J. Med. Genet. 2007, 44, 424–428. [Google Scholar] [CrossRef] [PubMed]
- Yao, L.; Alahmari, M.; Temel, Y.; Hovinga, K. Therapy of Sporadic and NF2-Related Vestibular Schwannoma. Cancers 2020, 12, 835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.W.; Tu, H.T.; Chuang, C.Y.; Chang, C.S.; Chou, H.H.; Lee, M.T.; Huang, C.F. Gamma Knife radiosurgery for large vestibular schwannomas greater than 3 cm in diameter. J. Neurosurg. 2018, 128, 1380–1387. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.-Y.; Wu, Y.-Y.; Cai, X.-Y.; Fang, W.-L.; Xiao, F.-L. Molecular Diagnosis of Neurofibromatosis by Multigene Panel Testing. Front. Genet. 2021, 12, 603195. [Google Scholar] [CrossRef]
- Teranishi, Y.; Miyawaki, S.; Hongo, H.; Dofuku, S.; Okano, A.; Takayanagi, S.; Ota, T.; Yoshimura, J.; Qu, W.; Mitsui, J.; et al. Targeted deep sequencing of DNA from multiple tissue types improves the diagnostic rate and reveals a highly diverse phenotype of mosaic neurofibromatosis type 2. J. Med. Genet. 2021, 58, 701–711. [Google Scholar] [CrossRef]
- Gladden, A.B.; Hebert, A.M.; Schneeberger, E.E.; McClatchey, A.I. The NF2 Tumor Suppressor, Merlin, Regulates Epidermal Development through the Establishment of a Junctional Polarity Complex. Dev. Cell 2010, 19, 727–739. [Google Scholar] [CrossRef] [Green Version]
- Pećina-Šlaus, N. Merlin, the NF2 Gene Product. Pathol. Oncol. Res. 2013, 19, 365–373. [Google Scholar] [CrossRef]
- Santarpia, L.; Lippman, S.M.; El-Naggar, A.K. Targeting the MAPK-RAS-RAF signaling pathway in cancer therapy. Expert Opin. Ther. Targets 2012, 16, 103–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agnihotri, S.; Jalali, S.; Wilson, M.R.; Danesh, A.; Li, M.; Klironomos, G.; Krieger, J.R.; Mansouri, A.; Khan, O.; Mamatjan, Y.; et al. The genomic landscape of schwannoma. Nat. Genet. 2016, 48, 1339–1348. [Google Scholar] [CrossRef] [PubMed]
- Taule-Sivertsen, P.; Bruland, O.; Håvik, A.L.; Bratland, E.; Lund-Johansen, M.; Knappskog, P.M. The SH3PXD2A-HTRA1 fusion transcript is extremely rare in Norwegian sporadic vestibular schwannoma patients. J. Neuro-Oncol. 2021, 154, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Simons, M.; Gordon, E.; Claesson-Welsh, L. Mechanisms and regulation of endothelial VEGF receptor signalling. Nat. Rev. Mol. Cell Biol. 2016, 17, 611–625. [Google Scholar] [CrossRef] [PubMed]
- Wong, H.K.; Shimizu, A.; Kirkpatrick, N.D.; Garkavtsev, I.; Chan, A.W.; di Tomaso, E.; Klagsbrun, M.; Jain, R.K. Merlin/NF2 regulates angiogenesis in schwannomas through a Rac1/semaphorin 3F-dependent mechanism. Neoplasia 2012, 14, 84–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uesaka, T.; Shono, T.; Suzuki, S.O.; Nakamizo, A.; Niiro, H.; Mizoguchi, M.; Iwaki, T.; Sasaki, T. Expression of VEGF and its receptor genes in intracranial schwannomas. J. Neuro-Oncol. 2007, 83, 259–266. [Google Scholar] [CrossRef]
- Plotkin, S.R.; Duda, D.G.; Muzikansky, A.; Allen, J.; Blakeley, J.; Rosser, T.; Campian, J.L.; Clapp, D.W.; Fisher, M.J.; Tonsgard, J.; et al. Multicenter, Prospective, Phase II and Biomarker Study of High-Dose Bevacizumab as Induction Therapy in Patients with Neurofibromatosis Type 2 and Progressive Vestibular Schwannoma. J. Clin. Oncol. 2019, 37, 3446–3454. [Google Scholar] [CrossRef]
- Shi, J.; Lu, D.; Gu, R.; Sun, H.; Yu, L.; Pan, R.; Zhang, Y. Reliability and toxicity of bevacizumab for neurofibromatosis type 2-related vestibular schwannomas: A systematic review and meta-analysis. Am. J. Otolaryngol. 2021, 42, 103148. [Google Scholar] [CrossRef]
- Morris, K.A.; Golding, J.F.; Axon, P.R.; Afridi, S.; Blesing, C.; Ferner, R.E.; Halliday, D.; Jena, R.; Pretorius, P.M.; Evans, G.; et al. Bevacizumab in neurofibromatosis type 2 (NF2) related vestibular schwannomas: A nationally coordinated approach to delivery and prospective evaluation. Neuro-Oncol. Pract. 2016, 3, 281–289. [Google Scholar] [CrossRef] [Green Version]
- Fujii, M.; Ichikawa, M.; Iwatate, K.; Bakhit, M.; Yamada, M.; Kuromi, Y.; Sato, T.; Sakuma, J.; Saito, K. Bevacizumab Therapy of Neurofibromatosis Type 2 Associated Vestibular Schwannoma in Japanese Patients. Neurol. Med. Chir. 2020, 60, 75–82. [Google Scholar] [CrossRef] [Green Version]
- Fujii, M.; Kobayakawa, M.; Saito, K.; Inano, A.; Morita, A.; Hasegawa, M.; Mukasa, A.; Mitsuhara, T.; Goto, T.; Yamaguchi, S.; et al. Rationale and Design of BeatNF2 Trial: A Clinical Trial to Assess the Efficacy and Safety of Bevacizumab in Patients with Neurofibromatosis Type 2 Related Vestibular Schwannoma. Curr. Oncol. 2021, 28, 726–739. [Google Scholar] [CrossRef] [PubMed]
- Karajannis, M.A.; Hagiwara, M.; Schreyer, M.; Haque, S. Sustained imaging response and hearing preservation with low-dose bevacizumab in sporadic vestibular schwannoma. Neuro-Oncology 2019, 21, 822–824. [Google Scholar] [CrossRef] [PubMed]
- Tamura, R.; Tanaka, T.; Miyake, K.; Yoshida, K.; Sasaki, H. Bevacizumab for malignant gliomas: Current indications, mechanisms of action and resistance, and markers of response. Brain Tumor Pathol. 2017, 34, 62–77. [Google Scholar] [CrossRef]
- Tamura, R.; Fujioka, M.; Morimoto, Y.; Ohara, K.; Kosugi, K.; Oishi, Y.; Sato, M.; Ueda, R.; Fujiwara, H.; Hikichi, T.; et al. A VEGF receptor vaccine demonstrates preliminary efficacy in neurofibromatosis type 2. Nat. Commun. 2019, 10, 5758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wee, P.; Wang, Z. Epidermal Growth Factor Receptor Cell Proliferation Signaling Pathways. Cancers 2017, 9, 52. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, Z.K.; Brown, C.M.; Cueva, R.A.; Ryan, A.F.; Doherty, J.K. ErbB Expression, Activation, and Inhibition with Lapatinib and Tyrphostin (AG825) in Human Vestibular Schwannomas. Otol. Neurotol. 2011, 32, 841–847. [Google Scholar] [CrossRef] [Green Version]
- Doherty, J.K.; Ongkeko, W.; Crawley, B.; Andalibi, A.; Ryan, A.F. ErbB and Nrg: Potential Molecular Targets for Vestibular Schwannoma Pharmacotherapy. Otol. Neurotol. 2008, 29, 50–57. [Google Scholar] [CrossRef]
- Zhang, Y.; Long, J.; Ren, J.; Huang, X.; Zhong, P.; Wang, B. Potential Molecular Biomarkers of Vestibular Schwannoma Growth: Progress and Prospects. Front. Oncol. 2021, 11, 731441. [Google Scholar] [CrossRef]
- Clark, J.J.; Provenzano, M.; Diggelmann, H.R.; Xu, N.; Hansen, S.S.; Hansen, M.R. The ErbB inhibitors trastuzumab and erlotinib inhibit growth of vestibular schwannoma xenografts in nude mice: A preliminary study. Otol. Neurotol. 2008, 29, 846–853. [Google Scholar] [CrossRef] [Green Version]
- Ammoun, S.; Cunliffe, C.H.; Allen, J.; Chiriboga, L.; Giancotti, F.G.; Zagzag, D.; Hanemann, C.O.; Karajannis, M.A. ErbB/HER receptor activation and preclinical efficacy of lapatinib in vestibular schwannoma. Neuro-Oncology 2010, 12, 834–843. [Google Scholar] [CrossRef]
- Plotkin, S.R.; Halpin, C.; McKenna, M.J.; Loeffler, J.S.; Batchelor, T.T.; Barker, F.G., 2nd. Erlotinib for progressive vestibular schwannoma in neurofibromatosis 2 patients. Otol. Neurotol. 2010, 31, 1135–1143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salha, S.; Gehmert, S.; Brébant, V.; Anker, A.; Loibl, M.; Prantl, L.; Gehmert, S. PDGF regulated migration of mesenchymal stem cells towards malignancy acts via the PI3K signaling pathway. Clin. Hemorheol. Microcirc. 2018, 70, 543–551. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, J.; Kamnasaran, D.; Balasubramaniam, A.; Radovanovic, I.; Zadeh, G.; Kiehl, T.-R.; Guha, A. Human Schwannomas Express Activated Platelet-Derived Growth Factor Receptors and c-kit and Are Growth Inhibited by Gleevec (Imatinib Mesylate). Cancer Res. 2009, 69, 5099–5107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Dwyer, M.E.; Druker, B.J. STI571: An inhibitor of the BCR-ABL tyrosine kinase for the treatment of chronic myelogenous leukaemia. Lancet Oncol. 2000, 1, 207–211. [Google Scholar] [CrossRef]
- Yener, U.; Avsar, T.; Akgün, E.; Şeker, A.; Bayri, Y.; Kılıç, T. Assessment of antiangiogenic effect of imatinib mesylate on vestibular schwannoma tumors using in vivo corneal angiogenesis assay laboratory investigation. J. Neurosurg. 2012, 117, 697–704. [Google Scholar] [CrossRef] [PubMed]
- Ammoun, S.; Schmid, M.C.; Triner, J.; Manley, P.; Hanemann, C.O. Nilotinib alone or in combination with selumetinib is a drug candidate for neurofibromatosis type 2. Neuro-Oncology 2011, 13, 759–766. [Google Scholar] [CrossRef]
- Petrilli, A.M.; Garcia, J.; Bott, M.; Plati, S.K.; Dinh, C.T.; Bracho, O.R.; Yan, D.; Zou, B.; Mittal, R.; Telischi, F.F.; et al. Ponatinib promotes a G1 cell-cycle arrest of merlin/NF2-deficient human schwann cells. Oncotarget 2017, 8, 31666–31681. [Google Scholar] [CrossRef]
- Fu, J.; Su, X.; Li, Z.; Deng, L.; Liu, X.; Feng, X.; Peng, J. HGF/c-MET pathway in cancer: From molecular characterization to clinical evidence. Oncogene 2021, 40, 4625–4651. [Google Scholar] [CrossRef]
- Konstorum, A.; Lowengrub, J.S. Activation of the HGF/c-Met axis in the tumor microenvironment: A multispecies model. J. Theor. Biol. 2018, 439, 86–99. [Google Scholar] [CrossRef]
- Dilwali, S.; Roberts, D.; Stankovic, K.M. Interplay between VEGF-A and cMET signaling in human vestibular schwannomas and schwann cells. Cancer Biol. Ther. 2015, 16, 170–175. [Google Scholar] [CrossRef] [Green Version]
- Hu, S.-Y.; Duan, H.-F.; Li, Q.-F.; Yang, Y.-F.; Chen, J.-L.; Wang, L.-S.; Wang, H. Hepatocyte growth factor protects endothelial cells against gamma ray irradiation-induced damage. Acta Pharmacol. Sin. 2009, 30, 1415–1420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Liu, P.; Zhang, N.; Chen, J.; Landegger, L.D.; Wu, L.; Zhao, F.; Zhao, Y.; Zhang, Y.; Zhang, J.; et al. Targeting the cMET pathway augments radiation response without adverse effect on hearing in NF2 schwannoma models. Proc. Natl. Acad. Sci. USA 2018, 115, E2077–E2084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuse, M.A.; Plati, S.K.; Burns, S.S.; Dinh, C.T.; Bracho, O.; Yan, D.; Mittal, R.; Shen, R.; Soulakova, J.N.; Copik, A.J.; et al. Combination Therapy with c-Met and Src Inhibitors Induces Caspase-Dependent Apoptosis of Merlin-Deficient Schwann Cells and Suppresses Growth of Schwannoma Cells. Mol. Cancer Ther. 2017, 16, 2387–2398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Welling, D.B.; Lasak, J.M.; Akhmametyeva, E.; Ghaheri, B.; Chang, L.-S. cDNA Microarray Analysis of Vestibular Schwannomas. Otol. Neurotol. 2002, 23, 736–748. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.X.; Packer, M.D.; Huang, J.; Akhmametyeva, E.M.; Kulp, S.K.; Chen, C.-S.; Giovannini, M.; Jacob, A.; Welling, D.B.; Chang, L.-S. Growth inhibitory and anti-tumour activities of OSU-03012, a novel PDK-1 inhibitor, on vestibular schwannoma and malignant schwannoma cells. Eur. J. Cancer 2009, 45, 1709–1720. [Google Scholar] [CrossRef] [Green Version]
- Bush, M.L.; Oblinger, J.; Brendel, V.; Santarelli, G.; Huang, J.; Akhmametyeva, E.M.; Burns, S.S.; Wheeler, J.; Davis, J.; Yates, C.W.; et al. AR42, a novel histone deacetylase inhibitor, as a potential therapy for vestibular schwannomas and meningiomas. Neuro-Oncology 2011, 13, 983–999. [Google Scholar] [CrossRef] [Green Version]
- Porta, C.; Paglino, C.; Mosca, A. Targeting PI3K/Akt/mTOR Signaling in Cancer. Front. Oncol. 2014, 4, 64. [Google Scholar] [CrossRef] [Green Version]
- James, M.F.; Stivison, E.; Beauchamp, R.L.; Han, S.; Li, H.; Wallace, M.R.; Gusella, J.F.; Stemmer-Rachamimov, A.O.; Ramesh, V. Regulation of mTOR Complex 2 Signaling in Neurofibromatosis 2–Deficient Target Cell Types. Mol. Cancer Res. 2012, 10, 649–659. [Google Scholar] [CrossRef] [Green Version]
- Lane, H.A.; Wood, J.M.; McSheehy, P.M.; Allegrini, P.R.; Boulay, A.; Brueggen, J.; Littlewood-Evans, A.; Maira, S.-M.; Martiny-Baron, G.; Schnell, C.R.; et al. mTOR Inhibitor RAD001 (Everolimus) Has Antiangiogenic/Vascular Properties Distinct from a VEGFR Tyrosine Kinase Inhibitor. Clin. Cancer Res. 2009, 15, 1612–1622. [Google Scholar] [CrossRef] [Green Version]
- Goutagny, S.; Raymond, E.; Esposito-Farese, M.; Trunet, S.; Mawrin, C.; Bernardeschi, D.; Larroque, B.; Sterkers, O.; Giovannini, M.; Kalamarides, M. Phase II study of mTORC1 inhibition by everolimus in neurofibromatosis type 2 patients with growing vestibular schwannomas. J. Neuro-Oncol. 2015, 122, 313–320. [Google Scholar] [CrossRef] [Green Version]
- Karajannis, M.A.; Legault, G.; Hagiwara, M.; Giancotti, F.G.; Filatov, A.; Derman, A.; Hochman, T.; Goldberg, J.D.; Vega, E.; Wisoff, J.H.; et al. Phase II study of everolimus in children and adults with neurofibromatosis type 2 and progressive vestibular schwannomas. Neuro-Oncology 2014, 16, 292–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breun, M.; Schwerdtfeger, A.; Martellotta, D.D.; Kessler, A.F.; Perez, J.M.; Monoranu, C.M.; Ernestus, R.-I.; Matthies, C.; Löhr, M.; Hagemann, C. CXCR4: A new player in vestibular schwannoma pathogenesis. Oncotarget 2018, 9, 9940–9950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breun, M.; Monoranu, C.M.; Kessler, A.F.; Matthies, C.; Löhr, M.; Hagemann, C.; Schirbel, A.; Rowe, S.P.; Pomper, M.G.; Buck, A.K.; et al. [68Ga]-Pentixafor PET/CT for CXCR4-Mediated Imaging of Vestibular Schwannomas. Front. Oncol. 2019, 9, 503. [Google Scholar] [CrossRef] [PubMed]
- Hannan, C.J.; Lewis, D.; O’Leary, C.; Donofrio, C.A.; Evans, D.G.; Roncaroli, F.; Brough, D.; King, A.T.; Coope, D.; Pathmanaban, O.N. The inflammatory microenvironment in vestibular schwannoma. Neuro-Oncol. Adv. 2020, 2, vdaa023. [Google Scholar] [CrossRef] [PubMed]
- Fujioka, M.; Okano, H.; Ogawa, K. Inflammatory and immune responses in the cochlea: Potential therapeutic targets for sensorineural hearing loss. Front. Pharmacol. 2014, 5, 287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helbing, D.-L.; Schulz, A.; Morrison, H. Pathomechanisms in schwannoma development and progression. Oncogene 2020, 39, 5421–5429. [Google Scholar] [CrossRef] [PubMed]
- Tamura, R.; Tanaka, T.; Yamamoto, Y.; Akasaki, Y.; Sasaki, H. Dual role of macrophage in tumor immunity. Immunotherapy 2018, 10, 899–909. [Google Scholar] [CrossRef]
- Tamura, R.; Tanaka, T.; Akasaki, Y.; Murayama, Y.; Yoshida, K.; Sasaki, H. The role of vascular endothelial growth factor in the hypoxic and immunosuppressive tumor microenvironment: Perspectives for therapeutic implications. Med. Oncol. 2019, 37, 2. [Google Scholar] [CrossRef] [Green Version]
- Tamura, R.; Morimoto, Y.; Sato, M.; Kuranari, Y.; Oishi, Y.; Kosugi, K.; Yoshida, K.; Toda, M. Difference in the hypoxic immunosuppressive microenvironment of patients with neurofibromatosis type 2 schwannomas and sporadic schwannomas. J. Neuro-Oncol. 2020, 146, 265–273. [Google Scholar] [CrossRef]
- Nisenbaum, E.; Misztalm, C.; Szczupak, M.; Thielhelm, T.; Peña, S.; Mei, C.; Goncalves, S.; Bracho, O.; Ma, R.; Ivan, M.E.; et al. Tumor-Associated Macrophages in Vestibular Schwannoma and Relationship to Hearing. OTO Open. 2021, 5, 2473974X211059111. [Google Scholar] [CrossRef]
- Li, Z.; Liu, X.; Guo, R.; Wang, P. TIM-3 plays a more important role than PD-1 in the functional impairments of cytotoxic T cells of malignant Schwannomas. Tumor Biol. 2017, 39, 1010428317698352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Liechty, B.; Patel, S.; Weber, J.S.; Hollmann, T.J.; Snuderl, M.; Karajannis, M.A. Programmed death ligand 1 expression and tumor infiltrating lymphocytes in neurofibromatosis type 1 and 2 associated tumors. J. Neuro-Oncol. 2018, 138, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, J.F.; Idema, A.J.; Bol, K.F.; Nierkens, S.; Grauer, O.M.; Wesseling, P.; Grotenhuis, J.A.; Hoogerbrugge, P.M.; de Vries, I.J.; Adema, G.J. Regulatory T cells and the PD-L1/PD-1 pathway mediate immune suppression in malignant human brain tumors. Neuro Oncol. 2009, 11, 394–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakanishi, M.; Rosenberg, D.W. Multifaceted roles of PGE2 in inflammation and cancer. Semin. Immunopathol. 2013, 35, 123–137. [Google Scholar] [CrossRef] [PubMed]
- Finetti, F.; Travelli, C.; Ercoli, J.; Colombo, G.; Buoso, E.; Trabalzini, L. Prostaglandin E2 and Cancer: Insight into Tumor Progression and Immunity. Biology 2020, 9, 434. [Google Scholar] [CrossRef] [PubMed]
- Hong, B.; Krusche, C.A.; Schwabe, K.; Friedrich, S.; Klein, R.; Krauss, J.K.; Nakamura, M. Cyclooxygenase-2 Supports Tumor Proliferation in Vestibular Schwannomas. Neurosurgery 2011, 68, 1112–1117. [Google Scholar] [CrossRef]
- Kandathil, C.K.; Dilwali, S.; Wu, C.-C.; Ibrahimov, M.; McKenna, M.J.; Lee, H.; Stankovic, K.M. Aspirin Intake Correlates with Halted Growth of Sporadic Vestibular Schwannoma In Vivo. Otol. Neurotol. 2014, 35, 353–357. [Google Scholar] [CrossRef] [Green Version]
- Mackeith, S.; Wasson, J.; Baker, C.; Guilfoyle, M.; John, D.; Donnelly, N.; Mannion, R.; Jefferies, S.; Axon, P.; Tysome, J.R. Aspirin does not prevent growth of vestibular schwannomas: A case-control study. Laryngoscope 2018, 128, 2139–2144. [Google Scholar] [CrossRef]
- Whitesell, L.; Lindquist, S.L. HSP90 and the chaperoning of cancer. Nat. Rev. Cancer 2005, 5, 761–772. [Google Scholar] [CrossRef]
- Chen, Z.; Li, S.; Mo, J.; Hawley, E.; Wang, Y.; He, Y.; Brosseau, J.-P.; Shipman, T.; Clapp, D.W.; Carroll, T.J.; et al. Schwannoma development is mediated by Hippo pathway dysregulation and modified by RAS/MAPK signaling. JCI Insight 2020, 5, e141514. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.; Eskin, A.; Chareyre, F.; Jessen, W.; Manent, J.; Kawakita, M.; Chen, R.; White, C.; Vitte, J.; Jaffer, Z.M.; et al. Therapeutic Potential of HSP90 Inhibition for Neurofibromatosis Type 2. Clin. Cancer Res. 2013, 19, 3856–3870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sagers, J.E.; Brown, A.S.; Vasilijic, S.; Lewis, R.M.; Sahin, M.I.; Landegger, L.D.; Perlis, R.H.; Kohane, I.S.; Welling, D.B.; Patel, C.J.; et al. Computational repositioning and preclinical validation of mifepristone for human vestibular schwannoma. Sci. Rep. 2018, 8, 5437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakanishi, H.; Kawashima, Y.; Kurima, K.; Chae, J.J.; Ross, A.M.; Pinto-Patarroyo, G.; Patel, S.K.; Muskett, J.A.; Ratay, J.S.; Chattaraj, P.; et al. NLRP3 mutation and cochlear autoinflammation cause syndromic and nonsyndromic hearing loss DFNA34 responsive to anakinra therapy. Proc. Natl. Acad. Sci. USA 2017, 114, E7766–E7775. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Landegger, L.D.; Stankovic, K.M. Gene Therapy for Human Sensorineural Hearing Loss. Front. Cell. Neurosci. 2019, 13, 323. [Google Scholar] [CrossRef] [Green Version]
- Prabhakar, S.; Taherian, M.; Gianni, D.; Conlon, T.J.; Fulci, G.; Brockmann, J.; Stemmer-Rachamimov, A.; Sena-Esteves, M.; Breakefield, X.O.; Brenner, G.J. Regression of Schwannomas Induced by Adeno-Associated Virus-Mediated Delivery of Caspase-1. Hum. Gene Ther. 2013, 24, 152–162. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, S.G.; Abdelnabi, A.; Maguire, C.A.; Doha, M.; Sagers, J.E.; Lewis, R.M.; Muzikansky, A.; Giovannini, M.; Stemmer-Rachamimov, A.; Stankovic, K.M.; et al. Gene therapy with apoptosis-associated speck-like protein, a newly described schwannoma tumor suppressor, inhibits schwannoma growth in vivo. Neuro-Oncology 2019, 21, 854–866. [Google Scholar] [CrossRef]
- Ignacio, K.H.D.; Espiritu, A.I.; Diestro, J.D.B.; Chan, K.I.; Dmytriw, A.A.; Omar, A.T., 2nd. Efficacy of aspirin for sporadic vestibular schwannoma: A meta-analysis. Neurol. Sci. 2021, 42, 5101–5106. [Google Scholar] [CrossRef]
- Tamura, R. Current Understanding of Neurofibromatosis Type 1, 2, and Schwannomatosis. Tamura R. Int. J. Mol. Sci. 2021, 22, 5850. [Google Scholar] [CrossRef]

{kind=link}
| Targeted Pathway | |
|---|---|
| NF2 (merlin)-related pathway | |
| 1 | Ras/Raf/MEK/ERK signaling |
| 2 | PI3K/Akt/mTORC1 signaling |
| SH3PXD2A-HTRA1-fusion-related pathway | |
| 1 | MAPK signaling |
| Protein-kinase-related pathway | |
| 1 | VEGF-A/VEGFR signaling |
| 2 | ErbB family signaling |
| 3 | PDGF/PDGFR signaling |
| 4 | HGF/HGFR (c-MET) signaling |
| Cytokines and chemokines | |
| 1 | CXCL12/CXCR4 signaling |
| 2 | IL-1β, IL-6, IL-34, M-CSF, TNF-α |
| Tumor microenvironment | |
| 1 | Tumor-associated macrophage |
| 2 | Regulatory T cell |
| 3 | PD-1/PD-L1 |
| 4 | Hypoxia |
| Inflammation and stress reaction | |
| 1 | COX2 |
| 2 | Hsp90 |
| ClinicalTrials.Gov Identifier | ID | RP | EE | Age | TS |
|---|---|---|---|---|---|
| NCT01083966 | 8, 2011 | Lenox Hill Brain Tumor Center | 30 | ≥18 | Superselective intraarterial intracranial infusion of bevacizumab |
| NCT04283669 | 2, 2020 | University of Alabama at Birmingham | 19 | ≥6 | Crizotinib |
| NCT03079999 | 6, 2018 | Massachusetts Eye and Ear Infirmary | 300 | ≥12 | Aspirin |
| NCT02282917 | 9, 2015 | Massachusetts Eye and Ear | 5 | ≥18 | AR-42 (OSU-HDAC42) |
| NCT01345136 | 7, 2015 | University of California | 4 | 16–65 | Everolimus |
| NCT03095248 | 5, 2017 | Children’s Hospital Medical Center | 34 | 3–45 | Selumetinib |
| NCT04163419 | 4, 2020 | Massachusetts General Hospital | 46 | ≥18 | Tanezumab |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tamura, R.; Toda, M. A Critical Overview of Targeted Therapies for Vestibular Schwannoma. Int. J. Mol. Sci. 2022, 23, 5462. https://doi.org/10.3390/ijms23105462
Tamura R, Toda M. A Critical Overview of Targeted Therapies for Vestibular Schwannoma. International Journal of Molecular Sciences. 2022; 23(10):5462. https://doi.org/10.3390/ijms23105462
Chicago/Turabian StyleTamura, Ryota, and Masahiro Toda. 2022. "A Critical Overview of Targeted Therapies for Vestibular Schwannoma" International Journal of Molecular Sciences 23, no. 10: 5462. https://doi.org/10.3390/ijms23105462
APA StyleTamura, R., & Toda, M. (2022). A Critical Overview of Targeted Therapies for Vestibular Schwannoma. International Journal of Molecular Sciences, 23(10), 5462. https://doi.org/10.3390/ijms23105462