
Repeated and Interrupted Resistance Exercise Induces the Desensitization and Re-Sensitization of mTOR-Related Signaling in Human Skeletal Muscle Fibers

 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
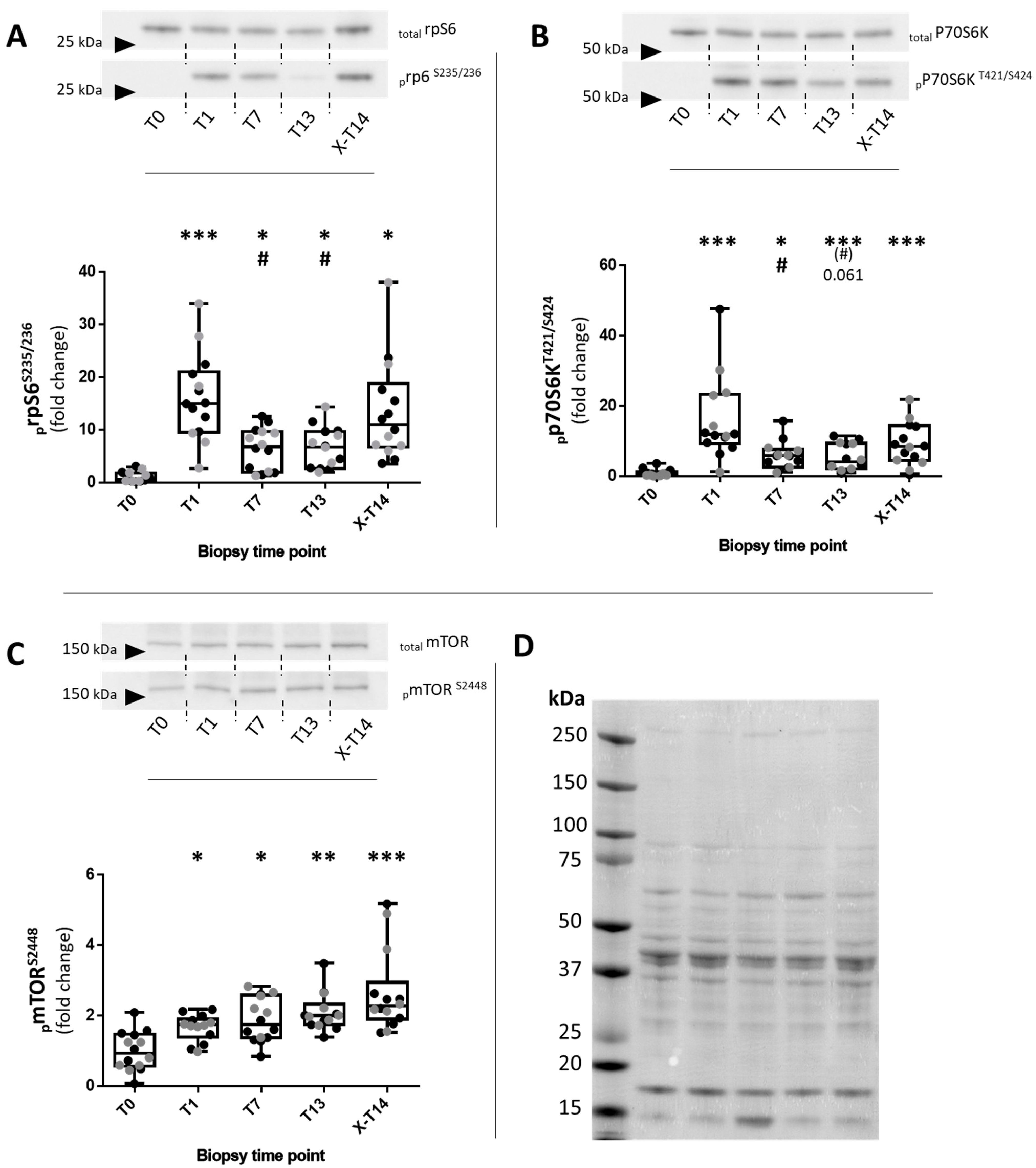
2.1. Western Blotting
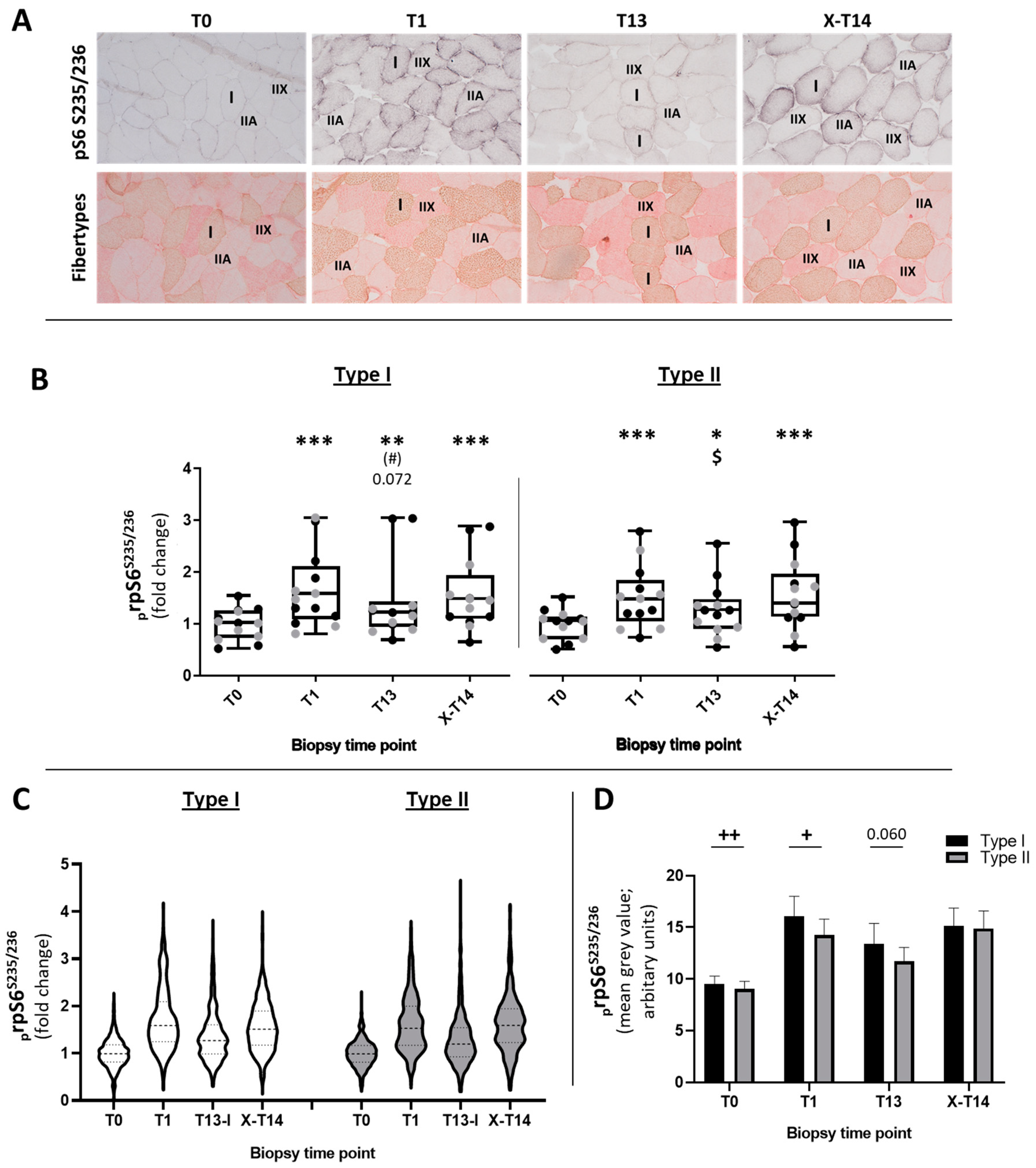
2.1.1. Fiber-Type-Specific Analysis of prpS6S235/236
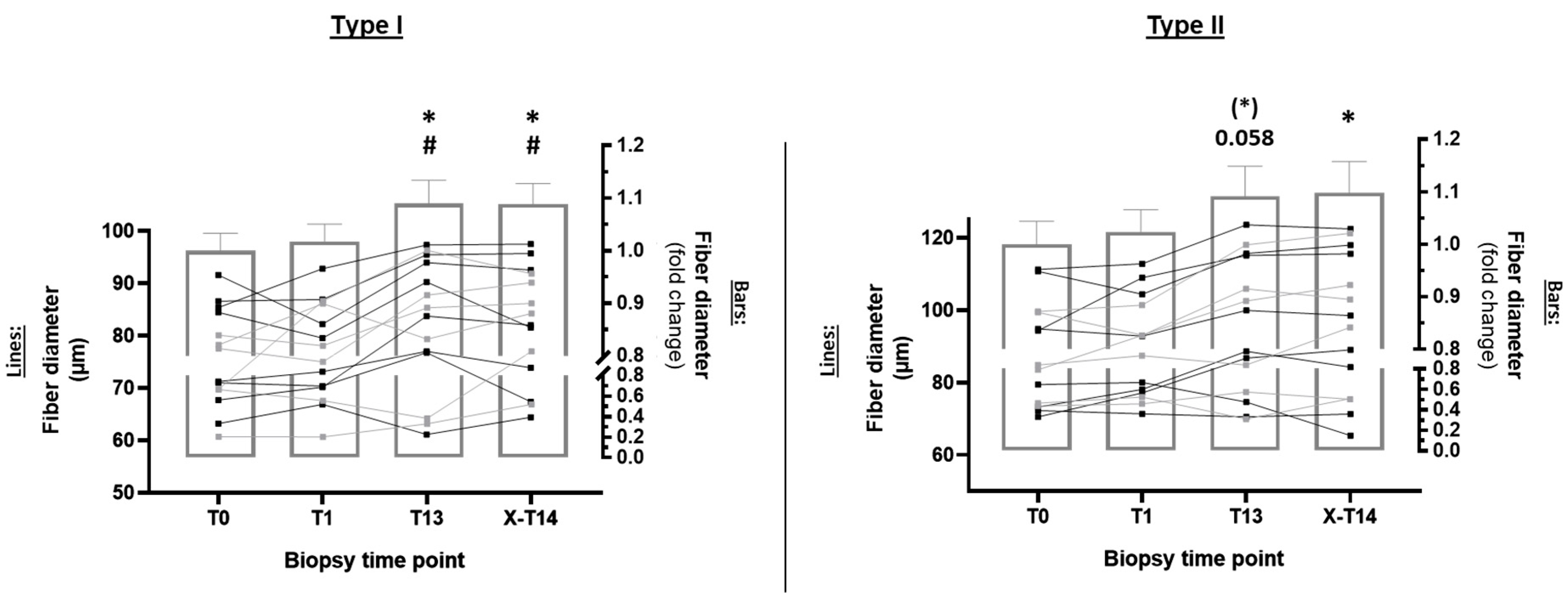
2.1.2. Type I and Type II Fiber Diameter
3. Discussion
4. Methods
4.1. Ethics Statement
4.2. Subjects
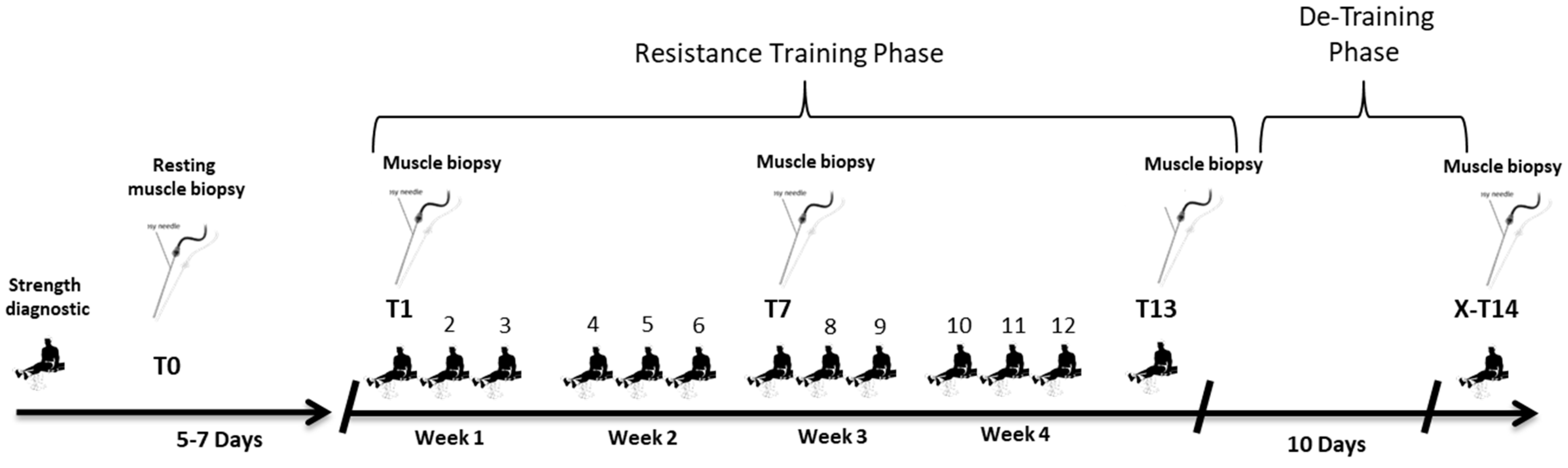
4.3. Training Intervention
4.4. Tissue Collection and Analysis
4.5. Tissue Analysis
4.5.1. Antibodies Used for Western Blotting
4.5.2. Antibodies Used for Immunohistochemistry
4.5.3. Western Blotting
4.5.4. Immunohistochemistry
4.5.5. Quantification of Sarcoplasmic pS6S235/236 Staining
4.6. Determination of Myofiber Diameter
4.7. Statistics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bull, F.C.; Al-Ansari, S.S.; Biddle, S.; Borodulin, K.; Buman, M.P.; Cardon, G.; Carty, C.; Chaput, J.-P.; Chastin, S.; Chou, R.; et al. World Health Organization 2020 Guidelines on Physical Activity and Sedentary Behaviour. Br. J. Sports Med. 2020, 54, 1451–1462. [Google Scholar] [CrossRef] [PubMed]
- Tipton, K.D.; Hamilton, D.L.; Gallagher, I.J. Assessing the Role of Muscle Protein Breakdown in Response to Nutrition and Exercise in Humans. Sports Med. 2018, 48, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Drummond, M.J.; Fry, C.S.; Glynn, E.L.; Dreyer, H.C.; Dhanani, S.; Timmerman, K.L.; Volpi, E.; Rasmussen, B.B. Rapamycin Administration in Humans Blocks the Contraction-Induced Increase in Skeletal Muscle Protein Synthesis. J. Physiol. 2009, 587, 1535–1546. [Google Scholar] [CrossRef] [PubMed]
- Inoki, K.; Zhu, T.; Guan, K.-L. TSC2 Mediates Cellular Energy Response to Control Cell Growth and Survival. Cell 2003, 115, 577–590. [Google Scholar] [CrossRef]
- Cantó, C.; Auwerx, J. PGC-1α, SIRT1 and AMPK, an Energy Sensing Network That Controls Energy Expenditure. Curr. Opin. Lipidol. 2009, 20, 98–105. [Google Scholar] [CrossRef]
- Atherton, P.J.; Babraj, J.A.; Smith, K.; Singh, J.; Rennie, M.J.; Wackerhage, H. Selective Activation of AMPK-PGC-1α or PKB-TSC2-mTOR Signaling Can Explain Specific Adaptive Responses to Endurance or Resistance Training-like Electrical Muscle Stimulation. FASEB J. 2016, 19, 1–23. [Google Scholar] [CrossRef]
- Baar, K.; Esser, K. Phosphorylation of P70(S6k) Correlates with Increased Skeletal Muscle Mass Following Resistance Exercise. Am. J. Physiol. 1999, 276, C120–C127. [Google Scholar] [CrossRef]
- Terzis, G.; Spengos, K.; Mascher, H.; Georgiadis, G.; Manta, P.; Blomstrand, E. The Degree of P70S6k and S6 Phosphorylation in Human Skeletal Muscle in Response to Resistance Exercise Depends on the Training Volume. Eur. J. Appl. Physiol. 2010, 110, 835–843. [Google Scholar] [CrossRef]
- Goodman, C.A.; Kotecki, J.A.; Jacobs, B.L.; Hornberger, T.A. Muscle Fiber Type-Dependent Differences in the Regulation of Protein Synthesis. PLoS ONE 2012, 7, e37890-11. [Google Scholar] [CrossRef]
- PROUD, C.G. Protein Phosphorylation in Translational Control. Curr. Top. Cell. Regul. 1992, 32, 243–369. [Google Scholar] [CrossRef]
- Goodman, C.A.; Pierre, P.; Hornberger, T.A. Imaging of Protein Synthesis with Puromycin. Proc. Natl. Acad. Sci. USA 2012, 109, E989. [Google Scholar] [CrossRef] [PubMed]
- Gehlert, S.; Suhr, F.; Gutsche, K.; Willkomm, L.; Kern, J.; Jacko, D.; Knicker, A.; Schiffer, T.; Wackerhage, H.; Bloch, W. High Force Development Augments Skeletal Muscle Signalling in Resistance Exercise Modes Equalized for Time under Tension. Pflug. Arch. Eur. J. Physiol. 2014, 467, 1343–1356. [Google Scholar] [CrossRef] [PubMed]
- D’Souza, R.F.; Markworth, J.F.; Figueiredo, V.C.; Gatta, P.A.D.; Petersen, A.C.; Mitchell, C.J.; Cameron-Smith, D. Dose-dependent Increases in P70S6K Phosphorylation and Intramuscular Branched-chain Amino Acids in Older Men Following Resistance Exercise and Protein Intake. Physiol. Rep. 2014, 2, e12112. [Google Scholar] [CrossRef] [PubMed]
- Kawasome, H.; Papst, P.; Webb, S.; Keller, G.M.; Johnson, G.L.; Gelfand, E.W.; Terada, N. Targeted Disruption of P70 S6k Defines Its Role in Protein Synthesis and Rapamycin Sensitivity. Proc. Natl. Acad. Sci. USA 1998, 95, 5033–5038. [Google Scholar] [CrossRef] [PubMed]
- Mayhew, D.L.; Kim, J.; Cross, J.M.; Ferrando, A.A.; Bamman, M.M. Translational Signaling Responses Preceding Resistance Training-Mediated Myofiber Hypertrophy in Young and Old Humans. J. Appl. Physiol. 2009, 107, 1655–1662. [Google Scholar] [CrossRef]
- Greenhaff, P.L.; Karagounis, L.G.; Peirce, N.; Simpson, E.J.; Hazell, M.; Layfield, R.; Wackerhage, H.; Smith, K.; Atherton, P.; Selby, A.; et al. Disassociation between the Effects of Amino Acids and Insulin on Signaling, Ubiquitin Ligases, and Protein Turnover in Human Muscle. Am. J. Physiol. Endocrinol. Metab. 2008, 295, E595–E604. [Google Scholar] [CrossRef]
- Mitchell, C.J.; Churchward-Venne, T.A.; Parise, G.; Bellamy, L.; Baker, S.K.; Smith, K.; Atherton, P.J.; Phillips, S.M. Acute Post-Exercise Myofibrillar Protein Synthesis Is Not Correlated with Resistance Training-Induced Muscle Hypertrophy in Young Men. PLoS ONE 2014, 9, e89431-7. [Google Scholar] [CrossRef]
- Terzis, G.; Georgiadis, G.; Stratakos, G.; Vogiatzis, I.; Kavouras, S.; Manta, P.; Mascher, H.; Blomstrand, E. Resistance Exercise-Induced Increase in Muscle Mass Correlates with P70S6 Kinase Phosphorylation in Human Subjects. Eur. J. Appl. Physiol. 2008, 102, 145–152. [Google Scholar] [CrossRef]
- Yoon, M.-S. MTOR as a Key Regulator in Maintaining Skeletal Muscle Mass. Front. Physiol. 2017, 8, 788. [Google Scholar] [CrossRef]
- Tannerstedt, J.; Apro, W.; Blomstrand, E. Maximal Lengthening Contractions Induce Different Signaling Responses in the Type I and Type II Fibers of Human Skeletal Muscle. J. Appl. Physiol. 2009, 106, 1412–1418. [Google Scholar] [CrossRef]
- Burd, N.A.; West, D.W.D.; Staples, A.W.; Atherton, P.J.; Baker, J.M.; Moore, D.R.; Holwerda, A.M.; Parise, G.; Rennie, M.J.; Baker, S.K.; et al. Low-Load High Volume Resistance Exercise Stimulates Muscle Protein Synthesis More Than High-Load Low Volume Resistance Exercise in Young Men. PLoS ONE 2010, 5, e12033. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, C.J.; Churchward-Venne, T.A.; West, D.W.D.; Burd, N.A.; Breen, L.; Baker, S.K.; Phillips, S.M. Resistance Exercise Load Does Not Determine Training-Mediated Hypertrophic Gains in Young Men. J. Appl. Physiol. 2012, 113, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Koopman, R. Increase in S6K1 Phosphorylation in Human Skeletal Muscle Following Resistance Exercise Occurs Mainly in Type II Muscle Fibers. Am. J. Physiol. Endocrinol. Metab. 2006, 290, E1245–E1252. [Google Scholar] [CrossRef] [PubMed]
- Ogasawara, R.; Kobayashi, K.; Tsutaki, A.; Lee, K.; Abe, T.; Fujita, S.; Nakazato, K.; Ishii, N. MTOR Signaling Response to Resistance Exercise Is Altered by Chronic Resistance Training and Detraining in Skeletal Muscle. J. Appl. Physiol. 2013, 114, 934–940. [Google Scholar] [CrossRef]
- Ulbricht, A.; Gehlert, S.; Leciejewski, B.; Schiffer, T.; Bloch, W.; Höhfeld, J. Induction and Adaptation of Chaperone-Assisted Selective Autophagy CASA in Response to Resistance Exercise in Human Skeletal Muscle. Autophagy 2015, 11, 538–546. [Google Scholar] [CrossRef] [PubMed]
- Jacko, D.; Bersiner, K.; Schulz, O.; Przyklenk, A.; Spahiu, F.; Höhfeld, J.; Bloch, W.; Gehlert, S. Coordinated Alpha-Crystallin B Phosphorylation and Desmin Expression Indicate Adaptation and Deadaptation to Resistance Exercise-Induced Loading in Human Skeletal Muscle. Am. J. Physiol. Physiol. 2020, 319, C300–C312. [Google Scholar] [CrossRef]
- Damas, F.; Phillips, S.M.; Libardi, C.A.; Vechin, F.C.; Lixandrão, M.E.; Jannig, P.R.; Costa, L.A.R.; Bacurau, A.V.; Snijders, T.; Parise, G.; et al. Resistance Training-Induced Changes in Integrated Myofibrillar Protein Synthesis Are Related to Hypertrophy Only after Attenuation of Muscle Damage. J. Physiol. 2016, 594, 5209–5222. [Google Scholar] [CrossRef]
- Collier, M.P.; Benesch, J.L.P. Small Heat-Shock Proteins and Their Role in Mechanical Stress. Cell Stress Chaperones 2020, 25, 601–613. [Google Scholar] [CrossRef]
- Paulin, D.; Li, Z. Desmin: A Major Intermediate Filament Protein Essential for the Structural Integrity and Function of Muscle. Exp. Cell Res. 2004, 301, 1–7. [Google Scholar] [CrossRef]
- Wilkinson, S.B.; Phillips, S.M.; Atherton, P.J.; Patel, R.; Yarasheski, K.E.; Tarnopolsky, M.A.; Rennie, M.J. Differential Effects of Resistance and Endurance Exercise in the Fed State on Signalling Molecule Phosphorylation and Protein Synthesis in Human Muscle. J. Physiol. 2008, 586, 3701–3717. [Google Scholar] [CrossRef]
- Figueiredo, V.C.; Caldow, M.K.; Massie, V.; Markworth, J.F.; Cameron-Smith, D.; Blazevich, A.J. Ribosome Biogenesis Adaptation in Resistance Training-Induced Human Skeletal Muscle Hypertrophy. Am. J. Physiol. Metab. 2015, 309, E72–E83. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Atherton, P.; Smith, K.; Rennie, M.J. Human Muscle Protein Synthesis and Breakdown during and after Exercise. J. Appl. Physiol. 2009, 106, 2026–2039. [Google Scholar] [CrossRef] [PubMed]
- MacDougall, J.D.; Gibala, M.J.; Tarnopolsky, M.A.; MacDonald, J.R.; Interisano, S.A.; Yarasheski, K.E. The Time Course for Elevated Muscle Protein Synthesis Following Heavy Resistance Exercise. Can. J. Appl. Physiol. 1995, 20, 480–486. [Google Scholar] [CrossRef] [PubMed]
- Miller, B.F.; Olesen, J.L.; Hansen, M.; Døssing, S.; Crameri, R.M.; Welling, R.J.; Langberg, H.; Flyvbjerg, A.; Kjaer, M.; Babraj, J.A.; et al. Coordinated Collagen and Muscle Protein Synthesis in Human Patella Tendon and Quadriceps Muscle after Exercise. J. Physiol. 2005, 567, 1021–1033. [Google Scholar] [CrossRef]
- Figueiredo, V.C.; McCarthy, J.J. Regulation of Ribosome Biogenesis in Skeletal Muscle Hypertrophy. Physiology 2019, 34, 30–42. [Google Scholar] [CrossRef]
- Lin, S.-S.; Liu, Y.-W. Mechanical Stretch Induces MTOR Recruitment and Activation at the Phosphatidic Acid-Enriched Macropinosome in Muscle Cell. Front. Cell Dev. Biol. 2019, 7, 78. [Google Scholar] [CrossRef]
- Jacobs, B.L.; Goodman, C.A.; Hornberger, T.A. The Mechanical Activation of MTOR Signaling: An Emerging Role for Late Endosome/Lysosomal Targeting. J. Muscle Res. Cell Motil. 2014, 35, 11–21. [Google Scholar] [CrossRef]
- Hornberger, T.A.; Chu, W.K.; Mak, Y.W.; Hsiung, J.W.; Huang, S.A.; Chien, S. The Role of Phospholipase D and Phosphatidic Acid in the Mechanical Activation of MTOR Signaling in Skeletal Muscle. Proc. Natl. Acad. Sci. USA 2006, 103, 4741–4746. [Google Scholar] [CrossRef]
- Lehman, N.; Ledford, B.; Fulvio, M.D.; Frondorf, K.; McPhail, L.C.; Gomez-Cambronero, J. Phospholipase D2-Derived Phosphatidic Acid Binds to and Activates Ribosomal P70 S6 Kinase Independently of MTOR. FASEB J. 2007, 21, 1075–1087. [Google Scholar] [CrossRef]
- Klossner, S.; Durieux, A.-C.; Freyssenet, D.; Flueck, M. Mechano-Transduction to Muscle Protein Synthesis Is Modulated by FAK. Eur. J. Appl. Physiol. 2009, 106, 389–398. [Google Scholar] [CrossRef]
- Martineau, L.C.; Gardiner, P.F. Insight into Skeletal Muscle Mechanotransduction: MAPK Activation Is Quantitatively Related to Tension. J. Appl. Physiol. 2001, 91, 693–702. [Google Scholar] [CrossRef] [PubMed]
- Martineau, L.C.; Gardiner, P.F. Skeletal Muscle Is Sensitive to the Tension–Time Integral but Not to the Rate of Change of Tension, as Assessed by Mechanically Induced Signaling. J. Biomech. 2002, 35, 657–663. [Google Scholar] [CrossRef]
- Martin, T.D.; Dennis, M.D.; Gordon, B.S.; Kimball, S.R.; Jefferson, L.S. MTORC1 and JNK Coordinate Phosphorylation of the P70S6K1 Autoinhibitory Domain in Skeletal Muscle Following Functional Overloading. Am. J. Physiol. Metab. 2014, 306, E1397–E1405. [Google Scholar] [CrossRef] [PubMed]
- Roux, P.P.; Richards, S.A.; Blenis, J. Phosphorylation of P90 Ribosomal S6 Kinase (RSK) Regulates Extracellular Signal-Regulated Kinase Docking and RSK Activity. Mol. Cell Biol. 2003, 23, 4796–4804. [Google Scholar] [CrossRef] [PubMed]
- Abe, Y.; Yoon, S.-O.; Kubota, K.; Mendoza, M.C.; Gygi, S.P.; Blenis, J. P90 Ribosomal S6 Kinase and P70 Ribosomal S6 Kinase Link Phosphorylation of the Eukaryotic Chaperonin Containing TCP-1 to Growth Factor, Insulin, and Nutrient Signaling. J. Biol. Chem. 2009, 284, 14939–14948. [Google Scholar] [CrossRef]
- Barash, I.A.; Peters, D.; Fridén, J.; Lutz, G.J.; Lieber, R.L. Desmin Cytoskeletal Modifications after a Bout of Eccentric Exercise in the Rat. Am. J. Physiol. Integr. Comp. Physiol. 2002, 283, R958–R963. [Google Scholar] [CrossRef]
- Meyer, G.A.; Kiss, B.; Ward, S.R.; Morgan, D.L.; Kellermayer, M.S.Z.; Lieber, R.L. Theoretical Predictions of the Effects of Force Transmission by Desmin on Intersarcomere Dynamics. Biophys. J. 2010, 98, 258–266. [Google Scholar] [CrossRef][Green Version]
- Jakob, U.; Gaestel, M.; Engel, K.; Buchner, J. Small Heat Shock Proteins Are Molecular Chaperones. J. Biol. Chem. 2001, 268, 1517–1520. [Google Scholar] [CrossRef]
- Boppart, M.D. 7beta1-Integrin Regulates Mechanotransduction and Prevents Skeletal Muscle Injury. Am. J. Physiol. Physiol. 2006, 290, C1660–C1665. [Google Scholar] [CrossRef]
- Camera, D.M.; EDGE, J.; SHORT, M.J.; Hawley, J.A.; Coffey, V.G. Early Time Course of Akt Phosphorylation after Endurance and Resistance Exercise. Med. Sci. Sports Exerc. 2010, 42, 1843–1852. [Google Scholar] [CrossRef]
- Höhfeld, J.; Benzing, T.; Bloch, W.; Fürst, D.O.; Gehlert, S.; Hesse, M.; Hoffmann, B.; Hoppe, T.; Huesgen, P.F.; Köhn, M.; et al. Maintaining Proteostasis under Mechanical Stress. EMBO Rep. 2021, 22, e52507. [Google Scholar] [CrossRef] [PubMed]
- Vijayan, K.; Thompson, J.L.; Norenberg, K.M.; Fitts, R.H.; Riley, D.A. Fiber-Type Susceptibility to Eccentric Contraction-Induced Damage of Hindlimb-Unloaded Rat AL Muscles. J. Appl. Physiol. 2001, 90, 770–776. [Google Scholar] [CrossRef] [PubMed]
- Brancaccio, P.; Lippi, G.; Maffulli, N. Biochemical Markers of Muscular Damage. Clin. Chem. Lab. Med. 2010, 48, 757–767. [Google Scholar] [CrossRef] [PubMed]
- Klimek, C.; Kathage, B.; Wördehoff, J.; Höhfeld, J. BAG3-Mediated Proteostasis at a Glance. J. Cell Sci. 2017, 130, 2781–2788. [Google Scholar] [CrossRef]
- Arndt, V.; Dick, N.; Tawo, R.; Dreiseidler, M.; Wenzel, D.; Hesse, M.; Fürst, D.O.; Saftig, P.; Saint, R.; Fleischmann, B.K.; et al. Chaperone-Assisted Selective Autophagy Is Essential for Muscle Maintenance. Curr. Biol. 2010, 20, 143–148. [Google Scholar] [CrossRef]
- Smith, D.A.; Carland, C.R.; Guo, Y.; Bernstein, S.I. Getting Folded: Chaperone Proteins in Muscle Development, Maintenance and Disease. Anat. Rec. 2014, 297, 1637–1649. [Google Scholar] [CrossRef]
- Kathage, B.; Gehlert, S.; Ulbricht, A.; Lüdecke, L.; Tapia, V.E.; Orfanos, Z.; Wenzel, D.; Bloch, W.; Volkmer, R.; Fleischmann, B.K.; et al. The Cochaperone BAG3 Coordinates Protein Synthesis and Autophagy under Mechanical Strain through Spatial Regulation of MTORC1. BBA Mol. Cell Res. 2017, 1864, 62–75. [Google Scholar] [CrossRef]
- Roux, P.P.; Shahbazian, D.; Vu, H.; Holz, M.K.; Cohen, M.S.; Taunton, J.; Sonenberg, N.; Blenis, J. RAS/ERK Signaling Promotes Site-Specific Ribosomal Protein S6 Phosphorylation via RSK and Stimulates Cap-Dependent Translation. J. Biol. Chem. 2007, 282, 14056–14064. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, Z. IRES-Mediated Cap-Independent Translation, a Path Leading to Hidden Proteome. J. Mol. Cell Biol 2019, 11, 911–919. [Google Scholar] [CrossRef]
- Péladeau, C.; Jasmin, B.J. Targeting IRES-Dependent Translation as a Novel Approach for Treating Duchenne Muscular Dystrophy. RNA Biol. 2020, 18, 1–14. [Google Scholar] [CrossRef]
- Makhnovskii, P.A.; Zgoda, V.G.; Bokov, R.O.; Shagimardanova, E.I.; Gazizova, G.R.; Gusev, O.A.; Lysenko, E.A.; Kolpakov, F.A.; Vinogradova, O.L.; Popov, D.V. Regulation of Proteins in Human Skeletal Muscle: The Role of Transcription. Sci. Rep. 2020, 10, 3514. [Google Scholar] [CrossRef] [PubMed]
- Jacko, D.; Bersiner, K.; Hebchen, J.; de Marées, M.; Bloch, W.; Gehlert, S. Phosphorylation of AB-Crystallin and Its Cytoskeleton Association Differs in Skeletal Myofiber Types Depending on Resistance Exercise Intensity and Volume. J. Appl. Physiol. 2019, 126, 1607–1618. [Google Scholar] [CrossRef] [PubMed]
- Duchateau, J.; Hainaut, K. Effects of Immobilization on Contractile Properties, Recruitment and Firing Rates of Human Motor Units. J. Physiol. 1990, 422, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Beltman, J.G.M.; de Haan, A.; Haan, H.; Gerrits, H.L.; van Mechelen, W.; Sargeant, A.J. Metabolically Assessed Muscle Fibre Recruitment in Brief Isometric Contractions at Different Intensities. Eur. J. Appl. Physiol. 2004, 92, 485–492. [Google Scholar] [CrossRef]
- Enoka, R.M. Eccentric Contractions Require Unique Activation Strategies by the Nervous System. J. Appl. Physiol. 1996, 81, 2339–2346. [Google Scholar] [CrossRef]
- Signorile, J.F.; Weber, B.; Roll, B.; Caruso, J.F.; Lowensteyn, I.; Perry, A.C. An Electromyographical Comparison of the Squat and Knee Extension Exercises. J. Strength Cond. Res. 1994, 8, 178. [Google Scholar] [CrossRef]
- Padulo, J.; Migliaccio, G.; Ardigò, L.; Leban, B.; Cosso, M.; Samozino, P. Lower Limb Force, Velocity, Power Capabilities during Leg Press and Squat Movements. Int. J. Sports Med. 2017, 38, 1083–1089. [Google Scholar] [CrossRef]
- Padulo, J.; Laffaye, G.; Chamari, K.; Concu, A. Concentric and Eccentric: Muscle Contraction or Exercise? Sports Health A Multidiscip. Approach 2013, 5, 306. [Google Scholar] [CrossRef]
- Bergström, J. Percutaneous Needle Biopsy of Skeletal Muscle in Physiological and Clinical Research. Scand. J. Clin. Lab. Investig. 1975, 35, 609–616. [Google Scholar] [CrossRef]
- Andersen, J.L.; Aagaard, P. Myosin Heavy Chain IIX Overshoot in Human Skeletal Muscle. Muscle Nerve 2000, 23, 1095–1104. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jacko, D.; Schaaf, K.; Masur, L.; Windoffer, H.; Aussieker, T.; Schiffer, T.; Zacher, J.; Bloch, W.; Gehlert, S. Repeated and Interrupted Resistance Exercise Induces the Desensitization and Re-Sensitization of mTOR-Related Signaling in Human Skeletal Muscle Fibers. Int. J. Mol. Sci. 2022, 23, 5431. https://doi.org/10.3390/ijms23105431
Jacko D, Schaaf K, Masur L, Windoffer H, Aussieker T, Schiffer T, Zacher J, Bloch W, Gehlert S. Repeated and Interrupted Resistance Exercise Induces the Desensitization and Re-Sensitization of mTOR-Related Signaling in Human Skeletal Muscle Fibers. International Journal of Molecular Sciences. 2022; 23(10):5431. https://doi.org/10.3390/ijms23105431
Chicago/Turabian StyleJacko, Daniel, Kirill Schaaf, Lukas Masur, Hannes Windoffer, Thorben Aussieker, Thorsten Schiffer, Jonas Zacher, Wilhelm Bloch, and Sebastian Gehlert. 2022. "Repeated and Interrupted Resistance Exercise Induces the Desensitization and Re-Sensitization of mTOR-Related Signaling in Human Skeletal Muscle Fibers" International Journal of Molecular Sciences 23, no. 10: 5431. https://doi.org/10.3390/ijms23105431
APA StyleJacko, D., Schaaf, K., Masur, L., Windoffer, H., Aussieker, T., Schiffer, T., Zacher, J., Bloch, W., & Gehlert, S. (2022). Repeated and Interrupted Resistance Exercise Induces the Desensitization and Re-Sensitization of mTOR-Related Signaling in Human Skeletal Muscle Fibers. International Journal of Molecular Sciences, 23(10), 5431. https://doi.org/10.3390/ijms23105431