
Genome-Wide Analysis Identified a Set of Conserved lncRNAs Associated with Domestication-Related Traits in Rice

 and
and
Abstract
1. Introduction
2. Results
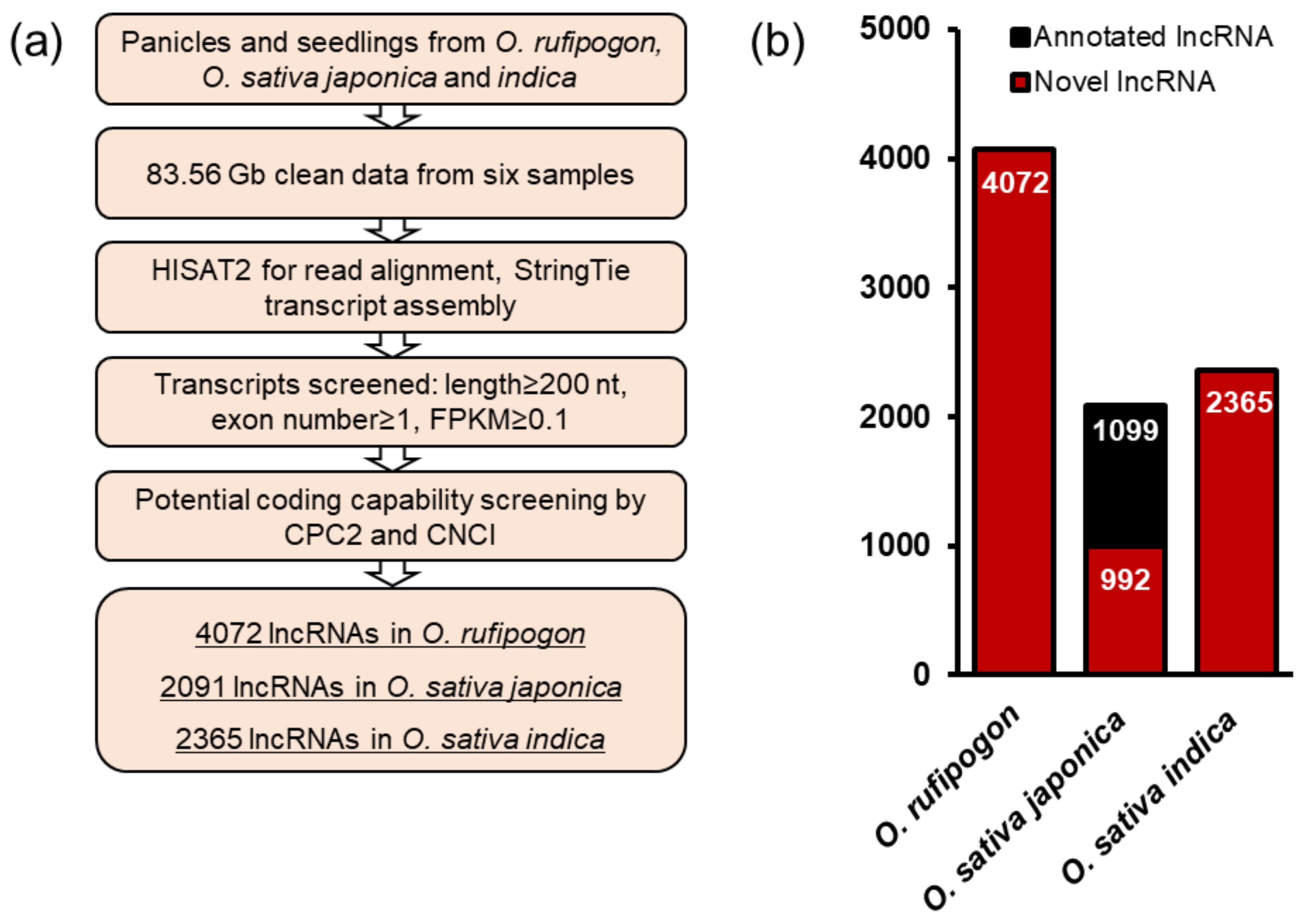
2.1. Transcriptome-Wide Analysis of lncRNAs in Wild Rice and Cultivated Rice
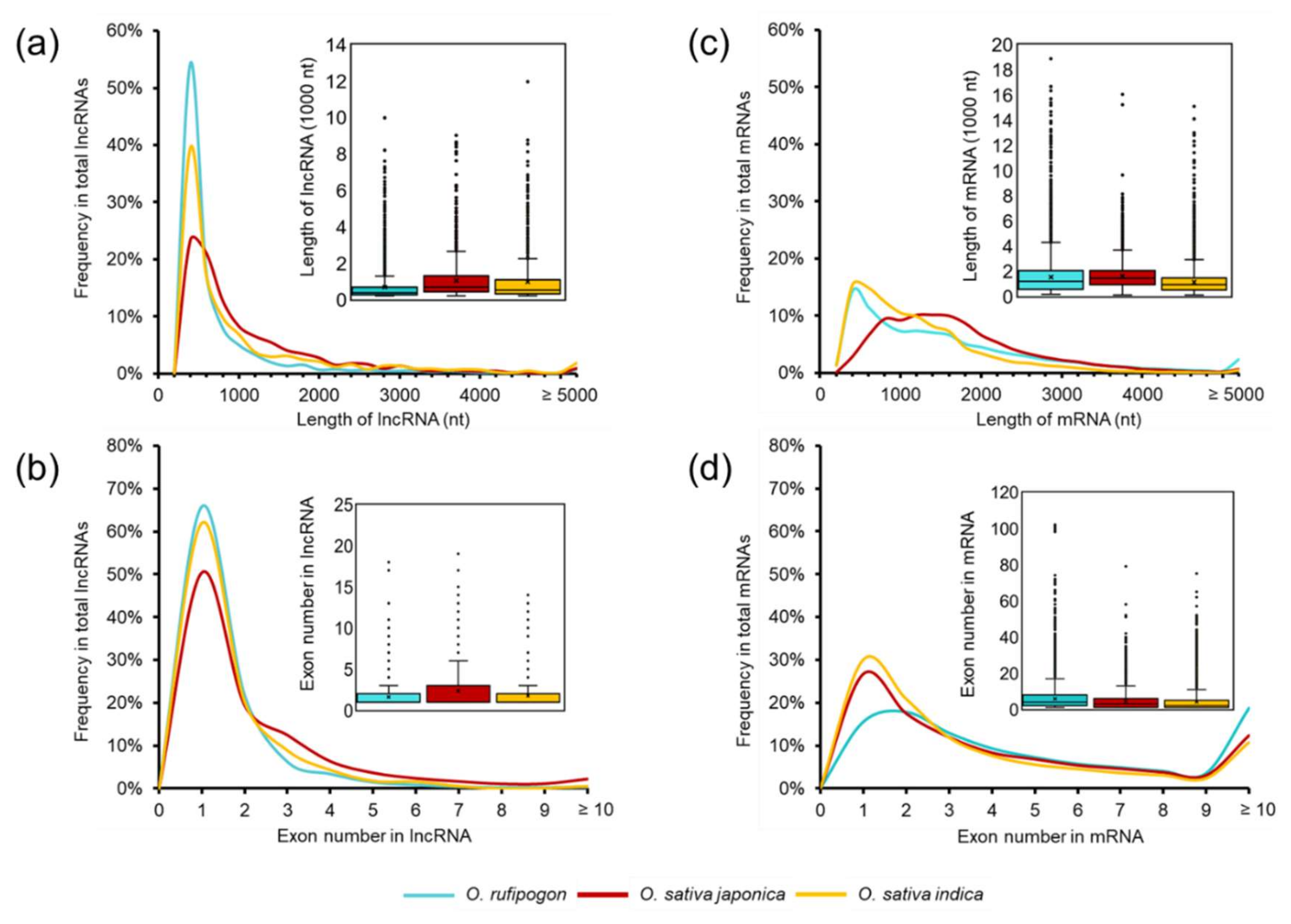
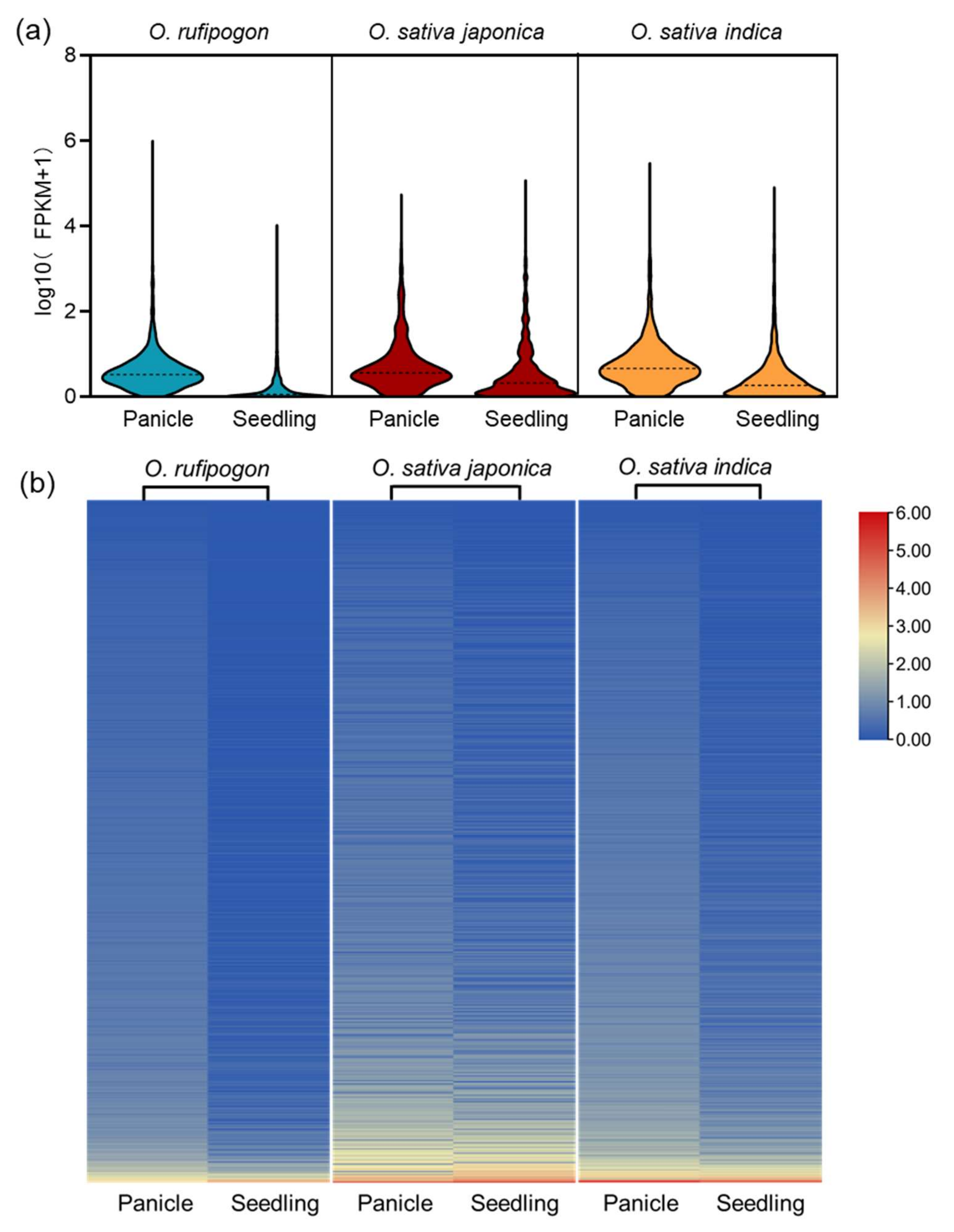
2.2. Characteristics Comparison of lncRNAs in Wild and Cultivated Rice
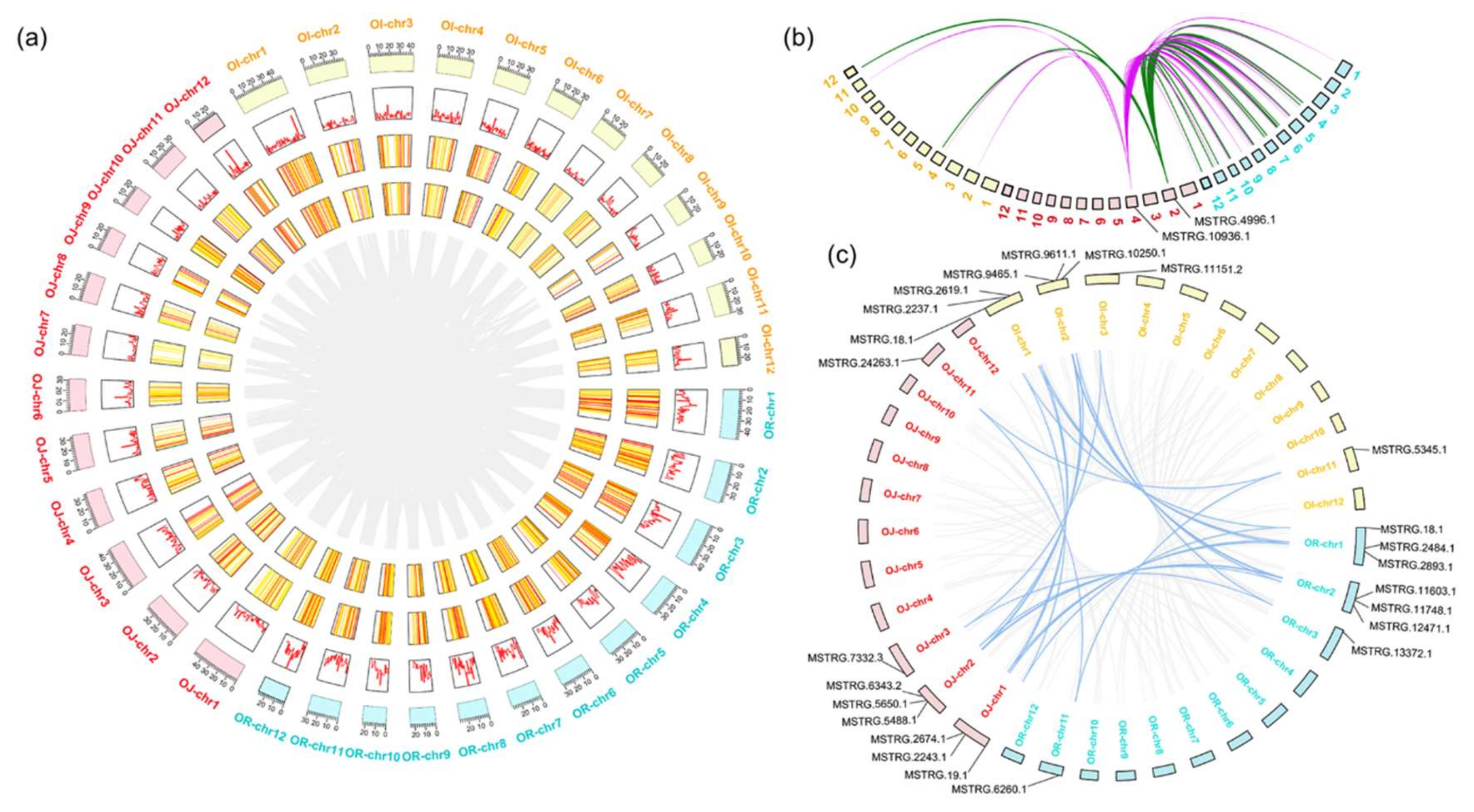
2.3. Conservation Analysis of lncRNAs That may be Associated with Rice Domestication
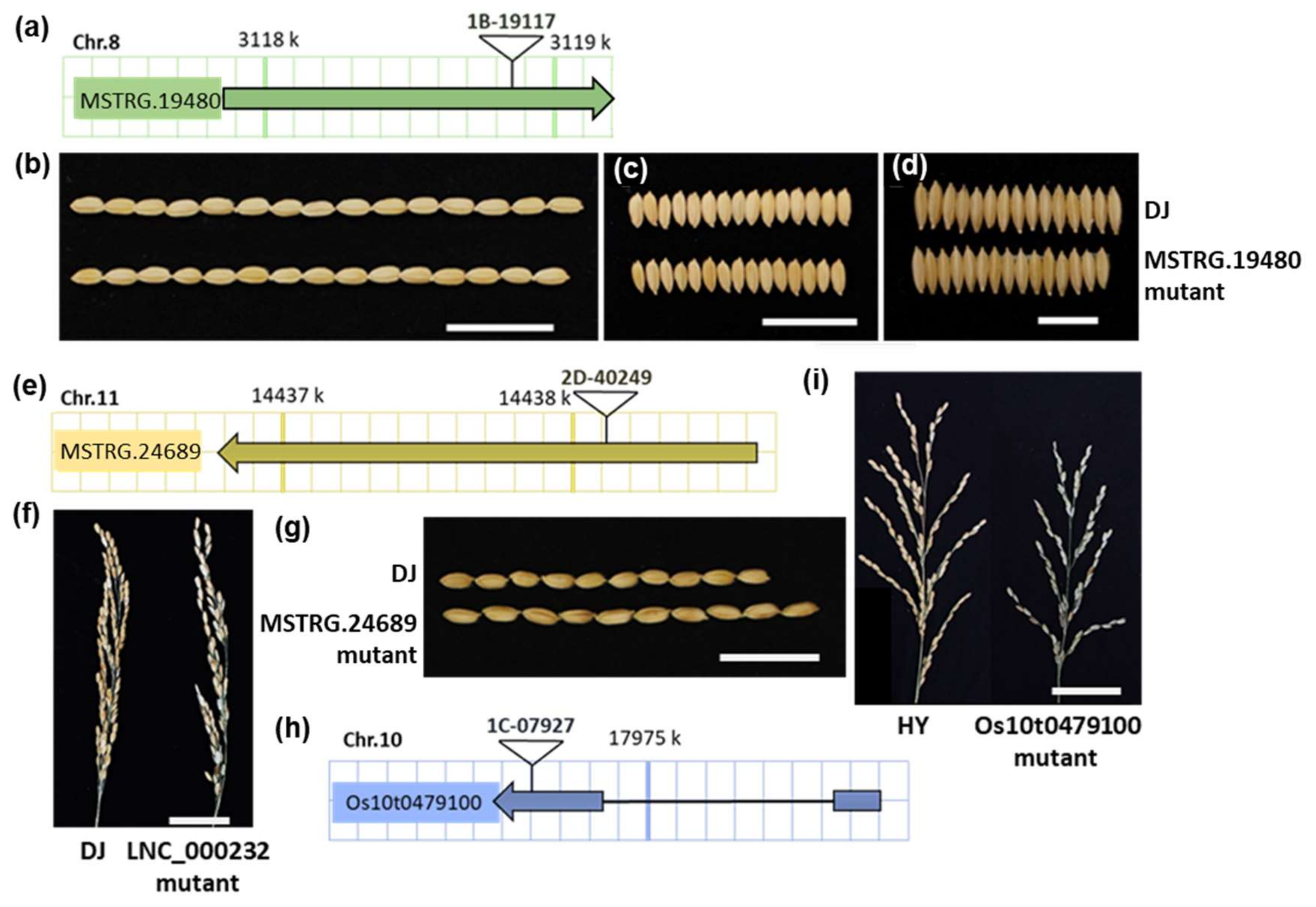
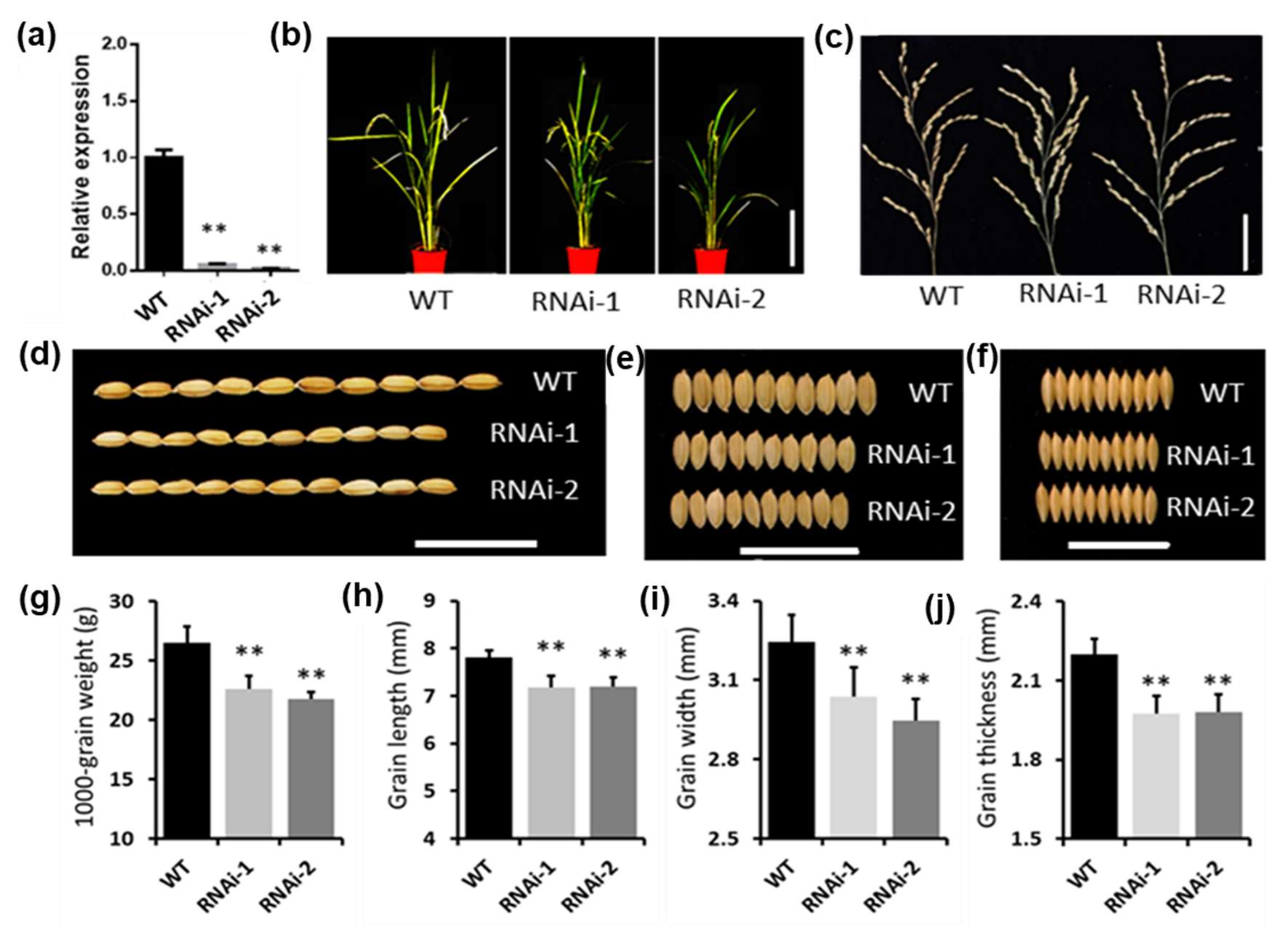
2.4. LncRNAs Are Potential Targets for the Selection of Yield-Related Traits during Domestication
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. RNA Extraction, Library Construction and Sequencing
4.3. LncRNA Identification
4.4. Expression Analysis
4.5. Conservation Analyses
4.6. Vector Construction and Rice Transformation
4.7. Plant Inoculations
4.8. Phenotypic Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Choi, J.Y.; Platts, A.E.; Fuller, D.Q.; Hsing, Y.L.; Wing, R.A.; Purugganan, M.D. The Rice Paradox: Multiple Origins but Single Domestication in Asian Rice. Mol. Biol. Evol. 2017, 34, 969–979. [Google Scholar] [CrossRef] [PubMed]
- Londo, J.P.; Chiang, Y.C.; Hung, K.H.; Chiang, T.Y.; Schaal, B.A. Phylogeography of Asian wild rice, Oryza rufipogon, reveals multiple independent domestications of cultivated rice, Oryza sativa. Proc. Natl. Acad. Sci. USA 2006, 103, 9578–9583. [Google Scholar] [CrossRef] [PubMed]
- Sweeney, M.; Mccouch, S. The complex history of the domestication of rice. Ann. Bot. 2007, 100, 951–957. [Google Scholar] [CrossRef] [PubMed]
- Usman, B.; Nawaz, G.; Zhao, N.; Liao, S.; Qin, B.; Liu, F.; Liu, Y.; Li, R. Programmed Editing of Rice (Oryza sativa L.) OsSPL16 Gene Using CRISPR/Cas9 Improves Grain Yield by Modulating the Expression of Pyruvate Enzymes and Cell Cycle Proteins. Int. J. Mol. Sci. 2020, 22, 249. [Google Scholar] [CrossRef] [PubMed]
- Meyer, R.S.; Purugganan, M.D. Evolution of crop species: Genetics of domestication and diversification. Nat. Rev. Genet. 2013, 14, 840–852. [Google Scholar] [CrossRef] [PubMed]
- Kovach, M.J.; Sweeney, M.T.; McCouch, S.R. New insights into the history of rice domestication. Trends Genet. 2007, 23, 578–587. [Google Scholar] [CrossRef]
- Liu, H.; Li, Q.; Xing, Y. Genes Contributing to Domestication of Rice Seed Traits and Its Global Expansion. Genes 2018, 9, 489. [Google Scholar] [CrossRef]
- Wang, M.; Yu, Y.; Haberer, G.; Marri, P.R.; Fan, C.; Goicoechea, J.L.; Zuccolo, A.; Song, X.; Kudrna, D.; Ammiraju, J.S.S.; et al. The genome sequence of African rice (Oryza glaberrima) and evidence for independent domestication. Nat. Genet. 2014, 46, 982–988. [Google Scholar] [CrossRef]
- Huang, X.; Kurata, N.; Wei, X.; Wang, Z.-X.; Wang, A.; Zhao, Q.; Zhao, Y.; Liu, K.; Lu, H.; Li, W.; et al. A map of rice genome variation reveals the origin of cultivated rice. Nature 2012, 490, 497–501. [Google Scholar] [CrossRef]
- Wang, W.; Mauleon, R.; Hu, Z.; Chebotarov, D.; Tai, S.; Wu, Z.; Li, M.; Zheng, T.; Fuentes, R.R.; Zhang, F.; et al. Genomic variation in 3010 diverse accessions of Asian cultivated rice. Nature 2018, 557, 43–49. [Google Scholar] [CrossRef]
- Zhao, Q.; Feng, Q.; Lu, H.; Li, Y.; Wang, A.; Tian, Q.; Zhan, Q.; Lu, Y.; Zhang, L.; Huang, T.; et al. Pan-genome analysis highlights the extent of genomic variation in cultivated and wild rice. Nat. Genet. 2018, 50, 278–284. [Google Scholar] [CrossRef]
- Shomura, A.; Izawa, T.; Ebana, K.; Ebitani, T.; Kanegae, H.; Konishi, S.; Yano, M. Deletion in a gene associated with grain size increased yields during rice domestication. Nat. Genet. 2008, 40, 1023–1028. [Google Scholar] [CrossRef]
- Tang, J.; Chu, C. MicroRNAs in crop improvement: Fine-tuners for complex traits. Nat. Plants 2017, 3, 17077. [Google Scholar] [CrossRef]
- Yu, Y.; Zhou, Y.; Zhang, Y.; Chen, Y. Grass phasiRNAs and male fertility. Sci. China Life Sci. 2018, 61, 148–154. [Google Scholar] [CrossRef]
- Ta, K.N.; Sabot, F.; Adam, H.; Vigouroux, Y.; De Mita, S.; Ghesquiere, A.; Do, N.V.; Gantet, P.; Jouannic, S. miR2118-triggered phased siRNAs are differentially expressed during the panicle development of wild and domesticated African rice species. Rice 2016, 9, 10. [Google Scholar] [CrossRef]
- Jiao, Y.; Wang, Y.; Xue, D.; Wang, J.; Yan, M.; Liu, G.; Dong, G.; Zeng, D.; Lu, Z.; Zhu, X.; et al. Regulation of OsSPL14 by OsmiR156 defines ideal plant architecture in rice. Nat. Genet. 2010, 42, 541–544. [Google Scholar] [CrossRef]
- Miura, K.; Ikeda, M.; Matsubara, A.; Song, X.-J.; Ito, M.; Asano, K.; Matsuoka, M.; Kitano, H.; Ashikari, M. OsSPL14 promotes panicle branching and higher grain productivity in rice. Nat. Genet. 2010, 42, 545–549. [Google Scholar] [CrossRef]
- Yu, Y.; Zhang, Y.; Chen, X.; Chen, Y. Plant Noncoding RNAs: Hidden Players in Development and Stress Responses. Annu. Rev. Cell Dev. Biol. 2019, 35, 407–431. [Google Scholar] [CrossRef]
- Yu, Y.; Zhou, Y.F.; Feng, Y.Z.; He, H.; Lian, J.P.; Yang, Y.W.; Lei, M.Q.; Zhang, Y.C.; Chen, Y.Q. Transcriptional landscape of pathogen-responsive lncRNAs in rice unveils the role of ALEX1 in jasmonate pathway and disease resistance. Plant. Biotechnol. J. 2020, 18, 679–690. [Google Scholar] [CrossRef]
- Waseem, M.; Liu, Y.; Xia, R. Long Non-Coding RNAs, the Dark Matter: An Emerging Regulatory Component in Plants. Int. J. Mol. Sci. 2020, 22, 86. [Google Scholar] [CrossRef]
- Zheng, X.M.; Chen, J.; Pang, H.B.; Liu, S.; Gao, Q.; Wang, J.R.; Qiao, W.H.; Wang, H.; Liu, J.; Olsen, K.M.; et al. Genome-wide analyses reveal the role of noncoding variation in complex traits during rice domestication. Sci. Adv. 2019, 5, eaax3619. [Google Scholar] [CrossRef]
- Deng, P.; Liu, S.; Nie, X.; Weining, S.; Wu, L. Conservation analysis of long non-coding RNAs in plants. Sci. China Life Sci. 2018, 61, 190–198. [Google Scholar] [CrossRef]
- Jacquemin, J.; Bhatia, D.; Singh, K.; Wing, R.A. The International Oryza Map Alignment Project: Development of a genus-wide comparative genomics platform to help solve the 9 billion-people question. Curr. Opin. Plant. Biol. 2013, 16, 147–156. [Google Scholar] [CrossRef]
- Kawahara, Y.; de la Bastide, M.; Hamilton, J.P.; Kanamori, H.; McCombie, W.R.; Ouyang, S.; Schwartz, D.C.; Tanaka, T.; Wu, J.; Zhou, S.; et al. Improvement of the Oryza sativa Nipponbare reference genome using next generation sequence and optical map data. Rice 2013, 6, 4. [Google Scholar] [CrossRef]
- Sakai, H.; Lee, S.S.; Tanaka, T.; Numa, H.; Kim, J.; Kawahara, Y.; Wakimoto, H.; Yang, C.C.; Iwamoto, M.; Abe, T.; et al. Rice Annotation Project Database (RAP-DB): An integrative and interactive database for rice genomics. Plant. Cell Physiol. 2013, 54, e6. [Google Scholar] [CrossRef]
- Yu, J.; Hu, S.; Wang, J.; Wong, G.K.; Li, S.; Liu, B.; Deng, Y.; Dai, L.; Zhou, Y.; Zhang, X.; et al. A draft sequence of the rice genome (Oryza sativa L. ssp. indica). Science 2002, 296, 79–92. [Google Scholar] [CrossRef]
- Zhao, W.; Wang, J.; He, X.; Huang, X.; Jiao, Y.; Dai, M.; Wei, S.; Fu, J.; Chen, Y.; Ren, X.; et al. BGI-RIS: An integrated information resource and comparative analysis workbench for rice genomics. Nucleic Acids Res. 2004, 32, D377–D382. [Google Scholar] [CrossRef]
- Nguyen, N.T.T.; Contreras-Moreira, B.; Castro-Mondragon, J.A.; Santana-Garcia, W.; Ossio, R.; Robles-Espinoza, C.D.; Bahin, M.; Collombet, S.; Vincens, P.; Thieffry, D.; et al. RSAT 2018: Regulatory sequence analysis tools 20th anniversary. Nucleic Acids Res. 2018, 46, W209–W214. [Google Scholar] [CrossRef]
- Nejat, N.; Mantri, N. Emerging roles of long non-coding RNAs in plant response to biotic and abiotic stresses. Crit. Rev. Biotechnol. 2018, 38, 93–105. [Google Scholar] [CrossRef]
- Zhang, J.; Li, C.; Wu, C.; Xiong, L.; Chen, G.; Zhang, Q.; Wang, S. RMD: A rice mutant database for functional analysis of the rice genome. Nucleic Acids Res. 2006, 34, D745–D748. [Google Scholar] [CrossRef]
- Jeong, D.H.; An, S.; Park, S.; Kang, H.G.; Park, G.G.; Kim, S.R.; Sim, J.; Kim, Y.O.; Kim, M.K.; Kim, S.R.; et al. Generation of a flanking sequence-tag database for activation-tagging lines in japonica rice. Plant J. 2006, 45, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Gauley, A.; Boden, S.A. Genetic pathways controlling inflorescence architecture and development in wheat and barley. J. Integr. Plant Biol. 2019, 61, 296–309. [Google Scholar] [CrossRef] [PubMed]
- Geshnizjani, N.; Sarikhani Khorami, S.; Willems, L.A.J.; Snoek, B.L.; Hilhorst, H.W.M.; Ligterink, W. The interaction between genotype and maternal nutritional environments affects tomato seed and seedling quality. J. Exp. Bot. 2019, 70, 2905–2918. [Google Scholar] [CrossRef] [PubMed]
- Golan, G.; Oksenberg, A.; Peleg, Z. Genetic evidence for differential selection of grain and embryo weight during wheat evolution under domestication. J. Exp. Bot. 2015, 66, 5703–5711. [Google Scholar] [CrossRef]
- Haas, M.; Schreiber, M.; Mascher, M. Domestication and crop evolution of wheat and barley: Genes, genomics, and future directions. J. Integr. Plant Biol. 2019, 61, 204–225. [Google Scholar] [CrossRef]
- Chen, Z.; Li, F.; Yang, S.; Dong, Y.; Yuan, Q.; Wang, F.; Li, W.; Jiang, Y.; Jia, S.; Pei, X. Identification and functional analysis of flowering related microRNAs in common wild rice (Oryza rufipogon Griff.). PLoS ONE 2013, 8, e82844. [Google Scholar] [CrossRef]
- Fan, Y.; Yang, J.; Mathioni, S.M.; Yu, J.; Shen, J.; Yang, X.; Wang, L.; Zhang, Q.; Cai, Z.; Xu, C.; et al. PMS1T, producing phased small-interfering RNAs, regulates photoperiod-sensitive male sterility in rice. Proc. Natl. Acad. Sci. USA 2016, 113, 15144–15149. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, Y.; Zhu, Q.H.; Fan, L. Identification of phasiRNAs in wild rice (Oryza rufipogon). Plant Signal. Behav. 2013, 8, e25079. [Google Scholar] [CrossRef][Green Version]
- Wang, Y.; Bai, X.; Yan, C.; Gui, Y.; Wei, X.; Zhu, Q.H.; Guo, L.; Fan, L. Genomic dissection of small RNAs in wild rice (Oryza rufipogon): Lessons for rice domestication. New Phytol. 2012, 196, 914–925. [Google Scholar] [CrossRef]
- Cho, J. Transposon-Derived Non-coding RNAs and Their Function in Plants. Front. Plant. Sci. 2018, 9, 600. [Google Scholar] [CrossRef]
- Wang, D.; Qu, Z.P.; Yang, L.; Zhang, Q.Z.; Liu, Z.H.; Do, T.; Adelson, D.L.; Wang, Z.Y.; Searle, I.; Zhu, J.K. Transposable elements (TEs) contribute to stress-related long intergenic noncoding RNAs in plants. Plant J. 2017, 90, 133–146. [Google Scholar] [CrossRef]
- Wang, X.; Ai, G.; Zhang, C.L.; Cui, L.; Wang, J.F.; Li, H.X.; Zhang, J.H.; Ye, Z.B.A. Expression and diversification analysis reveals transposable elements play important roles in the origin of Lycopersicon-specific lncRNAs in tomato. New Phytol. 2016, 209, 1442–1455. [Google Scholar] [CrossRef]
- Liu, W.; Chen, L.; Zhang, S.L.; Hu, F.Y.; Wang, Z.; Lyu, J.; Wang, B.; Xiang, H.; Zhao, R.P.; Tian, Z.X.; et al. Decrease of gene expression diversity during domestication of animals and plants. BMC Evol. Biol. 2019, 19. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index | O. rufipogon | O. sativa ssp. japonica | O. sativa ssp. indica | |||
|---|---|---|---|---|---|---|
| Panicle | Seedling | Panicle | Seedling | Panicle | Seedling | |
| Raw reads | 82,586,298 | 90,574,892 | 88,408,434 | 97,907,538 | 93,570,604 | 104,287,794 |
| Clean reads | 82,578,958 | 90,569,210 | 88,402,696 | 97,902,546 | 93,564,262 | 104,279,248 |
| Clean bases | 12.38 G | 13.58 G | 13.25 G | 14.68 G | 14.03 G | 15.64 G |
| GC content | 51.09% | 46.50% | 49.66% | 48.10% | 49.67% | 48.91% |
| Total mapped reads (%) | 62,020,407 (80.10%) | 75,227,980 (83.74%) | 80,629,086 (92.84%) | 91,211,211 (94.06%) | 81,866,743 (89.13%) | 97,171,107 (95.20%) |
| Unique mapped reads (%) | 61,024,157 (78.82%) | 73,364,876 (81.67%) | 67,553,409 (77.79%) | 60,706,447 (62.60%) | 74,933,904 (81.59%) | 78,992,468 (77.39%) |
| Multiple mapped reads (%) | 996,250 (1.29%) | 1,863,104 (2.07%) | 13,075,677 (15.06%) | 30,504,764 (31.46%) | 6,932,839 (7.55%) | 18,178,639 (17.81%) |
| Reads map to ‘exon’ (%) | 32,898,026 (53.04%) | 30,524,827 (40.58%) | 34,735,578 (43.08%) | 17,715,152 (19.42%) | 31,530,937 (38.51%) | 30,706,672 (31.60%) |
| Reads map to ‘intron’ (%) | 8,676,838 (13.99%) | 6149,971 (8.18%) | 5,247,590 (6.51%) | 2,080,358 (2.28%) | 6,717,453 (8.21%) | 2,175,025 (2.24%) |
| Reads map to ‘intergenic’ (%) | 20,445,543 (32.97%) | 38,553,182 (51.25%) | 40,645,918 (50.41%) | 71,415,701 (78.30%) | 43,618,353 (53.28%) | 64,289,410 (66.16%) |
| lncRNA ID | WT-Dongjin | MSTRG.19480 | MSTRG.24689 | WT-Hwayoung | Os10t0479100 |
|---|---|---|---|---|---|
| Mutant ID (wild-type) | Dongjin (DJ) | 1B-19117 (DJ) | 2D-40249 (DJ) | Hwayoung (HY) | 1C-07927 (HY) |
| Plant height (cm) | 64.38 ± 3.56 | 51.38 ± 3.87 (**↓) | 59.62 ± 4.45 (**↓) | 64.50 ± 4.54 | 56.92 ± 3.04 (**↓) |
| Panicle length (cm) | 16.80 ± 0.56 | 16.62 ± 2.04 (n.s.) | 15.76 ± 1.14 (**↓) | 16.35 ± 1.44 | 15.73 ± 1.32 (n.s.) |
| Number of primary branches | 6.57 ± 0.82 | 6.06 ± 1.57 (n.s.) | 6.67 ± 0.85 | 5.82 ± 0.57 | 6.57 ± 0.82 (n.s.) |
| Seed-setting rate (%) | 90.52 ± 5.05 | 90.61 ± 5.01 (n.s.) | 67.53 ± 5.08 (**↓) | 95.00 ± 4.57 | 11.01 ± 3.93 (**↓) |
| 1000-grain weight (g) | 25.56 ± 0.56 | 20.66 ± 2.93 (**↓) | 26.40 ± 1.34 (*↑) | 24.75 ± 1.21 | 24.80 ± 0.64 (n.s.) |
| Grain length (mm) | 7.64 ± 0.33 | 6.93 ± 0.27 (**↓) | 8.28 ± 0.26 (**↑) | 7.94 ± 0.29 | 8.02 ± 0.25 (n.s.) |
| Grain width (mm) | 3.49 ± 0.14 | 2.72 ± 0.17 (**↓) | 3.43 ± 0.14 (n.s.) | 3.56 ± 0.12 | 3.61 ± 0.16 (n.s.) |
| Grain thickness (mm) | 2.33 ± 0.09 | 2.08 ± 0.19 (**↓) | 2.31 ± 0.06 (n.s.) | 2.18 ± 0.08 | 2.14 ± 0.08 (n.s.) |
| Lesion length of Xoo infection (cm) | 6.56 ± 1.99 | 4.73 ± 1.68 (**↓) | 5.71 ± 2.97 (n.s.) | 5.31 ± 2.29 | 9.10 ± 2.99 (**↑) |
| Lesion length of Xoc infection (cm) | 2.11 ± 0.76 | 2.30 ± 0.22 (n.s.) | 0.47 ± 0.18 (**↓) | 1.15 ± 0.76 | 2.19 ± 0.83 (**↑) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, H.; Zhou, Y.-F.; Yang, Y.-W.; Zhang, Z.; Lei, M.-Q.; Feng, Y.-Z.; Zhang, Y.-C.; Chen, Y.-Q.; Lian, J.-P.; Yu, Y. Genome-Wide Analysis Identified a Set of Conserved lncRNAs Associated with Domestication-Related Traits in Rice. Int. J. Mol. Sci. 2021, 22, 4742. https://doi.org/10.3390/ijms22094742
He H, Zhou Y-F, Yang Y-W, Zhang Z, Lei M-Q, Feng Y-Z, Zhang Y-C, Chen Y-Q, Lian J-P, Yu Y. Genome-Wide Analysis Identified a Set of Conserved lncRNAs Associated with Domestication-Related Traits in Rice. International Journal of Molecular Sciences. 2021; 22(9):4742. https://doi.org/10.3390/ijms22094742
Chicago/Turabian StyleHe, Huang, Yan-Fei Zhou, Yu-Wei Yang, Zhi Zhang, Meng-Qi Lei, Yan-Zhao Feng, Yu-Chan Zhang, Yue-Qin Chen, Jian-Ping Lian, and Yang Yu. 2021. "Genome-Wide Analysis Identified a Set of Conserved lncRNAs Associated with Domestication-Related Traits in Rice" International Journal of Molecular Sciences 22, no. 9: 4742. https://doi.org/10.3390/ijms22094742
APA StyleHe, H., Zhou, Y.-F., Yang, Y.-W., Zhang, Z., Lei, M.-Q., Feng, Y.-Z., Zhang, Y.-C., Chen, Y.-Q., Lian, J.-P., & Yu, Y. (2021). Genome-Wide Analysis Identified a Set of Conserved lncRNAs Associated with Domestication-Related Traits in Rice. International Journal of Molecular Sciences, 22(9), 4742. https://doi.org/10.3390/ijms22094742