
A Comprehensive Analysis into the Therapeutic Application of Natural Products as SIRT6 Modulators in Alzheimer’s Disease, Aging, Cancer, Inflammation, and Diabetes

 ,
,  ,
,
Abstract
1. Introduction
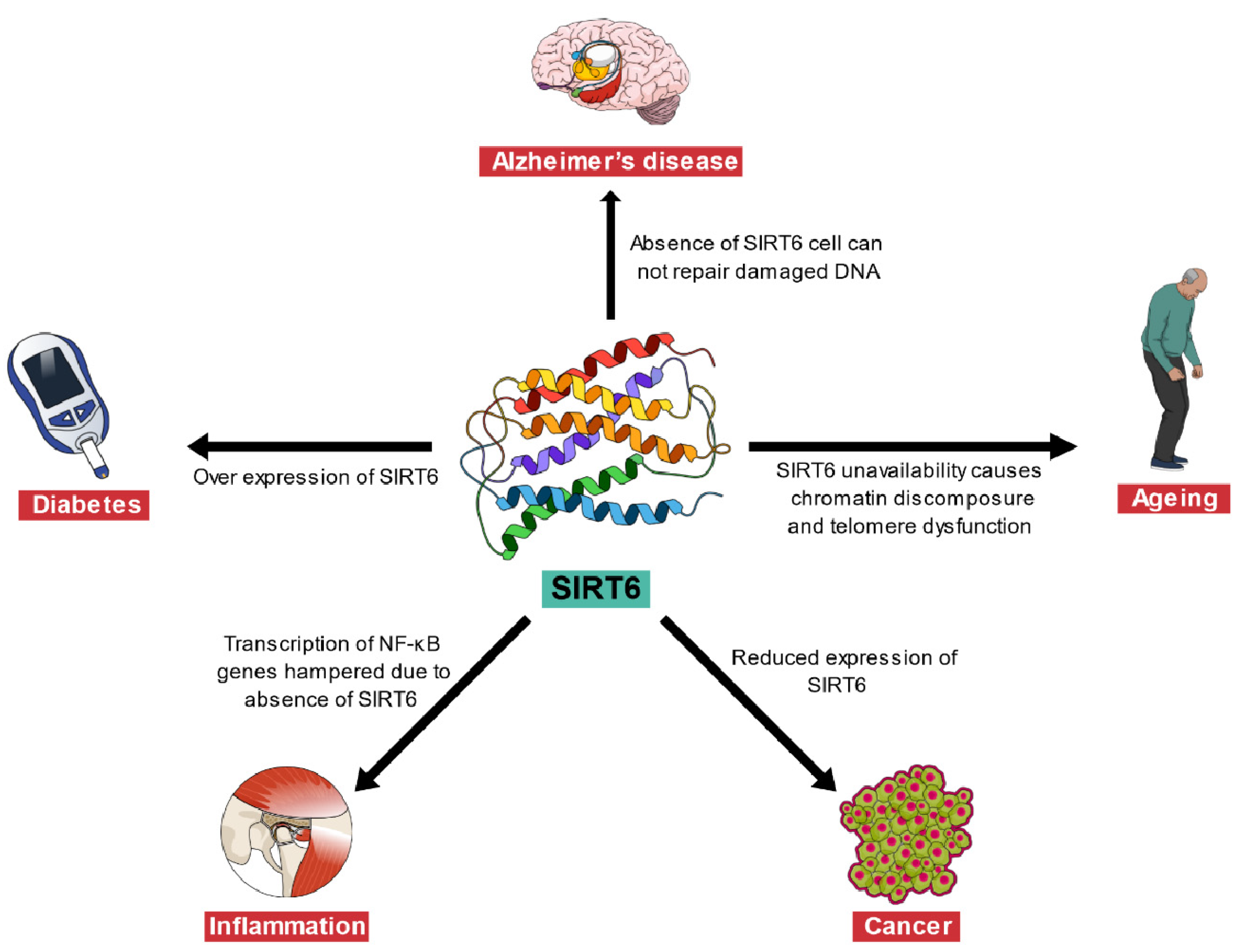
2. Sirtuin 6 (SIRT6) and Its Association with Different Diseases
2.1. Association of SIRT6 with Alzheimer’s Disease and Aging
2.2. Association of SIRT6 with Cancer
2.3. Association of SIRT6 with Inflammation
2.4. Association of SIRT6 with Diabetes
3. Natural Products as Modulators of SIRT6: Promising Therapeutic Targets
3.1. Natural Products Acting as SIRT6 Activators
3.1.1. Polyphenols and Flavonoids
Quercetin
Isoquercetin
Kaempferol
Luteolin
Cyanidin
Fisetin
Delphinidin and Its Derivative
Icariin
3.1.2. Polysaccharides
Fucoidan
Arabinoxylans
3.1.3. Fatty Acids
Oleic Acid and Linoleic Acid
Fatty Acid Derivatives (N-acylethanolamines)
3.2. Natural Products Acting as SIRT6 Inhibitors
3.2.1. Polyphenols and Flavonoids
Quercetin, Its Derivatives and Luteolin
Vitexin
Catechin and Its Derivatives
Scutellarin
3.2.2. Vitamins
Nicotinamide
3.2.3. Peptides
3.2.4. Fatty Acid
Myristic Acid
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ji, H.-F.; Li, X.-J.; Zhang, H.-Y. Natural products and drug discovery. Can thousands of years of ancient medical knowledge lead us to new and powerful drug combinations in the fight against cancer and dementia? EMBO Rep. 2009, 10, 194–200. [Google Scholar] [CrossRef]
- Mathur, S.; Hoskins, C. Drug development: Lessons from nature. Biomed. Rep. 2017, 6, 612–614. [Google Scholar] [CrossRef] [PubMed]
- Dobson, P.D.; Patel, Y.; Kell, D.B. ‘Metabolite-likeness’ as a criterion in the design and selection of pharmaceutical drug libraries. Drug Discov. Today 2009, 14, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Carlson, E.E. Natural products as chemical probes. ACS Chem. Biol. 2010, 5, 639–653. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, B.B.; Shishodia, S.; Sandur, S.K.; Pandey, M.K.; Sethi, G. Inflammation and cancer: How hot is the link? Biochem. Pharmacol. 2006, 72, 1605–1621. [Google Scholar] [CrossRef]
- Ambrosino, P.; Soldovieri, M.; de Maria, M.; Russo, C.; Taglialatela, M. Functional and biochemical interaction between PPARα receptors and TRPV1 channels: Potential role in PPARα agonists-mediated analgesia. Pharmacol. Res. 2014, 87, 113–122. [Google Scholar] [CrossRef]
- Aggarwal, B. Nuclear factor-κB: The enemy within. Cancer Cell 2004, 6, 203–208. [Google Scholar] [CrossRef]
- Ambrosino, P.; Soldovieri, M.; Russo, C.; Taglialatela, M. Activation and desensitization of TRPV1 channels in sensory neurons by the PPARα agonist palmitoylethanolamide. Br. J. Pharmacol. 2013, 168, 1430–1444. [Google Scholar] [CrossRef]
- Michishita, E.; McCord, R.A.; Berber, E.; Kioi, M.; Padilla-Nash, H.; Damian, M.; Cheung, P.; Kusumoto, R.; Kawahara, T.L.A.; Barrett, J.C.; et al. SIRT6 is a histone H3 lysine 9 deacetylase that modulates telomeric chromatin. Nature 2008, 452, 492–496. [Google Scholar] [CrossRef]
- Michishita, E.; McCord, R.A.; Boxer, L.D.; Barber, M.F.; Hong, T.; Gozani, O.; Chua, K.F. Cell cycle-dependent deacetylation of telomeric histone H3 lysine K56 by human SIRT6. Cell Cycle 2009, 8, 2664–2666. [Google Scholar] [CrossRef]
- Tasselli, L.; Zheng, W.; Chua, K.F. SIRT6: Novel Mechanisms and Links to Aging and Disease. Trends Endocrinol. Metab. 2017, 28, 168–185. [Google Scholar] [CrossRef]
- Nirzhor, S.; Khan, R.; Neelotpol, S. The Biology of Glial Cells and Their Complex Roles in Alzheimer’s Disease: New Opportunities in Therapy. Biomolecules 2018, 8, 93. [Google Scholar] [CrossRef] [PubMed]
- Heger, V.; Tyni, J.; Hunyadi, A.; Horáková, L.; Lahtela-Kakkonen, M.; Rahnasto-Rilla, M. Quercetin based derivatives as sirtuin inhibitors. Biomed. Pharmacother. 2019, 111, 1326–1333. [Google Scholar] [CrossRef]
- Khan, R.I.; Nirzhor, S.S.R.; Akter, R. A Review of the Recent Advances Made with SIRT6 and its Implications on Aging Related Processes, Major Human Diseases, and Possible Therapeutic Targets. Biomolecules 2018, 8, 44. [Google Scholar] [CrossRef] [PubMed]
- Mao, Z.; Hine, C.; Tian, X.; van Meter, M.; Au, M.; Vaidya, A.; Seluanov, A.; Gorbunova, V. SIRT6 promotes DNA repair under stress by activating PARP1. Science 2011, 332, 1443–1446. [Google Scholar] [CrossRef] [PubMed]
- Uddin, M.J.; Khan, R.I.; Nirzhor, S.S.R. A review of the promising new advances in the combat against cholesterol trafficking defect and consequent occurrence of the rare Niemann-pick type C disease. Int. J. Biomed. Adv. Res. 2018, 9, 275–285. [Google Scholar]
- Whyte, L.S.; Lau, A.A.; Hemsley, K.M.; Hopwood, J.J.; Sargeant, T.J. Endo-lysosomal and autophagic dysfunction: A driving factor in Alzheimer’s disease? J. Neurochem. 2017, 140, 703–717. [Google Scholar] [CrossRef] [PubMed]
- Islam Khan, R.; Nirzhor, S.S.R.; Rashid, B. A Closer Look into the Role of Protein Tau in the Identification of Promising Therapeutic Targets for Alzheimer’s Disease. Brain Sci. 2018, 8, 162. [Google Scholar] [CrossRef]
- Kaluski, S.; Portillo, M.; Besnard, A.; Stein, D.; Einav, M.; Zhong, L.; Ueberham, U.; Arendt, T.; Mostoslavsky, R.; Sahay, A.; et al. Neuroprotective Functions for the Histone Deacetylase SIRT6. Cell Rep. 2017, 18, 3052–3062. [Google Scholar] [CrossRef]
- Van Meter, M.; Kashyap, M.; Rezazadeh, S.; Geneva, A.; Morello, T.; Seluanov, A.; Gorbunova, V. SIRT6 represses LINE1 retrotransposons by ribosylating KAP1 but this repression fails with stress and age. Nat. Commun. 2014, 5, 5011. [Google Scholar] [CrossRef]
- Tennen, R.; Bua, D.; Wright, W.; Chua, K. SIRT6 is required for maintenance of telomere position effect in human cells. Nat. Commun. 2011, 2, 433. [Google Scholar] [CrossRef] [PubMed]
- Endisha, H.; Merrill-Schools, J.; Zhao, M.; Bristol, M.; Wang, X.; Kubben, N.; Elmore, L.W. Restoring SIRT6 Expression in Hutchinson-Gilford Progeria Syndrome Cells Impedes Premature Senescence and Formation of Dysmorphic Nuclei. Pathobiology 2015, 82, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Kugel, S.; Mostoslavsky, R. Chromatin and beyond: The multitasking roles for SIRT6. Trends Biochem. Sci. 2014, 39, 72–81. [Google Scholar] [CrossRef]
- Shin, J.; He, M.; Liu, Y.; Paredes, S.; Villanova, L.; Brown, K.; Qiu, X.; Nabavi, N.; Mohrin, M.; Wojnoonski, K.; et al. SIRT7 Represses Myc Activity to Suppress ER Stress and Prevent Fatty Liver Disease. Cell Rep. 2013, 5, 654–665. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Tu, B.; Wang, H.; Cao, Z.; Tang, M.; Zhang, C.; Gu, B.; Li, Z.; Wang, L.; Yang, Y.; et al. Tumor suppressor p53 cooperates with SIRT6 to regulate gluconeogenesis by promoting FoxO1 nuclear exclusion. Proc. Natl. Acad. Sci. USA 2014, 111, 10684–10689. [Google Scholar] [CrossRef]
- You, W.; Rotili, D.; Li, T.-M.; Kambach, C.; Meleshin, M.; Schutkowski, M.; Chua, K.F.; Mai, A.; Steegborn, C. Structural Basis of Sirtuin 6 Activation by Synthetic Small Molecules. Angew. Chem. Int. Ed. 2017, 56, 1007–1011. [Google Scholar] [CrossRef]
- Jackson, M.D.; Denu, J.M. Structural identification of 2′- and 3′-O-acetyl-ADP-ribose as novel metabolites derived from the Sir2 family β-NAD+ -dependent histone/protein deacetylases. J. Biol. Chem. 2002, 277, 8535–18544. [Google Scholar] [CrossRef]
- Denu, J.M. The Sir2 family of protein deacetylases. Curr. Opin. Chem. Biol. 2005, 9, 431–440. [Google Scholar] [CrossRef]
- Beauharnois, J.M.; Bolívar, B.E.; Welch, J.T. Sirtuin 6: A review of biological effects and potential therapeutic properties. Mol. BioSyst. 2013, 9, 1789–1806. [Google Scholar] [CrossRef]
- Michishita, E.; Park, J.; Burneskis, J.; Barrett, C.; Horikawa, I. Evolutionarily conserved and nonconserved cellular localizations and functions of human SIRT proteins. Mol. Biol. Cell 2005, 16, 4623–4635. [Google Scholar] [CrossRef]
- Pereira, C.V.; Lebiedzinska, M.; Wieckowski, M.R.; Oliveira, P.J. Regulation and protection of mitochondrial physiology by sirtuins. Mitochondrion 2012, 12, 66–76. [Google Scholar] [CrossRef] [PubMed]
- Lombard, D.; Schwer, B.; Alt, F.; Mostoslavsky, R. SIRT6 in DNA repair, metabolism and ageing. J. Intern. Med. 2008, 263, 128–141. [Google Scholar] [CrossRef] [PubMed]
- Mostoslavsky, R.; Chua, K.F.; Lombard, D.B.; Pang, W.W.; Fischer, M.R.; Gellon, L.; Liu, P.; Mostoslavsky, G.; Franco, S.; Murphy, M.M.; et al. Genomic Instability and Aging-like Phenotype in the Absence of Mammalian SIRT6. Cell 2006, 124, 315–329. [Google Scholar] [CrossRef] [PubMed]
- Peshti, V.; Obolensky, A.; Nahum, L.; Kanfi, Y.; Rathaus, M.; Avraham, M.; Tinman, S.; Alt, F.W.; Banin, E.; Cohen, H.Y. Characterization of physiological defects in adult SIRT6-/- mice. PLoS ONE 2017, 12, e0176371. [Google Scholar] [CrossRef]
- Finkel, T.; Deng, C.-X.; Mostoslavsky, R. Recent progress in the biology and physiology of Sirtuins. Nature 2009, 460, 587–591. [Google Scholar] [CrossRef]
- Dimauro, T.; David, G. Chromatin modifications: The driving force of senescence and aging? Aging 2009, 1, 182–190. [Google Scholar] [CrossRef][Green Version]
- Desantis, V.; Lamanuzzi, A.; Vacca, A. The role of SIRT6 in tumors. Haematologica 2018, 103, 1–4. [Google Scholar] [CrossRef]
- Losson, H.; Schnekenburger, M.; Dicato, M.; Diederich, M. Natural Compound Histone Deacetylase Inhibitors (HDACi): Synergy with Inflammatory Signaling Pathway Modulators and Clinical Applications in Cancer. Molecules 2016, 21, 1608. [Google Scholar] [CrossRef]
- Carafa, V.; Altucci, L.; Nebbioso, A. Dual Tumor Suppressor and Tumor Promoter Action of Sirtuins in Determining Malignant Phenotype. Front. Pharmacol. 2019, 10, 38. [Google Scholar] [CrossRef]
- Kugel, S.; Sebastián, C.; Fitamant, J.; Ross, K.; Saha, S.K.; Jain, E.; Gladden-Young, A.; Arora, K.S.; Kato, Y.; Rivera, M.N. SIRT6 Suppresses Pancreatic Cancer through Control of Lin28b. Cell 2016, 165, 1401–1415. [Google Scholar] [CrossRef]
- Kuang, J.; Chen, L.; Tang, Q.; Zhang, J.; Li, Y.; He, J. The Role of Sirt6 in Obesity and Diabetes. Front. Physiol. 2018, 9, 135. [Google Scholar] [CrossRef]
- Xiao, C.; Wang, R.-H.; Lahusen, T.J.; Park, O.; Bertola, A.; Maruyama, T.; Reynolds, D.; Chen, Q.; Xu, X.; Young, H.A.; et al. Progression of Chronic Liver Inflammation and Fibrosis Driven by Activation of c-JUN Signaling in Sirt6 Mutant Mice. J. Biol. Chem. 2012, 287, 41903–41913. [Google Scholar] [CrossRef]
- Sánchez-Fidalgo, S.; Villegas, I.; Sánchez-Hidalgo, M.; Lastra, C. Sirtuin Modulators: Mechanisms and Potential Clinical Implications. Curr. Med. Chem. 2012, 19, 2414–2441. [Google Scholar] [CrossRef]
- Sociali, G.; Magnone, M.; Ravera, S.; Damonte, P.; Vigliarolo, T.; Holtey, M.; Vellone, V.; Millo, E.; Caffa, I.; Cea, M.; et al. Pharmacological Sirt6 inhibition improves glucose tolerance in a type 2 diabetes mouse model. FASEB J. 2017, 31, 3138–3149. [Google Scholar] [CrossRef] [PubMed]
- Zhong, L.; Mostoslavsky, R. SIRT6: A master epigenetic gatekeeper of glucose metabolism. Transcription 2010, 1, 17–21. [Google Scholar] [CrossRef] [PubMed]
- Zhong, L.; D’Urso, A.; Toiber, D.; Sebastian, C.; Henry, R.; Vadysirisack, D.; Guimaraes, A.; Marinelli, B.; Wikstrom, J.; Nir, T.; et al. The Histone Deacetylase SIRT6 Regulates Glucose Homeostasis via Hif1α. Cell 2010, 140, 280–293. [Google Scholar] [CrossRef] [PubMed]
- Dominy, J.; Lee, Y.; Jedrychowski, M.; Chim, H.; Jurczak, J.; Camporez, J.P.; Ruan, H.-B.; Feldman, J.; Pierce, K.; Mostoslavsky, R.; et al. The Deacetylase Sirt6 Activates the Acetyltransferase GCN5 and Suppresses Hepatic Gluconeogenesis. Mol. Cell 2012, 48, 900–913. [Google Scholar] [CrossRef]
- Song, M.-Y.; Wang, J.; Ka, S.-O.; Bae, E.J.; Park, B.-H. Insulin secretion impairment in Sirt6 knockout pancreatic β cells is mediated by suppression of the FoxO1-Pdx1-Glut2 pathway. Sci. Rep. 2016, 6, 30321. [Google Scholar] [CrossRef] [PubMed]
- Shishir, T.A.; Khan, R.I.; Nirzhor, S.S.R. The critical role of tumor microenvironment in cancer evolution and metastasis. Int. J. Biomed. Res. 2018, 9, 244–258. [Google Scholar]
- De Céu, T.M.; Sanchez-Lopez, E.; Espina, M.; Garcia, M.L.; Durazzo, A.; Lucarini, M.; Novellino, E.; Souto, S.B.; Santini, A.; Souto, E.B. Sirtuins and SIRT6 in Carcinogenesis and in Diet. Int. J. Mol. Sci. 2019, 20, 4945. [Google Scholar] [CrossRef]
- Bae, E.J. Sirtuin 6, a possible therapeutic target for type 2 diabetes. Arch. Pharm. Res. 2017, 40, 1380–1389. [Google Scholar] [CrossRef] [PubMed]
- Rahnasto-Rilla, M.; Tyni, J.; Huovinen, M.; Jarho, E.; Kulikowicz, T.; Ravichandran, S.A.; Bohr, V.; Ferrucci, L.; Lahtela-Kakkonen, M.; Moaddel, R. Natural polyphenols as sirtuin 6 modulators. Sci. Rep. 2018, 8, 4163. [Google Scholar] [CrossRef]
- Karaman Mayack, B.S.W.; Ntie-Kang, F. Natural Products as Modulators of Sirtuins. Molecules 2020, 25, 3287. [Google Scholar] [CrossRef] [PubMed]
- Miguel, M.G.; Nunes, S.; Dandlen, S.A.; Cavaco, A.M.; Antunes, M.D. Phenols and antioxidant activity of hydro-alcoholic extracts of propolis from Algarve, South of Portugal. Food Chem. Toxicol. 2010, 48, 3418–3423. [Google Scholar] [CrossRef] [PubMed]
- Zainab, S.A.; Nowrin, T.; Hossain, M.H.; Nasrin, T.; Akter, R. Profiling of Crotalaria verrucosa Leaf Extract and Evaluation of its Antioxidant and Cytotoxic Potency. Res. J. Chem. 2018, 12, 60–70. [Google Scholar]
- Mlcek, J.; Jurikova, T.; Skrovankova, S.; Sochor, J. Quercetin and Its Anti-Allergic Immune Response. Molecules 2016, 21, 623. [Google Scholar] [CrossRef] [PubMed]
- You, W.; Zheng, W.; Weiss, S.; Chua, K.F.; Steegborn, C. Structural basis for the activation and inhibition of Sirtuin 6 by quercetin and its derivatives. Sci. Rep. 2019, 9, 19176. [Google Scholar] [CrossRef]
- Rahnasto-Rilla, M.K.; McLoughlin, P.; Kulikowicz, T.; Doyle, M.; Bohr, V.A.; Lahtela-Kakkonen, M.; Ferrucci, L.; Hayes, M.; Moaddel, R. The Identification of a SIRT6 Activator from Brown Algae Fucus distichus. Mar. Drugs 2017, 15, 190. [Google Scholar] [CrossRef]
- Hałasa, M.; Wawruszak, A.; Przybyszewska, A.; Jaruga, A.; Guz, M.; Kałafut, J.; Stepulak, A.; Cybulski, M. H3K18Ac as a Marker of Cancer Progression and Potential Target of Anti-Cancer Therapy. Cells 2019, 8, 485. [Google Scholar] [CrossRef]
- Rahnasto-Rilla, M.; Kokkola, T.; Jarho, E.; Lahtela-Kakkonen, M.; Moaddel, R. N-Acylethanolamines Bind to SIRT6. ChemBioChem 2016, 17, 77–81. [Google Scholar] [CrossRef]
- Ravichandran, S.; Singh, N.; Donnelly, D.; Migliore, M.; Johnson, P.; Fishwick, C.; Luke, B.T.; Martin, B.; Maudsley, S.; Fugmann, S.D.; et al. Pharmacophore model of the quercetin binding site of the SIRT6 protein. J. Mol. Graph. Model 2014, 49, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Jurikova, T.; Sochor, J.; Rop, O.; Mlcek, J.; Balla, S.; Szekeres, L.; Adam, V.; Kizek, R. Polyphenolic Profile and Biological Activity of Chinese Hawthorn (Crataegus pinnatifida BUNGE) Fruits. Molecules 2012, 17, 14490–14509. [Google Scholar] [CrossRef]
- Valentová, K.; Vrba, J.; Bancirova, M.; Ulrichova, J.; Křen, V. Isoquercitrin: Pharmacology, toxicology, and metabolism. Food Chem. Toxicol. 2014, 68, 267–282. [Google Scholar] [CrossRef] [PubMed]
- Rubin, T.M.; Jae, K.S. A Review on Structure, Modifications and Structure-Activity Relation of Quercetin and Its Derivatives. J. Microbiol. Biotechnol. 2020, 30, 11–20. [Google Scholar]
- Zhang, R.; Yao, Y.; Wang, Y.; Ren, G. Antidiabetic activity of isoquercetin in diabetic KK -Ay mice. Nutr. Metab. 2011, 8, 85. [Google Scholar] [CrossRef] [PubMed]
- Maiyo, F.; Moodley, R.; Singh, M. Cytotoxicity, Antioxidant and Apoptosis Studies of Quercetin-3-O Glucoside and 4-(β-D-Glucopyranosyl-1→4-α-L-Rhamnopyranosyloxy)-Benzyl Isothiocyanate from Moringa oleifera. Anticancer Agents Med. Chem. 2016, 16, 648–656. [Google Scholar] [CrossRef]
- Jayachandran, M.; Wu, Z.; Ganesan, K.; Khalid, S.; Chung, S.M.; Xu, B. Isoquercetin upregulates antioxidant genes, suppresses inflammatory cytokines and regulates AMPK pathway in streptozotocin-induced diabetic rats. Chem-Biol. Interact. 2019, 303, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Dabeek, W.M.; Marra, M.V. Dietary Quercetin and Kaempferol: Bioavailability and Potential Cardiovascular-Related Bioactivity in Humans. Nutrients 2019, 11, 2288. [Google Scholar] [CrossRef]
- Calderon-Montano, M.J.; Burgos-Moron, E.; Perez-Guerrero, C.; Lopez-Lazaro, M. A Review on the Dietary Flavonoid Kaempferol. Mini-Rev. Med. Chem. 2011, 11, 298–344. [Google Scholar] [CrossRef]
- Kim, J.H.; Cho, E.J.; Kim, H.Y. Neuroprotective Effects of Kaempferol, Quercetin, and Its Glycosides by Regulation of Apoptosis. J. Korea Acad. Industr. Coop Soc. 2019, 20, 286–293. [Google Scholar]
- Lopez-Lazaro, M. Distribution and Biological Activities of the Flavonoid Luteolin. Mini-Rev. Med. Chem. 2009, 9, 31–59. [Google Scholar] [CrossRef] [PubMed]
- Shimoi, K.; Okada, H.; Furugori, M.; Goda, T.; Takase, S.; Suzuki, M.; Hara, Y.; Yamamoto, H.; Kinae, N. Intestinal absorption of luteolin and luteolin 7-O-β-glucoside in rats and humans. FEBS Lett. 1998, 438, 220–224. [Google Scholar] [CrossRef]
- Kim, A.; Lee, W.; Yun, J.-M. Luteolin and fisetin suppress oxidative stress by modulating sirtuins and forkhead box O3a expression under in vitro diabetic conditions. Nutr. Res. Pract. 2017, 11, 430–434. [Google Scholar] [CrossRef]
- Razavi, S.M.; Zahri, S.; Zarrini, G.; Nazemiyeh, H.; Mohammadi, S. Biological activity of quercetin-3-O-glucoside, a known plant flavonoid. Russ. J. Bioorganic Chem. 2009, 35, 376–378. [Google Scholar] [CrossRef]
- Khan, F.; Niaz, K.; Maqbool, F.; Ismail Hassan, F.; Abdollahi, M.; Nagulapalli Venkata, K.C.; Nabavi, S.M.; Bishayee, A. Molecular Targets Underlying the Anticancer Effects of Quercetin: An Update. Nutrients 2016, 8, 529. [Google Scholar] [CrossRef] [PubMed]
- Galvano, F.; Fauci, L.; Lazzarino, G.; Fogliano, V.; Ritieni, A.; Ciappellano, S.; Battistini, N.C.; Tavazzi, B.; Galvano, G. Cyanidins: Metabolism and biological properties. J. Nutr. Biochem. 2004, 15, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Corona, G.; Tang, F.; Vauzour, D.; Rodriguez-Mateos, A.; Spencer, J.P.E. Assessment of the anthocyanidin content of common fruits and development of a test diet rich in a range of anthocyanins. J. Berry Res. 2011, 1, 209–216. [Google Scholar] [CrossRef]
- Kwak, S.; Ku, S.-K.; Bae, J.-S. Fisetin inhibits high-glucose-induced vascular inflammation in vitro and in vivo. Inflamm. Res. 2014, 63, 779–787. [Google Scholar] [CrossRef]
- Khan, N.; Syed, D.N.; Ahmad, N.; Mukhtar, H. Fisetin: A dietary antioxidant for health promotion. Antioxid. Redox Signal 2013, 19, 151–162. [Google Scholar] [CrossRef]
- Cerit, H.; Engin, K.; Ipek, B.; Ayse, Z.A. Stop the Time for Your Skin: A Search for Ageing Process. Glob. J. Med. Res. 2018, 18, 1–7. [Google Scholar]
- Murakami, A.; Ohnishi, K. Target molecules of food phytochemicals: Food science bound for the next dimension. Food Funct. 2012, 3, 462–476. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zhu, Y.; Zhang, W.; Peng, X.; Zhou, J.; Li, F.; Han, B.; Liu, X.; Ou, Y.; Yu, X. Delphinidin induced protective autophagy via mTOR pathway suppression and AMPK pathway activation in HER-2 positive breast cancer cells. BMC Cancer 2018, 18, 342. [Google Scholar] [CrossRef] [PubMed]
- Jessica, T.; Emin, O.; Leslie, W.; George, A. Delphinidin and cyanidin exhibit antiproliferative and apoptotic effects in MCF7 human breast cancer cells. Integr. Cancer Sci. Ther. 2015, 2, 82–86. [Google Scholar]
- Patel, K.; Jain, A.; Patel, D.K. Medicinal significance, pharmacological activities, and analytical aspects of anthocyanidins ‘delphinidin’: A concise report. J. Acute Dis. 2013, 2, 169–178. [Google Scholar] [CrossRef]
- Xu, C.; Huang, X.; Tong, Y.; Feng, X.; Wang, Y.; Wang, C.; Jiang, Y. Icariin modulates the sirtuin/NF-κB pathway and exerts anti-aging effects in human lung fibroblasts. Mol. Med. Rep. 2020, 22, 3833–3839. [Google Scholar]
- Zhang, Z.; Yao, C.; Xu, S.; Yuan, G.Z.; Liu, Z.; Zhang, Z.; Shi, L. Determination of Six Main Flavonoids in Herba Epimedii by Self Contrast with Correction Factor. China Pharm. 2015, 6, 904–907. [Google Scholar]
- Vo, T.-S.; Kim, S.-K. Fucoidans as a natural bioactive ingredient for functional foods. J. Funct. Foods 2013, 5, 16–27. [Google Scholar] [CrossRef]
- Ganesan, A.R.; Tiwari, U.; Rajauria, G. Seaweed nutraceuticals and their therapeutic role in disease prevention. Food Sci. Hum. Wellness 2019, 8, 252–263. [Google Scholar] [CrossRef]
- Laurienzo, P. Marine Polysccharides; MDPI: Basel, Switzerland, 2018; Volume 1, p. 6. [Google Scholar]
- Park, H.Y.; Han, M.H.; Park, C.; Jin, C.-Y.; Kim, G.-Y.; Choi, I.-W.; Kim, N.D.; Nam, T.-J.; Kwon, T.K.; Choi, Y.H. Anti-inflammatory effects of fucoidan through inhibition of NF-κB, MAPK and Akt activation in lipopolysaccharide-induced BV2 microglia cells. Food Chem. Toxicol. 2011, 49, 1745–1752. [Google Scholar] [CrossRef]
- Mabate, B.; Daub, C.D.; Malgas, S.; Edkins, A.L.; Pletschke, B.I. Fucoidan Structure and Its Impact on Glucose Metabolism: Implications for Diabetes and Cancer Therapy. Mar. Drugs 2021, 19, 30. [Google Scholar] [CrossRef]
- Degn, M.; Lambertsen, K.L.; Petersen, G.; Meldgaard, M.; Artmann, A.; Clausen, B.H.; Hansen, S.H.; Finsen, B.; Hansen, H.S.; Lund, T.M. Changes in brain levels of N-acylethanolamines and 2-arachidonoylglycerol in focal cerebral ischemia in mice. J. Neurochem. 2007, 103, 1907–1916. [Google Scholar] [CrossRef] [PubMed]
- Diep, T.A.; Madsen, A.N.; Holst, B.; Kristiansen, M.M.; Wellner, N.; Hansen, S.H.; Hansen, H.S. Dietary fat decreases intestinal levels of the anorectic lipids through a fat sensor. FASEB J. 2011, 25, 765–774. [Google Scholar] [CrossRef] [PubMed]
- Elagbar, Z.A.; Naik, R.R.; Shakya, A.K.; Bardaweel, S.K. Fatty Acids Analysis, Antioxidant and Biological Activity of Fixed Oil of Annona muricata L. Seeds. J. Chem. 2016, 2016, 6948098. [Google Scholar] [CrossRef]
- Feldman, J.L.; Baeza, J.; Denu, J.M. Activation of the Protein Deacetylase SIRT6 by Long-chain Fatty Acids and Widespread Deacylation by Mammalian Sirtuins. J. Biol. Chem. 2013, 288, 31350–31356. [Google Scholar] [CrossRef]
- Blasbalg, T.L.; Hibbeln, J.R.; Ramsden, C.E.; Majchrzak, S.F.; Rawlings, R.R. Changes in consumption of omega-3 and omega-6 fatty acids in the united states during the 20th century. Am. J. Clin. Nutr. 2011, 93, 950–962. [Google Scholar] [CrossRef]
- Chen, Y.; Sun, T.; Wu, J.; Kalionis, B.; Zhang, C.; Yuan, D.; Huang, J.; Cai, W.; Fang, H.; Xia, S. Icariin intervenes in cardiac inflammaging through upregulation of SIRT6 enzyme activity and inhibition of the NF-kappa B pathway. BioMed. Res. Int. 2015, 2015, 895976. [Google Scholar] [CrossRef]
- Li, N.; Wang, J.; Wang, X.; Sun, J.; Li, Z. Icariin exerts a protective effect against d-galactose induced premature ovarian failure via promoting DNA damage repair. Biomed. Pharmacother. 2019, 118, 109218. [Google Scholar] [CrossRef]
- Li, J.; Jiang, K.; Zhao, F. Icariin regulates the proliferation and apoptosis of human ovarian cancer cells through microRNA-21 by targeting PTEN, RECK and Bcl-2. Oncol. Rep. 2015, 33, 2829–2836. [Google Scholar] [CrossRef]
- Wang, Q.; Hao, J.; Pu, J.; Zhao, L.; Lü, Z.; Hu, J.; Yu, Q.; Wang, Y.; Xie, Y.; Li, G. Icariin induces apoptosis in mouse MLTC-10 Leydig tumor cells through activation of the mitochondrial pathway and down-regulation of the expression of piwil4. Int. J. Oncol. 2011, 39, 973–980. [Google Scholar]
- Ran, L.K.; Chen, Y.; Zhang, Z.Z.; Tao, N.N.; Ren, J.H.; Zhou, L.; Tang, H.; Chen, X.; Chen, K.; Li, W.Y.; et al. SIRT6 Overexpression Potentiates Apoptosis Evasion in Hepatocellular Carcinoma via BCL2-Associated X Protein-Dependent Apoptotic Pathway. Clin. Cancer Res. 2016, 22, 3372–3382. [Google Scholar] [CrossRef]
- De Jesus Raposo, M.F.; de Morais, A.M.B.; de Morais, R.M.S.C. Marine polysaccharides from algae with potential biomedical applications. Mar. Drugs 2015, 13, 2967–3028. [Google Scholar] [CrossRef]
- Costa, L.S.; Fidelis, G.P.; Cordeiro, S.L.; Oliveira, R.M.; Sabry, D.A.; Câmara, R.B.G.; Nobre, L.T.D.B.; Costa, M.S.S.P.; Almeida-Lima, J.; Farias, E.H.C.; et al. Biological activities of sulfated polysaccharides from tropical seaweeds. Biomed. Pharmacother. 2010, 64, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, Q.; Zhang, Z.; Li, Z. Antioxidant activity of sulfated polysaccharide fractions extracted from Laminaria japonica. Int. J. Biol. Macromol. 2008, 42, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Zhang, Z.; Jason, A.; Andrew, F.; Qiuyu, W.; Christopher, S. Bioactive fiber: Bioactivity of Cereal arabinoxylans in Relation to Their Sources and Structure. In Proceedings of the 5th World Congress on Advances in Nutrition, Food Science & Technology, Edinburgh, UK, 11–12 September 2017. [Google Scholar]
- Lim, S.H.; Lee, J. Supplementation with psyllium seed husk reduces myocardial damage in a rat model of ischemia/reperfusion. Nutr. Res. Pract. 2019, 13, 205–213. [Google Scholar] [CrossRef]
- Zhang, S.; Li, W.; Smith, C.J.; Musa, H. Cereal-Derived Arabinoxylans as Biological Response Modifiers: Extraction, Molecular Features, and Immune-Stimulating Properties. Crit. Rev. Food Sci. Nutr. 2015, 55, 1035–1052. [Google Scholar] [CrossRef]
- Huang, Z.; Zhao, J.; Deng, W.; Chen, Y.; Shang, J.; Song, K.; Zhang, L.; Wang, C.; Lu, S.; Yang, X.; et al. Identification of a cellularly active SIRT6 allosteric activator. Nat. Chem. Biol. 2018, 14, 1118–1126. [Google Scholar] [CrossRef]
- Klein, M.A.; Liu, C.; Kuznetsov, V.I.; Feltenberger, J.B.; Tang, W.; Denu, J.M. Mechanism of activation for the sirtuin 6 protein deacylase. J. Biol. Chem. 2020, 295, 1385–1399. [Google Scholar] [CrossRef]
- Li, Z.; Xu, K.; Guo, Y.; Ping, L.; Gao, Y.; Qiu, Y.; Ni, J.; Liu, Q.; Wang, Z. A high-fat diet reverses metabolic disorders and premature aging by modulating insulin and IGF1 signaling in SIRT6 knockout mice. Aging Cell 2020, 19, e13104. [Google Scholar] [CrossRef]
- Masri, S.; Rigor, P.; Cervantes, M.; Ceglia, N.; Sebastian, C.; Xiao, C.; Roqueta-Rivera, M.; Deng, C.; Osborne, T.; Mostoslavsky, R.; et al. Partitioning Circadian Transcription by SIRT6 Leads to Segregated Control of Cellular Metabolism. Cell 2014, 158, 659–672. [Google Scholar] [CrossRef]
- Sun, N.; Yu, Y.; Zhou, L.; Zhang, D.; Wu, X.; Song, C.; Miao, W.; Yang, Z.; Gao, W.; Zhang, C.; et al. Ginsenosides Rc, as a novel SIRT6 activator, protects mice against high fat diet induced NAFLD. J. Ginseng Res. 2020. [Google Scholar] [CrossRef]
- Carreño, M.; Bresque, M.; Machado, M.R.; Santos, L.; Durán, R.; Vitturi, D.A.; Escande, C.; Denicola, A. Nitro-fatty acids as activators of hSIRT6 deacetylase activity. J. Biol. Chem. 2020, 295, 18355–18366. [Google Scholar] [CrossRef]
- Singh, N.; Ravichandran, S.; Norton, D.D.; Fugmann, S.D.; Moaddel, R. Synthesis and characterization of a SIRT6 open tubular column: Predicting deacetylation activity using frontal chromatography. Anal. Biochem. 2013, 436, 78–83. [Google Scholar] [CrossRef]
- Xiong, X.; Wang, G.; Tao, R.; Wu, P.; Kono, T.; Li, K.; Ding, W.-X.; Tong, X.; Tersey, S.; Harris, R.; et al. Sirtuin 6 regulates glucose-stimulated insulin secretion in mouse pancreatic beta cells. Diabetologia 2016, 59, 151–160. [Google Scholar] [CrossRef]
- Babaei, F.; Moafizad, A.; Darvishvand, Z.; Mirzababaei, M.; Hosseinzadeh, H.; Nassiri-Asl, M. Review of the effects of vitexin in oxidative stress-related diseases. Food Sci. Nutr. 2020, 8, 2569–2580. [Google Scholar] [CrossRef]
- Hou, D.; Yousaf, L.; Xue, Y.; Hu, J.; Wu, J.; Hu, X.; Feng, N.; Shen, Q. Mung Bean (Vigna radiata L.): Bioactive Polyphenols, Polysaccharides, Peptides, and Health Benefits. Nutrients 2019, 11, 1238. [Google Scholar] [CrossRef]
- Chen, L.; Zhang, B.; Shan, S.; Zhao, X. Neuroprotective effects of vitexin against isoflurane-induced neurotoxicity by targeting the TRPV1 and NR2B signaling pathways. Mol. Med. Rep. 2016, 14, 5607–5613. [Google Scholar] [CrossRef]
- Singh, C.K.; Chhabra, G.; Ndiaye, M.A.; Garcia-Peterson, L.M.; Mack, N.J.; Ahmad, N. The Role of Sirtuins in Antioxidant and Redox Signaling. Antioxid. Redox Signal 2018, 28, 643–661. [Google Scholar] [CrossRef]
- Scarpa, E.S.; Emanuelli, M.; Frati, A.; Pozzi, V.; Antonini, E.; Diamantini, G.; Di Ruscio, G.; Sartini, D.; Armeni, T.; Palma, F.; et al. Betacyanins enhance vitexin-2-O-xyloside mediated inhibition of proliferation of T24 bladder cancer cells. Food Funct. 2016, 7, 4772–4780. [Google Scholar] [CrossRef]
- Isemura, M. Catechin in Human Health and Disease. Molecules 2019, 24, 528. [Google Scholar] [CrossRef]
- Tao, L.; Park, J.Y.; Lambert, J.D. Differential prooxidative effects of the green tea polyphenol, (-)-epigallocatechin-3-gallate, in normal and oral cancer cells are related to differences in sirtuin 3 signaling. Mol. Nutr. Food Res. 2015, 59, 203–211. [Google Scholar] [CrossRef]
- Zhao, S.; Zhu, Y.-Y.; Wang, X.-Y.; Liu, Y.-S.; Sun, Y.-X.; Zhao, Q.-J.; Li, H.-Y. Structural Insight into the Interactions between Structurally Similar Inhibitors and SIRT6. Int. J. Mol. Sci. 2020, 21, 2601. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Wu, X.; Zhao, B.; Wang, Y. Scutellarin protects human retinal pigment epithelial cells against hydrogen peroxide (H2O2)-induced oxidative damage. Cell Biosci. 2019, 9, 12. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Yan, D.; Zhao, H.Y.; Zhang, F.C.; Zhao, W.H.; Feng, M. SIRT6 suppresses glioma cell growth via induction of apoptosis, inhibition of oxidative stress and suppression of JAK2/STAT3 signaling pathway activation. Oncol. Rep. 2015, 35, 1395–1402. [Google Scholar] [CrossRef]
- Sun, C.-Y.; Nie, J.; Zheng, Z.-L.; Zhao, J.; Wu, L.-M.; Zhu, Y.; Su, Z.-Q.; Zheng, G.-J.; Feng, B. Renoprotective effect of scutellarin on cisplatin-induced renal injury in mice: Impact on inflammation, apoptosis, and autophagy. Biomed. Pharmacother. 2019, 112, 108647. [Google Scholar] [CrossRef] [PubMed]
- Fricker, R.A.; Green, E.L.; Jenkins, S.I.; Griffin, S.M. The Influence of Nicotinamide on Health and Disease in the Central Nervous System. Int. J. Tryptophan Res. 2018, 11, 1178646918776658. [Google Scholar] [CrossRef] [PubMed]
- Alenzi, F. Effect of Nicotinamide on Experimental Induced Diabetes. Iran. J. Allergy Asthma Immunol. 2009, 8, 11–18. [Google Scholar]
- Verruck, S.; Balthazar, C.F.; Rocha, R.S.; Silva, R.; Esmerino, E.A.; Pimentel, T.C.; Freitas, M.Q.; Silva, M.C.; da Cruz, A.G.; Prudencio, E.S. Chapter Three—Dairy foods and positive impact on the consumer’s health. In Advances in Food and Nutrition Research; Toldrá, F., Ed.; Academic Press: Cambridge, MA, USA, 2019; Volume 89, pp. 95–164. [Google Scholar]
- Chowdhury, R.; Steur, M.; Patel, P.S.; Franco, O.H. Chapter 10—Individual Fatty Acids in Cardiometabolic Disease. In Handbook of Lipids in Human Function; Watson, R.R., de Meester, F., Eds.; AOCS Press: Cambridge, MA, USA, 2016; pp. 207–318. [Google Scholar]
- Hu, J.; Jing, H.; Lin, H. Sirtuin inhibitors as anticancer agents. Future Med. Chem. 2014, 6, 945–966. [Google Scholar] [CrossRef] [PubMed]
- Kokkonen, P.; Rahnasto-Rilla, M.; Kiviranta, P.H.; Huhtiniemi, T.; Laitinen, T.; Poso, A.; Jarho, E.; Lahtela-Kakkonen, M. Peptides and Pseudopeptides as SIRT6 Deacetylation Inhibitors. ACS Med. Chem. Lett. 2012, 3, 969–974. [Google Scholar] [CrossRef][Green Version]
- Schepper, S.; Bruwiere, H.; Verhulst, T.; Steller, U.; Andries, L.; Wouters, W.; Janicot, M.; Arts, J.; Heusden, J. Inhibition of Histone Deacetylases by Chlamydocin Induces Apoptosis and Proteasome-Mediated Degradation of Survivin. J. Pharmacol. Exp. Ther. 2003, 304, 881–888. [Google Scholar] [CrossRef]


{kind=link}
| Disease | Molecular Mechanism of Disease | In Vitro/In Vivo Studies | Expression of SIRT6 Level | Role of SIRT6 | References |
|---|---|---|---|---|---|
| Alzheimer’s disease | DNA damage | Study was conducted on mice | Deficiency or reduced level of SIRT6 | SIRT6 repairs DNA damage recruiting chromatin remodeler SNF2H and deacylating H3K56 on those damaged sites | [12,14,19] |
| Aging | DNA damage, abnormal transcription and destabilization of retrotransposon, telomeres, and centromeres, dysfunction of telomeres, impaired heterochromatin sustentation | Study was carried out on aged tissues and cells of SIRT6 KO mice, human fibroblasts, and premature cells; SIRT6 KD cells | Absence or deficiency or decreased level of SIRT6 | SIRT6 stabilizes repressive heterochromatin at the sub-telomeric region through silencing transcription of telomere proximal genes/activates PARP1/provides chromatin-regulatory activity/deacylates whole chromatin and nucleosomes | [11,21,22,26,27,28,31,32,34,35,36] |
| Cancer | Activation of HIF-1α factor, suppression of Lin28 and Lin28b oncofetal proteins, suppression of survivin protein, activation of Bax protein transcription | Studies conducted in patients with liver and chronic lymphocytic leukemia (CLL); patients, human cancer tissues or cells are studied | Downregulation in colorectal, breast, ovarian, hepatocellular, and lung cancers and p-regulation of SIRT6 in prostate, skin, CLL, and pancreatic cancers | SIRT6 inhibits the HIF-1α factor; activates Lin28, Lin28b, and survivin proteins; suppresses Bax protein transcription | [37,38,39,40,41,42] |
| Inflammation | NF-κB signal deactivation leads to a reduction in proinflammatory cytokines and anti-apoptotic gene generation, reduced expression of IL8 and TNF | FKO mice, mouse immune cells | Absence of SIRT6/deficiency of SIRT6 | SIRT6 can regulate several NF-κB gene transcriptions, SIRT6 has a role in promoting the expression of proinflammatory cyto-chemokines such as interleukin-8 (IL8) and TNF | [14,29,41,42,43] |
| Diabetes | Decreased uptake of glucose by tissue, low expression of GLUT1; increased glycolysis and mitochondrial respiration suppression; increased level of HIF1-α, decreased acetylation levels of PGC-1 α; inhibition of protein kinase, Akt, low expression of Pdx1 and Gult2 | Studies are conducted in β-specific KO mice | Enhanced level of SIRT6 | Reduced level of SIRT6 upregulates GLUT1, Pdx1, and glut2; downregulates the level of HIF1-α; increases acetylation of PGC-1 α; stimulates protein kinase Akt, thus regulates glucose homeostasis | [43,44,45,46,47,48] |
| Compound (Class) | Source | Mode of Action against the Selected Diseases | Role in SIRT6 Activation | References |
|---|---|---|---|---|
| Quercetin (flavonoid) | Onions, shallots, broccoli, peppers, caper fruits, apples, berries, grapes, herbs, tea, and wine | Anticancer activity: Inhibition of tyrosine kinase in vivo, inhibition of PI3K (phosphatidylinisitol-3-kinase)-Akt-PKB (protein kinase B) pathway Antidiabetic effect: Scavenging reactive oxygen species (ROS) such as peroxynitrile, hydroxyl radicals, and superoxide anions Anti-inflammatory action: Prevention of cytokines including IL12, IL8, INFγ, INFα, COX-2, PGE | To activate SIRT6 quercetin binds to SIRT6-selective acyl binding channel and activation of SIRT6 provides all these bioactivities though different mechanisms | [56,74,75] |
| Isoquercetin (flavonoid) | Medicinal herbs, fruits, beverages, vegetables, onions, mangoes, Tartary buckwheat bran, Chinese and hawthorn fruits | Cytotoxic/anticancer activity: Inhibition of protein kinase B phopsphorylation, and thus surviving protein activates caspases and reduces anti-apoptotic proteins, i.e., Bcl-2 and Mcl-1 Antioxidant effect: Scavenging ROS Anti-inflammatory action: Regulation of Nrf2 pathway-associated protein and gene expression Antidiabetic effect: Reduction of oxidative stress, and also regulation of proteins and genes that associated with Nrf2 pathway. | Isoquercetin binds with SIRT6-selective acyl binding channel through its bulky sugar moiety to activate SIRT6 and activation of SIRT6 exerts these bioactivities | [56,61,63,64,65,66,67,74] |
| Kaempferol (flavonoid) | Green leafy vegetables such as spinach, kale, herbs, dills, chives, tarragon, wild leeks, and ramps | Neuroprotective effect in AD: Induction of anti-apoptotic activity in Aβ-induced SH-SY5Y neuronal cells. Antioxidant activity: Scavenging superoxide anions, hydroxyl radicals, decrease peroxynitrile levels, inhibition of xanthine oxidase enzyme. Anticancer potential: Mechanism involves apoptosis, cell cycle arrest at the G2/M phase, downregulation of epithelial-mesenchymal transition (EMT)-related markers, and phosphoinositide 3-kinase/protein kinase B signaling pathways. Anti-inflammatory effect: Inhibition of NF-κB activity and TNF activity, reduction of expression of IL-1 and IL-8, inhibition of the activation of AP-1, COX-2 Antidiabetic activity: Reduction of ROS level | To activate SIRT6 kaempferol binds to the SIRT6-specific acyl bonding channel and activation of SIRT6 results in these biological actions | [68,69,70] |
| Luteolin (flavonoid) | Carrots, peppers, celery, olive oil, peppermint, thyme, rosemary, lettuce, pomegranate, turnip, capers, cucumber, lemon, beets, brussels sprouts, cabbage, cauliflower, chives, fennel, harwort, horseradish, kohlrabi, parsley, spinach, and green tea | Neuroprotective action: ROS scavenging Antioxidant activity: ROS scavenging through elevation of SIRT6 expression Anticancer potential: Inhibition of topoisomerase I and II Anti-inflammatory action: Selective inhibition of COX-2, reduction of NFkappaB and AP-1 activity | Luteolin activates SIRT6 by binding with SIRT6-specific acyl binding channel. Stimulation of SIRT6 deacetylation activity contributes to these biological effects | [71,72,73] |
| Cyanidin (anthocyanidin) | Berries, black currant, grapes | Anti-aging property: Inhibition of ROS and nitrogen species production Anticancer activity: Upregulation of SIRT6 associated gene, FOXO3α; downregulation of SIRT6 associated Twist1 and GLUT1 genes; inhibition of epidermal growth factor receptor Antioxidant activity: ROS scavenging | To activate SIRT6 cyanidin binds with SIRT6-selective acyl binding site and in the same mood as quercetin. Stimulation of SIRT6 contributes to anti-aging, antiancer, and antioxidant effects | [52,76,77] |
| Fisetin (flavonoid) | Apples, grapes, persimmons, strawberries, cucumbers, and onions | Anti-aging property: Reduction of Ckd5 activator p35 cleavage product, p25 in the brains of Alzheimer’s disease patient Anticancer effect: Inhibition of Akt/mTOR signaling pathways, activation of Caspase-7 and-9 Antidiabetic activity: Reduction of methylglyoxal dependent protein glycation Antioxidant potential: Inhibition of ROS production Anti-inflammatory action: Suppression of NF-κB in vascular inflammatory responses | Structural basis of SIRT6 activation by fisetin is not well known, however, activation of SIRT6 brings these biological responses | [53,78,79] |
| Delphinidin (anthocyanidin) | Flowers, fruits, vegetables, and grains | Anti-aging action: ROS and free radical scavenging Anti-inflammatory activity: Reduction of IL-1β and suppression of NF-κ B pathway in vitro Anticancer effect: Induction of apoptosis, autophagy thorough mTOR signaling pathway suppression, and AMPK pathway activation | Delphinidin activates SIRT6 by binding to a binding site which is next to a loop near the acetylated peptide substrate binding site of SIRT6 and activation of SIRT6 results in anti-aging, anti-inflammatory, and anticancer effects | [80,81,82,83,84] |
| Icariin (prenylated flavonoid glycoside) | Herba epimedii | Anticancer potential: Inhibition of cell proliferation, accelerated apoptosis, and caspase-3 activity by targeting PTEN, RECK, and Bcl-2 protein expression in ovarian cancer cells (A2780); regulation of the expression of Bcl-2/Bax and cytochrome c, activation of caspase-9, and -3 in MLTC-1 mouse tumor cells. Anti-inflammatory effect: Reduction of NF-κB protein expression Anti-aging effect: Inhibition of p53/p21 and NF-κB signaling pathways, overexpression of SIRT6 | Structural basis of SIRT6 activation by icariin is not well known, however, activation of SIRT6 brings all these biological responses | [85,86] |
| Fucoidan (polysaccharide) | Seaweeds and brown algae | Antioxidant effect: NO scavenging Anticancer potential: Induction of apoptosis in HepG2 liver cancer cells by upregulation of p53 and p14 and stimulation of caspases activity Anti-inflammatory action: Inhibition of NO production; downregulation of iNOS, COX-2, IL-1 β, TNF-α, NF- κB expression, ERK, JNK, MAPK, and Akt pathways. Antidiabetic effect: Stimulation of SIRT6 by p53, and enhancement of FoxO1 level. Activation of PI3K/PKB pathway, which regulates insulin production and stimulate GLUT4 translocation | Structural basis of SIRT6 activation by fucoidan is not well known, however, activation of SIRT6 brings several biological responses | [87,88,89,90,91] |
| N-acylethanolamines (NAEs) (lipid) | Endogenous molecules | Neuroprotection: Regulation of neuroprotection in focal cerebral ischemia in mice Anticancer and anti-inflammatory action: Upregulation of SIRT6 | These compounds bind with a large hydrophobic pocket of SIRT6 and they stimulate SIRT6 to produce neuroprotection, anticancer, and anti-inflammatory effects | [60,92] |
| Oleic acid (fatty acid) | Plant-based oil (olive oil), nuts, and vegetable | Antioxidant activity: Free radical scavenging | Oleic acid stimulates deacetylation activity of SIRT6 by binding with a large hydrophobic pocket of SIRT6 and provides antioxidant activity | [60,92,93,94,95] |
| Linoleic acid (fatty acid) | Plant-based oil, nut, meat, and animal products | Antioxidant activity: Free radical scavenging | Same as oleic acid | [60,93,94,96] |
| Compound (Class) | Source | Mode of Action against the Selected Diseases | Role in SIRT6 Inhibition | References |
|---|---|---|---|---|
| Quercetin (flavonoid) | Onions, shallots, broccoli, peppers, caper fruits, apples, berries, grapes, herbs, tea, and wine | Anticancer and antidiabetic activity: Inhibition of SERCA (sarco/endoplasmic reticulum Ca2+-ATPase) | To inhibit SIRT6 quercetin binds with an alternative site on the entrance of active site of SIRT6 and inhibition of SIRT6 produces anticancer and antidiabetic effects | [56,115] |
| Vitexin (apigenin flavones glycoside) | Hawthorn berry, mung beans, bamboo, buckwheat, echinodorus, and passiflora | Antioxidant effect: Reduction of ROS level and enhancement of GSH and SOD level Neuroprotective effect in AD: Down egulation of HIF1-α and VEGF and maintenance of blood brain barrier (BBB) integrity, reduction of brain edema Anticancer activity: Apoptosis, regulation of apoptosis-related gene expression of p53 and bcl-2 Anti-inflammatory effect: Inhibition of IL-1β, IL-6, IL-8, IL-17, and IL-33, TNF-α, NFκ-B, and iNOS | Vitexin inhibits SIRT6 by binding to a site close to the NAM binding site of NAD+ and inhibition of SIRT6 contributes to therapeutic potential in different diseases | [52,53,61,116] |
| Catechin (polyphenol) | Variety of foods and herbs including tea, apples, persimmons, cacaos, grapes, and berries | Cytoprotective, anticancer, antidiabetic and neuroprotective effects: Reduction of oxidative stress and inhibition of DNA damage | Catechin inhibits SIRT6 by hydrophobic and π-stacking interactions and configuration matching which results in reduction of oxidative stress and DNA damage | [52,121,122,123] |
| Scutellarin (flavone) | Scutellaria baicalensis | Chemotherapeutic effect: Activation of JAK2/STAT3 signaling pathway Antioxidant activity: Reduction of ROS and MDA level, enhancement of SOD and GSH level Anti-inflammatory effect: Inhibition of proinflammatory cytokines, TNF-α, and IL-6 release | Scutellarin inhibits SIRT6 activity through hydrophobic and, potentially, π-stacking interaction, and thus provides chemotherapeutic, antioxidant and anti-inflammatory effects | [124,125,126] |
| Nicotinamide (amide form of vitamin B3-Niacin) | Eggs, meat, fish, and mushrooms | Anticancer activity: Apoptosis, DNA repairing, and genome stability Antidiabetic action: Induction of antidiabetic action by protecting pancreatic β-cells through prevention of apoptosis and NO generation Anti-aging: Reduction of ROS level | As a non-competitive inhibitor of SIRT6 nicotinamide reforms substrates by binding to the enzyme and reacting with the substrate-ADP-ribose adduct. Inhibition of SIRT6 is associated with various health benefits | [109,127,128] |
| Myristic acid (fatty acid) | Milk fat, nutmeg, palm kernel, and butter | Anti-inflammatory effect: Suppression of NF-κB signaling pathway and emission of TNF-α Anticancer potential: Genome stabilization, DNA repairing | Myristic acid inhibits SIRT6 competing with peptide-conjugated myristoyl chain for the same hydrophobic pocket of SIRT6 protein, and thus it plays a role in inhibiting inflammation and cancer | [109,129,130] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akter, R.; Afrose, A.; Rahman, M.R.; Chowdhury, R.; Nirzhor, S.S.R.; Khan, R.I.; Kabir, M.T. A Comprehensive Analysis into the Therapeutic Application of Natural Products as SIRT6 Modulators in Alzheimer’s Disease, Aging, Cancer, Inflammation, and Diabetes. Int. J. Mol. Sci. 2021, 22, 4180. https://doi.org/10.3390/ijms22084180
Akter R, Afrose A, Rahman MR, Chowdhury R, Nirzhor SSR, Khan RI, Kabir MT. A Comprehensive Analysis into the Therapeutic Application of Natural Products as SIRT6 Modulators in Alzheimer’s Disease, Aging, Cancer, Inflammation, and Diabetes. International Journal of Molecular Sciences. 2021; 22(8):4180. https://doi.org/10.3390/ijms22084180
Chicago/Turabian StyleAkter, Raushanara, Afrina Afrose, Md. Rashidur Rahman, Rakhi Chowdhury, Saif Shahriar Rahman Nirzhor, Rubayat Islam Khan, and Md. Tanvir Kabir. 2021. "A Comprehensive Analysis into the Therapeutic Application of Natural Products as SIRT6 Modulators in Alzheimer’s Disease, Aging, Cancer, Inflammation, and Diabetes" International Journal of Molecular Sciences 22, no. 8: 4180. https://doi.org/10.3390/ijms22084180
APA StyleAkter, R., Afrose, A., Rahman, M. R., Chowdhury, R., Nirzhor, S. S. R., Khan, R. I., & Kabir, M. T. (2021). A Comprehensive Analysis into the Therapeutic Application of Natural Products as SIRT6 Modulators in Alzheimer’s Disease, Aging, Cancer, Inflammation, and Diabetes. International Journal of Molecular Sciences, 22(8), 4180. https://doi.org/10.3390/ijms22084180