
Whole Alga, Algal Extracts, and Compounds as Ingredients of Functional Foods: Composition and Action Mechanism Relationships in the Prevention and Treatment of Type-2 Diabetes Mellitus

 , ,
, ,
Abstract
1. Introduction
2. Type-2 Diabetes Mellitus
2.1. Microbiota Dysbiosis
2.2. Genetics
3. Algae
3.1. Algae Definition and Classification
3.2. Algal Consumption and Commercial Importance
3.3. Algae as Functional Foods or as a Potential Raw Material for Bioactive Ingredients
4. Algal Composition, Structure, and Beneficial Effects on Type-2 Diabetes Mellitus
4.1. Energy
4.2. Polysaccharides and Fiber
4.3. Protein and Amino Acid Contents
4.4. Lipids, Fatty Acids, and the Unsaponifiable Fraction
4.5. Mineral and Trace Elements
4.6. Vitamins and Related Compounds
4.7. Antioxidants: Polyphenols and Related Compounds
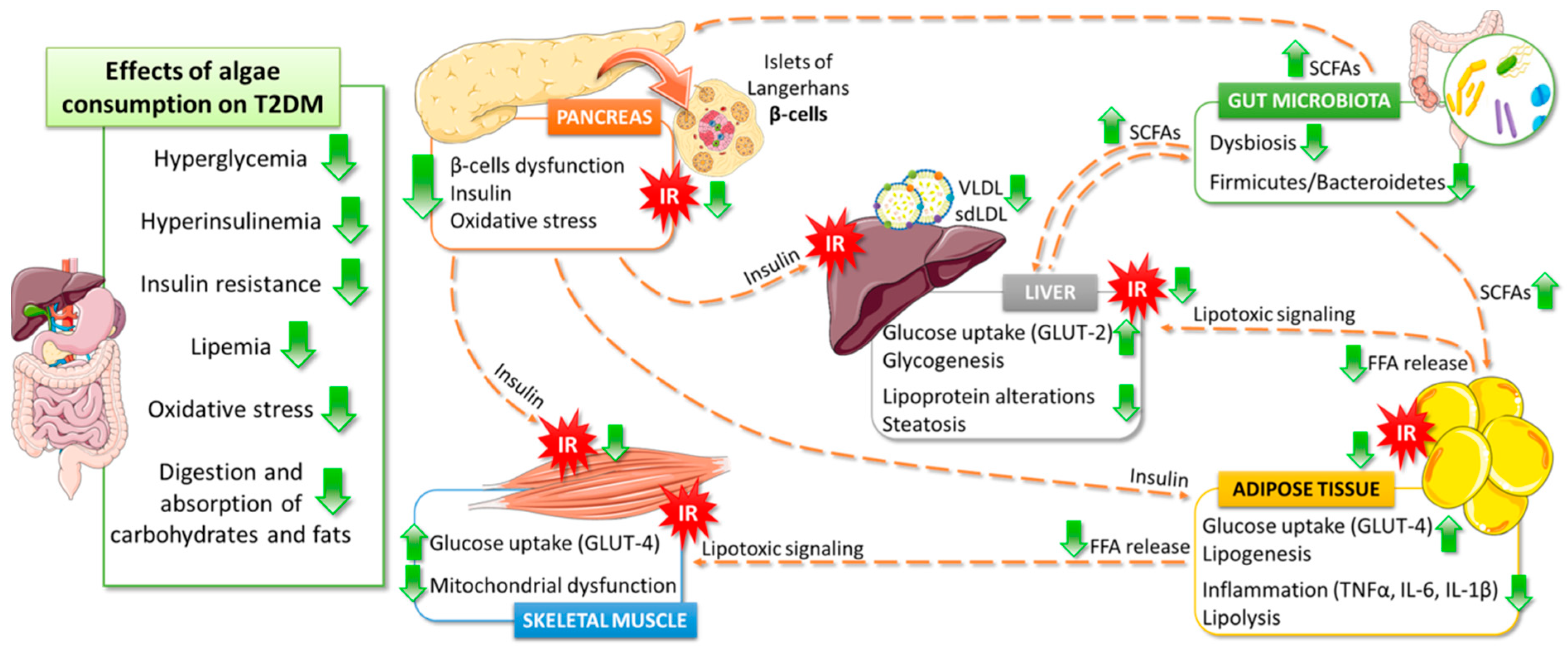
5. Effects of Algae Consumption on T2DM Pathophysiology
5.1. Glucose Homeostasis
5.2. Lipids Metabolism
5.3. Gut Microbiota
5.4. Antioxidant Properties
6. Conclusions and Future Remarks
- The ingredients, fiber, polysaccharides, PUFAs, and polyphenol compounds have been most associated with improvement in T2DM.
- Mechanisms have been proposed to explain the beneficial effects of the algae on T2DM. However, not all studies fully elucidate the factors involved in beneficial properties of algae dietary intake in managing diabetes.
- Algae have been postulated as promising antidiabetic agents, effectively reducing carbohydrates digestion and absorption, regulating the α-glucosidase activity.
- Green and brown algae seem to modulate and restore intestinal dysbiosis associated with T2DM, however, there are no studies evaluating the potential of red algae in relation to this aspect.
- The large number of bioactive compounds present in algae makes them powerful antioxidant agents, alleviating the oxidative stress associated with T2DM.
- A variety of algae can be useful for developing functional foods aimed at people with T2DM, reducing the risks of acute and chronic diabetic complications. However, it is important to study the optimization of algae extraction conditions to maximize yields of the active compounds and on the properties of algal compounds useful for the antidiabetic benefits required.
- Clean habitat where algae grow should be absolutely guaranteed.
- Further investigations in diabetic individuals are required to elucidate the mechanisms involved in preventing, reducing, and controlling diabetes.
- Identification of genetic factors affecting dietary response to algal compounds may assist in the development of targeted and potentially more efficient dietary interventions.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Global Report on Diabetes. 2016. Available online: https://www.who.int/diabetes/publications/grd-2016/en/ (accessed on 10 January 2021).
- Kolb, H.; Martin, S. Environmental/lifestyle factors in the pathogenesis and prevention of type 2 diabetes. BMC Med. 2017, 15, 1–11. [Google Scholar] [CrossRef]
- Laakso, M. Biomarkers for type 2 diabetes. Mol. Metab. 2019, 27, S139–S146. [Google Scholar] [CrossRef]
- Stančáková, A.; Laakso, M. Genetics of type 2 diabetes. Endocr. Dev. 2016, 31, 203–220. [Google Scholar] [CrossRef]
- Sharma, S.; Tripathi, P. Gut microbiome and type 2 diabetes: Where we are and where to go? J. Nutr. Biochem. 2019, 63, 101–108. [Google Scholar] [CrossRef] [PubMed]
- PR Newswire. Transparency Market Research. Available online: https://www.prnewswire.com/news-releases/global-algae-market-is-projected-to-be-worth-us11-bn-by-2024-at-a-cagr-of-739-global-industry-analysis-size-share-growth-trends-and-forecast-2016---2024-tmr-594253011.html (accessed on 12 January 2021).
- Koyande, A.K.; Chew, K.W.; Rambabu, K.; Tao, Y.; Chu, D.-T.; Show, P.-L. Microalgae: A potential alternative to health supplementation for humans. Food Sci. Hum. Wellness 2019, 8, 16–24. [Google Scholar] [CrossRef]
- Hannon, B.A.; Fairfield, W.D.; Adams, B.; Kyle, T.; Crow, M.; Thomas, D.M. Use and abuse of dietary supplements in persons with diabetes. Nutr. Diabetes 2020, 10, 14. [Google Scholar] [CrossRef]
- Jiménez-Escrig, A.; Goñi, I. Nutritional evaluation and physiological effects of edible seaweeds. Arch. Latinoam. Nutr. 1999, 49, 114–120. [Google Scholar] [PubMed]
- Bocanegra, A.; Bastida, S.; Benedí, J.; Ródenas, S.; Sánchez-Muniz, F.J. Characteristics and nutritional and cardiovascular-health properties of seaweeds. J. Med. Food 2009, 12, 236–258. [Google Scholar] [CrossRef]
- Brown, E.M.; Allsopp, P.J.; Magee, P.J.; Gill, C.I.R.; Nitecki, S.; Strain, C.R.; McSorley, E.M. Seaweed and human health. Nutr. Rev. 2014, 72, 205–216. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.-Y.; Huang, X.; Cheong, K.-L. Recent advances in marine algae polysaccharides: Isolation, structure, and activities. Mar. Drugs 2017, 15, 388. [Google Scholar] [CrossRef]
- Peña-Romero, A.C.; Navas-Carrillo, D.; Marín, F.; Orenes-Piñero, E. The future of nutrition: Nutrigenomics and nutrigenetics in obesity and cardiovascular diseases. Crit. Rev. Food Sci. Nutr. 2018, 58, 3030–3041. [Google Scholar] [CrossRef]
- Mathers, J.C. Nutrigenomics in the modern era. Proc. Nutr. Soc. 2017, 76, 265–275. [Google Scholar] [CrossRef]
- Tiffon, C. The impact of nutrition and environmental epigenetics on human health and disease. Int. J. Mol. Sci. 2018, 19, 3425. [Google Scholar] [CrossRef]
- Amar, J.; Chabo, C.; Waget, A.; Klopp, P.; Vachoux, C.; Bermúdez-Humarán, L.G.; Smirnova, N.; Bergé, M.; Sulpice, T.; Lahtinen, S.; et al. Intestinal mucosal adherence and translocation of commensal bacteria at the early onset of type 2 diabetes: Molecular mechanisms and probiotic treatment. EMBO Mol. Med. 2011, 3, 559–572. [Google Scholar] [CrossRef] [PubMed]
- Kolodziejczyk, A.A.; Zheng, D.; Shibolet, O.; Elinav, E. The role of the microbiome in NAFLD and NASH. EMBO Mol. Med. 2019, 11, e9302. [Google Scholar] [CrossRef]
- Barbier, M.; Charrier, B.; Araujo, R.; Holdt, S.L.; Jacquemin, B.; Rebours, C. PEGASUS—Phycomorph European Guidelines for a Sustainable Aquaculture of Seaweeds; Barbier, M., Charrier, B., Eds.; COST Action FA1406: Roscoff, France, 2019. [Google Scholar] [CrossRef]
- Holdt, S.L.; Kraan, S. Bioactive compounds in seaweed: Functional food applications and legislation. J. Appl. Phycol. 2011, 23, 543–597. [Google Scholar] [CrossRef]
- Macho-González, A.; Bastida, S.; Garcimartín, A.; López-Oliva, M.E.; González, P.; Benedí, J.; González-Muñoz, M.J.; Sánchez-Muniz, F.J. Functional meat products as oxidative stress modulators: A review. Adv. Nutr. 2021, nmaa182. [Google Scholar] [CrossRef] [PubMed]
- Mirmiran, P.; Bahadoran, Z.; Azizi, F. Functional foods-based diet as a novel dietary approach for management of type 2 diabetes and its complications: A review. World J. Diabetes 2014, 5, 267–281. [Google Scholar] [CrossRef] [PubMed]
- Schultz, A.R.; Garcimartín, A.; Bastida, S.; Jiménez-Escrig, A.; Rupérez, P.; Green, B.D.; Rafferty, E.; Sánchez-Muniz, F.J.; Benedí, J. Effects of Undaria pinnatifida, Himanthalia elongata and Porphyra umbilicalis extracts on in vitro α-glucosidase activity and glucose diffusion. Nutr. Hosp. 2014, 29, 1434–1446. [Google Scholar]
- Garcimartín, A.; Benedi, J.; Bastida, S.; Sánchez-Muniz, F.J. Aqueous extracts and suspensions of restructured pork formulated with Undaria pinnatifida, Himanthalia elongata and Porphyra umbilicalis distinctly affect the in vitro α-glucosidase activity and glucose diffusion. LWT Food Sci. Technol. 2015, 64, 720–726. [Google Scholar] [CrossRef]
- Ródenas de la Rocha, S.; Sánchez-Muniz, F.; Gómez-Juaristi, M.; Larrea-Marín, M. Trace elements determination in edible seaweeds by an optimized and validated ICP-MS method. J. Food Compos. Anal. 2009, 22, 330–336. [Google Scholar] [CrossRef]
- Larrea-Marín, M.; Pomares-Alfonso, M.; Gómez-Juaristi, M.; Sánchez-Muniz, F.; de la Rocha, S.R. Validation of an ICP-OES method for macro and trace element determination in Laminaria and Porphyra seaweeds from four different countries. J. Food Compos. Anal. 2010, 23, 814–820. [Google Scholar] [CrossRef]
- Cherry, P.; O’Hara, C.; Magee, P.J.; McSorley, E.M.; Allsopp, P.J. Risks and benefits of consuming edible seaweeds. Nutr. Rev. 2019, 77, 307–329. [Google Scholar] [CrossRef] [PubMed]
- American Diabetes Association. Diagnosis and classification of diabetes mellitus. Diabetes Care 2013, 37, S81–S90. [Google Scholar] [CrossRef]
- Schlienger, J.-L. Complications du diabète de type 2. Presse Med. 2013, 42, 839–848. [Google Scholar] [CrossRef]
- Macho-González, A.; López-Oliva, M.E.; Merino, J.J.; García-Fernández, R.A.; Garcimartín, A.; Redondo-Castillejo, R.; Bastida, S.; Sánchez-Muniz, F.J.; Benedí, J. Carob fruit extract-enriched meat improves pancreatic beta-cell dysfunction, hepatic insulin signaling and lipogenesis in late-stage type 2 diabetes mellitus model. J. Nutr. Biochem. 2020, 84, 108461. [Google Scholar] [CrossRef]
- Beale, E.G. Insulin signaling and insulin resistance. J. Investig. Med. 2013, 61, 11–14. [Google Scholar] [CrossRef]
- Sánchez-Muniz, F.J.; García-Quismondo, A.; del Cañizo, F.J.; Dorado, J. Classical and emergent cardiovascular disease risk factors in type 2 diabetics from the Vallecas area (DICARIVA study). Nutr. Hosp. 2017, 34, 1432–1441. [Google Scholar] [CrossRef]
- Bays, H.E. “Sick Fat,” metabolic disease, and atherosclerosis. Am. J. Med. 2009, 122, S26–S37. [Google Scholar] [CrossRef]
- Carmena, R. Dyslipemia in type 2 diabetes mellitus. In Type 2 Diabetes Mellitus; Serrano, M., Gutiérrez, J.A., Eds.; Elsevier España S.L.: Barcelona, Spain, 2010; pp. 219–230. [Google Scholar]
- Chehade, J.M.; Gladysz, M.; Mooradian, A.D. Dyslipidemia in type 2 diabetes: Prevalence, pathophysiology, and management. Drugs 2013, 73, 327–339. [Google Scholar] [CrossRef]
- Athyros, V.G.; Doumas, M.; Imprialos, K.P.; Stavropoulos, K.; Georgianou, E.; Katsimardou, A.; Karagiannis, A. Diabetes and lipid metabolism. Hormones 2018, 17, 61–67. [Google Scholar] [CrossRef]
- Tangvarasittichai, S. Oxidative stress, insulin resistance, dyslipidemia and type 2 diabetes mellitus. World J Diabetes. 2015, 6, 456–480. [Google Scholar] [CrossRef]
- Qin, H.; Chen, Z.; Zhang, Y.; Wang, L.; Ouyang, P.; Cheng, L.; Zhang, Y. Triglyceride to high-density lipoprotein cholesterol ratio is associated with incident diabetes in men: A retrospective study of Chinese individuals. J. Diabetes Investig. 2020, 11, 192–198. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Hu, H.; Chen, M.; Luo, X.; Yao, W.; Liang, Q.; Yang, F.; Wang, X. Association of Triglyceride to high-density lipoprotein cholesterol ratio and incident of diabetes mellitus: A secondary retrospective analysis based on a Chinese cohort study. Lipids Health Dis. 2020, 19, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Ojo, O. Dietary intake and type 2 diabetes. Nutrients 2019, 11, 2177. [Google Scholar] [CrossRef] [PubMed]
- Burgio, E.; Lopomo, A.; Migliore, L. Obesity and diabetes: From genetics to epigenetics. Mol. Biol. Rep. 2014, 42, 799–818. [Google Scholar] [CrossRef] [PubMed]
- Gesteiro, E.; Bastida, S.; Vázquez-Velasco, M.; Corella, D.; Guillén, M.; Ordovas, J.M.; Sánchez-Muniz, F.J. Effects of APOA5 S19W polymorphism on growth, insulin sensitivity and lipoproteins in normoweight neonates. Eur. J. Pediatr. 2011, 170, 1551–1558. [Google Scholar] [CrossRef] [PubMed]
- Gesteiro, E.; Sánchez-Muniz, F.J.; Ortega-Azorín, C.; Guillén, M.; Corella, D.; Bastida, S. Maternal and neonatal FTO rs9939609 polymorphism affect insulin sensitivity markers and lipoprotein profile at birth in appropriate-for-gestational-age term neonates. J. Physiol. Biochem. 2016, 72, 169–181. [Google Scholar] [CrossRef]
- Alkhatib, A.; Tsang, C.; Tiss, A.; Bahorun, T.; Arefanian, H.; Barake, R.; Khadir, A.; Tuomilehto, J. Functional Foods and Lifestyle Approaches for Diabetes Prevention and Management. Nutrients 2017, 9, 1310. [Google Scholar] [CrossRef]
- Zhao, M.; Liao, D.; Zhao, J. Diabetes-induced mechanophysiological changes in the small intestine and colon. World J. Diabetes 2017, 8, 249–269. [Google Scholar] [CrossRef]
- Leylabadlo, H.E.; Sanaie, S.; Heravi, F.S.; Ahmadian, Z.; Ghotaslou, R. From role of gut microbiota to microbial-based therapies in type 2-diabetes. Infect. Genet. Evol. 2020, 81, 104268. [Google Scholar] [CrossRef] [PubMed]
- Yassour, M.; Lim, M.Y.; Yun, H.S.; Tickle, T.L.; Sung, J.; Song, Y.-M.; Lee, K.; Franzosa, E.A.; Morgan, X.C.; Gevers, D.; et al. Sub-clinical detection of gut microbial biomarkers of obesity and type 2 diabetes. Genome Med. 2016, 8, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Camargo, A.; Jimenez-Lucena, R.; Alcala-Diaz, J.F.; Rangel-Zuñiga, O.A.; Garcia-Carpintero, S.; Lopez-Moreno, J.; Blanco-Rojo, R.; Delgado-Lista, J.; Perez-Martinez, P.; van Ommen, B.; et al. Postprandial endotoxemia may influence the development of type 2 diabetes mellitus: From the CORDIOPREV study. Clin. Nutr. 2019, 38, 529–538. [Google Scholar] [CrossRef] [PubMed]
- Singer-Englar, T.; Barlow, G.; Mathur, R. Obesity, diabetes, and the gut microbiome: An updated review. Expert Rev. Gastroenterol. Hepatol. 2019, 13, 3–15. [Google Scholar] [CrossRef]
- Gurung, M.; Li, Z.; You, H.; Rodrigues, R.; Jump, D.B.; Morgun, A.; Shulzhenko, N. Role of gut microbiota in type 2 diabetes pathophysiology. EBioMedicine 2020, 51, 102590. [Google Scholar] [CrossRef]
- Sircana, A.; Framarin, L.; Leone, N.; Berrutti, M.; Castellino, F.; Parente, R.; de Michieli, F.; Paschetta, E.; Musso, G. Altered gut microbiota in type 2 diabetes: Just a coincidence? Curr. Diabetes Rep. 2018, 18, 98. [Google Scholar] [CrossRef] [PubMed]
- Gérard, C.; Vidal, H. Impact of gut microbiota on host glycemic control. Front. Endocrinol. 2019, 10, 29. [Google Scholar] [CrossRef]
- Mohlke, K.L.; Boehnke, M. Recent advances in understanding the genetic architecture of type 2 diabetes. Hum. Mol. Genet. 2015, 24, R85–R92. [Google Scholar] [CrossRef]
- Merino, J.; Udler, M.S.; Leong, A.; Meigs, J.B. A decade of genetic and metabolomic contributions to type 2 diabetes risk prediction. Curr. Diabetes Rep. 2017, 17, 135. [Google Scholar] [CrossRef]
- Scott, R.A.; Scott, L.J.; Mägi, R.; Marullo, L.; Gaulton, K.J.; Kaakinen, M.; Pervjakova, N.; Pers, T.H.; Johnson, A.D.; Eicher, J.D.; et al. an expanded genome-wide association study of type 2 diabetes in Europeans. Diabetes 2017, 66, 2888–2902. [Google Scholar] [CrossRef] [PubMed]
- Ingelsson, E.; McCarthy, M.I. Human genetics of obesity and type 2 diabetes mellitus: Past, present, and future. Circ. Genom. Precis. Med. 2018, 11, e002090. [Google Scholar] [CrossRef]
- Brown, A.E.; Walker, M. Genetics of insulin resistance and the metabolic syndrome. Curr. Cardiol. Rep. 2016, 18, 1–8. [Google Scholar] [CrossRef]
- Kahn, S.E.; Hull, R.L.; Utzschneider, K.M. Mechanisms linking obesity to insulin resistance and type 2 diabetes. Nature 2006, 444, 840–846. [Google Scholar] [CrossRef] [PubMed]
- Dupuis, J.; Langenberg, C.; Prokopenko, I.; Saxena, R.; Soranzo, N.; Jackson, A.U.; Wheeler, E.; Glazer, N.L.; Bouatia-Naji, N.; Gloyn, A.L.; et al. New genetic loci implicated in fasting glucose homeostasis and their impact on type 2 diabetes risk. Nat. Genet. 2010, 42, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Fuchsberger, C.; Flannick, J.; Teslovich, T.M.; Mahajan, A.; Agarwala, V.; Gaulton, K.J.; Ma, C.; Fontanillas, P.; Moutsianas, L.; McCarthy, D.J.; et al. The genetic architecture of type 2 diabetes. Nature 2016, 536, 41–47. [Google Scholar] [CrossRef]
- Mahajan, A.; Taliun, D.; Thurner, M.; Robertson, N.R.; Torres, J.M.; Rayner, N.W.; Payne, A.J.; Steinthorsdottir, V.; Scott, R.A.; Grarup, N.; et al. Fine-mapping type 2 diabetes loci to single-variant resolution using high-density imputation and islet-specific epigenome maps. Nat. Genet. 2018, 50, 1505–1513. [Google Scholar] [CrossRef] [PubMed]
- Florez, J.C. Pharmacogenetics in type 2 diabetes: Precision medicine or discovery tool? Diabetologia 2017, 60, 800–807. [Google Scholar] [CrossRef] [PubMed]
- Zeevi, D.; Korem, T.; Zmora, N.; Israeli, D.; Rothschild, D.; Weinberger, A.; Ben-Yacov, O.; Lador, D.; Avnit-Sagi, T.; Lotan-Pompan, M.; et al. Personalized nutrition by prediction of glycemic responses. Cell 2015, 163, 1079–1094. [Google Scholar] [CrossRef]
- Wehr, J.D.; Sheath, R.G.; Kociolek, J.P. Freshwater Algae of North America. Ecology and Classification, 2nd ed.; Academia Press: San Diego, CA, USA; Elsevier: Oxford, UK, 2015; A volume in Aquatic Ecology; ISBN 9780123858771. [Google Scholar]
- Ścieszka, S.; Klewicka, E. Algae in food: A general review. Crit. Rev. Food Sci. Nutr. 2019, 59, 3538–3547. [Google Scholar] [CrossRef]
- Tiwari, B.K.; Troy, D.J. Seaweed Sustainability. Food and Non-Food Applications, 1st ed.; Academic Press: Waltham, MA, USA; Elsevier: Oxford, UK, 2015; ISBN 978-0-12-418697-2. [Google Scholar]
- Van den Hoek, C.; Mann, D.G.; Jahns, H.M. Algae: An Introduction to Phycology; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar]
- Ferdouse, F.; Løvstad-Holdt, S.; Smith, R.; Murúa, P.; Yang, Z. Food and Agriculture Organization of the United Nations. Available online: http://www.fao.org/in-action/globefish/publications/details-publication/en/c/1154074/ (accessed on 25 January 2021).
- Wells, M.L.; Potin, P.; Craigie, J.S.; Raven, J.A.; Merchant, S.S.; Helliwell, K.E.; Smith, A.G.; Camire, M.E.; Brawley, S.H. Algae as nutritional and functional food sources: Revisiting our understanding. Appl. Phycol. 2017, 29, 949–982. [Google Scholar] [CrossRef]
- EUMOFA. Observatorio Europeo del Mercado de los Productos de la Pesca y de la Acuicultura. Available online: https://www.eumofa.eu/documents/20178/136822/Analisis+de+especies.pdf (accessed on 25 January 2021).
- Tseng, C.K. Commercial cultivation. In The Biology of Seaweeds; Lobban, C.S., Wynne, M.J., Eds.; University of California Press: Berkeley, CA, USA; Blackwell Science Publications: Oxford, UK, 1981; Volume 17, pp. 680–725. [Google Scholar]
- Afonso, N.C.; Catarino, M.D.; Silva, A.M.S.; Cardoso, S.M. Brown Macroalgae as Valuable Food Ingredients. Antioxidants 2019, 8, 365. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations (FAO). The State of World Fisheries and Aquaculture 2018—Meeting the Sustainable Development Goals; FAO: Rome, Italy, 2018; ISBN 978-92-5-130562-1. [Google Scholar]
- Hafting, J.T.; Critchley, A.T.; Cornish, M.L.; Hubley, S.A.; Archibald, A.F. On-land cultivation of functional seaweed products for human usage. J. Appl. Phycol. 2012, 24, 385–392. [Google Scholar] [CrossRef]
- Stengel, D.B.; Connan, S.; Popper, Z.A. Algal chemodiversity and bioactivity: Sources of natural variability and implications for commercial application. Biotechnol. Adv. 2011, 29, 483–501. [Google Scholar] [CrossRef]
- Zheng, Y.; Ley, S.H.; Hu, F.B. Global aetiology and epidemiology of type 2 diabetes mellitus and its complications. Nat. Rev. Endocrinol. 2018, 14, 88–98. [Google Scholar] [CrossRef] [PubMed]
- Villanueva-Peñacarrillo, M.L.; Martín-Duce, A.; Ramos-Álvarez, I.; Gutiérrez-Rojas, I.; Moreno, P.; Nuche-Berenguer, B.; Acitores, A.; Sancho, V.; Valverde, I.; González, N. Characteristic of GLP-1 effects on glucose metabolism in human skeletal muscle from obese patients. Regul. Pept. 2011, 168, 39–44. [Google Scholar] [CrossRef]
- Cofrades, S.; Benedí, J.; Garcimartin, A.; Sánchez-Muniz, F.; Jimenez-Colmenero, F. A comprehensive approach to formulation of seaweed-enriched meat products: From technological development to assessment of healthy properties. Food Res. Int. 2017, 99, 1084–1094. [Google Scholar] [CrossRef] [PubMed]
- Shang, Q.; Song, G.; Zhang, M.; Shi, J.; Xu, C.; Hao, J.; Li, G.; Yu, G. Dietary fucoidan improves metabolic syndrome in association with increased Akkermansia population in the gut microbiota of high-fat diet-fed mice. J. Funct. Foods 2017, 28, 138–146. [Google Scholar] [CrossRef]
- Granado-Lorencio, F.; Hernández-Alvarez, E. Functional foods and health effects: A nutritional biochemistry perspective. Curr. Med. Chem. 2016, 23, 2929–2957. [Google Scholar] [CrossRef] [PubMed]
- Imamura, F.; Micha, R.; Wu, J.H.; de Oliveira Otto, M.C.; Otite, F.O.; Abioye, A.I.; Mozaffarian, D. Effects of saturated fat, polyunsaturated fat, monounsaturated fat, and carbohydrate on glucose-insulin homeostasis: A systematic review and meta-analysis of randomised controlled feeding trials. PLoS Med. 2016, 13, e1002087. [Google Scholar] [CrossRef]
- Shah, M.; Garg, A. The relationships between macronutrient and micronutrient intakes and type 2 diabetes mellitus in South Asians: A review. J. Diabetes Complicat. 2019, 33, 500–507. [Google Scholar] [CrossRef]
- Galbete, C.; Schwingshackl, L.; Schwedhelm, C.; Boeing, H.; Schulze, M.B. Evaluating Mediterranean diet and risk of chronic disease in cohort studies: An umbrella review of meta-analyses. Eur. J. Epidemiol. 2018, 33, 909–931. [Google Scholar] [CrossRef]
- The InterAct Consortium. Dietary fibre and incidence of type 2 diabetes in eight European countries: The EPIC-InterAct Study and a meta-analysis of prospective studies. Diabetologia 2015, 58, 1394–1408. [Google Scholar] [CrossRef] [PubMed]
- Malik, V.S.; Li, Y.; Tobias, D.K.; Pan, A.; Hu, F.B. Dietary Protein Intake and Risk of Type 2 Diabetes in US Men and Women. Am. J. Epidemiol. 2016, 183, 715–728. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Ordóñez, E.; Jiménez-Escrig, A.; Rupérez, P. Dietary fibre and physicochemical properties of several edible seaweeds from the northwestern Spanish coast. Food Res. Int. 2010, 43, 2289–2294. [Google Scholar] [CrossRef]
- Micha, R.; Shulkin, M.L.; Peñalvo, J.L.; Khatibzadeh, S.; Singh, G.M.; Rao, M.; Fahimi, S.; Powles, J.; Mozaffarian, D. Etiologic effects and optimal intakes of foods and nutrients for risk of cardiovascular diseases and diabetes: Systematic reviews and meta-analyses from the Nutrition and Chronic Diseases Expert Group (NutriCoDE). PLoS ONE 2017, 12, e0175149. [Google Scholar] [CrossRef] [PubMed]
- Azadbakht, L.; Haghighatdoost, F.; Keshteli, A.H.; Larijani, B.; Esmaillzadeh, A. Consumption of energy-dense diets in relation to metabolic syndrome and inflammatory markers in Iranian female nurses. Public Health Nutr. 2016, 20, 893–901. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Sánchez-Muniz, J.; Sanz, B. Diet and hydration in obesity prevention and treatment. An. Real Acad. Nac. Farm. 2016, 82, 106–128. [Google Scholar]
- Sánchez-Muniz, F.J.; Bocanegra, A.; Bastida, S.; Benedí, J. Algae and cardiovascular health. In Functional Ingredients from Algae for Foods and Nutraceuticals, 1st ed.; Domínguez, H., Ed.; Woodhead Publishing Limited: Cambridge, UK, 2013; pp. 369–415. [Google Scholar]
- Maeda, H. Anti-obesity and anti-diabetic activities of algae. In Functional Ingredients from Algae for Foods and Nutraceuticals, 1st ed.; Domínguez, H., Ed.; Woodhead Publishing Limited: Cambridge, UK, 2013; pp. 453–472. [Google Scholar]
- Usov, A.I.; Zelinsky, N.D. Chemical structures of algal polysaccharides. In Functional Ingredients from Algae for Foods and Nutraceuticals, 1st ed.; Domínguez, H., Ed.; Woodhead Publishing Limited: Cambridge, UK, 2013; pp. 23–86. [Google Scholar]
- Jiménez-Escrig, A.; Sánchez-Muniz, F. Dietary fibre from edible seaweeds: Chemical structure, physicochemical properties and effects on cholesterol metabolism. Nutr. Res. 2000, 20, 585–598. [Google Scholar] [CrossRef]
- Gotteland, M.; Riveros, K.; Gasaly, N.; Carcamo, C.; Magne, F.; Liabeuf, G.; Beattie, A.; Rosenfeld, S. The pros and cons of using algal polysaccharides as prebiotics. Front. Nutr. 2020, 7. [Google Scholar] [CrossRef]
- Goñi, I.; Valdivieso, L.; Gudiel-Urbano, M. Capacity of edible seaweeds to modify in vitro starch digestibility of wheat bread. Food/Nahrung 2002, 46, 18–20. [Google Scholar] [CrossRef]
- Jung, C.-H.; Choi, K.M. Impact of high-carbohydrate diet on metabolic parameters in patients with type 2 diabetes. Nutrients 2017, 9, 322. [Google Scholar] [CrossRef]
- Ojo, O.; Ojo, O.O.; Adebowale, F.; Wang, X.-H. The effect of dietary glycaemic index on glycaemia in patients with type 2 diabetes: A systematic review and meta-analysis of randomized controlled trials. Nutrients 2018, 10, 373. [Google Scholar] [CrossRef]
- Zafar, M.I.; Mills, K.E.; Zheng, J.; Regmi, A.; Hu, S.Q.; Gou, L.; Chen, L.-L. Low-glycemic index diets as an intervention for diabetes: A systematic review and meta-analysis. Am. J. Clin. Nutr. 2019, 110, 891–902. [Google Scholar] [CrossRef]
- Ojo, O.; Ojo, O.O.; Wang, X.-H.; Adegboye, A.R.A. The Effects of a low GI diet on cardiometabolic and inflammatory parameters in patients with type 2 and gestational diabetes: A systematic review and meta-analysis of randomised controlled trials. Nutrients 2019, 11, 1584. [Google Scholar] [CrossRef]
- O’Sullivan, L.; Murphy, B.; McLoughlin, P.; Duggan, P.; Lawlor, P.G.; Hughes, H.; Gardiner, G.E. Prebiotics from marine macroalgae for human and animal health applications. Mar. Drugs 2010, 8, 2038–2064. [Google Scholar] [CrossRef]
- Cherry, P.; Yadav, S.; Strain, C.R.; Allsopp, P.J.; McSorley, E.M.; Ross, R.P.; Stanton, C. Prebiotics from seaweeds: An ocean of opportunity? Mar. Drugs 2019, 17, 327. [Google Scholar] [CrossRef]
- American Diabetes Association. (5) Prevention or delay of type 2 diabetes. Diabetes Care 2015, 38, 31–32. [Google Scholar] [CrossRef]
- Kumar, V.; Sinha, A.K.; Makkar, H.P.S.; de Boeck, G.; Becker, K. Dietary roles of non-starch polysachharides in human nutrition: A review. Crit. Rev. Food Sci. Nutr. 2012, 52, 899–935. [Google Scholar] [CrossRef]
- Dvir, I.; Chayoth, R.; Sod-Moriah, U.; Shany, S.; Nyska, A.; Stark, A.H.; Madar, Z.; Arad, S.M. Soluble polysaccharide and biomass of red microalga Porphyridium sp. alter intestinal morphology and reduce serum cholesterol in rats. Br. J. Nutr. 2000, 84, 469–476. [Google Scholar] [CrossRef] [PubMed]
- Usov, A.I. Polysaccharides of the red algae. Adv. Carbohydr. Chem. Biochem. 2011, 65, 115–217. [Google Scholar] [CrossRef]
- Necas, J.; Bartosikova, L. Carrageenan: A review. Vet. Med. 2013, 58, 187–205. [Google Scholar] [CrossRef]
- Wang, X.; Yang, Z.; Xu, X.; Jiang, H.; Cai, C.; Yu, G. Odd-numbered agaro-oligosaccharides alleviate type 2 diabetes mellitus and related colonic microbiota dysbiosis in mice. Carbohydr. Polym. 2020, 240, 116261. [Google Scholar] [CrossRef]
- Gupta, S.; Abu-Ghannam, N. Bioactive potential and possible health effects of edible brown seaweeds. Trends Food Sci. Technol. 2011, 22, 315–326. [Google Scholar] [CrossRef]
- Dobrinčić, A.; Balbino, S.; Zorić, Z.; Pedisić, S.; Kovačević, D.B.; Garofulić, I.E.; Dragović-Uzelac, V. Advanced technologies for the extraction of marine brown algal polysaccharides. Mar. Drugs 2020, 18, 168. [Google Scholar] [CrossRef] [PubMed]
- Zargarzadeh, M.; Amaral, A.J.; Custódio, C.A.; Mano, J.F. Biomedical applications of laminarin. Carbohydr. Polym. 2020, 232, 115774. [Google Scholar] [CrossRef] [PubMed]
- Commission implementing regulation (EU) 2018/1023 of 23 July 2018 correcting Implementing Regulation (EU) 2017/2470 establishing the Union list of novel foods. Off. J. Eur. Union 2018, 187, 1–133.
- Wang, Y.; Xing, M.; Cao, Q.; Ji, A.; Liang, H.; Song, S. Biological activities of fucoidan and the factors mediating its therapeutic effects: A review of recent studies. Mar. Drugs 2019, 17, 183. [Google Scholar] [CrossRef]
- Guo, J.; Gao, Y.; Cao, X.; Zhang, J.; Chen, W. Cholesterol-lowing effect of taurine in HepG2 cell. Lipids Health Dis. 2017, 16, 56. [Google Scholar] [CrossRef]
- Terpstra, A.H.; Sánchez-Muniz, F.J. Time course of the development of hypercholesterolemia in rabbits fed semipurified diets containing casein or soybean protein. Atherosclerosis 1981, 39, 217–227. [Google Scholar] [CrossRef]
- Vázquez, J.; Sánchez-Muniz, F.J. Proteína de pescado y metabolismo de colesterol. Rev. Española Cienc. Tecnol. Aliment. 1994, 34, 589–608. [Google Scholar]
- Promintzer, M.; Krebs, M. Effects of dietary protein on glucose homeostasis. Curr. Opin. Clin. Nutr. Metab. Care 2006, 9, 463–468. [Google Scholar] [CrossRef]
- Shang, X.; Scott, D.; Hodge, A.M.; English, D.R.; Giles, G.G.; Ebeling, P.R.; Sanders, K.M. Dietary protein intake and risk of type 2 diabetes: Results from the Melbourne Collaborative Cohort Study and a meta-analysis of prospective studies. Am. J. Clin. Nutr. 2016, 104, 1352–1365. [Google Scholar] [CrossRef] [PubMed]
- Kumari, P. Algal lipids, fatty acids and sterols. In Functional Ingredients from Algae for Foods and Nutraceuticals, 1st ed.; Domínguez, H., Ed.; Woodhead Publishing Limited: Cambridge, UK, 2013; pp. 87–132. [Google Scholar]
- Murray, M.; Dordevic, A.L.; Ryan, L.; Bonham, M.P. An emerging trend in functional foods for the prevention of cardiovascular disease and diabetes: Marine algal polyphenols. Crit. Rev. Food Sci. Nutr. 2018, 58, 1342–1358. [Google Scholar] [CrossRef] [PubMed]
- Barkia, I.; Saari, N.; Manning, S.R. Microalgae for high-value products towards human health and nutrition. Mar. Drugs 2019, 17, 304. [Google Scholar] [CrossRef]
- Takahashi, K.; Kamada, C.; Yoshimura, H.; Okumura, R.; Iimuro, S.; Ohashi, Y.; Araki, A.; Umegaki, H.; Sakurai, T.; Yoshimura, Y.; et al. Effects of total and green vegetable intakes on glycated hemoglobin A1c and triglycerides in elderly patients with type 2 diabetes mellitus: The Japanese Elderly Intervention Trial. Geriatr. Gerontol. Int. 2012, 12, 50–58. [Google Scholar] [CrossRef]
- Telle-Hansen, V.H.; Gaundal, L.; Myhrstad, M.C. Polyunsaturated fatty acids and glycemic control in type 2 diabetes. Nutrients 2019, 11, 1067. [Google Scholar] [CrossRef]
- Ros, E.; López-Miranda, J.; Rubio, M.Á.; Babio, N.; Sala-Vila, A.; Pérez-Jiménez, F.; Escrich, E.; Bulló, M.; Solanas, M.; on behalf FESNAD. Consensus on fats and oils in the diet of Spanish adults; position paper of the Spanish Federation of Food, Nutrition and Dietetics Societies. Nutr. Hosp. 2015, 32, 435–477. [Google Scholar]
- Forouhi, N.G.; Imamura, F.; Sharp, S.J.; Koulman, A.; Schulze, M.B.; Zheng, J.; Ye, Z.; Sluijs, I.; Guevara, M.; Huerta, J.M.; et al. Association of plasma phospholipid n-3 and n-6 polyunsaturated fatty acids with type 2 diabetes: The EPIC-InterAct case-cohort study. PLoS Med. 2016, 13, e1002094. [Google Scholar] [CrossRef]
- Brown, T.J.; Brainard, J.; Song, F.; Wang, X.; Abdelhamid, A.; Hooper, L.; PUFAH Group. Omega-3, omega-6, and total dietary polyunsaturated fat for prevention and treatment of type 2 diabetes mellitus: Systematic review and meta-analysis of randomised controlled trials. BMJ 2019, 366, l4697. [Google Scholar] [CrossRef]
- Figuiredo, P.S.; Inada, A.C.; Marcelino, G.; Lopes Cardozo, C.M.; de Cássia Freitas, K.; de Cássia Avellaneda Guimarães, R.; de Castro, A.P.; do Nascimento, V.A.; Hiane, P.A. Fatty acids consumption: The role metabolic aspects involved in obesity and its associated disorders. Nutrients 2017, 9, 1158. [Google Scholar] [CrossRef]
- Wang, M.; Ma, L.-J.; Yang, Y.; Xiao, Z.; Wan, J.-B. n-3 Polyunsaturated fatty acids for the management of alcoholic liver disease: A critical review. Crit. Rev. Food Sci. Nutr. 2019, 59, S116–S129. [Google Scholar] [CrossRef]
- Salas-Salvadó, J.; Bulló, M.; Babio, N.; Martínez-González, M.Á.; Ibarrola-Jurado, N.; Basora, J.; Estruch, R.; Covas, M.I.; Corella, D.; Arós, F.; et al. Reduction in the incidence of type 2 diabetes with the mediterranean diet: Results of the PREDIMED-Reus nutrition intervention randomized trial. Diabetes Care 2010, 34, 14–19. [Google Scholar] [CrossRef]
- Gil, A. Tratado de Nutrición, 3rd ed.; Editorial Médica Panamericana: Barcelona, Spain, 2010. [Google Scholar]
- Calder, P.C. Marine omega-3 fatty acids and inflammatory processes: Effects, mechanisms and clinical relevance. Biochim. Biophys. Acta 2015, 1851, 469–484. [Google Scholar] [CrossRef]
- Cofrades, S.; Serdaroglu, M.; Jiménez-Colmenero, F. Design of healthier foods and beverages containing whole algae. In Functional Ingredients from Algae for Foods and Nutraceuticals, 1st ed.; Domínguez, H., Ed.; Woodhead Publishing Limited: Cambridge, UK, 2013; pp. 609–633. [Google Scholar]
- Masuda, T.; Yamamoto, A.; Toyohara, H. The iron content and ferritin contribution in fresh, dried, and toasted nori, Pyropia yezoensis. Biosci. Biotechnol. Biochem. 2015, 79, 74–81. [Google Scholar] [CrossRef]
- Siddiqui, K.; Bawazeer, N.; Joy, S.S. Variation in macro and trace elements in progression of type 2 diabetes. Sci. World J. 2014, 2014, 1–9. [Google Scholar] [CrossRef]
- Fernández-Cao, J.C.; Warthon-Medina, M.; Moran, V.H.; Arija, V.; Doepking, C.; Serra-Majem, L.; Lowe, N.M. zinc intake and status and risk of type 2 diabetes mellitus: A systematic review and meta-analysis. Nutrients 2019, 11, 1027. [Google Scholar] [CrossRef]
- Fukunaka, A.; Fujitani, Y. Role of zinc homeostasis in the pathogenesis of diabetes and obesity. Int. J. Mol. Sci. 2018, 19, 476. [Google Scholar] [CrossRef] [PubMed]
- Chu, A.; Foster, M.; Samman, S. Zinc status and risk of cardiovascular diseases and type 2 diabetes mellitus—A systematic review of prospective cohort studies. Nutrients 2016, 8, 707. [Google Scholar] [CrossRef] [PubMed]
- Eshak, E.S.; Iso, H.; Maruyama, K.; Muraki, I.; Tamakoshi, A. Associations between dietary intakes of iron, copper and zinc with risk of type 2 diabetes mellitus: A large population-based prospective cohort study. Clin. Nutr. 2018, 37, 667–674. [Google Scholar] [CrossRef] [PubMed]
- Vaquero, M.P.; Martínez-Suárez, M.; García-Quismondo, Á.; del Cañizo, F.J.; Sánchez-Muniz, F.J. Diabesity negatively affects transferrin saturation and iron status. The DICARIVA study. Diabetes Res. Clin. Pr. 2021, 172, 108653. [Google Scholar] [CrossRef] [PubMed]
- Ozcaliskan Ilkay, H.; Sahin, H.; Tanriverdi, F.; Samur, G. Association between magnesium status, dietary magnesium intake, and metabolic control in patients with type 2 diabetes mellitus. J. Am. Coll. Nutr. 2019, 38, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Brandão-Lima, P.N.; de Carvalho, G.B.; Santos, R.K.F.; Santos, B.D.C.; Dias-Vasconcelos, N.L.; Rocha, V.D.S.; Barbosa, K.B.F.; Pires, L.V. Intakes of zinc, potassium, calcium, and magnesium of individuals with type 2 diabetes mellitus and the relationship with glycemic control. Nutrients 2018, 10, 1948. [Google Scholar] [CrossRef] [PubMed]
- Solovyev, N.; Vanhaecke, F.; Michalke, B. Selenium and iodine in diabetes mellitus with a focus on the interplay and speciation of the elements. J. Trace Elem. Med. Biol. 2019, 56, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Farebrother, J.; Zimmermann, M.B.; Andersson, M. Excess iodine intake: Sources, assessment, and effects on thyroid function. Ann. N. Y. Acad. Sci. 2019, 1446, 44–65. [Google Scholar] [CrossRef]
- Hua, Y.; Clark, S.; Ren, J.; Sreejayan, N. Molecular mechanisms of chromium in alleviating insulin resistance. J. Nutr. Biochem. 2012, 23, 313–319. [Google Scholar] [CrossRef]
- Vincent, J.B. The biochemistry of chromium. J. Nutr. 2000, 130, 715–718. [Google Scholar] [CrossRef] [PubMed]
- Maret, W. Chromium supplementation in human health, metabolic syndrome, and diabetes. Met. Ions Life Sci. 2019, 19, 393–412. [Google Scholar] [CrossRef]
- Sánchez-Muniz, F.J.; Macho-González, A.; Garcimartín, A.; Santos-López, J.A.; Benedí, J.; Bastida, S.; González-Muñoz, M.J. The nutritional components of beer and its relationship with neurodegeneration and Alzheimer’s Disease. Nutrients 2019, 11, 1558. [Google Scholar] [CrossRef] [PubMed]
- Garcimartín, A.; López-Oliva, M.E.; Sántos-López, J.A.; García-Fernández, R.A.; Macho-González, A.; Bastida, S.; Benedí, J.; Sánchez-Muniz, F.J. Silicon alleviates nonalcoholic steatohepatitis by reducing apoptosis in aged wistar rats fed a high–saturated fat, high-cholesterol diet. J. Nutr. 2017, 147, 1104–1112. [Google Scholar] [CrossRef] [PubMed]
- Garcimartín, A.; López-Oliva, M.E.; Macho-González, A.; Bastida, S.; Benedí, J.; Sánchez-Muniz, F.J. Hypoglycaemic and hypotriglyceridaemic postprandial properties of organic silicon. J. Funct. Foods 2017, 29, 290–294. [Google Scholar] [CrossRef]
- Santos-López, J.A.; Garcimartín, A.; Merino, P.; López-Oliva, M.E.; Bastida, S.; Benedí, J.; Sánchez-Muniz, F.J. Effects of silicon vs. hydroxytyrosol-enriched restructured pork on liver oxidation status of aged rats fed high-saturated/high-cholesterol diets. PLoS ONE 2016, 11, e0147469. [Google Scholar] [CrossRef]
- Bocanegra, A.; Beneduí, J.; Nchez-Muniz, F.J.S. Differential effects of konbu and nori seaweed dietary supplementation on liver glutathione status in normo- and hypercholesterolaemic growing rats. Br. J. Nutr. 2006, 95, 696–702. [Google Scholar] [CrossRef]
- Kanazawa, S.; Herbert, V. Noncobalamin vitamin B12 analogues in human red cells, liver, and brain. Am. J. Clin. Nutr. 1983, 37, 774–777. [Google Scholar] [CrossRef]
- Ito, M.; Koba, K.; Hikihara, R.; Ishimaru, M.; Shibata, T.; Hatate, H.; Tanaka, R. Analysis of functional components and radical scavenging activity of 21 algae species collected from the Japanese coast. Food Chem. 2018, 255, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Dakshinamurti, K. Vitamins and their derivatives in the prevention and treatment of metabolic syndrome diseases (diabetes). Can. J. Physiol. Pharmacol. 2015, 93, 355–362. [Google Scholar] [CrossRef]
- Valdes-Ramos, R.; Guadarrama-López, A.L.; Martínez-Carrillo, B.E.; Benítez-Arciniega, A.D. Vitamins and type 2 diabetes mellitus. Endocr. Metab. Immune Disord. Drug Targets 2015, 15, 54–63. [Google Scholar] [CrossRef]
- Wijekoon, E.; Brosnan, M.; Brosnan, J. Homocysteine metabolism in diabetes. Biochem. Soc. Trans. 2007, 35, 1175–1179. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Ahn, C.W.; Fang, S.; Lee, H.S.; Park, J.S. Association between metformin dose and vitamin B12 deficiency in patients with type 2 diabetes. Medicines 2019, 98, e17918. [Google Scholar] [CrossRef] [PubMed]
- Mason, S.A.; Rasmussen, B.; van Loon, L.J.; Salmon, J.; Wadley, G.D. Ascorbic acid supplementation improves postprandial glycaemic control and blood pressure in individuals with type 2 diabetes: Findings of a randomized cross-over trial. Diabetes Obes. Metab. 2019, 21, 674–682. [Google Scholar] [CrossRef]
- Wilson, R.; Willis, J.; Gearry, R.; Skidmore, P.; Fleming, E.; Frampton, C.; Carr, A. Inadequate vitamin C status in prediabetes and type 2 diabetes mellitus: Associations with glycaemic control, obesity, and smoking. Nutrients 2017, 9, 997. [Google Scholar] [CrossRef]
- Pazdro, R.; Burgess, J.R. The role of vitamin E and oxidative stress in diabetes complications. Mech. Ageing Dev. 2010, 131, 276–286. [Google Scholar] [CrossRef]
- Faure, P. Protective effects of antioxidant micronutrients (vitamin E, Zinc and Selenium) in type 2 diabetes mellitus. Clin. Chem. Lab. Med. 2003, 41, 995–998. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Garach, A.; García-Fontana, B.; Muñoz-Torres, M. Vitamin D status, calcium intake and risk of developing type 2 diabetes: An unresolved issue. Nutrients 2019, 11, 642. [Google Scholar] [CrossRef]
- Lips, P.; Eekhoff, M.; van Schoor, N.; Oosterwerff, M.; de Jongh, R.; Krul-Poel, Y.; Simsek, S. Vitamin D and type 2 diabetes. J. Steroid Biochem. Mol. Biol. 2017, 173, 280–285. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chen, J.P.; Duan, L.; Li, S. Effect of vitamin K2 on type 2 diabetes mellitus: A review. Diabetes Res. Clin. Pr. 2018, 136, 39–51. [Google Scholar] [CrossRef]
- Guasch-Ferré, M.; Merino, J.; Sun, Q.; Fitó, M.; Salas-Salvadó, J. Dietary polyphenols, Mediterranean diet, prediabetes, and type 2 diabetes: A narrative review of the evidence. Oxidative Med. Cell. Longev. 2017, 2017, 1–16. [Google Scholar] [CrossRef]
- Hussain, T.; Tan, B.; Murtaza, G.; Liu, G.; Rahu, N.; Saleem Kalhoro, M.; Hussain-Kalhoro, D.; Adebowale, T.O.; Usman Mazhar, M.; Rehman, Z.U.; et al. Flavonoids and type 2 diabetes: Evidence of efficacy in clinical and animal studies and delivery strategies to enhance their therapeutic efficacy. Pharmacol. Res. 2020, 152, 104629. [Google Scholar] [CrossRef] [PubMed]
- Putta, S.; Yarla, N.S.; Kumar K, E.; Lakkappa, D.B.; Kamal, M.A.; Scotti, L.; Scotti, M.T.; Ashraf, G.M.; Rao, B.S.B.; Reddy, G.V.; et al. Preventive and therapeutic potentials of anthocyanins in diabetes and associated complications. Curr. Med. Chem. 2019, 25, 5347–5371. [Google Scholar] [CrossRef]
- Serrano, J.; Puupponen-Pimiä, R.; Dauer, A.; Aura, A.-M.; Calixto, F.D.S. Tannins: Current knowledge of food sources, intake, bioavailability and biological effects. Mol. Nutr. Food Res. 2009, 53, S310–S329. [Google Scholar] [CrossRef] [PubMed]
- Scoggan, K.A.; Gruber, H.; Chen, Q.; Plouffe, L.J.; Lefebvre, J.M.; Wang, B.; Bertinato, J.; L’Abbé, M.R.; Hayward, S.; Ratnayake, W.N. Increased incorporation of dietary plant sterols and cholesterol correlates with decreased expression of hepatic and intestinal Abcg5 and Abcg8 in diabetic BB rats. J. Nutr. Biochem. 2009, 20, 177–186. [Google Scholar] [CrossRef]
- Sluijs, I.; Cadier, E.; Beulens, J.; van der A, D.L.; Spijkerman, A.; van der Schouw, Y. Dietary intake of carotenoids and risk of type 2 diabetes. Nutr. Metab. Cardiovasc. Dis. 2015, 25, 376–381. [Google Scholar] [CrossRef]
- She, C.; Shang, F.; Zhou, K.; Liu, N. Serum carotenoids and risks of diabetes and diabetic retinopathy in a Chinese population sample. Curr. Mol. Med. 2017, 17, 287–297. [Google Scholar] [CrossRef]
- Gomez-Zavaglia, A.; Lage, M.A.P.; Jimenez-Lopez, C.; Mejuto, J.C.; Simal-Gandara, J. El potencial de las algas marinas como fuente de ingredientes funcionales de valor prebiótico y antioxidante. Antioxidantes 2019, 8, 406. [Google Scholar] [CrossRef] [PubMed]
- Cofrades, S.; López-López, I.; Bravo, L.; Ruiz-Capillas, C.; Bastida, S.; Larrea, M.T.; Jiménez-Colmenero, F. Nutritional and antioxidant properties of different brown and red Spanish edible seaweeds. Food Sci. Technol. Int. 2010, 16, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.W.; Woo, H.D.; Cho, M.J.; Park, J.K.; Kim, S.S. Identification of dietary patterns associated with incidence of hyperglycemia in middle-aged and older Korean adults. Nutrients 2019, 11, 1801. [Google Scholar] [CrossRef]
- Lee, H.J.; Kim, H.C.; Vitek, L.; Nam, C.M. Algae consumption and risk of type 2 diabetes: Korean National Health and Nutrition Examination Survey in 2005. J. Nutr. Sci. Vitaminol. 2010, 56, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Oba, S.; Nanri, A.; Kurotani, K.; Goto, A.; Kato, M.; Mizoue, T.; Noda, M.; Inoue, M.; Tsugane, S.; Japan Public Health Center-based Prospective Study Group. Dietary glycemic index, glycemic load and incidence of type 2 diabetes in Japanese men and women: The Japan public health center-based prospective study. Nutr. J. 2013, 12, 165. [Google Scholar] [CrossRef]
- Paradis, M.-E.; Couture, P.; Lamarche, B. A randomised crossover placebo-controlled trial investigating the effect of brown seaweed (Ascophyllum nodosum and Fucus vesiculosus) on postchallenge plasma glucose and insulin levels in men and women. Appl. Physiol. Nutr. Metab. 2011, 36, 913–919. [Google Scholar] [CrossRef] [PubMed]
- Yoshinaga, K.; Mitamura, R. Effects of Undaria pinnatifida (Wakame) on postprandial glycemia and insulin levels in humans: A randomized crossover trial. Plant Foods Hum. Nutr. 2019, 74, 461–467. [Google Scholar] [CrossRef]
- Jensen, M.G.; Pedersen, C.; Kristensen, M.; Frost, G.; Astrup, A. Review: Efficacy of alginate supplementation in relation to appetite regulation and metabolic risk factors: Evidence from animal and human studies. Obes. Rev. 2013, 14, 129–144. [Google Scholar] [CrossRef]
- Tanemura, Y.; Yamanaka-Okumura, H.; Sakuma, M.; Nii, Y.; Taketani, Y.; Takeda, E. Effects of the intake of Undaria pinnatifida (Wakame) and its sporophylls (Mekabu) on postprandial glucose and insulin metabolism. J. Med Investig. 2014, 61, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.-T.; Rioux, L.-E.; Turgeon, S.L. Alpha-amylase and alpha-glucosidase inhibition is differentially modulated by fucoidan obtained from Fucus vesiculosus and Ascophyllum nodosum. Phytochemistry 2014, 98, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Murray, M.; Dordevic, A.L.; Bonham, M.P.; Ryan, L. Do marine algal polyphenols have antidiabetic, antihyperlipidemic or anti-inflammatory effects in humans? A systematic review. Crit. Rev. Food Sci. Nutr. 2017, 58, 2039–2054. [Google Scholar] [CrossRef]
- Seung-Hong, L.; You-Jin, J. Anti-diabetic effects of brown algae derived phlorotannins, marine polyphenols through diverse mechanisms. Fitoterapia 2013, 86, 129–136. [Google Scholar] [CrossRef]
- Hall, A.; Fairclough, A.; Mahadevan, K.; Paxman, J. Ascophyllum nodosum enriched bread reduces subsequent energy intake with no effect on post-prandial glucose and cholesterol in healthy, overweight males. A pilot study. Appetite 2012, 58, 379–386. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, S.; O-Sullivan, I.; Katyal, S.; Unterman, T.; Tobacman, J.K. Exposure to the common food additive carrageenan leads to glucose intolerance, insulin resistance and inhibition of insulin signalling in HepG2 cells and C57BL/6J mice. Diabetologia 2011, 55, 194–203. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, S.; Liu, H.; Zhang, Z.; Jam, M.; Dudeja, P.K.; Michel, G.; Linhardt, R.J.; Tobacman, J.K. Carrageenan-induced innate immune response is modified by enzymes that hydrolyze distinct galactosidic bonds. J. Nutr. Biochem. 2010, 21, 906–913. [Google Scholar] [CrossRef]
- Du Preez, R.; Paul, N.; Mouatt, P.; Majzoub, M.E.; Thomas, T.; Panchal, S.K.; Brown, L. Carrageenans from the red seaweed Sarconema filiforme attenuate symptoms of diet-induced metabolic syndrome in rats. Mar. Drugs 2020, 18, 97. [Google Scholar] [CrossRef]
- Liu, J.; Kandasamy, S.; Zhang, J.; Kirby, C.W.; Karakach, T.K.; Hafting, J.; Critchley, A.T.; Evans, F.; Prithiviraj, B. Prebiotic effects of diet supplemented with the cultivated red seaweed Chondrus crispus or with fructo-oligo-saccharide on host immunity, colonic microbiota and gut microbial metabolites. BMC Complement. Altern. Med. 2015, 15, 1–12. [Google Scholar] [CrossRef]
- Sanaka, M.; Yamamoto, T.; Anjiki, H.; Nagasawa, K.; Kuyama, Y. Effects of agar and pectin on gastric emptying and post-prandial glycaemic profiles in healthy human volunteers. Clin. Exp. Pharmacol. Physiol. 2007, 34, 1151–1155. [Google Scholar] [CrossRef]
- Maeda, H.; Yamamoto, R.; Hirao, K.; Tochikubo, O. Effects of agar (kanten) diet on obese patients with impaired glucose tolerance and type 2 diabetes. Diabetes Obes. Metab. 2004, 7, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Wang, L.; Zhu, C.; Wu, J.; Yuan, Y.; Yu, L.; Xu, Y.; Xu, J.; Wang, T.; Liao, Z.; et al. Laminarin counteracts diet-induced obesity associated with glucagon-like peptide-1 secretion. Oncotarget 2017, 8, 99470–99481. [Google Scholar] [CrossRef]
- Kimura, Y.; Watanabe, K.; Okuda, H. Effects of soluble sodium alginate on cholesterol excretion and glucose tolerance in rats. J. Ethnopharmacol. 1996, 54, 47–54. [Google Scholar] [CrossRef]
- De La Garza, A.R.M.; Tapia-Salazar, M.; Maldonado-Muñiz, M.; de la Rosa-Millán, J.; Gutiérrez-Uribe, J.A.; Santos-Zea, L.; Barba-Dávila, B.A.; Ricque-Marie, D.; Cruz-Suárez, L.E. Nutraceutical potential of five Mexican brown seaweeds. BioMed Res. Int. 2019, 2019, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Torsdottir, I.; Alpsten, M.; Holm, G.; Sandberg, A.-S.; Tölli, J. A small dose of soluble alginate-fiber affects postprandial glycemia and gastric emptying in humans with diabetes. J. Nutr. 1991, 121, 795–799. [Google Scholar] [CrossRef] [PubMed]
- Idota, Y.; Kato, T.; Shiragami, K.; Koike, M.; Yokoyama, A.; Takahashi, H.; Yano, K.; Ogihara, T. Mechanism of suppression of blood glucose level by calcium alginate in rats. Biol. Pharm. Bull. 2018, 41, 1362–1366. [Google Scholar] [CrossRef]
- Wan-Loy, C.; Siew-Moi, P. Marine algae as a potential source for anti-obesity agents. Mar. Drugs 2016, 14, 222. [Google Scholar] [CrossRef]
- Odunsi, S.T.; Vázquez-Roque, M.I.; Camilleri, M.; Papathanasopoulos, A.; Clark, M.M.; Wodrich, L.; Lempke, M.; McKinzie, S.; Ryks, M.; Burton, D.; et al. Effect of alginate on satiation, appetite, gastric function, and selected gut satiety hormones in overweight and obesity. Obesity 2010, 18, 1579–1584. [Google Scholar] [CrossRef]
- Birketvedt, G.S.; Shimshi, M.; Erling, T.; Florholmen, J. Experiences with three different fiber supplements in weight reduction. Med Sci. Monit. 2005, 11, PI5–PI8. [Google Scholar]
- Vaugelade, P.; Hoebler, C.; Bernard, F.; Guillon, F.; Lahaye, M.; Duee, P.-H.; Darcy-Vrillon, B. Non-starch polysaccharides extracted from seaweed can modulate intestinal absorption of glucose and insulin response in the pig. Reprod. Nutr. Dev. 2000, 40, 33–47. [Google Scholar] [CrossRef]
- Zayed, A.; Ulber, R. Fucoidans: Downstream Processes and Recent Applications. Mar. drugs. 2020, 18, 170. [Google Scholar] [CrossRef]
- Luthuli, S.; Wu, S.; Cheng, Y.; Zheng, X.; Wu, M.; Tong, H. Therapeutic effects of fucoidan: A review on recent studies. Mar. Drugs 2019, 17, 487. [Google Scholar] [CrossRef]
- Pradhan, B.; Patra, S.; Nayak, R.; Behera, C.; Dash, S.R.; Nayak, S.; Sahu, B.B.; Bhutia, S.K.; Jena, M. Multifunctional role of fucoidan, sulfated polysaccharides in human health and disease: A journey under the sea in pursuit of potent therapeutic agents. Int. J. Biol. Macromol. 2020, 164, 4263–4278. [Google Scholar] [CrossRef] [PubMed]
- Willcox, D.C.; Scapagnini, G.; Willcox, B.J. Healthy aging diets other than the Mediterranean: A focus on the Okinawan diet. Mech. Ageing Dev. 2014, 136–137, 148–162. [Google Scholar] [CrossRef] [PubMed]
- Mabate, B.; Daub, C.D.; Malgas, S.; Edkins, A.L.; Pletschke, B.I. Fucoidan structure and its impact on glucose metabolism: Implications for diabetes and cancer therapy. Mar. Drugs 2021, 19, 30. [Google Scholar] [CrossRef] [PubMed]
- Shan, X.; Liu, X.; Hao, J.; Cai, C.; Fan, F.; Dun, Y.; Zhao, X.; Liu, X.; Li, C.; Yu, G. In vitro and in vivo hypoglycemic effects of brown algal fucoidans. Int. J. Biol. Macromol. 2016, 82, 249–255. [Google Scholar] [CrossRef]
- Kim, K.-J.; Yoon, K.-Y.; Lee, B.-Y. Fucoidan regulate blood glucose homeostasis in C57BL/KSJ m+/+db and C57BL/KSJ db/db mice. Fitoterapia 2012, 83, 1105–1109. [Google Scholar] [CrossRef]
- Apostolidis, E.; Lee, C.M. In vitro potential of ascophyllum nodosum phenolic antioxidant-mediated α-glucosidase and α-amylase inhibition. J. Food Sci. 2010, 75, H97–H102. [Google Scholar] [CrossRef]
- Lordan, S.; Smyth, T.J.; Soler-Vila, A.; Stanton, C.; Ross, R.P. The α-amylase and α-glucosidase inhibitory effects of Irish seaweed extracts. Food Chem. 2013, 141, 2170–2176. [Google Scholar] [CrossRef]
- Nwosu, F.; Morris, J.; Lund, V.A.; Stewart, D.; Ross, H.A.; McDougall, G.J. Anti-proliferative and potential anti-diabetic effects of phenolic-rich extracts from edible marine algae. Food Chem. 2011, 126, 1006–1012. [Google Scholar] [CrossRef]
- Sim, S.-Y.; Shin, Y.-E.; Kim, H.-K. Fucoidan from Undaria pinnatifida has anti-diabetic effects by stimulation of glucose uptake and reduction of basal lipolysis in 3T3-L1 adipocytes. Nutr. Res. 2019, 65, 54–62. [Google Scholar] [CrossRef]
- Yu, W.-C.; Chen, Y.-L.; Hwang, P.-A.; Chen, T.-H.; Chou, T.-C. Fucoidan ameliorates pancreatic β-cell death and impaired insulin synthesis in streptozotocin-treated β cells and mice via a Sirt-1-dependent manner. Mol. Nutr. Food Res. 2017, 61, 1700136. [Google Scholar] [CrossRef] [PubMed]
- Song, W.; Wang, Z.; Zhang, X.; Li, Y. Ethanol extract from ulva prolifera prevents high-fat diet-induced insulin resistance, oxidative stress, and inflammation response in mice. BioMed Res. Int. 2018, 2018, 1–9. [Google Scholar] [CrossRef]
- Taboada, C.; Millã¡n, R.; Míguez, I. Composition, nutritional aspects and effect on serum parameters of marine algaeUlva rigida. J. Sci. Food Agric. 2009, 90, 445–449. [Google Scholar] [CrossRef]
- Xu, H.-L.; Kitajima, C.; Ito, H.; Miyazaki, T.; Baba, M.; Okuyama, T.; Okada, Y. Antidiabetic effect of polyphenols from brown alga Ecklonia kuromein genetically diabetic KK-Aymice. Pharm. Biol. 2011, 50, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Iwai, K. Antidiabetic and Antioxidant Effects of Polyphenols in Brown Alga Ecklonia stolonifera in Genetically Diabetic KK-Ay Mice. Plant Foods Hum. Nutr. 2008, 63, 163–169. [Google Scholar] [CrossRef]
- Roy, M.-C.; Anguenot, R.; Fillion, C.; Beaulieu, M.; Bérubé, J.; Richard, D. Effect of a commercially-available algal phlorotannins extract on digestive enzymes and carbohydrate absorption in vivo. Food Res. Int. 2011, 44, 3026–3029. [Google Scholar] [CrossRef]
- Murray, M.; Dordevic, A.L.; Ryan, L.; Bonham, M.P. The impact of a single dose of a polyphenol-rich seaweed extract on postprandial glycaemic control in healthy adults: A randomised cross-over trial. Nutrients 2018, 10, 270. [Google Scholar] [CrossRef] [PubMed]
- Motshakeri, M.; Ebrahimi, M.; Goh, Y.M.; Matanjun, P.; Mohamed, S. Sargassum polycystumreduces hyperglycaemia, dyslipidaemia and oxidative stress via increasing insulin sensitivity in a rat model of type 2 diabetes. J. Sci. Food Agric. 2012, 93, 1772–1778. [Google Scholar] [CrossRef] [PubMed]
- Maeda, H.; Hosokawa, M.; Sashima, T.; Murakami-Funayama, K.; Miyashita, K. Anti-obesity and anti-diabetic effects of fucoxanthin on diet-induced obesity conditions in a murine model. Mol. Med. Rep. 2009, 2, 897–902. [Google Scholar] [CrossRef]
- Jung, H.A.; Bhakta, H.K.; Min, B.-S.; Choi, J.S. Fucosterol activates the insulin signaling pathway in insulin resistant HepG2 cells via inhibiting PTP1B. Arch. Pharmacal Res. 2016, 39, 1454–1464. [Google Scholar] [CrossRef]
- Jung, H.A.; Islam, N.; Lee, C.M.; Oh, S.H.; Lee, S.; Jung, J.H.; Choi, J.S. Kinetics and molecular docking studies of an anti-diabetic complication inhibitor fucosterol from edible brown algae Eisenia bicyclis and Ecklonia stolonifera. Chem. Biol. Interact. 2013, 206, 55–62. [Google Scholar] [CrossRef]
- González-Torres, L.; Matos, C.; Vázquez-Velasco, M.; Santos-López, J.A.; Sánchez-Martínez, I.; García–Fernández, C.; Bastida, S.; Benedí, J.; Sánchez-Muniz, F.J. Glucomannan- and glucomannan plus spirulina-enriched pork affect liver fatty acid profile, LDL receptor expression and antioxidant status in Zucker fa/fa rats fed atherogenic diets. Food Nutr. Res. 2016, 61, 1264710–1264714. [Google Scholar] [CrossRef] [PubMed]
- Vázquez-Velasco, M.; González-Torres, L.; Méndez, M.T.; Bastida, S.; Benedí, J.; González-Muñoz, M.J.; Sánchez-Muniz, F.J. Glucomannan and glucomannan plus spirulina-enriched squid-surimi added to high saturated diet affect glycemia, plasma and adipose leptin and adiponectin levels in growing fa/fa rats. Nutr. Hosp. 2015, 32, 2718–2724. [Google Scholar]
- Nesto, R.W. Beyond low-density lipoprotein: Addressing the atherogenic lipid triad in type 2 diabetes mellitus and the metabolic syndrome. Am. J. Cardiovasc. Drugs 2005, 5, 379–387. [Google Scholar] [CrossRef] [PubMed]
- Ormazabal, V.; Nair, S.; Elfeky, O.; Aguayo, C.; Salomon, C.; Zuñiga, F.A. Association between insulin resistance and the development of cardiovascular disease. Cardiovasc. Diabetol. 2018, 17, 122. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Muniz, F.J. IV y V Curso Avanzado en Obesidad y Síndrome Metabólico. Monografía 46; Instituto de España; Real Academia Nacional de Farmacia: Madrid, Spain, 2018. [Google Scholar]
- Bocanegra, A.; Bastida, S.; Benedí, J.; Nus, M.; Sánchez-Montero, J.M.; Sánchez-Muniz, F.J. Effect of seaweed and cholesterol-enriched diets on postprandial lipoproteinaemia in rats. Br. J. Nutr. 2009, 102, 1728–1739. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Muniz, F.J. Dietary fibre and cardiovascular health. Nutr. Hosp. 2012, 27, 31–45. [Google Scholar] [PubMed]
- González-Torres, L.; Vázquez-Velasco, M.; Olivero-David, R.; Bastida, S.; Benedi, J.; González, R.R.; González-Muñoz, M.J.; Sánchez-Muniz, F.J. Glucomannan and glucomannan plus spirulina added to pork significantly block dietary cholesterol effects on lipoproteinemia, arylesterase activity, and CYP7A1 expression in Zucker fa/fa rats. J. Physiol. Biochem. 2015, 71, 773–784. [Google Scholar] [CrossRef]
- Vázquez-Velasco, M.; González-Torres, L.; Olivero-David, R.; Bastida, S.; Benedi, J.; Sánchez-Reus, M.I.; González-Muñoz, M.J.; Sánchez-Muniz, F.J. Lipoproteinemia and arylesterase activity in Zucker Fa/Fa rats fed glucomannan/spirulina-enriched squid-surimi. Eur. J. Lipid Sci. 2013, 115, 1274–1283. [Google Scholar] [CrossRef]
- Ježek, P. Possible physiological roles of mitochondrial uncoupling proteins—UCPn. Int. J. Biochem. Cell Biol. 2002, 34, 1190–1206. [Google Scholar] [CrossRef]
- Ye, J. Mechanisms of insulin resistance in obesity. Front. Med. 2013, 7, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Backes, J.; Anzalone, D.; Hilleman, D.; Catini, J. The clinical relevance of omega-3 fatty acids in the management of hypertriglyceridemia. Lipids Health Dis. 2016, 15, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Shahidi, F.; Ambigaipalan, P. Omega-3 polyunsaturated fatty acids and their health benefits. Annu. Rev. Food Sci. Technol. 2018, 9, 345–381. [Google Scholar] [CrossRef] [PubMed]
- Abdelhamid, A.S.; Brown, T.J.; Brainard, J.S.; Biswas, P.; Thorpe, G.C.; Moore, H.J.; Deane, K.H.; AlAbdulghafoor, F.K.; Summerbell, C.D.; Worthington, H.V.; et al. Omega-3 fatty acids for the primary and secondary prevention of cardiovascular disease. Cochrane Database Syst. Rev. 2018, 7, CD003177. [Google Scholar] [CrossRef]
- Mišurcová, L.; Ambrožová, J.; Samek, D. Seaweed lipids as nutraceuticals. Adv. Food Nutr. Res. 2011, 64, 339–355. [Google Scholar] [CrossRef]
- Bermano, G.; Stoyanova, T.; Hennequart, F.; Wainwright, C.L. Seaweed-derived bioactives as potential energy regulators in obesity and type 2 diabetes. Adv. Pharmacol. 2020, 87, 205–256. [Google Scholar] [CrossRef]
- Lopez-Santamarina, A.; Miranda, J.M.; Mondragon, A.D.C.; Lamas, A.; Cardelle-Cobas, A.; Franco, C.M.; Cepeda, A. Potential use of marine seaweeds as prebiotics: A review. Molecules 2020, 25, 1004. [Google Scholar] [CrossRef]
- De Jesus Raposo, M.F.; de Morais, A.M.M.B.; de Morais, R.M.S.C. Emergent sources of prebiotics: Seaweeds and microalgae. Mar. Drugs 2016, 14, 27. [Google Scholar] [CrossRef]
- Lin, G.; Liu, X.; Yan, X.; Liu, D.; Yang, C.; Liu, B.; Huang, Y.; Zhao, C. Role of green macroalgae enteromorpha prolifera polyphenols in the modulation of gene expression and intestinal microflora profiles in type 2 diabetic mice. Int. J. Mol. Sci. 2018, 20, 25. [Google Scholar] [CrossRef]
- Yan, X.; Yang, C.; Lin, G.; Chen, Y.; Miao, S.; Liu, B.; Zhao, C. Antidiabetic potential of green seaweed enteromorpha prolifera flavonoids regulating insulin signaling pathway and gut microbiota in type 2 diabetic mice. J. Food Sci. 2019, 84, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Keleszade, E.; Patterson, M.; Trangmar, S.; Guinan, K.; Costabile, A. Clinical efficacy of brown seaweeds Ascophyllum nodosum and Fucus vesiculosus in the prevention or delay progression of the metabolic syndrome: A review of clinical trials. Molecules 2021, 26, 714. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Sibusiso, L.; Hou, L.; Jiang, H.; Chen, P.; Zhang, X.; Wu, M.; Tong, H. Sargassum fusiforme fucoidan modifies the gut microbiota during alleviation of streptozotocin-induced hyperglycemia in mice. Int. J. Biol. Macromol. 2019, 131, 1162–1170. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Zheng, Y.; Zhou, J.; Geng, Y.; Zou, P.; Li, Y.; Zhang, C. Polyphenol-rich extracts from brown macroalgae lessonia trabeculate attenuate hyperglycemia and modulate gut microbiota in high-fat diet and streptozotocin-induced diabetic rats. J. Agric. Food Chem. 2019, 67, 12472–12480. [Google Scholar] [CrossRef] [PubMed]
- Erpel, F.; Mateos, R.; Pérez-Jiménez, J.; Pérez-Correa, J.R. Phlorotannins: From isolation and structural characterization, to the evaluation of their antidiabetic and anticancer potential. Food Res. Int. 2020, 137, 109589. [Google Scholar] [CrossRef]
- Koh, A.; de Vadder, F.; Kovatcheva-Datchary, P.; Bäckhed, F. From dietary fiber to host physiology: Short-chain fatty acids as key bacterial metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef]
- Pirian, K.; Moein, S.; Sohrabipour, J.; Rabiei, R.; Blomster, J. Antidiabetic and antioxidant activities of brown and red macroalgae from the Persian Gulf. J. Appl. Phycol. 2017, 29, 3151–3159. [Google Scholar] [CrossRef]
- Nguyen, T.H.; Nguyen, T.H.; Nguyen, V.M.; Nguyen, T.L.P.; Tran, T.V.A.; Do Anh Duy, K.S.M. Antidiabetic and antioxidant activities of red seaweed Laurencia dendroidea. Asian Pac. J. Trop. Biomed. 2019, 9, 501–509. [Google Scholar] [CrossRef]
- Alves, M.; Barreto, F.; de Vasconcelos, M.A.; Nascimento Neto, L.; Carneiro, R.F.; da Silva, L.T.; Nagano, C.S.; Sampaio, A.H.; Teixeira, E.H. Antihyperglycemic and antioxidant activities of a lectin from the marine red algae, Bryothamnion seaforthii, in rats with streptozotocin-induced diabetes. Int. J. Biol. Macromol. 2020, 158, 773–780. [Google Scholar] [CrossRef] [PubMed]
- Vuppalapati, L.; Velayudam, R.; Ahamed, K.N.; Cherukuri, S.; Kesavan, B.R. The protective effect of dietary flavonoid fraction from Acanthophora spicifera on streptozotocin induced oxidative stress in diabetic rats. Food Sci. Hum. Wellness 2016, 5, 57–64. [Google Scholar] [CrossRef]
- Lee, S.-H.; Kang, S.-M.; Ko, S.-C.; Kang, M.-C.; Jeon, Y.-J. Octaphlorethol A, a novel phenolic compound isolated from Ishige foliacea, protects against streptozotocin-induced pancreatic β cell damage by reducing oxidative stress and apoptosis. Food Chem. Toxicol. 2013, 59, 643–649. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-H.; Park, M.-H.; Park, S.-J.; Kim, J.; Kim, Y.-T.; Oh, M.-C.; Jeong, Y.; Kim, M.; Han, J.-S.; Jeon, Y.-J. Bioactive compounds extracted from ecklonia cava by using enzymatic hydrolysis protects high glucose-induced damage in ins-1 pancreatic β-cells. Appl. Biochem. Biotechnol. 2012, 167, 1973–1985. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.-C.; Wijesinghe, W.; Lee, S.-H.; Kang, S.-M.; Ko, S.-C.; Yang, X.; Kang, N.; Jeon, B.-T.; Kim, J.; Lee, D.-H.; et al. Dieckol isolated from brown seaweed Ecklonia cava attenuates type II diabetes in db/db mouse model. Food Chem. Toxicol. 2013, 53, 294–298. [Google Scholar] [CrossRef] [PubMed]
- Gheda, S.; Naby, M.A.; Mohamed, T.; Pereira, L.; Khamis, A. Antidiabetic and antioxidant activity of phlorotannins extracted from the brown seaweed Cystoseira compressa in streptozotocin-induced diabetic rats. Environ. Sci. Pollut. Res. Int. 2021, 1–16. [Google Scholar] [CrossRef]
- Shannon, E.; Abu-Ghannam, N. Seaweeds as nutraceuticals for health and nutrition. Phycologia 2019, 58, 563–577. [Google Scholar] [CrossRef]
- Jayasri, M.A.; Unnikrishnan, P.S.; Suthindhiran, K. Alpha-amylase inhibition and antioxidant activity of marine green algae and its possible role in diabetes management. Pharmacogn. Mag. 2015, 11, 511–S515. [Google Scholar] [CrossRef] [PubMed]
- Mesbahzadeh, B.; Rajaei, S.A.; Tarahomi, P.; Seyedinia, S.A.; Rahmani, M.; Rezamohamadi, F.; Kakar, M.A.; Moradi-Kor, N. Beneficial effects of Spirogyra Neglecta Extract on antioxidant and anti-inflammatory factors in streptozotocin-induced diabetic rats. Biomol. Concepts 2018, 9, 184–189. [Google Scholar] [CrossRef]
- Kim, M.S.; Kim, J.Y.; Choi, W.H.; Lee, S.S. Effects of seaweed supplementation on blood glucose concentration, lipid profile, and antioxidant enzyme activities in patients with type 2 diabetes mellitus. Nutr. Res. Pr. 2008, 2, 62–67. [Google Scholar] [CrossRef]
- Kim, M.Y.; Cheong, S.H.; Lee, J.H.; Kim, M.J.; Sok, D.-E.; Kim, M.R. spirulina improves antioxidant status by reducing oxidative stress in rabbits fed a high-cholesterol diet. J. Med. Food 2010, 13, 420–426. [Google Scholar] [CrossRef] [PubMed]
- Riss, J.; Décordé, K.; Sutra, T.; Delage, M.; Baccou, J.-C.; Jouy, N.; Brune, J.-P.; Oréal, H.; Cristol, J.-P.; Rouanet, J.-M. Phycobiliprotein C-Phycocyanin from Spirulina platensis is powerfully responsible for reducing oxidative stress and NADPH oxidase expression induced by an atherogenic diet in hamsters. J. Agric. Food Chem. 2007, 55, 7962–7967. [Google Scholar] [CrossRef]
- Gargouri, M.; Hamed, H.; Akrouti, A.; Dauvergne, X.; Magné, C.; el Feki, A. Effects ofSpirulina platensison lipid peroxidation, antioxidant defenses, and tissue damage in kidney of alloxan-induced diabetic rats. Appl. Physiol. Nutr. Metab. 2018, 43, 345–354. [Google Scholar] [CrossRef]
- Gargouri, M.; Magné, C.; el Feki, A. Hyperglycemia, oxidative stress, liver damage and dysfunction in alloxan-induced diabetic rat are prevented by Spirulina supplementation. Nutr. Res. 2016, 36, 1255–1268. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.H.; Park, J.-E.; Choi, Y.-J.; Huh, K.-B.; Kim, W.-Y. A randomized study to establish the effects of spirulina in type 2 diabetes mellitus patients. Nutr. Res. Pract. 2008, 2, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Ku, C.S.; Yang, Y.; Park, Y.; Lee, J. Health benefits of blue-green algae: Prevention of cardiovascular disease and nonalcoholic fatty liver disease. J. Med. Food 2013, 16, 103–111. [Google Scholar] [CrossRef]
- Balboa, E.M.; Conde, E.; Moure, A.; Falqué, E.; Domínguez, H. In vitro antioxidant properties of crude extracts and compounds from brown algae. Food Chem. 2013, 138, 1764–1785. [Google Scholar] [CrossRef] [PubMed]
- Domínguez, H. Functional Ingredients from Algae for Foods and Nutraceuticals; Woodhead Publishing Limited: Cambridge, UK, 2013. [Google Scholar]
- Wang, R.; Paul, V.J.; Luesch, H. Seaweed extracts and unsaturated fatty acid constituents from the green alga Ulva lactuca as activators of the cytoprotective Nrf2–ARE pathway. Free Radic. Biol. Med. 2013, 57, 141–153. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Division/Common Name | Specie | Pigments | Storage Product | Cell Structural Wall | Intercellular Mucilage |
|---|---|---|---|---|---|
| Phaeophyta/Brown algae | Sargassum polycystum | Chlorophyll a, c Fucoxanthins β-carotenes Xanthophylls | Laminarans Mannitol | Cellulose, Chitin | Alginic acid/Alginates, Sulfated polysaccharides (Fucoidans) |
| Himanthalia elongata | |||||
| Undaria pinnatifida | |||||
| Laminaria spp. | |||||
| Laminaria japónica, L digitata | |||||
| Hizikia fusiforme | |||||
| Chlorophyta/Green algae | Ulva spp. | Chlorophyll a, b Xanthophylls | Starch | Cellulose, Xylans, Mannans | Sulfated polysaccharides |
| Ulva lactuta | |||||
| Ulva pertusa | |||||
| Enteromorpha spp. (E. compresa) | |||||
| Rhodophyta/Red algae | Pyropia spp. | Phycoerythrin Phycocyanin Chlorophyll a β-carotene Xanthophylls | Florideans Starch | Cellulose, Xylans, Mannans | Sulfated polysaccharides (Agar, Carrageenans, Porphyrans) |
| Pyropia tenera | |||||
| Pyropia yezoensis | |||||
| Chondrus crispu | |||||
| Gracilaria verrucosa |
| Study Assay | Study Characteristics | Source and Content | Bioactivity | References |
|---|---|---|---|---|
| Intervention trial | Diet with large amount of seaweed 417 male Japanese T2DM 65 years or older | Total vegetable intake | ↓ HbA1c, Tg, waist circumference | [120] |
| >150 g of daily total vegetable | ↓ HbA1c | |||
| >200 g of total vegetable intake | ↓ Serum Tg ↓ HbA1c | |||
| Green vegetable intake | ↓ Body mass index, Tg, waist circumference | |||
| Double blind, randomized, placebo-controlled crossover study | 23 participants 19–59 year old | Blend of Ascophyllum nodosum and Fucus vesiculosus | ↓ Insulin concentrations ↑ Insulin sensitive | [215] |
| Randomized Crossover Trial | 26 participants | Undaria pinnatifida (4 g, dry alga) + Rice (200 g) | ↓Postprandial glycemia ↓ Insulin levels | [176] |
| Randomized crossover study | 12 participants | 70 g Mekabu(sporophylls of Undaria pinnatifida) | ↓ Postprandial glycemia ↓ Glucose area under the curve | [178] |
| Randomized controlled trial | 12 overweight, healthy males. Aged 40 year | Ascophyllum nodosum enriched bread (4%) | ↓ Energy intake | [182] |
| In vitro assay | Brown marine algae from Eastern Canada. | Fucoidan extracted from Ascophyllum nodosum | Inhibit α-glucosidade and α-amylase activities | [179] |
| Mouse model and human hepatic cells | Male C57BL/6J mice 12 weeks | Food additive carrageenan (E-407) Drinking water (10 mg carrageenan/L) HepG2 Cells (1 mg carrageenan/L × 24 h) | ↓ Glucose tolerance ↑ Insulin resistance Inhibit insulin signalling | [183] |
| In vivo assay | Male Wistar rats 16 weeks | Lota-Carrageenans from Sarconema filiforme (5%, last 8 weeks) | ↓ Body weight ↓ Abdominal and liver fat Improve symptoms of high-carbohydrate, high-fat diet-induced metabolic syndrome. | [185] |
| Randomized controlled trial | 10 healthy male volunteers, studied on three occasions | Agar (2.0 g) | ↓ Delay gastric emptying No effect on the postprandial glucose response | [187] |
| Randomized controlled trial | 76 obese patients with type 2 diabetes 12 weeks | Agar (180 g) + Traditional Japanese food | ↓ HbA1c, ↓ Visceral fat area, subcutaneous fat area, total body fat, ↓ Insulin area under the curve after oral glucose tolerance test ↓Total cholesterol p. | [188] |
| In vitro and in vivo assay | Normal C57/BL6 mice 4 weeks | Laminarin (50 mg/mL) | ↑ GLP-1 secretion and c-Fos protein expression in STC-1 cells | [189] |
| In vivo assay | Adult male Wistar rats | Sodium alginate from Laminaria angustata. Natural and three water-soluble low-molecular weight | Natural and 50 and 100 kDa molecular weights of alginates: ↓ Glucose tolerance ↓ Cholesterol excretion | [190] |
| In vitro and vivo assay | Wistar rats | Calcium alginate | Inhibited α-glucosidase activity in vitro Suppression postprandial increase of blood glucose | [193] |
| Randomized controlled trial | 48 overweight or obese participants 10 days | Sodium alginate from Laminaria digitata | No effect on gastric motor functions, satiation, appetite, or gut hormones | [195] |
| Randomized placebo-controlled trial | 176 participants 5 weeks | Fiber supplements of alginate + balanced 1200 Kcal diet | ↓ Body weight | [196] |
| In vitro and in vivo assay | IEC-6 cell line Male db/db mice | Fucoidans from eleven species of brown algae | Specially from Fucus vesiculosus: ↓ α-glucosidase activity ↓ Fasting blood glucose ↓ HbA1c | [203] |
| In vitro assay | Human colonic carcinoma Caco-2 cells | Five brown species of alga | Cold water and ethanol extracts of Ascophyllum nodosum: ↓ α-glucosidase activity | [206] |
| In vitro assay | 3T3-L1 cells | Fucoidan from Undaria pinnatifida Several concentrations | ↑ Glucose uptake ↓ Lipolysis ↓ Expression of PPARϒ | [208] |
| In vivo assay | Diabetic KK-A(y) mice 5 weeks | Polyphenols from Ecklonia kurome (0.1%) | ↓ α-glucosidase and α-amylase activity ↓ Postprandial blood glucose ↑ Glucose tolerance ↓ Fasting blood glucose ↓ Insulin levels | [212] |
| In vivo assay | Male diabetic KK-A(y) mice 5 weeks | Polyphenols (Phlorotannins) from Ecklonia stolonifera (0–1%) | ↓ Increase in plasma glucose ↓ Increase in lipid peroxidation in plasma | [213] |
| In vivo assay | Male Wistar rats | Ascophyllum nodosum and Fucus vesiculosus extract (10% polyphenols) (7.5 mg/kg body weight) | ↓ α-glucosidase and α-amylase activity ↓ Postprandial blood glucose | [214] |
| Double-blind, placebo-controlled, randomised croos-overal trial | 38 healthy adults (Asian and non-Asian) | Polyphenol-rich Fucus vesiculosus extract (500–200 mg) | ↑ Risk of insulin resistance among Asian populations | [215] |
| In vivo | Rat model of type 2 diabetes | Sargassum polycystum, alcohol (150 and 300 mg kg (−1) body weight) water extract (150 and 300 mg Kg (−1)) | ↓ Blood glucose ↓ HbA1c levels ↑ Response to insulin | [216] |
| In vivo | Obese murine model 10 weeks | Fucoxanthin-rich Undaria pinnatifida (Wakame) lipids (carotenoid) | ↓ Alterations in lipid metabolism and IR induced by a HF diet | [217] |
| In vitro assay | Insulin-resistant HepG2 cells | Fucosterol from Ecklonia stolonifera | ↑ Glucose uptake ↑ Insulin resistance by down regulating expression of PTP1B ↑ Insulin signaling pathway | [218] |
| In vivo assay | Growing Zucker fa-fa rats 8 weeks | Modified AIM-93 diets containing 30% of freeze-dried skid surimi formulated with glucomannan (30%) or with glucomannan plus Spirulina (30%) without added cholesterol | ↓ Hyperglycemia (glucomannan + Spirulina), ↑ Adiponectin/leptin ratio in adipose tissue increase | [221] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bocanegra, A.; Macho-González, A.; Garcimartín, A.; Benedí, J.; Sánchez-Muniz, F.J. Whole Alga, Algal Extracts, and Compounds as Ingredients of Functional Foods: Composition and Action Mechanism Relationships in the Prevention and Treatment of Type-2 Diabetes Mellitus. Int. J. Mol. Sci. 2021, 22, 3816. https://doi.org/10.3390/ijms22083816
Bocanegra A, Macho-González A, Garcimartín A, Benedí J, Sánchez-Muniz FJ. Whole Alga, Algal Extracts, and Compounds as Ingredients of Functional Foods: Composition and Action Mechanism Relationships in the Prevention and Treatment of Type-2 Diabetes Mellitus. International Journal of Molecular Sciences. 2021; 22(8):3816. https://doi.org/10.3390/ijms22083816
Chicago/Turabian StyleBocanegra, Aránzazu, Adrián Macho-González, Alba Garcimartín, Juana Benedí, and Francisco José Sánchez-Muniz. 2021. "Whole Alga, Algal Extracts, and Compounds as Ingredients of Functional Foods: Composition and Action Mechanism Relationships in the Prevention and Treatment of Type-2 Diabetes Mellitus" International Journal of Molecular Sciences 22, no. 8: 3816. https://doi.org/10.3390/ijms22083816
APA StyleBocanegra, A., Macho-González, A., Garcimartín, A., Benedí, J., & Sánchez-Muniz, F. J. (2021). Whole Alga, Algal Extracts, and Compounds as Ingredients of Functional Foods: Composition and Action Mechanism Relationships in the Prevention and Treatment of Type-2 Diabetes Mellitus. International Journal of Molecular Sciences, 22(8), 3816. https://doi.org/10.3390/ijms22083816