
Heterogeneous Nuclear Ribonucleoprotein K Is Involved in the Estrogen-Signaling Pathway in Breast Cancer

 , ,
, ,
Abstract
1. Introduction
2. Results
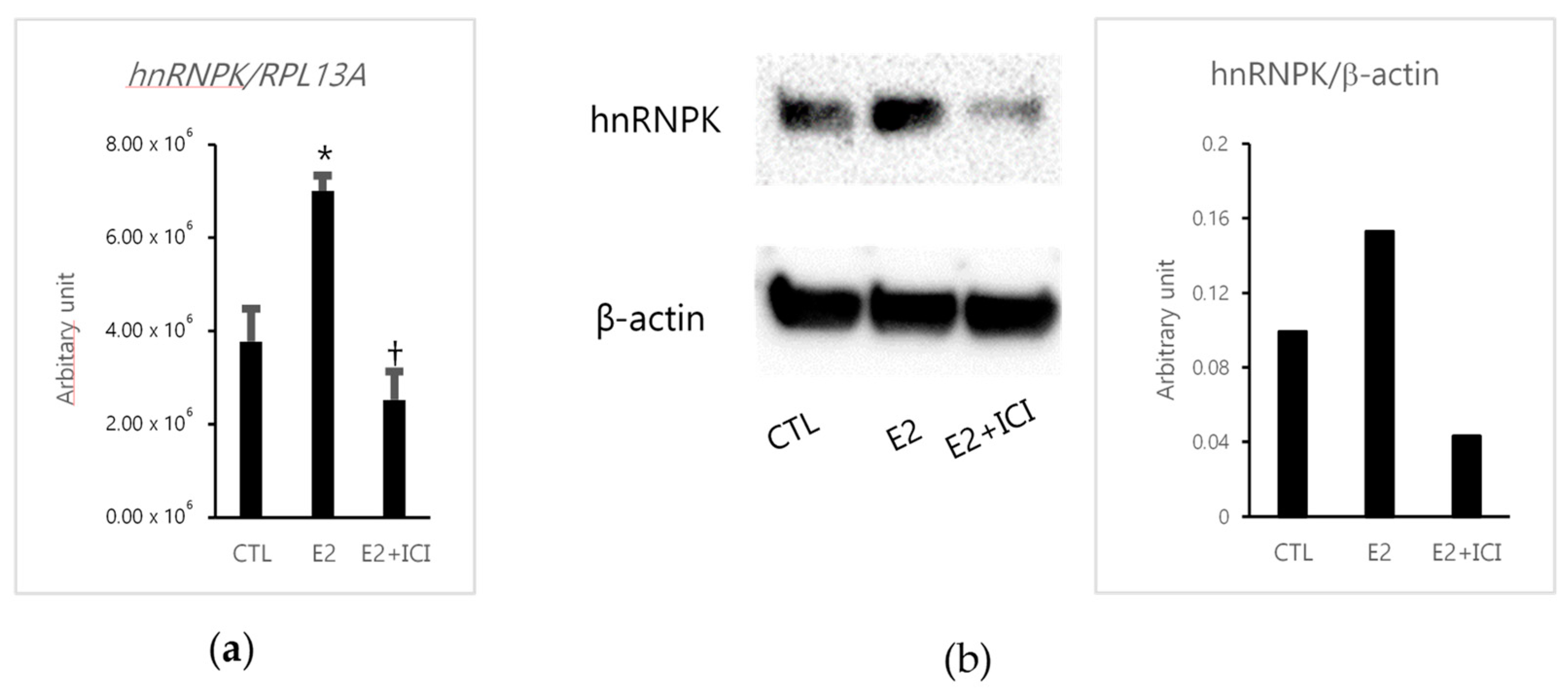
2.1. The Effects of Estradiol (E2) in MCF-7 Cells
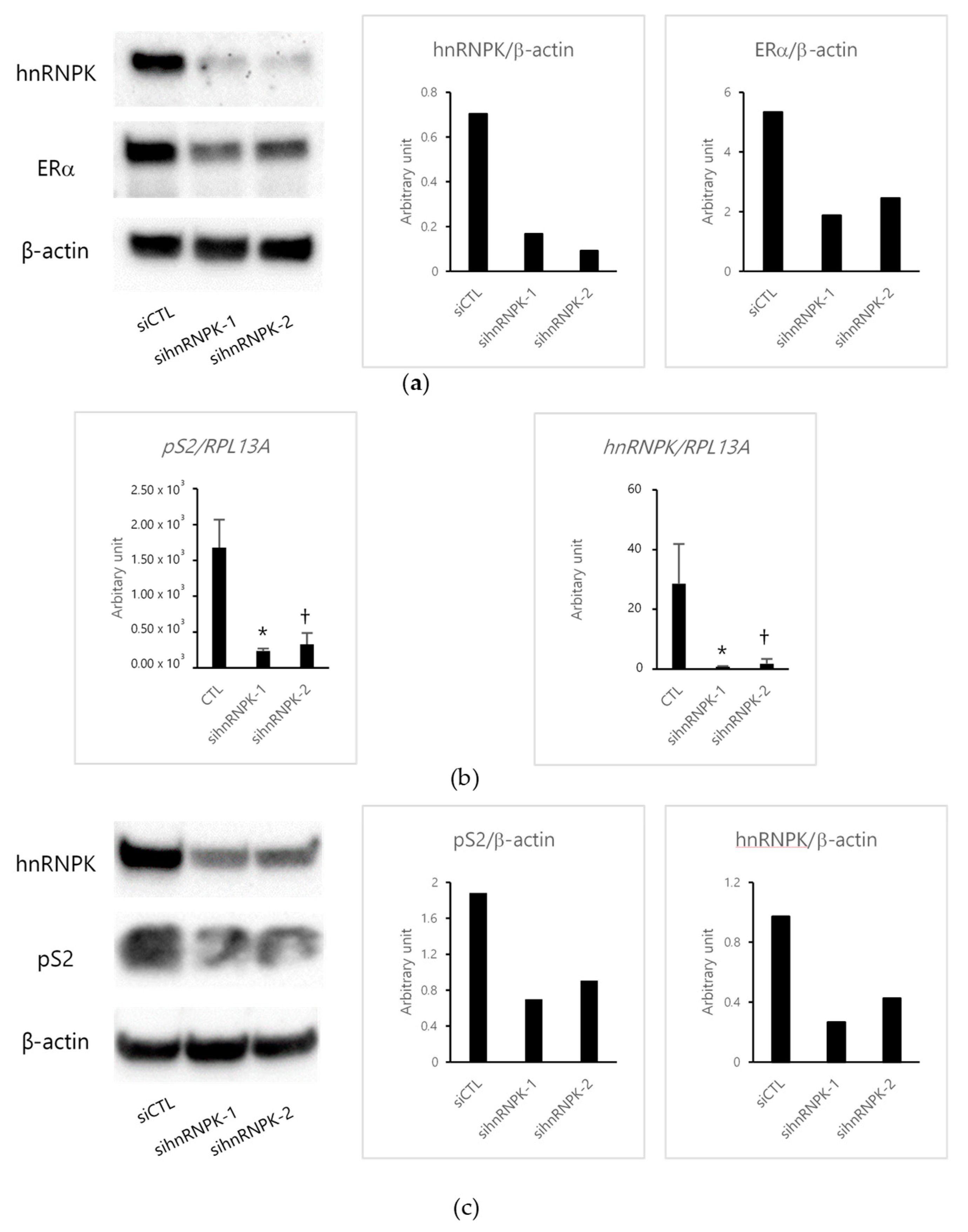
2.2. The Effects of E2 in hnRNPK-Knockdown-MCF-7 Cells
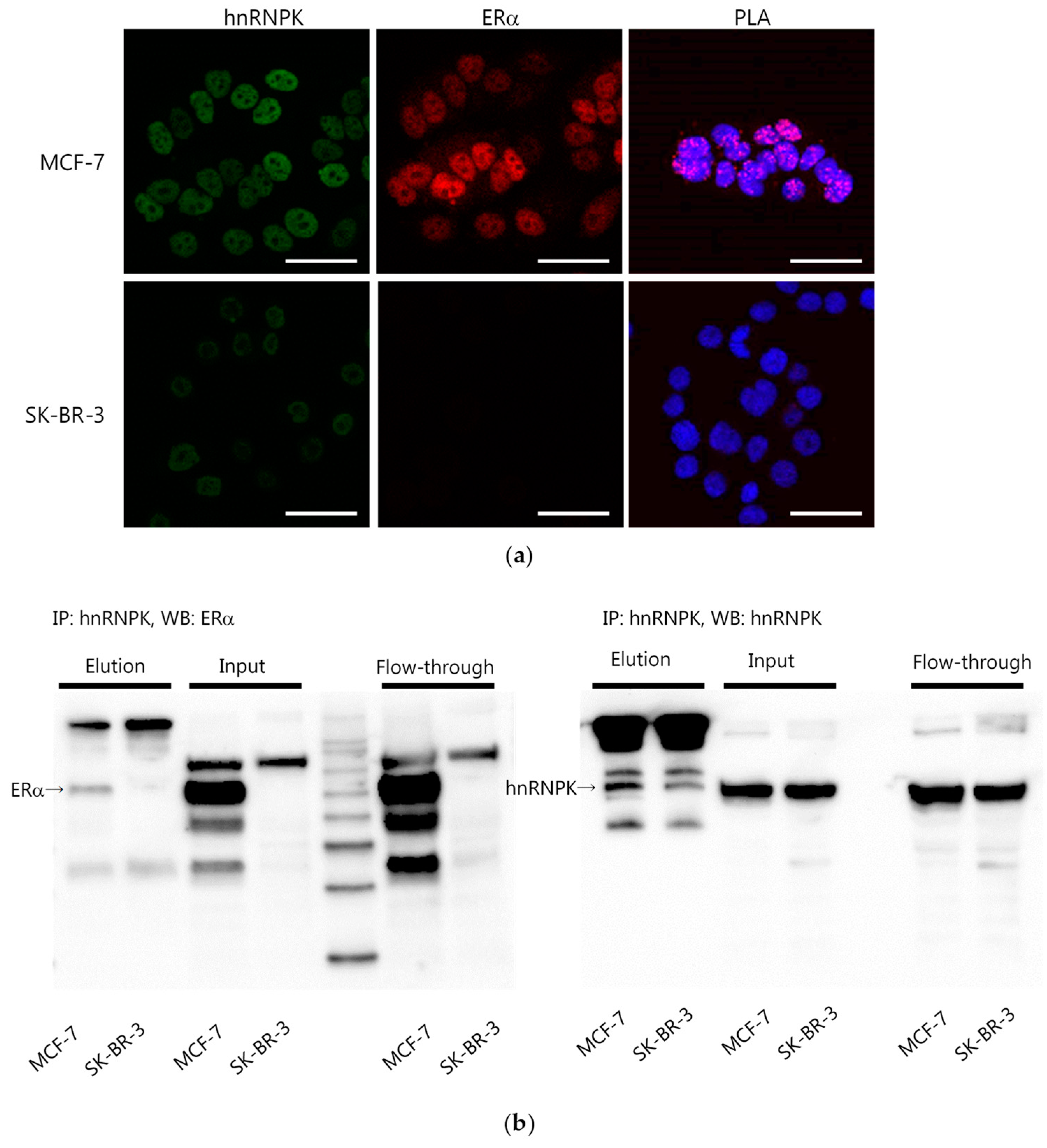
2.3. hnRNPK and ERα Interaction
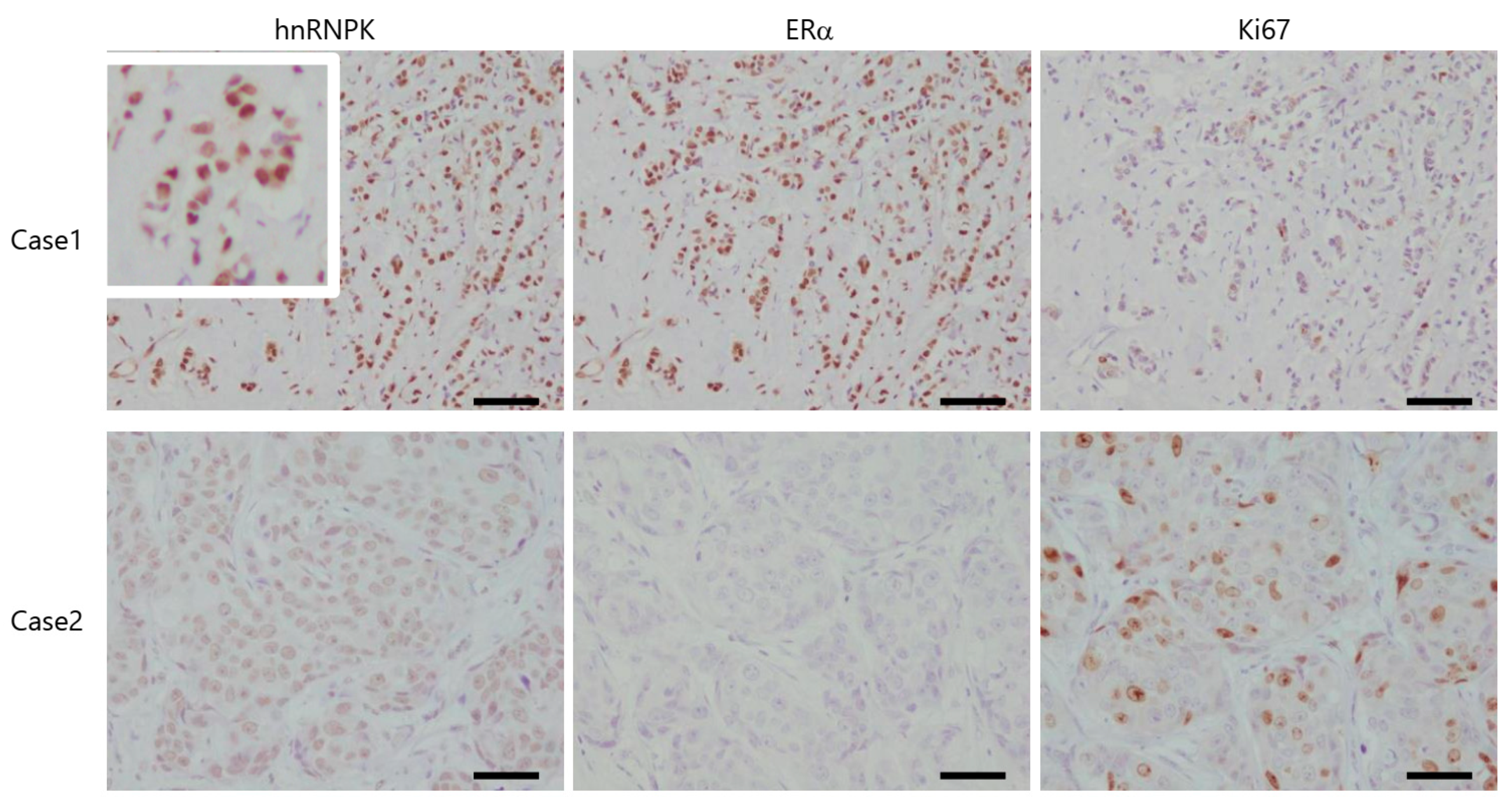
2.4. hnRNPK and ERα and Its Association with Clinicopathological Parameters of Patients with Breast Cancer
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Breast Cancer Tissues
4.3. The Effects of E2 in MCF-7 Cells
4.4. The Effects of E2 in hnRNPK-Knockdown-MCF-7 Cells
4.5. Quantitative Reverse Transcription Real-Time PCR
4.6. Western Blotting
4.7. Immunofluorescence
4.8. In Situ PLA
4.9. Immunoprecipitation
4.10. Immunohistochemistry
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| E2 | Estradiol |
| ER | Estrogen receptor |
| HER2 | Human epidermal growth factor receptor 2 |
| hnRNPK | Heterogeneous nuclear ribonucleoprotein |
| LI | Labeling index |
| SERM | Selective estrogen receptor modulator |
| siRNA | Small interfering RNA |
| PLA | Proximity ligation assay |
| PR | Progesterone receptor |
References
- Madak-Erdogan, Z.; Charn, T.-H.; Jiang, Y.; Liu, E.T.; Katzenellenbogen, J.A.; Katzenellenbogen, B.S. Integrative genomics of gene and metabolic regulation by estrogen receptors α and β, and their coregulators. Mol. Syst. Biol. 2013, 9, 676. [Google Scholar] [CrossRef] [PubMed]
- Mehta, R.; Jain, R.K.; Badve, S. Personalized Medicine: The Road Ahead. Clin. Breast Cancer 2011, 11, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Rondón-Lagos, M.; Villegas, V.E.; Rangel, N.; Sánchez, M.C.; Zaphiropoulos, P.G. Tamoxifen resistance: Emerging molecular targets. Int. J. Mol. Sci. 2016, 17, 1357. [Google Scholar] [CrossRef]
- Bomsztyk, K.; Denisenko, O.; Ostrowski, J. hnRNP K: One protein multiple processes. Bioessays 2004, 26, 629–638. [Google Scholar] [CrossRef]
- Matta, A.; Tripathi, S.C.; DeSouza, L.V.; Grigull, J.; Kaur, J.; Chauhan, S.S.; Srivastava, A.; Thakar, A.; Shukla, N.K.; Duggal, R.; et al. Heterogeneous ribonucleoprotein K is a marker of oral leukoplakia and correlates with poor prognosis of squamous cell carcinoma. Int. J. Cancer 2009, 125, 1398–1406. [Google Scholar] [CrossRef] [PubMed]
- Barboro, P.; Ferrari, N.; Balbi, C. Emerging roles of heterogeneous nuclear ribonucleoprotein K (hnRNP K) in cancer progression. Cancer Lett. 2014, 352, 152–159. [Google Scholar] [CrossRef]
- Chen, X.; Gu, P.; Xie, R.; Han, J.; Liu, H.; Wang, B.; Xie, W.; Xie, W.; Zhong, G.; Chen, C.; et al. Heterogeneous nuclear ribonucleoprotein K is associated with poor prognosis and regulates proliferation and apoptosis in bladder cancer. J. Cell. Mol. Med. 2017, 21, 1266–1279. [Google Scholar] [CrossRef]
- Carpenter, B.; Mckay, M.; Dundas, S.R.; Lawrie, L.C.; Telfer, C.; Murray, G.I. Heterogeneous nuclear ribonucleoprotein K is over expressed, aberrantly localised and is associated with poor prognosis in colorectal cancer. Br. J. Cancer 2006, 95, 921–927. [Google Scholar] [CrossRef]
- Huang, H.; Han, Y.; Yang, X.; Li, M.; Zhu, R.; Hu, J.; Zhang, X.; Wei, R.; Li, K.; Gao, R. HNRNPK inhibits gastric cancer cell proliferation through p53/p21/CCND1 pathway. Oncotarget 2017, 8, 103364–103374. [Google Scholar] [CrossRef] [PubMed]
- Mandal, M.; Vadlamudi, R.; Nguyen, D.; Wang, R.A.; Costa, L.; Bagheri-Yarmand, R.; Mendelsohn, J.; Kumar, R. Growth Factors Regulate Heterogeneous Nuclear Ribonucleoprotein K Expression and Function. J. Biol. Chem. 2001, 276, 9699–9704. [Google Scholar] [CrossRef] [PubMed]
- Nagai, M.A.; Da Rós, N.; Neto, M.M.; de Faria Junior, S.R.; Brentani, M.M.; Hirata, R.; Neves, E.J. Gene expression profiles in breast tumors regarding the presence or absence of estrogen and progesterone receptors. Int. J. Cancer 2004, 111, 892–899. [Google Scholar] [CrossRef]
- Ostrowski, J.; Schullery, D.S.; Denisenko, O.N.; Higaki, Y.; Watts, J.; Aebersold, R.; Stempka, L.; Gschwendt, M.; Bomsztykt, K. Role of tyrosine phosphorylation in the regulation of the interaction of heterogenous nuclear ribonucleoprotein K protein with its protein and RNA partners. J. Biol. Chem. 2000, 275, 3619–3628. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Wu, W.; Han, Q.; Wang, Y.; Li, C.; Zhang, P.; Xu, H. New Insights into the Interplay between Non-Coding RNAs and RNA-Binding Protein HnRNPK in Regulating Cellular Functions. Cells 2019, 8, 62. [Google Scholar] [CrossRef]
- Li, D.; Wang, X.; Mei, H.; Fang, E.; Ye, L.; Song, H.; Yang, F.; Li, H.; Huang, K.; Zheng, L.; et al. Long Noncoding RNA pancEts-1 Promotes Neuroblastoma Progression through hnRNPK-Mediated β-Catenin Stabilization. Cancer Res. 2018, 78, 1169–1183. [Google Scholar] [CrossRef] [PubMed]
- Onodera, Y.; Miki, Y.; Suzuki, T.; Takagi, K.; Akahira, J.I.; Sakyu, T.; Watanabe, M.; Inoue, S.; Ishida, T.; Ohuchi, N.; et al. Runx2 in human breast carcinoma: Its potential roles in cancer progression. Cancer Sci. 2010, 101, 2670–2675. [Google Scholar] [CrossRef] [PubMed]
- Ciarlo, M.; Benelli, R.; Barbieri, O.; Minghelli, S.; Barboro, P.; Balbi, C.; Ferrari, N. Regulation of neuroendocrine differentiation by AKT/hnRNPK/AR/β-catenin signaling in prostate cancer cells. Int. J. Cancer 2012, 131, 582–590. [Google Scholar] [CrossRef]
- Caligiuri, I.; Toffoli, G.; Giordano, A.; Rizzolio, F. pRb controls Estrogen Receptor alpha protein stability and activity. Oncotarget 2013, 4, 875–883. [Google Scholar] [CrossRef] [PubMed]
- Fliss, A.E.; Benzeno, S.; Rao, J.; Caplan, A.J. Control of estrogen receptor ligand binding by Hsp90. J. Steroid Biochem. Mol. Biol. 2000, 72, 223–230. [Google Scholar] [CrossRef]
- Rosa, F.E.; Caldeira, J.R.F.; Felipes, J.; Bertonha, F.B.; Quevedo, F.C.; Domingues, M.A.C.; Moraes Neto, F.A.; Rogatto, S.R. Evaluation of estrogen receptor α and β and progesterone receptor expression and correlation with clinicopathologic factors and proliferative marker Ki-67 in breast cancers. Hum. Pathol. 2008, 39, 720–730. [Google Scholar] [CrossRef]
- Tecalco-Cruz, A.C.; Ramírez-Jarquín, J.O. Mechanisms that Increase Stability of Estrogen Receptor Alpha in Breast Cancer. Clin. Breast Cancer 2017, 17. [Google Scholar] [CrossRef] [PubMed]
- Iwabuchi, E.; Miki, Y.; Ono, K.; Onodera, Y.; Suzuki, T.; Hirakawa, H.; Ishida, T.; Ohuchi, N.; Sasano, H. In situ detection of estrogen receptor dimers in breast carcinoma cells in archival materials using proximity ligation assay (PLA). J. Steroid Biochem. Mol. Biol. 2017, 165, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Iwabuchi, E.; Miki, Y.; Onodera, Y.; Shibahara, Y.; Takagi, K.; Suzuki, T.; Ishida, T.; Sasano, H. Co-expression of carcinoembryonic antigen-related cell adhesion molecule 6 and 8 inhibits proliferation and invasiveness of breast carcinoma cells. Clin. Exp. Metastasis 2019. [Google Scholar] [CrossRef] [PubMed]
- Iwabuchi, E.; Miki, Y.; Ono, K.; Onodera, Y.; Sasano, H. In Situ Evaluation of Estrogen Receptor Dimers in Breast Carcinoma Cells: Visualization of Protein-Protein Interactions. Acta Histochem. Cytochem. 2017, 50, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Iwabuchi, E.; Miki, Y.; Kanai, A.; Miyashita, M.; Kijima, G.; Hirakawa, H.; Suzuki, T.; Ishida, T.; Sasano, H. The interaction between carcinoembryonic antigen-related cell adhesion molecule 6 and HER2 is associated with therapeutic efficacy of trastuzumab in breast cancer. J. Pathol. 2018, 246, 379–389. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| hnRNPK | ||||
|---|---|---|---|---|
| Positive (n = 37) | Negative (n = 37) | p Value | ||
| Histological Grade | 1 | 9 | 5 | p = 0.0536 |
| 2 | 19 | 14 | ||
| 3 | 8 | 18 | ||
| Stage | 1 | 14 | 4 | p = 0.0232 |
| 2 | 14 | 21 | ||
| 3 | 7 | 9 | ||
| Pathologic T factor | 1 | 20 | 9 | p = 0.0092 |
| ≥2 | 15 | 25 | ||
| Lymph node metastasis | Positive | 12 | 22 | p = 0.0175 |
| Negative | 24 | 14 | ||
| ERα LI Median (Range) | 72 (0–98) | 18 (0–95) | p = 0.0027 | |
| PR LI Median (Range) | 13 (0–83) | 8 (0–90) | p = 0.7136 | |
| HER2 | Positive | 8 | 6 | p = 0.5522 |
| Negative | 29 | 31 | ||
| Ki67 LI Median (Range) | 12 (0–31) | 19 (3–53) | p = 0.0057 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iwabuchi, E.; Miki, Y.; Suzuki, T.; Hirakawa, H.; Ishida, T.; Sasano, H. Heterogeneous Nuclear Ribonucleoprotein K Is Involved in the Estrogen-Signaling Pathway in Breast Cancer. Int. J. Mol. Sci. 2021, 22, 2581. https://doi.org/10.3390/ijms22052581
Iwabuchi E, Miki Y, Suzuki T, Hirakawa H, Ishida T, Sasano H. Heterogeneous Nuclear Ribonucleoprotein K Is Involved in the Estrogen-Signaling Pathway in Breast Cancer. International Journal of Molecular Sciences. 2021; 22(5):2581. https://doi.org/10.3390/ijms22052581
Chicago/Turabian StyleIwabuchi, Erina, Yasuhiro Miki, Takashi Suzuki, Hisashi Hirakawa, Takanori Ishida, and Hironobu Sasano. 2021. "Heterogeneous Nuclear Ribonucleoprotein K Is Involved in the Estrogen-Signaling Pathway in Breast Cancer" International Journal of Molecular Sciences 22, no. 5: 2581. https://doi.org/10.3390/ijms22052581
APA StyleIwabuchi, E., Miki, Y., Suzuki, T., Hirakawa, H., Ishida, T., & Sasano, H. (2021). Heterogeneous Nuclear Ribonucleoprotein K Is Involved in the Estrogen-Signaling Pathway in Breast Cancer. International Journal of Molecular Sciences, 22(5), 2581. https://doi.org/10.3390/ijms22052581