
Aged Skeletal Muscle Retains the Ability to Remodel Extracellular Matrix for Degradation of Collagen Deposition after Muscle Injury



Abstract
1. Introduction
2. Results
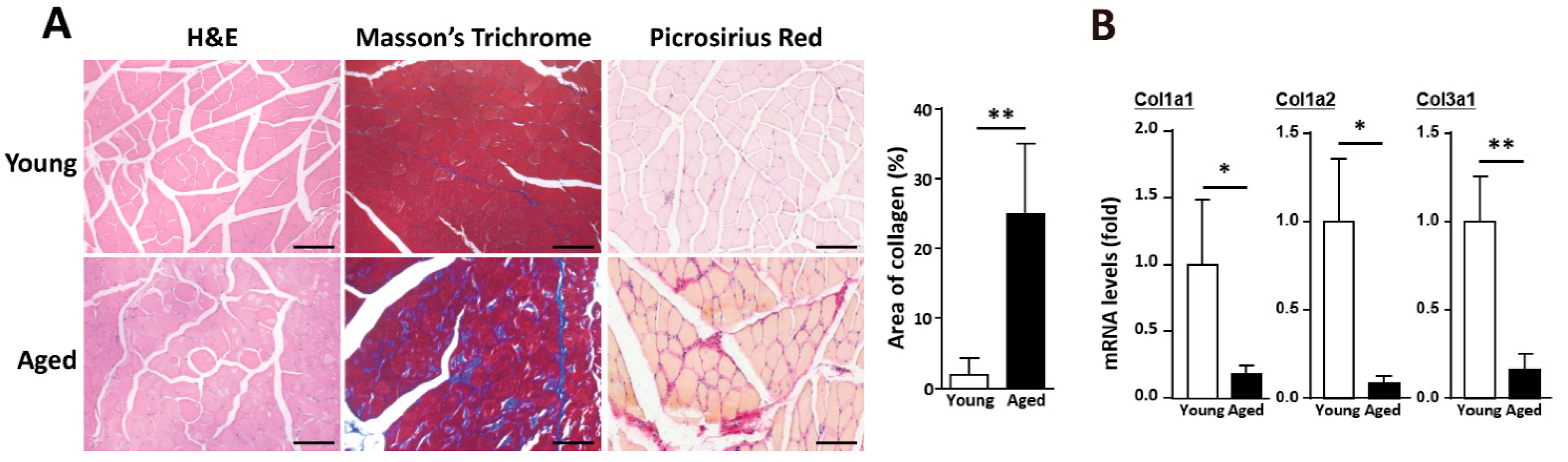
2.1. Alteration of ECM Components in Skeletal Muscle with Age
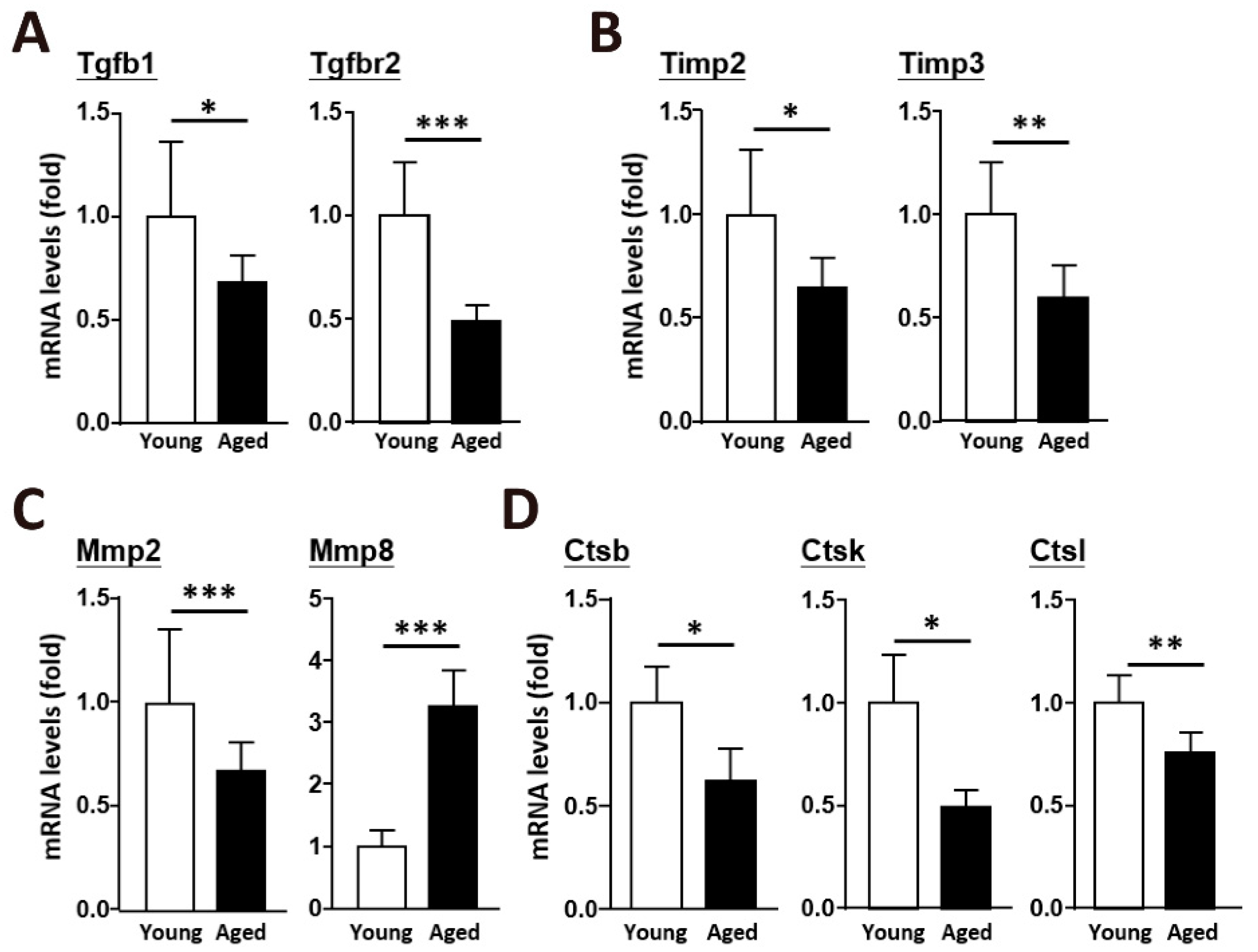
2.2. An Age-Dependent Decline in the Enzymes Involved in ECM Remodeling
2.3. Clearance of Accumulated Collagen in Aged Skeletal Muscle after Muscle Injury
2.4. Possible Mechanisms of the Effects of Aging on ECM Components in Skeletal Muscle
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. RNA Analysis
4.3. Protein Analysis
4.4. Histological Analysis
4.5. Statistical Aanalysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ahmad, K.; Shaikh, S.; Ahmad, S.S.; Lee, E.J.; Choi, I. Cross-Talk between Extracellular Matrix and Skeletal Muscle: Implications for Myopathies. Front. Pharmacol. 2020, 11, 142. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, K.; Lee, E.J.; Moon, J.S.; Park, S.Y.; Choi, I. Multifaceted Interweaving Between Extracellular Matrix, Insulin Resistance, and Skeletal Muscle. Cells 2018, 7, 148. [Google Scholar] [CrossRef]
- Gillies, A.R.; Lieber, R.L. Structure and function of the skeletal muscle extracellular matrix. Muscle Nerve 2011, 44, 318–331. [Google Scholar] [CrossRef]
- Kragstrup, T.W.; Kjaer, M.; Mackey, A.L. Structural, biochemical, cellular, and functional changes in skeletal muscle extracellular matrix with aging. Scand. J. Med. Sci. Sports 2011, 21, 749–757. [Google Scholar] [CrossRef]
- Frantz, C.; Stewart, K.M.; Weaver, V.M. The extracellular matrix at a glance. J. Cell Sci. 2010, 123, 4195–4200. [Google Scholar] [CrossRef] [PubMed]
- Sanes, J.R. The basement membrane/basal lamina of skeletal muscle. J. Biol. Chem. 2003, 278, 12601–12604. [Google Scholar] [CrossRef]
- Thomas, K.; Engler, A.J.; Meyer, G.A. Extracellular matrix regulation in the muscle satellite cell niche. Connect. Tissue Res. 2015, 56, 1–8. [Google Scholar] [CrossRef]
- Bella, J.; Hulmes, D.J. Fibrillar Collagens. Subcell Biochem. 2017, 82, 457–490. [Google Scholar] [PubMed]
- Gelse, K.; Poschl, E.; Aigner, T. Collagens–Structure, function, and biosynthesis. Adv. Drug Deliv. Rev. 2003, 55, 1531–1546. [Google Scholar] [CrossRef]
- Light, N.; Champion, A.E. Characterization of muscle epimysium, perimysium and endomysium collagens. Biochem. J. 1984, 219, 1017–1026. [Google Scholar] [CrossRef] [PubMed]
- Kang, L.; Ayala, J.E.; Lee-Young, R.S.; Zhang, Z.; James, F.D.; Neufer, P.D.; Pozzi, A.; Zutter, M.M.; Wasserman, D.H. Diet-induced muscle insulin resistance is associated with extracellular matrix remodeling and interaction with integrin alpha2beta1 in mice. Diabetes 2011, 60, 416–426. [Google Scholar] [CrossRef]
- Ricard-Blum, S.; Ruggiero, F. The collagen superfamily: From the extracellular matrix to the cell membrane. Pathol. Biol. 2005, 53, 430–442. [Google Scholar] [CrossRef]
- Halfter, W.; Dong, S.; Schurer, B.; Cole, G.J. Collagen XVIII is a basement membrane heparan sulfate proteoglycan. J. Biol. Chem. 1998, 273, 25404–25412. [Google Scholar] [CrossRef] [PubMed]
- Arthur, S.T.; Cooley, I.D. The effect of physiological stimuli on sarcopenia; impact of Notch and Wnt signaling on impaired aged skeletal muscle repair. Int. J. Biol. Sci. 2012, 8, 731–760. [Google Scholar] [CrossRef] [PubMed]
- Dumont, N.A.; Wang, Y.X.; Rudnicki, M.A. Intrinsic and extrinsic mechanisms regulating satellite cell function. Development 2015, 142, 1572–1581. [Google Scholar] [CrossRef] [PubMed]
- Beauchamp, J.R.; Heslop, L.; Yu, D.S.; Tajbakhsh, S.; Kelly, R.G.; Wernig, A.; Buckingham, M.E.; Partridge, T.A.; Zammit, P.S. Expression of CD34 and Myf5 defines the majority of quiescent adult skeletal muscle satellite cells. J. Cell Biol. 2000, 151, 1221–1234. [Google Scholar] [CrossRef] [PubMed]
- Maesner, C.C.; Almada, A.E.; Wagers, A.J. Established cell surface markers efficiently isolate highly overlapping populations of skeletal muscle satellite cells by fluorescence-activated cell sorting. Skelet Muscle 2016, 6, 35. [Google Scholar] [CrossRef]
- Almada, A.E.; Wagers, A.J. Molecular circuitry of stem cell fate in skeletal muscle regeneration, ageing and disease. Nat. Rev. Mol. Cell Biol. 2016, 17, 267–279. [Google Scholar] [CrossRef]
- Cooper, R.N.; Tajbakhsh, S.; Mouly, V.; Cossu, G.; Buckingham, M.; Butler-Browne, G.S. In vivo satellite cell activation via Myf5 and MyoD in regenerating mouse skeletal muscle. J. Cell Sci. 1999, 112, 2895–2901. [Google Scholar]
- Cornelison, D.D.; Wold, B.J. Single-cell analysis of regulatory gene expression in quiescent and activated mouse skeletal muscle satellite cells. Dev. Biol. 1997, 191, 270–283. [Google Scholar] [CrossRef]
- Abmayr, S.M.; Pavlath, G.K. Myoblast fusion: Lessons from flies and mice. Development 2012, 139, 641–656. [Google Scholar] [CrossRef]
- Zammit, P.S. Function of the myogenic regulatory factors Myf5, MyoD, Myogenin and MRF4 in skeletal muscle, satellite cells and regenerative myogenesis. Semin. Cell Dev. Biol. 2017, 72, 19–32. [Google Scholar] [CrossRef]
- Davis, M.E.; Gumucio, J.P.; Sugg, K.B.; Bedi, A.; Mendias, C.L. MMP inhibition as a potential method to augment the healing of skeletal muscle and tendon extracellular matrix. J. Appl. Physiol. 2013, 115, 884–891. [Google Scholar] [CrossRef]
- Lin, I.H.; Chang, J.L.; Hua, K.; Huang, W.C.; Hsu, M.T.; Chen, Y.F. Skeletal muscle in aged mice reveals extensive transformation of muscle gene expression. BMC Genet 2018, 19, 55. [Google Scholar] [CrossRef]
- Goldspink, G.; Fernandes, K.; Williams, P.E.; Wells, D.J. Age-related changes in collagen gene expression in the muscles of mdx dystrophic and normal mice. Neuromuscul. Disord. 1994, 4, 183–191. [Google Scholar] [CrossRef]
- Lieber, R.L.; Ward, S.R. Cellular mechanisms of tissue fibrosis. 4. Structural and functional consequences of skeletal muscle fibrosis. Am. J. Physiol. Cell Physiol. 2013, 305, C241–C252. [Google Scholar] [CrossRef] [PubMed]
- Stefano, C.; Stefano, S. Corrigendum: Regeneration of Mammalian Skeltal Muscle: Basic Mechanisms and Clinical Implications. Curr. Pharm. Des. 2015, 21, 4657. [Google Scholar]
- Zhou, Y.; Lovell, D.; Bethea, M.; Yoseph, B.; Poteracki, J.; Soker, S.; Criswell, T. The Impact of Age on Skeletal Muscle Progenitor Cell Survival and Fate after Injury. Tissue Eng. Methods 2017, 23, 1012–1021. [Google Scholar] [CrossRef] [PubMed]
- Serrano, A.L.; Munoz-Canoves, P. Regulation and dysregulation of fibrosis in skeletal muscle. Exp. Cell Res. 2010, 316, 3050–3058. [Google Scholar] [CrossRef] [PubMed]
- Brack, A.S.; Conboy, M.J.; Roy, S.; Lee, M.; Kuo, C.J.; Keller, C.; Rando, T.A. Increased Wnt signaling during aging alters muscle stem cell fate and increases fibrosis. Science 2007, 317, 807–810. [Google Scholar] [CrossRef]
- Mackey, A.L.; Donnelly, A.E.; Turpeenniemi-Hujanen, T.; Roper, H.P. Skeletal muscle collagen content in humans after high-force eccentric contractions. J. Appl. Physiol. 2004, 97, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Csapo, R.; Gumpenberger, M.; Wessner, B. Skeletal Muscle Extracellular Matrix-What Do We Know About Its Composition, Regulation, and Physiological Roles? A Narrative Review. Front. Physiol. 2020, 11, 253. [Google Scholar] [CrossRef]
- Thorsteinsdottir, S.; Deries, M.; Cachaco, A.S.; Bajanca, F. The extracellular matrix dimension of skeletal muscle development. Dev. Biol. 2011, 354, 191–207. [Google Scholar] [CrossRef]
- Alameddine, H.S. Matrix metalloproteinases in skeletal muscles: Friends or foes? Neurobiol. Dis. 2012, 48, 508–518. [Google Scholar] [CrossRef] [PubMed]
- Visse, R.; Nagase, H. Matrix metalloproteinases and tissue inhibitors of metalloproteinases: Structure, function, and biochemistry. Circ. Res. 2003, 92, 827–839. [Google Scholar] [CrossRef] [PubMed]
- Chuang, H.M.; Chen, Y.S.; Harn, H.J. The Versatile Role of Matrix Metalloproteinase for the Diverse Results of Fibrosis Treatment. Molecules 2019, 24, 4188. [Google Scholar] [CrossRef] [PubMed]
- Christensen, S.; Purslow, P.P. The role of matrix metalloproteinases in muscle and adipose tissue development and meat quality: A review. Meat Sci. 2016, 119, 138–146. [Google Scholar] [CrossRef]
- Vidak, E.; Javorsek, U.; Vizovisek, M.; Turk, B. Cysteine Cathepsins and their Extracellular Roles: Shaping the Microenvironment. Cells 2019, 8, 264. [Google Scholar] [CrossRef] [PubMed]
- Mondino, A.; Blasi, F. uPA and uPAR in fibrinolysis, immunity and pathology. Trends Immunol. 2004, 25, 450–455. [Google Scholar] [CrossRef]
- Daley, W.P.; Peters, S.B.; Larsen, M. Extracellular matrix dynamics in development and regenerative medicine. J. Cell Sci. 2008, 121, 255–264. [Google Scholar] [CrossRef]
- De Sousa Neto, I.V.; Durigan, J.L.Q.; Guzzoni, V.; Tibana, R.A.; Prestes, J.; de Araujo, H.S.S.; Marqueti, R.C. Effects of Resistance Training on Matrix Metalloproteinase Activity in Skeletal Muscles and Blood Circulation During Aging. Front. Physiol. 2018, 9, 190. [Google Scholar] [CrossRef]
- Labat-Robert, J.; Robert, L. Interaction between cells and extracellular matrix: Signaling by integrins and the elastin-laminin receptor. Prog. Mol. Subcell Biol. 2000, 25, 57–70. [Google Scholar]
- Stoka, V.; Turk, V.; Turk, B. Lysosomal cathepsins and their regulation in aging and neurodegeneration. Ageing Res. Rev. 2016, 32, 22–37. [Google Scholar] [CrossRef]
- Zhou, S.; Qian, B.; Wang, L.; Zhang, C.; Hogan, M.V.; Li, H. Altered bone-regulating myokine expression in skeletal muscle Of Duchenne muscular dystrophy mouse models. Muscle Nerve 2018, 58, 573–582. [Google Scholar] [CrossRef] [PubMed]
- Melouane, A.; Carbonell, A.; Yoshioka, M.; Puymirat, J.; St-Amand, J. Implication of SPARC in the modulation of the extracellular matrix and mitochondrial function in muscle cells. PLoS ONE 2018, 13, e0192714. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.J.; Christofidou, E.D.; Keene, D.R.; Hassan Milde, M.; Adams, J.C. Intermolecular interactions of thrombospondins drive their accumulation in extracellular matrix. Mol. Biol. Cell 2015, 26, 2640–2654. [Google Scholar] [CrossRef]
- Chung, C.Y.; Erickson, H.P. Glycosaminoglycans modulate fibronectin matrix assembly and are essential for matrix incorporation of tenascin-C. J. Cell Sci. 1997, 110, 1413–1419. [Google Scholar] [PubMed]
- Cifuentes-Diaz, C.; Faille, L.; Goudou, D.; Schachner, M.; Rieger, F.; Angaut-Petit, D. Abnormal reinnervation of skeletal muscle in a tenascin-C-deficient mouse. J. Neurosci. Res. 2002, 67, 93–99. [Google Scholar] [CrossRef]
- Tierney, M.T.; Gromova, A.; Sesillo, F.B.; Sala, D.; Spenle, C.; Orend, G.; Sacco, A. Autonomous Extracellular Matrix Remodeling Controls a Progressive Adaptation in Muscle Stem Cell Regenerative Capacity during Development. Cell Rep. 2016, 14, 1940–1952. [Google Scholar] [CrossRef]
- Calve, S.; Odelberg, S.J.; Simon, H.G. A transitional extracellular matrix instructs cell behavior during muscle regeneration. Dev. Biol. 2010, 344, 259–271. [Google Scholar] [CrossRef]
- Lukjanenko, L.; Karaz, S.; Stuelsatz, P.; Gurriaran-Rodriguez, U.; Michaud, J.; Dammone, G.; Sizzano, F.; Mashinchian, O.; Ancel, S.; Migliavacca, E.; et al. Aging Disrupts Muscle Stem Cell Function by Impairing Matricellular WISP1 Secretion from Fibro-Adipogenic Progenitors. Cell Stem Cell 2019, 24, 433–446. [Google Scholar] [CrossRef]
- Heinemeier, K.M.; Bjerrum, S.S.; Schjerling, P.; Kjaer, M. Expression of extracellular matrix components and related growth factors in human tendon and muscle after acute exercise. Scand. J. Med. Sci. Sports 2013, 23, e150–e161. [Google Scholar] [CrossRef] [PubMed]
- Hjorth, M.; Norheim, F.; Meen, A.J.; Pourteymour, S.; Lee, S.; Holen, T.; Jensen, J.; Birkeland, K.I.; Martinov, V.N.; Langleite, T.M.; et al. The effect of acute and long-term physical activity on extracellular matrix and serglycin in human skeletal muscle. Physiol. Rep. 2015, 3, e12473. [Google Scholar] [CrossRef] [PubMed]
- Hyldahl, R.D.; Nelson, B.; Xin, L.; Welling, T.; Groscost, L.; Hubal, M.J.; Chipkin, S.; Clarkson, P.M.; Parcell, A.C. Extracellular matrix remodeling and its contribution to protective adaptation following lengthening contractions in human muscle. FASEB J. 2015, 29, 2894–2904. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, C.; Weigert, C. Skeletal Muscle as an Endocrine Organ: The Role of Myokines in Exercise Adaptations. Cold Spring Harb. Perspect. Med. 2017, 7, a029793. [Google Scholar] [CrossRef] [PubMed]
- Reisenauer, A.; Eickelberg, O.; Wille, A.; Heimburg, A.; Reinhold, A.; Sloane, B.F.; Welte, T.; Buhling, F. Increased carcinogenic potential of myeloid tumor cells induced by aberrant TGF-beta1-signaling and upregulation of cathepsin B. Biol. Chem. 2007, 388, 639–650. [Google Scholar] [CrossRef]
- Mahdy, M.A.A. Skeletal muscle fibrosis: An overview. Cell Tissue Res. 2019, 375, 575–588. [Google Scholar] [CrossRef] [PubMed]
- Gumpenberger, M.; Wessner, B.; Graf, A.; Narici, M.V.; Fink, C.; Braun, S.; Hoser, C.; Blazevich, A.J.; Csapo, R. Remodeling the Skeletal Muscle Extracellular Matrix in Older Age-Effects of Acute Exercise Stimuli on Gene Expression. Int. J. Mol. Sci. 2020, 21, 7089. [Google Scholar] [CrossRef]
- Wessner, B.; Liebensteiner, M.; Nachbauer, W.; Csapo, R. Age-specific response of skeletal muscle extracellular matrix to acute resistance exercise: A pilot study. Eur. J. Sport Sci. 2019, 19, 354–364. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| chr | Gene | Name | SM-Young | SM-Aged | Aged to Young (logfc) |
|---|---|---|---|---|---|
| Gene Expression Involved in Fibrillar and Fibril-Associated Collagen | |||||
| chr6 | Col1a2 | ENSMUSG00000029661.9 | 506.20 | 188.90 | −1.418 |
| chr11 | Col1a1 | ENSMUSG00000001506.9 | 252.60 | 88.43 | −1.504 |
| chr15 | Col2a1 | ENSMUSG00000022483.9 | 0.24 | 0.20 | −0.045 |
| chr1 | Col3a1 | ENSMUSG00000026043.11 | 604.40 | 224.10 | −1.427 |
| chr2 | Col5a1 | ENSMUSG00000026837.9 | 65.31 | 44.19 | −0.553 |
| chr1 | Col5a2 | ENSMUSG00000026042.9 | 69.95 | 44.04 | −0.656 |
| chr3 | Col11a1 | ENSMUSG00000027966.10 | 5.08 | 1.25 | −1.435 |
| chr4 | Col27a1 | ENSMUSG00000045672.7 | 6.59 | 5.29 | −0.272 |
| chr9 | Col12a1 | ENSMUSG00000032332.10 | 18.83 | 8.73 | −1.027 |
| chr15 | Col14a1 | ENSMUSG00000022371.8 | 25.70 | 19.70 | −0.367 |
| Gene Expression Involved in Networking Collagen | |||||
| chr8 | Col4a1 | ENSMUSG00000031502.9 | 202.00 | 191.70 | −0.075 |
| chr1 | Col4a4 | ENSMUSG00000067158.7 | 5.22 | 4.74 | −0.116 |
| chrX | Col4a5 | ENSMUSG00000031274.9 | 13.42 | 6.98 | −0.853 |
| chr10 | Col6a1 | ENSMUSG00000001119.6 | 116.50 | 85.94 | −0.434 |
| chr10 | Col6a2 | ENSMUSG00000020241.7 | 105.40 | 79.30 | −0.406 |
| chr1 | Col6a3 | ENSMUSG00000048126.9 | 113.20 | 76.96 | −0.550 |
| chr9 | Col6a4 | ENSMUSG00000032572.5 | 1.80 | 0.00 | −1.487 |
| chr9 | Col6a5 | ENSMUSG00000091345.1 | 2.23 | 1.20 | −0.556 |
| chr10 | Col18a1 | ENSMUSG00000001435.8 | 26.61 | 20.90 | −0.334 |
| Down-Regulation of the Gene Expression which Encodes BM Glycoproteins | |||||
| chr2 | Lamc3 | ENSMUSG00000026840.8 | 4.17 | 4.14 | −0.009 |
| chr12 | Lamb1 | ENSMUSG00000002900.8 | 57.48 | 57.06 | −0.010 |
| chr17 | Lama1 | ENSMUSG00000032796.7 | 0.09 | 0.05 | −0.061 |
| chr10 | Lama2 | ENSMUSG00000019899.8 | 80.81 | 67.73 | −0.251 |
| chr1 | Lamc1 | ENSMUSG00000026478.8 | 180.50 | 150.80 | −0.258 |
| chr1 | Fn1 | ENSMUSG00000026193.8 | 210.20 | 122.10 | −0.779 |
| chr5 | Eln | ENSMUSG00000029675.9 | 27.27 | 23.69 | −0.195 |
| chr14 | Nid2 | ENSMUSG00000021806.3 | 18.83 | 18.55 | −0.020 |
| chr13 | Nid1 | ENSMUSG00000005397.7 | 217.50 | 161.70 | −0.425 |
| Down-Regulation of the Gene Expression which Encodes Proteoglycans and GAGs | |||||
| chr4 | Hspg2 | ENSMUSG00000028763.10 | 267.70 | 263.70 | −0.022 |
| chr10 | Dcn | ENSMUSG00000019929.8 | 892.00 | 792.50 | −0.171 |
| chrX | Bgn | ENSMUSG00000031375.10 | 113.70 | 76.01 | −0.575 |
| chr1 | Fmod | ENSMUSG00000041559.7 | 152.20 | 82.30 | −0.879 |
| Down-Regulation of the Gene Expression which Encodes Growth Factors | |||||
| chr7 | Tgfb1 | ENSMUSG00000002603.9 | 12.62 | 7.73 | −0.641 |
| chr9 | Tgfbr2 | ENSMUSG00000032440.5 | 78.77 | 68.03 | −0.209 |
| chr10 | Igf1 | ENSMUSG00000020053.11 | 64.03 | 36.46 | −0.796 |
| chr10 | Ctgf | ENSMUSG00000019997.4 | 40.17 | 29.33 | −0.441 |
| Gene Expression which Encodes Matrix Matricellular Proteins | |||||
| chr11 | Sparc | ENSMUSG00000018593.6 | 583.80 | 351.50 | −0.730 |
| chr2 | Thbs1 | ENSMUSG00000040152.8 | 15.98 | 23.54 | 0.531 |
| chr17 | Thbs2 | ENSMUSG00000023885.7 | 35.19 | 34.12 | −0.043 |
| chr3 | Thbs3 | ENSMUSG00000028047.5 | 10.72 | 8.98 | −0.232 |
| chr13 | Thbs4 | ENSMUSG00000021702.7 | 160.50 | 100.70 | −0.667 |
| chr4 | Tnc | ENSMUSG00000028364.8 | 11.10 | 4.79 | −1.064 |
| chr5 | Spp1 | ENSMUSG00000029304.8 | 4.51 | 10.67 | 1.084 |
| chr15 | Wisp1 | ENSMUSG00000005124.3 | 1.47 | 0.50 | −0.721 |
| Gene Expression which Encodes Cathepsins | |||||
| chr3 | Ctsk | ENSMUSG00000028111.3 | 14.28 | 9.28 | −0.572 |
| chr13 | Ctsl | ENSMUSG00000021477.7 | 153.80 | 149.50 | −0.040 |
| chr14 | Ctsb | ENSMUSG00000021939.6 | 523.70 | 435.90 | −0.264 |
| chr3 | Ctss | ENSMUSG00000038642.4 | 40.98 | 15.56 | −1.342 |
| chr2 | Ctsz | ENSMUSG00000016256.9 | 52.07 | 42.69 | −0.281 |
| chr7 | Ctsc | ENSMUSG00000030560.9 | 95.28 | 74.07 | −0.359 |
| chr19 | Ctsf | ENSMUSG00000083282.2 | 31.87 | 28.53 | −0.155 |
| chr9 | Ctsh | ENSMUSG00000032359.8 | 34.95 | 18.95 | −0.850 |
| chr3 | Ctso | ENSMUSG00000028015.2 | 20.91 | 19.90 | −0.068 |
| chr19 | Ctsw | ENSMUSG00000024910.4 | 2.09 | 0.10 | −1.489 |
| chr11 | Stat3 | ENSMUSG00000004040.10 | 95.90 | 93.57 | −0.035 |
| chr10 | Stat6 | ENSMUSG00000002147.11 | 47.33 | 41.15 | −0.198 |
| Gene Expression which Encodes Matrix Remodeling Enzymes | |||||
| chr9 | Mmp3 | ENSMUSG00000043613.7 | 9.39 | 3.24 | −1.292 |
| chr8 | Mmp2 | ENSMUSG00000031740.7 | 95.14 | 61.30 | −0.626 |
| chrX | Timp1 | ENSMUSG00000001131.5 | 0.95 | 0.75 | −0.157 |
| chr11 | Timp2 | ENSMUSG00000017466.3 | 261.40 | 226.70 | −0.205 |
| chr10 | Timp3 | ENSMUSG00000020044.7 | 184.30 | 177.20 | −0.057 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, W.-J.; Lin, I.-H.; Lee, C.-W.; Chen, Y.-F. Aged Skeletal Muscle Retains the Ability to Remodel Extracellular Matrix for Degradation of Collagen Deposition after Muscle Injury. Int. J. Mol. Sci. 2021, 22, 2123. https://doi.org/10.3390/ijms22042123
Chen W-J, Lin I-H, Lee C-W, Chen Y-F. Aged Skeletal Muscle Retains the Ability to Remodel Extracellular Matrix for Degradation of Collagen Deposition after Muscle Injury. International Journal of Molecular Sciences. 2021; 22(4):2123. https://doi.org/10.3390/ijms22042123
Chicago/Turabian StyleChen, Wan-Jing, I-Hsuan Lin, Chien-Wei Lee, and Yi-Fan Chen. 2021. "Aged Skeletal Muscle Retains the Ability to Remodel Extracellular Matrix for Degradation of Collagen Deposition after Muscle Injury" International Journal of Molecular Sciences 22, no. 4: 2123. https://doi.org/10.3390/ijms22042123
APA StyleChen, W.-J., Lin, I.-H., Lee, C.-W., & Chen, Y.-F. (2021). Aged Skeletal Muscle Retains the Ability to Remodel Extracellular Matrix for Degradation of Collagen Deposition after Muscle Injury. International Journal of Molecular Sciences, 22(4), 2123. https://doi.org/10.3390/ijms22042123