
Immune-Mediated Aggravation of the Campylobacter concisus-Induced Epithelial Barrier Dysfunction




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
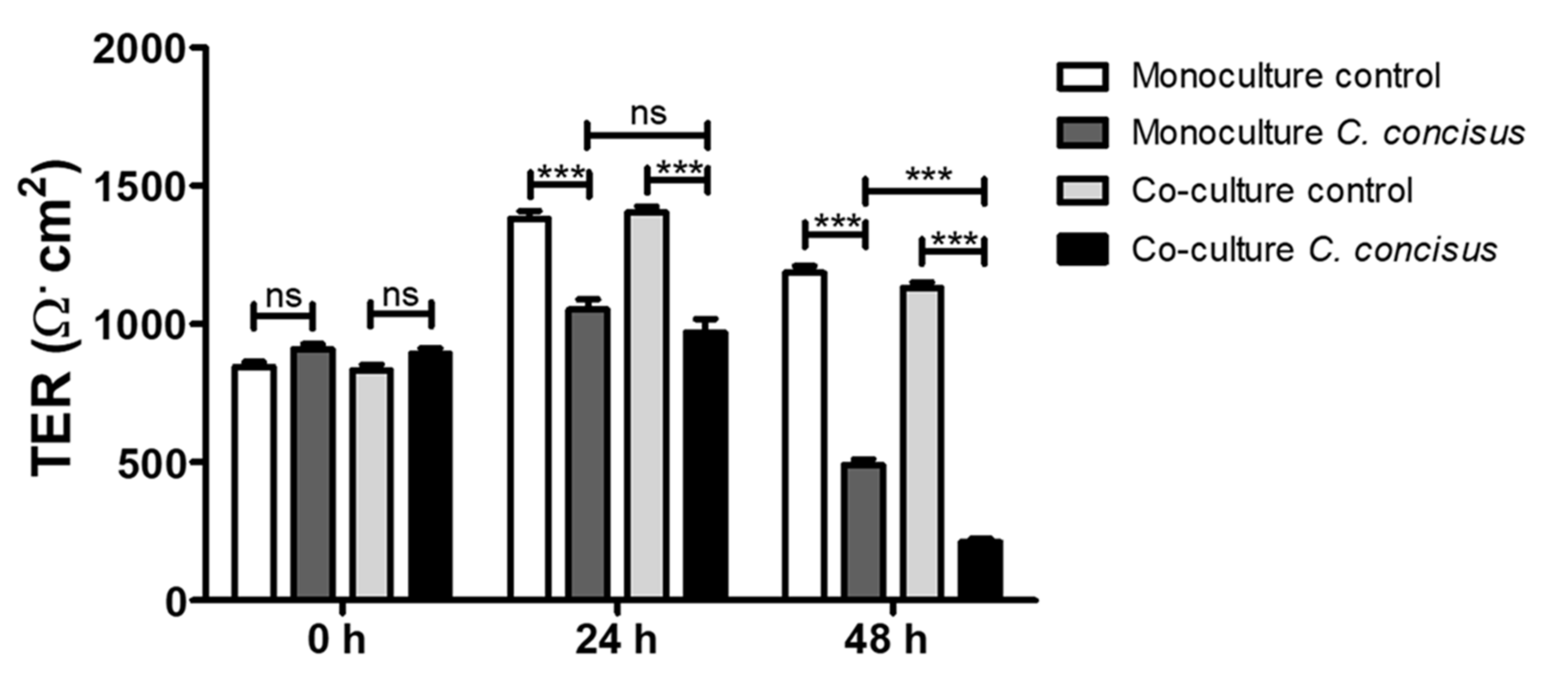
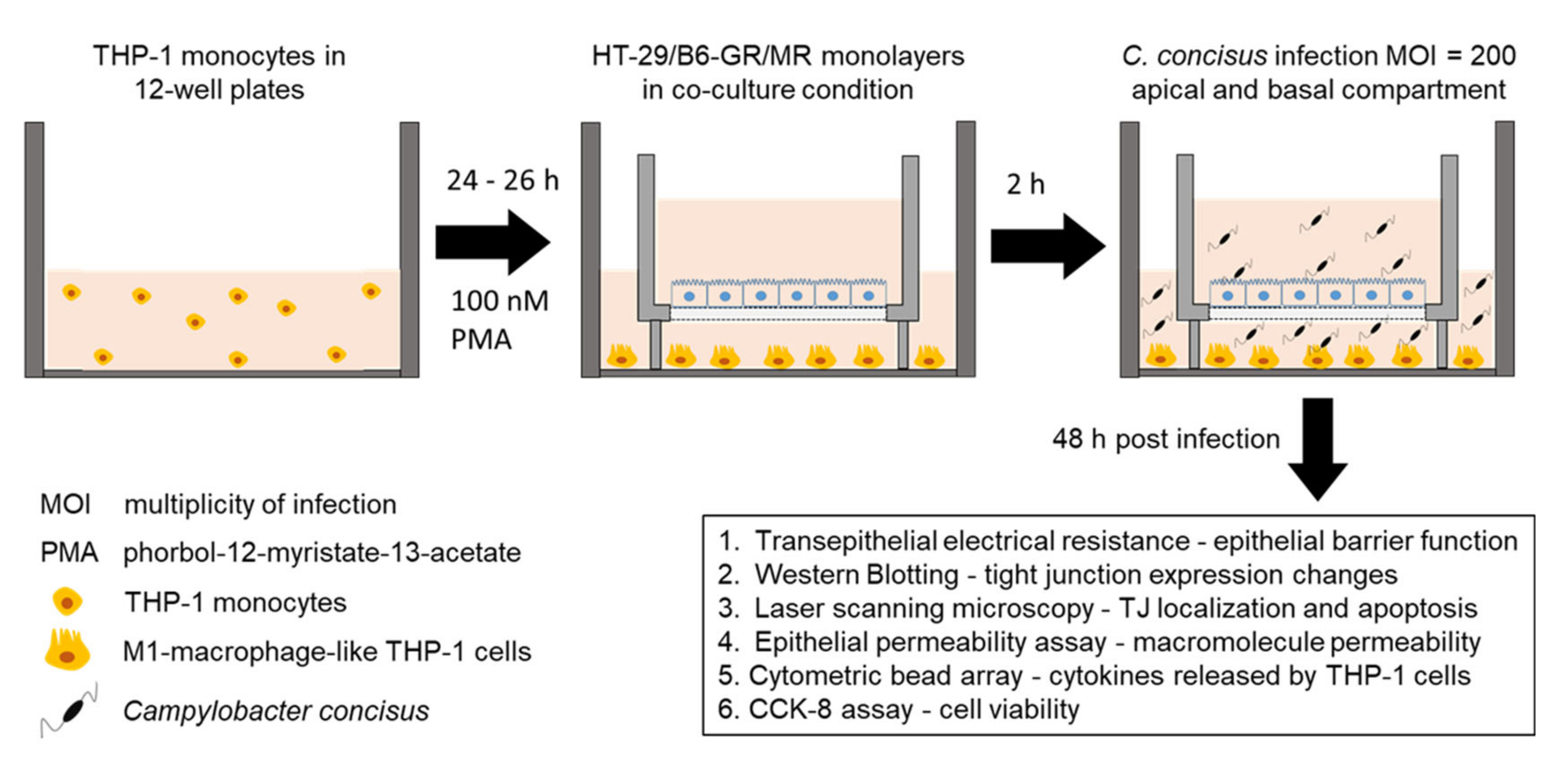
2.1. Campylobacter concisus Aggravates the Barrier Disturbance of Intestinal Epithelial Cells in Co-Culture with Immune Cells
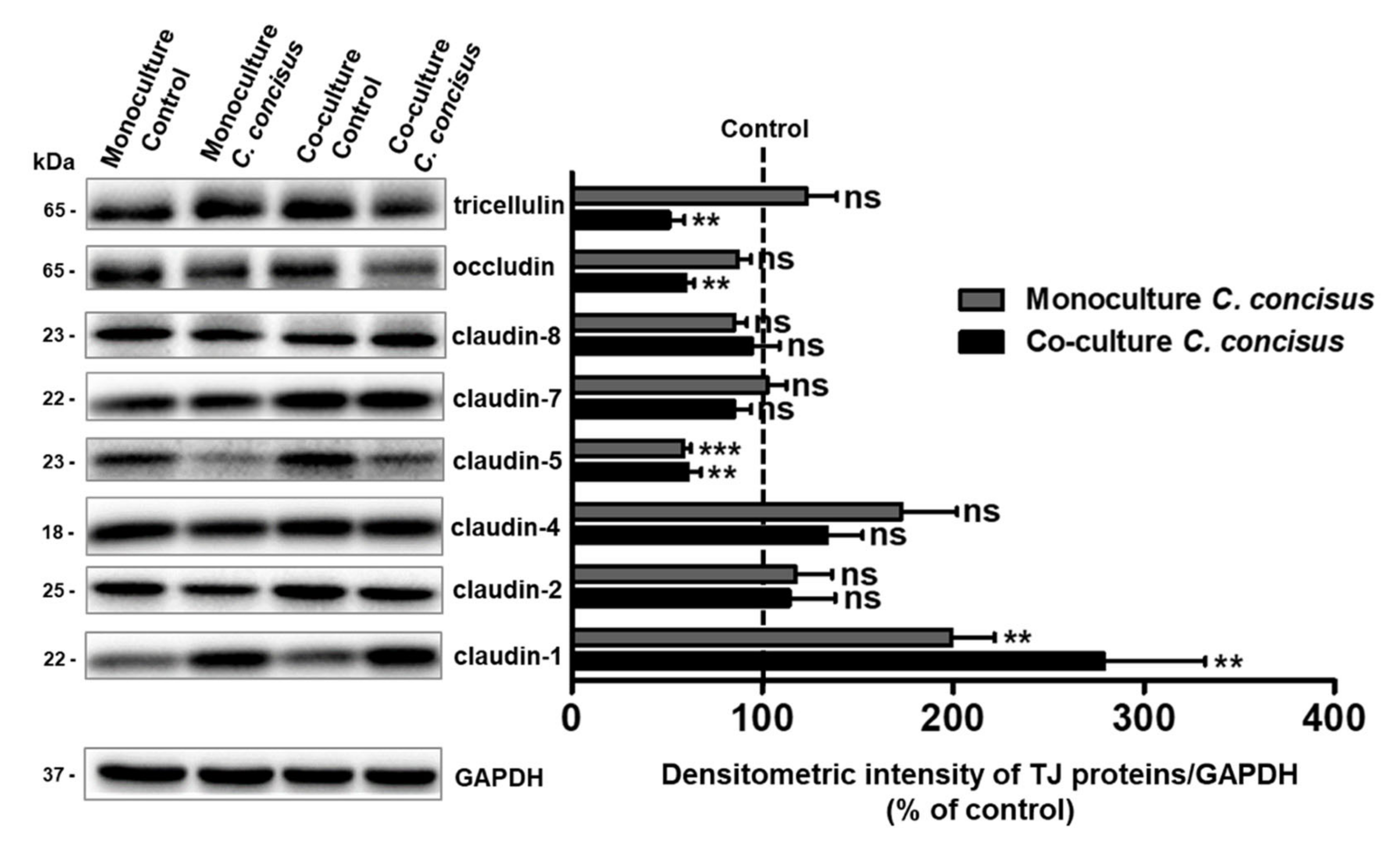
2.2. Protein Expression Changes of Claudins, Occludin and Tricellulin in Colonic Epithelial Cells after C. concisus Infection
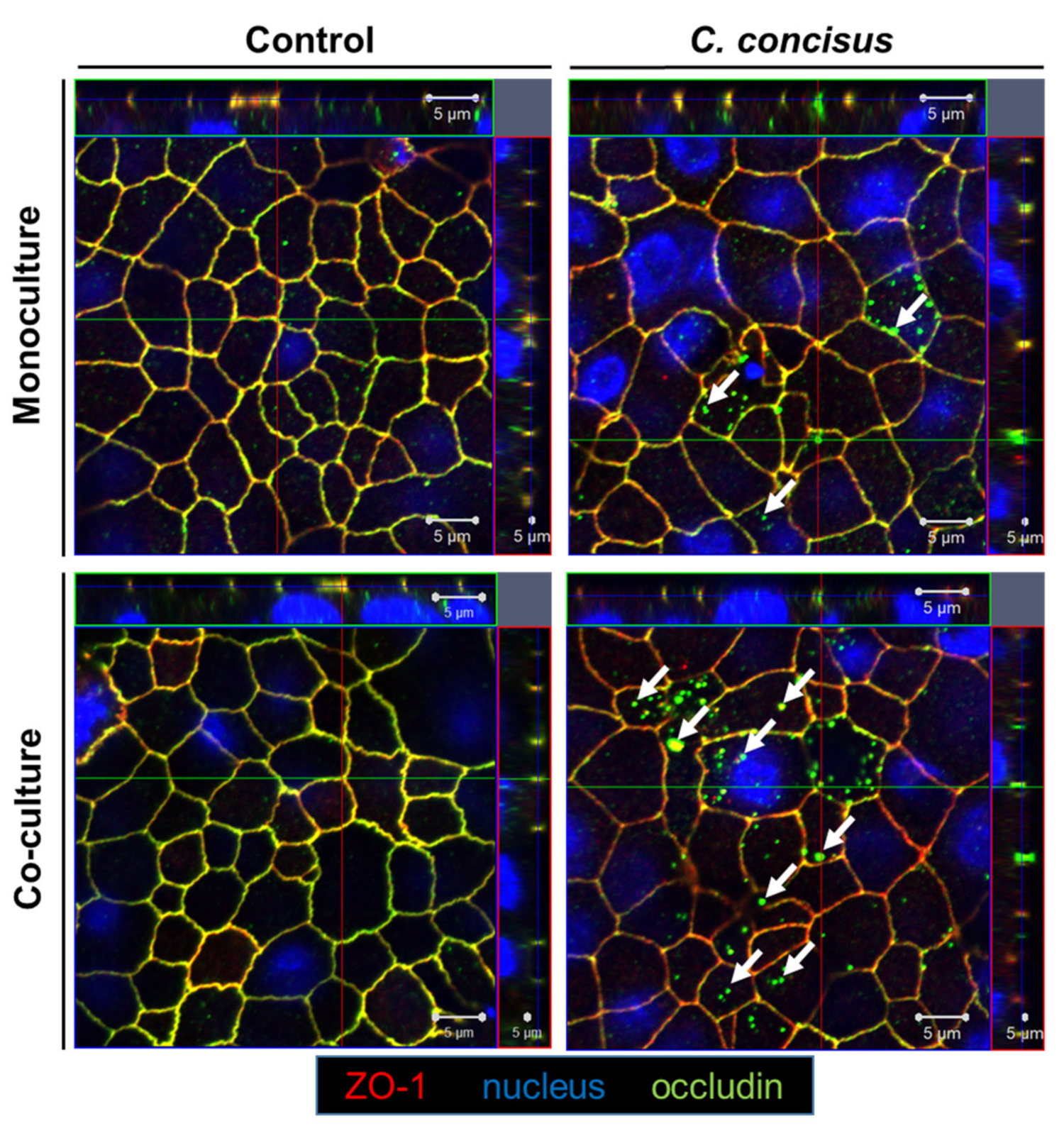
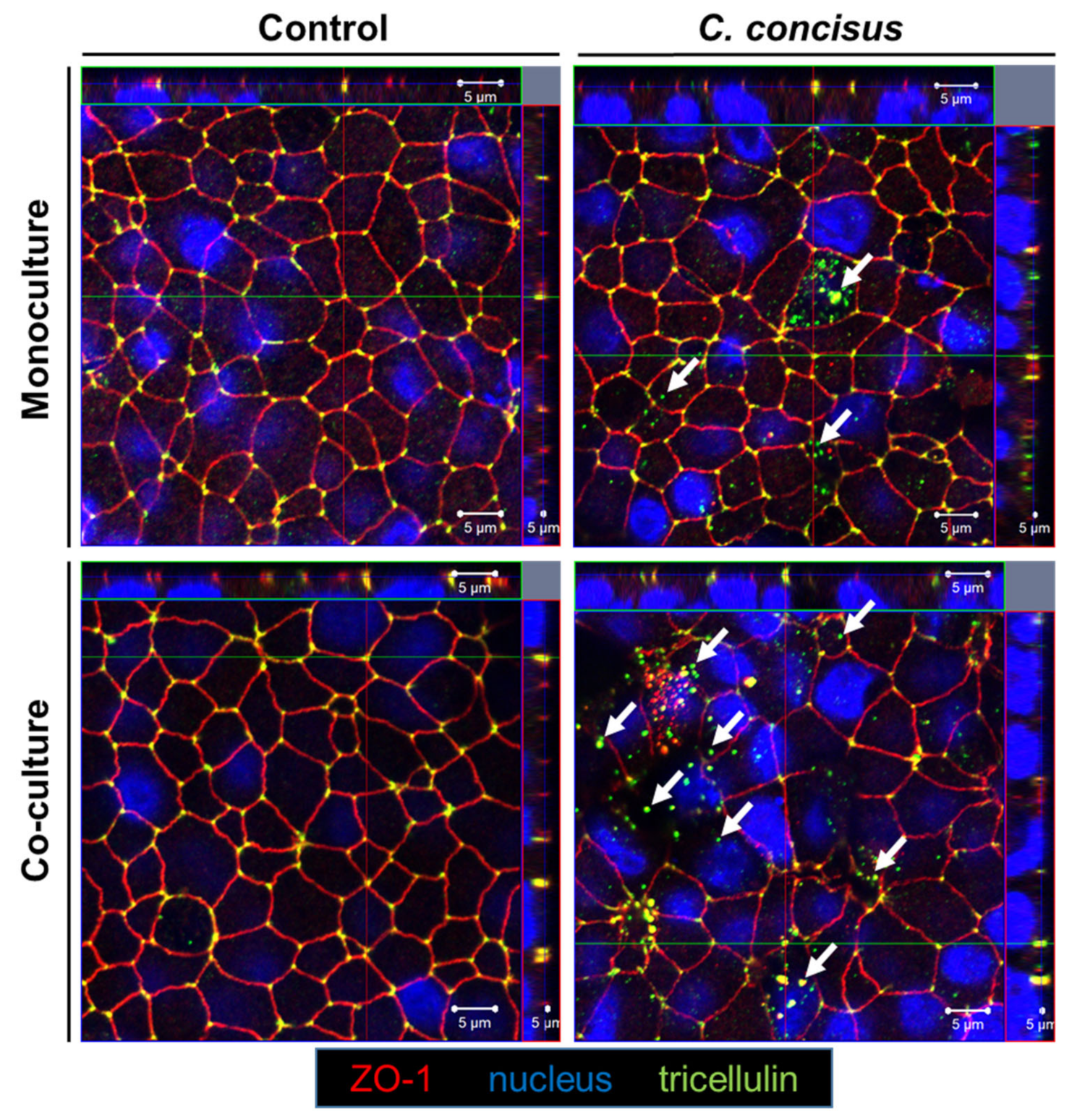
2.3. Subcellular Localization of Occludin and Tricellulin in Colonic Epithelial Cells after Campylobacter concisus Infection
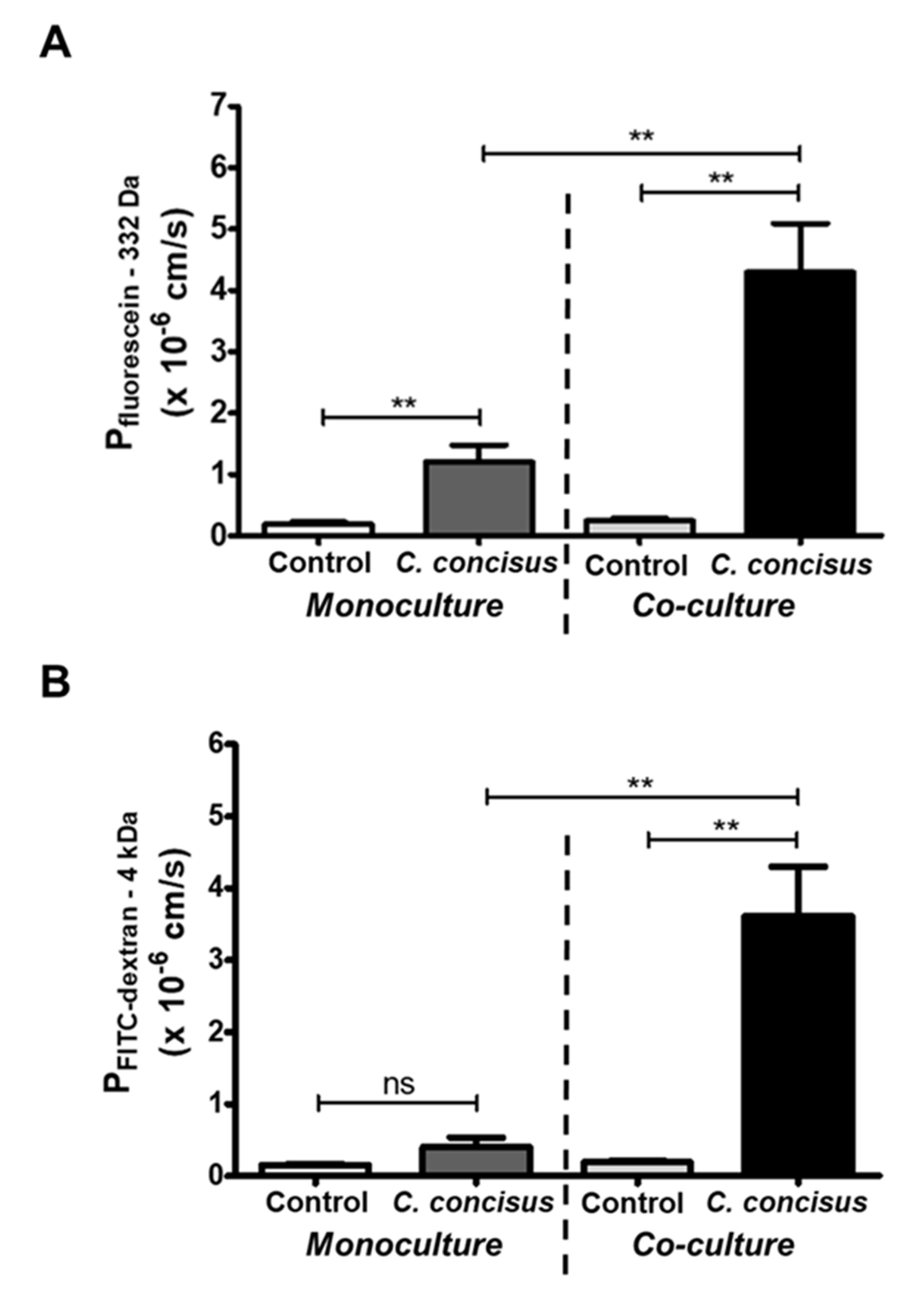
2.4. Epithelial Permeability in Campylobacter concisus-Infected Colonic Epithelial Cell Monolayers
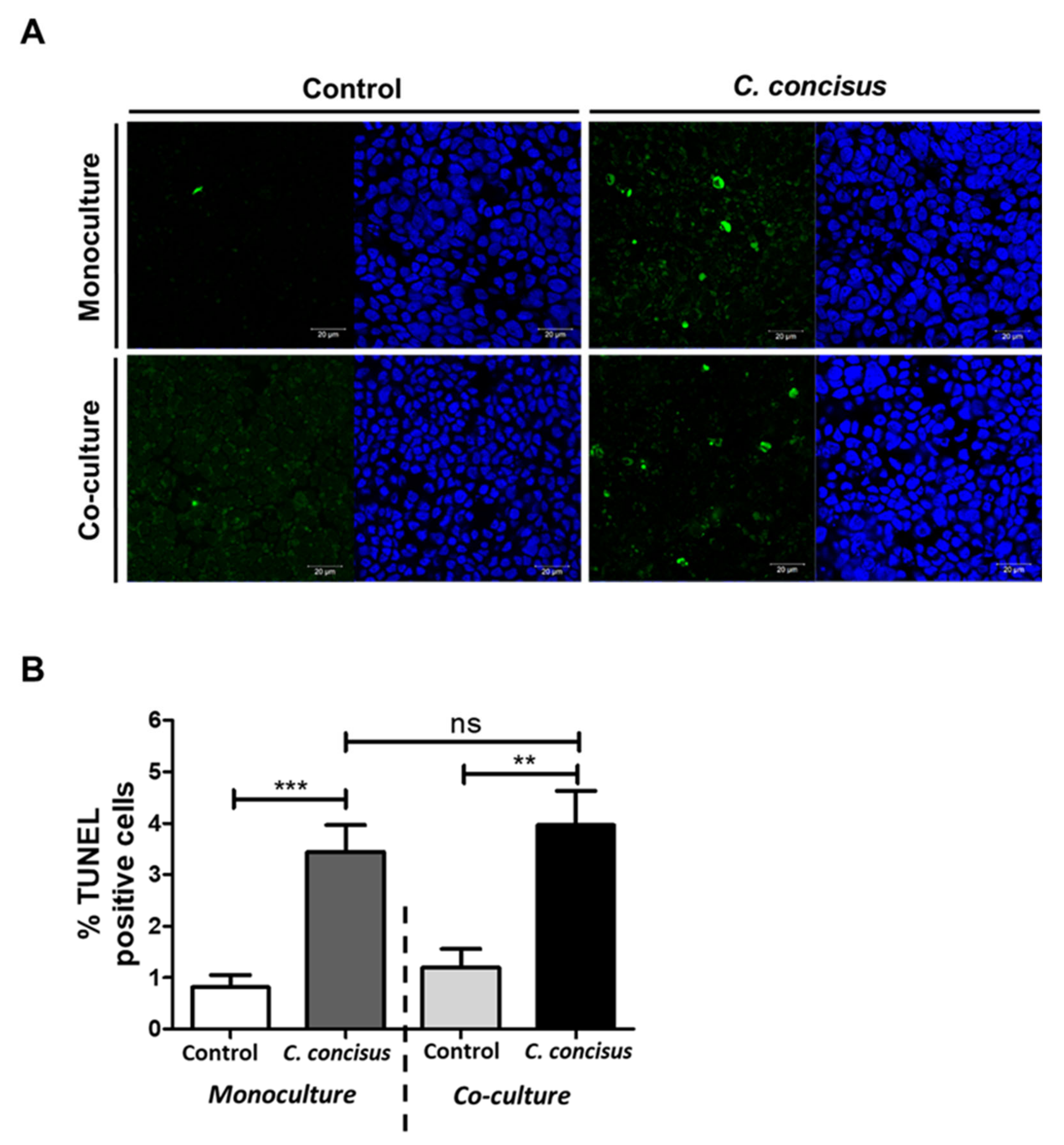
2.5. Epithelial Apoptosis in HT-29/B6-GR/MR Cells after Campylobacter concisus Infection
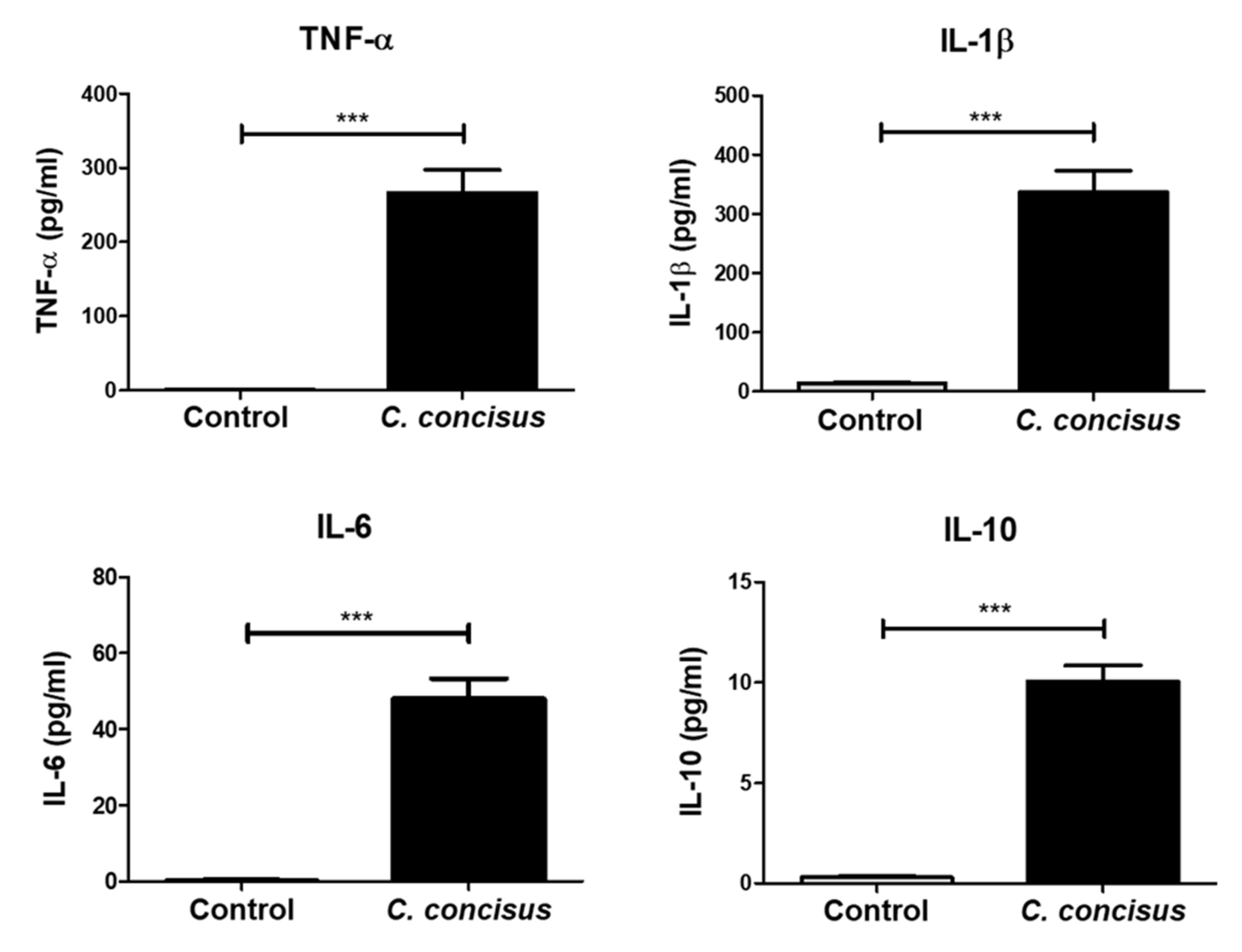
2.6. Inflammatory Response by M1-Macrophage-Like THP-1 Cells after Campylobacter concisus Infection in the Co-Culture Setting
2.7. Cell Viability in Colonic Epithelial Cell Monolayers after C. concisus Infection
3. Discussion
3.1. Immune-Mediated Aggravation of Campylobacter concisus-Induced Colonic Epithelial Barrier Dysfunction
3.2. Tight Junction Modifications in Colonic Epithelial Barrier Dysfunction Induced by Campylobacter concisus
3.2.1. Functional Loss of Occludin in Colonic Epithelial Cells in Co-culture with Macrophages after Campylobacter concisus Infection
3.2.2. Campylobacter concisus Induces Tricellulin Downregulation and Redistribution from Tricellular Tight Junctions of Colonic Epithelial Cells in Co-culture with Macrophages
3.2.3. The Role of Claudins in Campylobacter concisus-Induced Colonic Epithelial Barrier Dysfunction
3.3. Epithelial Apoptosis in the Regulation of Intestinal Epithelial Barrier Dysfunction Induced by Campylobacter concisus
4. Materials and Methods
4.1. Cell Culture
4.2. Transepithelial Electrical Resistance Measurements and Campylobacter concisus Infection
4.3. Western Blotting to Determine the Expression of Different Tight Junction Proteins
4.4. Immunofluorescence Staining and Confocal Laser-Scanning Microscopy to Determine Tight Junction Protein Localization
4.5. Epithelial Permeability Assay
4.6. Quantification of Epithelial Apoptosis
4.7. Cytometric Bead Array to Measure the Cytokine Release
4.8. Cell Viability Assay
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ATCC | American Type Culture Collection |
| BCA | Bicinchoninic acid |
| cAMP | Adenosine 3′,5′ -cyclic monophosphate |
| CBA | C Cytometric Bead Array |
| CCK-8 | Cell counting kit-8 |
| CD | Crohn’s Disease |
| CFU | Colony forming units |
| CLSM | Confocal laser scanning microscopy |
| CREB1 | cAMP responsive element binding protein 1 |
| CRP | C-Reactive Protein |
| CWD | Classical Whipple’s Disease |
| DAPI | 4′-6-diamidino-2-phenylindole dihydrochloride |
| DBA | DexaDexamethasone, butyrate-sodium salt, and aldosterone |
| DSS | Dextran sodium sulfate |
| EDTA | Ethylenediaminetetraacetic acid |
| EGTA | Ethylene glycol-bis(β-aminoethyl ether)-N,N,N′,N′-tetraacetic acid |
| ENaC | Epithelial sodium channel |
| EPEC | Enteropathogenic Escherichia coli |
| ERK | Extracellular signal-regulated kinases |
| FACS | Fluorescence-Activated Cell Sorting |
| FCS | Fetal calf serum |
| FITC | Fluorescein isothiocyanate |
| GAPDH | Glyceraldehyde 3-phosphate dehydrogenase |
| GR | Glucocorticoid receptor |
| HIV | Human immunodeficiency virus |
| IBD | Inflammatory bowel disease |
| IFN | Interferon |
| IL | Interleukin |
| LPS | Lipopolysaccharide |
| MOI | Multiplicity of infection |
| MR | Mineralocorticoid receptor |
| mRNA | messengerRNA |
| NF-κB | Nuclear factor kappa light chain enhancer of activated B cells |
| p.i. | Post infection |
| PBMC | Peripheral blood mononuclear cells |
| PBS | Phosphate buffered saline |
| PCF | Polycarbonate membrane filters |
| PFA | Paraformaldehyde |
| PMA | phorbol 12-myristate 13-acetate |
| PVDF | Polyvinylidene fluoride |
| PVP | Polyvinylpyrrolidone |
| RNA-Seq | RNA sequencing |
| RPMI | Rosewell Park Memorial Institute |
| RT | Room temperature |
| SDS | Sodium dodecyl sulfate |
| T3SS | Type-III secretion system |
| T6SS | Type-VI secretion system |
| TBS | Tris-buffered saline |
| TER | Transepithelial electrical resistance |
| TJ | Tight junction |
| tTJ | Tricellular TJ |
| TNF | Tumor necrosis factor |
| TUNEL | Terminal deoxynucleotidyl transferase dUTP nick end labeling |
| UC | Ulcerative colitis |
| ZO-1 | Zonula occludens protein-1 |
References
- Tanner, A.C.; Badger, S.; Lai, C.-H.; Listgarten, M.A.; Visconti, R.A.; Socransky, S.S. Wo Zinella gen. nov., WoZinella succinogenes (Vibrio succinogenes Wolin et al.) comb. nov., and Description of Bacteroides gracilis sp. nov., Wolinella recta sp. nov., Campylobacter concisus sp. nov., and Eikenella corrodens from Humans with Periodontal Disease. Int. J. Syst. Bacteriol. 1981, 31, 432–435. [Google Scholar]
- Lindblom, G.B.; Sjogren, E.; Hansson-Westerberg, J.; Kaijser, B. Campylobacter upsaliensis, C. sputorum sputorum and C. concisus as common causes of diarrhoea in Swedish children. Scand. J. Infect. Dis. 1995, 27, 187–188. [Google Scholar] [CrossRef]
- Aabenhus, R.; Permin, H.; On, S.L.; Andersen, L.P. Prevalence of Campylobacter concisus in diarrhoea of immunocompromised patients. Scand. J. Infect. Dis. 2002, 34, 248–252. [Google Scholar] [CrossRef]
- Nielsen, H.L.; Engberg, J.; Ejlertsen, T.; Bucker, R.; Nielsen, H. Short-term and medium-term clinical outcomes of Campylobacter concisus infection. Clin. Microbiol. Infect. 2012, 18, E459–E465. [Google Scholar] [CrossRef] [PubMed]
- Nattramilarasu, P.K.; Bücker, R.; Lobo de Sa, F.D.; Fromm, A.; Nagel, O.; Lee, I.M.; Butkevych, E.; Mousavi, S.; Genger, C.; Klove, S.; et al. Campylobacter concisus Impairs Sodium Absorption in Colonic Epithelium via ENaC Dysfunction and Claudin-8 Disruption. Int. J. Mol. Sci. 2020, 21, 373. [Google Scholar] [CrossRef] [PubMed]
- Barmeyer, C.; Erko, I.; Awad, K.; Fromm, A.; Bojarski, C.; Meissner, S.; Loddenkemper, C.; Kerick, M.; Siegmund, B.; Fromm, M.; et al. Epithelial barrier dysfunction in lymphocytic colitis through cytokine-dependent internalization of claudin-5 and -8. J. Gastroenterol. 2017, 52, 1090–1100. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, H.L.; Dalager-Pedersen, M.; Nielsen, H. High risk of microscopic colitis after Campylobacter concisus infection: Population-based cohort study. Gut 2020, 69, 1952–1958. [Google Scholar] [CrossRef]
- Yde Aagaard, M.E.; Frahm Kirk, K.; Linde Nielsen, H.; Harder Tarpgaard, I.; Bach Hansen, J.; Nielsen, H. Campylobacter concisus is prevalent in the gastrointestinal tract of patients with microscopic colitis. Scand. J. Gastroenterol. 2020, 55, 924–930. [Google Scholar] [CrossRef]
- Barmeyer, C.; Fromm, M.; Schulzke, J.D. Active and passive involvement of claudins in the pathophysiology of intestinal inflammatory diseases. Pflugers Arch. 2017, 469, 15–26. [Google Scholar] [CrossRef]
- Chiba, H.; Osanai, M.; Murata, M.; Kojima, T.; Sawada, N. Transmembrane proteins of tight junctions. Biochim. Biophys. Acta 2008, 1778, 588–600. [Google Scholar] [CrossRef] [PubMed]
- Otani, T.; Furuse, M. Tight Junction Structure and Function Revisited. Trends Cell Biol. 2020, 30, 805–817. [Google Scholar] [CrossRef]
- Tamura, A.; Hayashi, H.; Imasato, M.; Yamazaki, Y.; Hagiwara, A.; Wada, M.; Noda, T.; Watanabe, M.; Suzuki, Y.; Tsukita, S. Loss of claudin-15, but not claudin-2, causes Na+ deficiency and glucose malabsorption in mouse small intestine. Gastroenterology 2011, 140, 913–923. [Google Scholar] [CrossRef] [PubMed]
- Krug, S.M.; Gunzel, D.; Conrad, M.P.; Lee, I.F.; Amasheh, S.; Fromm, M.; Yu, A.S. Charge-selective claudin channels. Ann. N. Y. Acad. Sci. 2012, 1257, 20–28. [Google Scholar] [CrossRef]
- Krug, S.M.; Amasheh, S.; Richter, J.F.; Milatz, S.; Gunzel, D.; Westphal, J.K.; Huber, O.; Schulzke, J.D.; Fromm, M. Tricellulin forms a barrier to macromolecules in tricellular tight junctions without affecting ion permeability. Mol. Biol. Cell 2009, 20, 3713–3724. [Google Scholar] [CrossRef]
- Buschmann, M.M.; Shen, L.; Rajapakse, H.; Raleigh, D.R.; Wang, Y.; Wang, Y.; Lingaraju, A.; Zha, J.; Abbott, E.; McAuley, E.M.; et al. Occludin OCEL-domain interactions are required for maintenance and regulation of the tight junction barrier to macromolecular flux. Mol. Biol. Cell 2013, 24, 3056–3068. [Google Scholar] [CrossRef]
- Krug, S.M.; Schulzke, J.D.; Fromm, M. Tight junction, selective permeability, and related diseases. Semin. Cell Dev. Biol. 2014, 36, 166–176. [Google Scholar] [CrossRef]
- Staehlin, L.A. Further observations on the fine structure of freeze-cleaved tight junctions. J. Cell Sci. 1973, 13, 763–786. [Google Scholar]
- Walker, D.C.; MacKenzie, A.; Hulbert, W.C.; Hogg, J.C. A Re-assessment of the Tricellular Region of Epithelial Cell Tight Junctions in Trachea of Guinea Pig. Acta Anat. 1985, 122, 35–38. [Google Scholar] [CrossRef]
- Ikenouchi, J.; Furuse, M.; Furuse, K.; Sasaki, H.; Tsukita, S.; Tsukita, S. Tricellulin constitutes a novel barrier at tricellular contacts of epithelial cells. J. Cell Biol. 2005, 171, 939–945. [Google Scholar] [CrossRef] [PubMed]
- Ikenouchi, J.; Sasaki, H.; Tsukita, S.; Furuse, M.; Tsukita, S. Loss of occludin affects tricellular localization of tricellulin. Mol. Biol. Cell 2008, 19, 4687–4693. [Google Scholar] [CrossRef] [PubMed]
- Pabst, R. The anatomical basis for the immune function of the gut. Anat. Emryol. 1987, 176, 135–144. [Google Scholar] [CrossRef]
- Biancone, L.; Monteleone, I.; Blanco, G.D.V.; Vavassori, P.; Pallone, F. Redident bacterial flora and immune system. Digest. Liver Dis. 2002, 34, S37–S43. [Google Scholar] [CrossRef]
- Zigmond, E.; Varol, C.; Farache, J.; Elmaliah, E.; Satpathy, A.T.; Friedlander, G.; Mack, M.; Shpigel, N.; Boneca, I.G.; Murphy, K.M.; et al. Ly6C hi monocytes in the inflamed colon give rise to proinflammatory effector cells and migratory antigen-presenting cells. Immunity 2012, 37, 1076–1090. [Google Scholar] [CrossRef]
- Bain, C.C.; Scott, C.L.; Uronen-Hansson, H.; Gudjonsson, S.; Jansson, O.; Grip, O.; Guilliams, M.; Malissen, B.; Agace, W.W.; Mowat, A.M. Resident and pro-inflammatory macrophages in the colon represent alternative context-dependent fates of the same Ly6Chi monocyte precursors. Mucosal. Immunol. 2013, 6, 498–510. [Google Scholar] [CrossRef]
- Raduolovic, K.; Mak’Anyengo, R.; Kaya, B.; Steinert, A.; Niess, J.H. Injections of Lipopolysaccharide into Mice to Mimic Entrance of Microbial-derived Products After Intestinal Barrier Breach. J. Vis. Exp. 2018. [Google Scholar] [CrossRef]
- Shi, Y.J.; Gong, H.F.; Zhao, Q.Q.; Liu, X.S.; Liu, C.; Wang, H. Critical role of toll-like receptor 4 (TLR4) in dextran sulfate sodium (DSS)-Induced intestinal injury and repair. Toxicol. Lett. 2019, 315, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Lissner, D.; Schumann, M.; Batra, A.; Kredel, L.I.; Kuhl, A.A.; Erben, U.; May, C.; Schulzke, J.D.; Siegmund, B. Monocyte and M1 Macrophage-induced Barrier Defect Contributes to Chronic Intestinal Inflammation in IBD. Inflamm. Bowel Dis. 2015, 21, 1297–1305. [Google Scholar] [CrossRef] [PubMed]
- Man, S.M.; Kaakoush, N.O.; Leach, S.T.; Nahidi, L.; Lu, H.K.; Norman, J.; Day, A.S.; Zhang, L.; Mitchell, H.M. Host attachment, invasion, and stimulation of proinflammatory cytokines by Campylobacter concisus and other non-Campylobacter jejuni Campylobacter species. J. Infect. Dis. 2010, 202, 1855–1865. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, H.L.; Nielsen, H.; Ejlertsen, T.; Engberg, J.; Gunzel, D.; Zeitz, M.; Hering, N.A.; Fromm, M.; Schulzke, J.D.; Bücker, R. Oral and fecal Campylobacter concisus strains perturb barrier function by apoptosis induction in HT-29/B6 intestinal epithelial cells. PLoS ONE 2011, 6, e23858. [Google Scholar] [CrossRef]
- Man, S.M.; Zhang, L.; Day, A.S.; Leach, S.T.; Lemberg, D.A.; Mitchell, H. Campylobacter concisus and other Campylobacter species in children with newly diagnosed Crohn’s disease. Inflamm. Bowel Dis. 2010, 16, 1008–1016. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhya, I.; Thomson, J.M.; Hansen, R.; Berry, S.H.; El-Omar, E.M.; Hold, G.L. Detection of Campylobacter concisus and other Campylobacter species in colonic biopsies from adults with ulcerative colitis. PLoS ONE 2011, 6, e21490. [Google Scholar] [CrossRef]
- Mahendran, V.; Riordan, S.M.; Grimm, M.C.; Tran, T.A.; Major, J.; Kaakoush, N.O.; Mitchell, H.; Zhang, L. Prevalence of Campylobacter species in adult Crohn’s disease and the preferential colonization sites of Campylobacter species in the human intestine. PLoS ONE 2011, 6, e25417. [Google Scholar] [CrossRef]
- Castano-Rodriguez, N.; Kaakoush, N.O.; Lee, W.S.; Mitchell, H.M. Dual role of Helicobacter and Campylobacter species in IBD: A systematic review and meta-analysis. Gut 2017, 66, 235–249. [Google Scholar] [CrossRef]
- Ismail, Y.; Mahendran, V.; Octavia, S.; Day, A.S.; Riordan, S.M.; Grimm, M.C.; Lan, R.; Lemberg, D.; Tran, T.A.; Zhang, L. Investigation of the enteric pathogenic potential of oral Campylobacter concisus strains isolated from patients with inflammatory bowel disease. PLoS ONE 2012, 7, e38217. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, H.L.; Nielsen, H.; Torpdahl, M. Multilocus sequence typing of Campylobacter concisus from Danish diarrheic patients. Gut Pathog. 2016, 8, 44. [Google Scholar] [CrossRef][Green Version]
- Lobo de Sa, F.D.; Butkevych, E.; Nattramilarasu, P.K.; Fromm, A.; Mousavi, S.; Moos, V.; Golz, J.C.; Stingl, K.; Kittler, S.; Seinige, D.; et al. Curcumin Mitigates Immune-Induced Epithelial Barrier Dysfunction by Campylobacter jejuni. Int. J. Mol. Sci. 2019, 20, 4830. [Google Scholar] [CrossRef] [PubMed]
- Butkevych, E.; Lobo de Sa, F.D.; Nattramilarasu, P.K.; Bücker, R. Contribution of Epithelial Apoptosis and Subepithelial Immune Responses in Campylobacter jejuni-Induced Barrier Disruption. Front. Microbiol. 2020, 11, 344. [Google Scholar] [CrossRef] [PubMed]
- Bücker, R.; Schulz, E.; Gunzel, D.; Bojarski, C.; Lee, I.F.; John, L.J.; Wiegand, S.; Janssen, T.; Wieler, L.H.; Dobrindt, U.; et al. alpha-Haemolysin of Escherichia coli in IBD: A potentiator of inflammatory activity in the colon. Gut 2014, 63, 1893–1901. [Google Scholar] [CrossRef]
- Bücker, R.; Krug, S.M.; Moos, V.; Bojarski, C.; Schweiger, M.R.; Kerick, M.; Fromm, A.; Janssen, S.; Fromm, M.; Hering, N.A.; et al. Campylobacter jejuni impairs sodium transport and epithelial barrier function via cytokine release in human colon. Mucosal. Immunol. 2018, 11, 474–485. [Google Scholar] [CrossRef]
- Kirk, K.F.; Meric, G.; Nielsen, H.L.; Pascoe, B.; Sheppard, S.K.; Thorlacius-Ussing, O.; Nielsen, H. Molecular epidemiology and comparative genomics of Campylobacter concisus strains from saliva, faeces and gut mucosal biopsies in inflammatory bowel disease. Sci. Rep. 2018, 8, 1902. [Google Scholar] [CrossRef]
- Kaakoush, N.O.; Deshpande, N.P.; Wilkins, M.R.; Tan, C.G.; Burgos-Portugal, J.A.; Raftery, M.J.; Day, A.S.; Lemberg, D.A.; Mitchell, H. The pathogenic potential of Campylobacter concisus strains associated with chronic intestinal diseases. PLoS ONE 2011, 6, e29045. [Google Scholar] [CrossRef] [PubMed]
- Pulimood, A.B.; Ramakrishna, B.S.; Rita, A.B.; Srinivasan, P.; Mohan, V.; Gupta, S.; Perakath, B.; Kang, G.; Chandy, G.; Balasubramanian, K.A. Early activation of mucosal dendritic cells and macrophages in acute Campylobacter colitis and cholera: An in vivo study. J. Gastroenterol. Hepatol. 2008, 23, 752–758. [Google Scholar] [CrossRef]
- Heimesaat, M.M.; Genger, C.; Klove, S.; Weschka, D.; Mousavi, S.; Bereswill, S. The Host-Specific Intestinal Microbiota Composition Impacts Campylobacter coli Infection in a Clinical Mouse Model of Campylobacteriosis. Pathogens 2020, 9, 804. [Google Scholar] [CrossRef] [PubMed]
- Stockmann, M.; Schmitz, H.; Fromm, M.; Schmidt, W.; Pauli, G.; Scholz, P.; Riecken, E.O.; Schulzke, J.D. Mechanisms of Epithelial Barrier Impairment in HIV Infection. Ann. N. Y. Acad. Sci. 2000, 915, 293–303. [Google Scholar] [CrossRef]
- Shifflett, D.E.; Clayburgh, D.R.; Koutsouris, A.; Turner, J.R.; Hecht, G.A. Enteropathogenic, E. coli disrupts tight junction barrier function and structure in vivo. Lab. Investig. 2005, 85, 1308–1324. [Google Scholar] [CrossRef] [PubMed]
- Ou, G.; Rompikuntal, P.K.; Bitar, A.; Lindmark, B.; Vaitkevicius, K.; Wai, S.N.; Hammarstrom, M.L. Vibrio cholerae cytolysin causes an inflammatory response in human intestinal epithelial cells that is modulated by the PrtV protease. PLoS ONE 2009, 4, e7806. [Google Scholar] [CrossRef]
- Nickerson, C.A.; Goodwin, T.J.; Terlonge, J.; Ott, C.M.; Buchanan, K.L.; Uicker, W.C.; Emami, K.; LeBlanc, C.L.; Ramamurthy, R.; Clarke, M.S.; et al. Three-dimensional tissue assemblies: Novel models for the study of Salmonella enterica serovar Typhimurium pathogenesis. Infect. Immun. 2001, 69, 7106–7120. [Google Scholar] [CrossRef] [PubMed]
- Prasad, S.; Mingrino, R.; Kaukinen, K.; Hayes, K.L.; Powell, R.M.; MacDonald, T.T.; Collins, J.E. Inflammatory processes have differential effects on claudins 2, 3 and 4 in colonic epithelial cells. Lab. Investig. 2005, 85, 1139–1162. [Google Scholar] [CrossRef]
- Mankertz, J.; Amasheh, M.; Krug, S.M.; Fromm, A.; Amasheh, S.; Hillenbrand, B.; Tavalali, S.; Fromm, M.; Schulzke, J.D. TNFalpha up-regulates claudin-2 expression in epithelial HT-29/B6 cells via phosphatidylinositol-3-kinase signaling. Cell Tissue Res. 2009, 336, 67–77. [Google Scholar] [CrossRef]
- Zeissig, S.; Burgel, N.; Gunzel, D.; Richter, J.; Mankertz, J.; Wahnschaffe, U.; Kroesen, A.J.; Zeitz, M.; Fromm, M.; Schulzke, J.D. Changes in expression and distribution of claudin 2, 5 and 8 lead to discontinuous tight junctions and barrier dysfunction in active Crohn’s disease. Gut 2007, 56, 61–72. [Google Scholar] [CrossRef]
- Siegmund, B.; Lehr, H.A.; Fantuzzi, G.; Dinarello, C.A. IL-1 beta-converting enzyme (caspase-1) in intestinal inflammation. Proc. Natl. Acid. Sci. USA 2001, 98(23), 13249–13254. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.A.; Totemeyer, S.; Maskell, D.J.; Bryant, C.E.; Barrow, P.A. Induction of proinflammatory responses in the human monocytic cell line THP-1 by Campylobacter jejuni. Infect. Immun. 2003, 71, 2626–2633. [Google Scholar] [CrossRef]
- Sun, X.; Threadgill, D.; Jobin, C. Campylobacter jejuni induces colitis through activation of mammalian target of rapamycin signaling. Gastroenterology 2012, 142, 86–95.e85. [Google Scholar] [CrossRef] [PubMed]
- Kaakoush, N.O.; Deshpande, N.P.; Man, S.M.; Burgos-Portugal, J.A.; Khattak, F.A.; Raftery, M.J.; Wilkins, M.R.; Mitchell, H.M. Transcriptomic and proteomic analyses reveal key innate immune signatures in the host response to the gastrointestinal pathogen Campylobacter concisus. Infect. Immun. 2015, 83, 832–845. [Google Scholar] [CrossRef]
- Krause, P.; Morris, V.; Greenbaum, J.A.; Park, Y.; Bjoerheden, U.; Mikulski, Z.; Muffley, T.; Shui, J.W.; Kim, G.; Cheroutre, H.; et al. IL-10-producing intestinal macrophages prevent excessive antibacterial innate immunity by limiting IL-23 synthesis. Nat. Commun. 2015, 6, 7055. [Google Scholar] [CrossRef]
- Kaakoush, N.O.; Mitchell, H.M.; Man, S.M. Role of emerging Campylobacter species in inflammatory bowel diseases. Inflamm. Bowel. Dis. 2014, 20, 2189–2197. [Google Scholar] [CrossRef]
- Kim, K.Y.; Oh, T.W.; Do, H.J.; Yang, J.H.; Yang, I.J.; Jeon, Y.H.; Go, Y.H.; Ahn, S.C.; Ma, J.Y.; Park, K.I. Acer palmatum thumb. Ethanol Extract Alleviates Interleukin-6-Induced Barrier Dysfunction and Dextran Sodium Sulfate-Induced Colitis by Improving Intestinal Barrier Function and Reducing Inflammation. J. Immunol. Res. 2018, 2018, 5718396. [Google Scholar] [CrossRef]
- Guo, S.; Gillingham, T.; Guo, Y.; Meng, D.; Zhu, W.; Walker, W.A.; Ganguli, K. Secretions of Bifidobacterium infantis and Lactobacillus acidophilus Protect Intestinal Epithelial Barrier Function. J. Pediatr. Gastroenterol. Nutr. 2017, 64, 404–412. [Google Scholar] [CrossRef]
- Ogawa, M.; Osada, H.; Hasegawa, A.; Ohno, H.; Yanuma, N.; Sasaki, K.; Shimoda, M.; Shirai, J.; Kondo, H.; Ohmori, K. Effect of interleukin-1beta on occludin mRNA expression in the duodenal and colonic mucosa of dogs with inflammatory bowel disease. J. Vet. Intern. Med. 2018, 32, 1019–1025. [Google Scholar] [CrossRef]
- Amasheh, M.; Grotjohann, I.; Amasheh, S.; Fromm, A.; Soderholm, J.D.; Zeitz, M.; Fromm, M.; Schulzke, J.D. Regulation of mucosal structure and barrier function in rat colon exposed to tumor necrosis factor alpha and interferon gamma in vitro: A novel model for studying the pathomechanisms of inflammatory bowel disease cytokines. Scand. J. Gastroenterol. 2009, 44, 1226–1235. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Chirlaque, C.; Aranda, C.J.; Ocon, B.; Capitan-Canadas, F.; Ortega-Gonzalez, M.; Carrero, J.J.; Suarez, M.D.; Zarzuelo, A.; Sanchez de Medina, F.; Martinez-Augustin, O. Germ-free and Antibiotic-treated Mice are Highly Susceptible to Epithelial Injury in DSS Colitis. J. Crohns. Colitis. 2016, 10, 1324–1335. [Google Scholar] [CrossRef]
- Fukumatsu, M.; Ogawa, M.; Arakawa, S.; Suzuki, M.; Nakayama, K.; Shimizu, S.; Kim, M.; Mimuro, H.; Sasakawa, C. Shigella targets epithelial tricellular junctions and uses a noncanonical clathrin-dependent endocytic pathway to spread between cells. Cell Host Microbe. 2012, 11, 325–336. [Google Scholar] [CrossRef]
- Morampudi, V.; Graef, F.A.; Stahl, M.; Dalwadi, U.; Conlin, V.S.; Huang, T.; Vallance, B.A.; Yu, H.B.; Jacobson, K. Tricellular Tight Junction Protein Tricellulin Is Targeted by the Enteropathogenic Escherichia coli Effector EspG1, Leading to Epithelial Barrier Disruption. Infect. Immun. 2017, 85. [Google Scholar] [CrossRef]
- Gemmell, M.R.; Berry, S.; Mukhopadhya, I.; Hansen, R.; Nielsen, H.L.; Bajaj-Elliott, M.; Nielsen, H.; Hold, G.L. Comparative genomics of Campylobacter concisus: Analysis of clinical strains reveals genome diversity and pathogenic potential. Emerg. Microbes. Infect. 2018, 7, 116. [Google Scholar] [CrossRef]
- Liu, F.; Chen, S.; Luu, L.D.W.; Lee, S.A.; Tay, A.C.Y.; Wu, R.; Riordan, S.M.; Lan, R.; Liu, L.; Zhang, L. Analysis of complete Campylobacter concisus genomes identifies genomospecies features, secretion systems and novel plasmids and their association with severe ulcerative colitis. Microb. Genom. 2020, 6. [Google Scholar] [CrossRef]
- Krug, S.M.; Bojarski, C.; Fromm, A.; Lee, I.M.; Dames, P.; Richter, J.F.; Turner, J.R.; Fromm, M.; Schulzke, J.D. Tricellulin is regulated via interleukin-13-receptor alpha2, affects macromolecule uptake, and is decreased in ulcerative colitis. Mucosal. Immunol. 2018, 11, 345–356. [Google Scholar] [CrossRef]
- Keita, A.V.; Lindqvist, C.M.; Ost, A.; Magana, C.D.L.; Schoultz, I.; Halfvarson, J. Gut Barrier Dysfunction-A Primary Defect in Twins with Crohn’s Disease Predominantly Caused by Genetic Predisposition. J. Crohns. Colitis. 2018, 12, 1200–1209. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Epple, H.J.; Friebel, J.; Moos, V.; Troeger, H.; Krug, S.M.; Allers, K.; Schinnerling, K.; Fromm, A.; Siegmund, B.; Fromm, M.; et al. Architectural and functional alterations of the small intestinal mucosa in classical Whipple’s disease. Mucosal. Immunol. 2017, 10, 1542–1552. [Google Scholar] [CrossRef] [PubMed]
- Bucker, R.; Krug, S.M.; Fromm, A.; Nielsen, H.L.; Fromm, M.; Nielsen, H.; Schulzke, J.D. Campylobacter fetus impairs barrier function in HT-29/B6 cells through focal tight junction alterations and leaks. Ann. N. Y. Acad. Sci. 2017, 1405, 189–201. [Google Scholar] [CrossRef]
- Amasheh, M.; Fromm, A.; Krug, S.M.; Amasheh, S.; Andres, S.; Zeitz, M.; Fromm, M.; Schulzke, J.D. TNFalpha-induced and berberine-antagonized tight junction barrier impairment via tyrosine kinase, Akt and NFkappaB signaling. J. Cell Sci. 2010, 123, 4145–4155. [Google Scholar] [CrossRef] [PubMed]
- Bergmann, S.; von Buenau, B.; Vidal, Y.S.S.; Haftek, M.; Wladykowski, E.; Houdek, P.; Lezius, S.; Duplan, H.; Basler, K.; Dahnhardt-Pfeiffer, S.; et al. Claudin-1 decrease impacts epidermal barrier function in atopic dermatitis lesions dose-dependently. Sci. Rep. 2020, 10, 2024. [Google Scholar] [CrossRef] [PubMed]
- Weber, C.R.; Nalle, S.C.; Tretiakova, M.; Rubin, D.T.; Turner, J.R. Claudin-1 and claudin-2 expression is elevated in inflammatory bowel disease and may contribute to early neoplastic transformation. Lab. Investig. 2008, 88, 1110–1120. [Google Scholar] [CrossRef] [PubMed]
- Pope, J.L.; Ahmed, R.; Bhat, A.A.; Washington, M.K.; Singh, A.B.; Dhawan, P. Claudin-1 overexpression in intestinal epithelial cells enhances susceptibility to adenomatous polyposis coli-mediated colon tumorigenesis. Mol. Cancer 2014, 13, 167. [Google Scholar] [CrossRef] [PubMed]
- Troeger, H.; Epple, H.J.; Schneider, T.; Wahnschaffe, U.; Ullrich, R.; Burchard, G.D.; Jelinek, T.; Zeitz, M.; Fromm, M.; Schulzke, J.D. Effect of chronic Giardia lamblia infection on epithelial transport and barrier function in human duodenum. Gut 2007, 56, 328–335. [Google Scholar] [CrossRef] [PubMed]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nattramilarasu, P.K.; Lobo de Sá, F.D.; Schulzke, J.-D.; Bücker, R. Immune-Mediated Aggravation of the Campylobacter concisus-Induced Epithelial Barrier Dysfunction. Int. J. Mol. Sci. 2021, 22, 2043. https://doi.org/10.3390/ijms22042043
Nattramilarasu PK, Lobo de Sá FD, Schulzke J-D, Bücker R. Immune-Mediated Aggravation of the Campylobacter concisus-Induced Epithelial Barrier Dysfunction. International Journal of Molecular Sciences. 2021; 22(4):2043. https://doi.org/10.3390/ijms22042043
Chicago/Turabian StyleNattramilarasu, Praveen Kumar, Fábia Daniela Lobo de Sá, Jörg-Dieter Schulzke, and Roland Bücker. 2021. "Immune-Mediated Aggravation of the Campylobacter concisus-Induced Epithelial Barrier Dysfunction" International Journal of Molecular Sciences 22, no. 4: 2043. https://doi.org/10.3390/ijms22042043
APA StyleNattramilarasu, P. K., Lobo de Sá, F. D., Schulzke, J.-D., & Bücker, R. (2021). Immune-Mediated Aggravation of the Campylobacter concisus-Induced Epithelial Barrier Dysfunction. International Journal of Molecular Sciences, 22(4), 2043. https://doi.org/10.3390/ijms22042043