
Comprehensive Comparison of Amnion Stromal Cells and Chorion Stromal Cells by RNA-Seq

Abstract
1. Introduction
2. Results
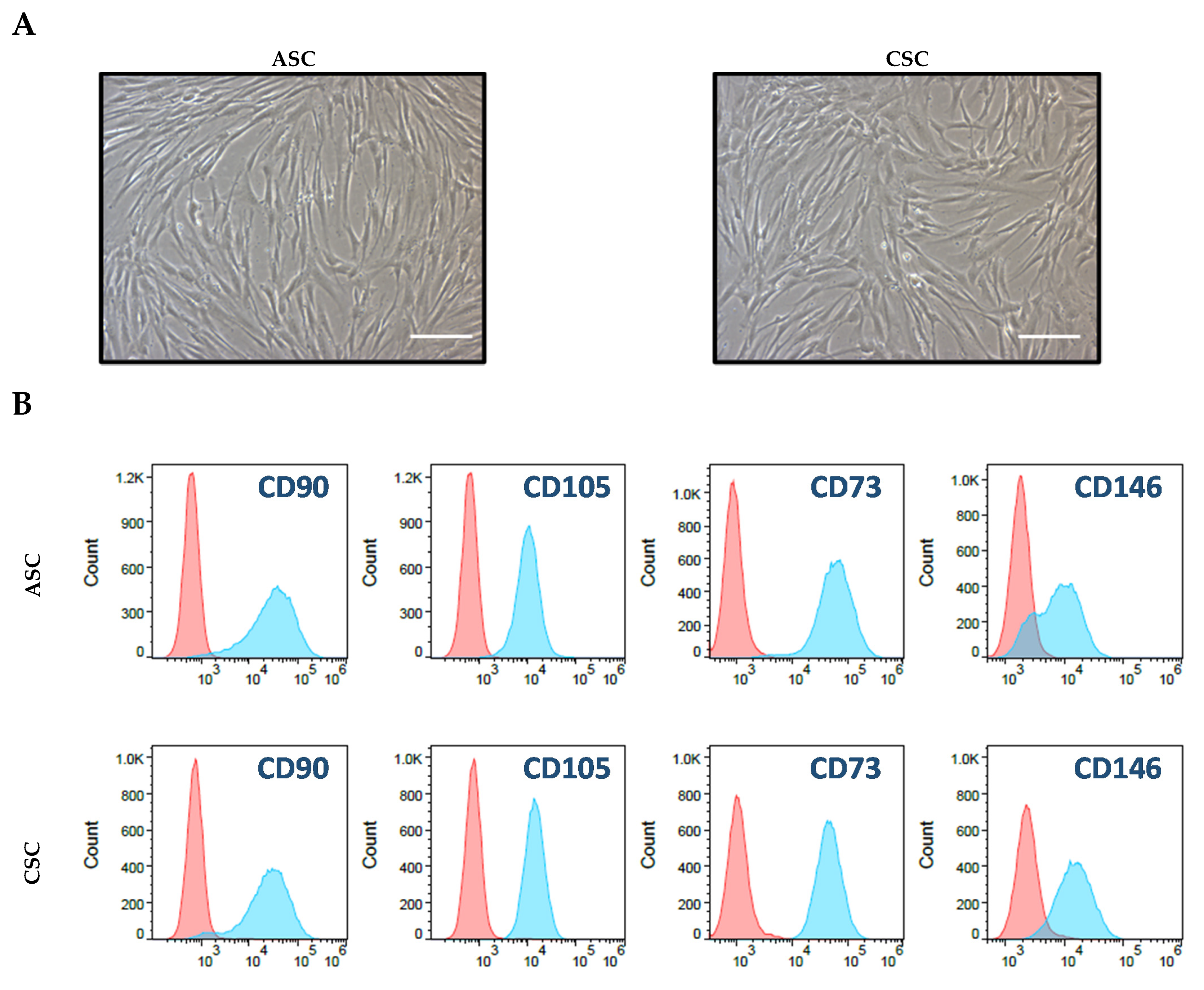
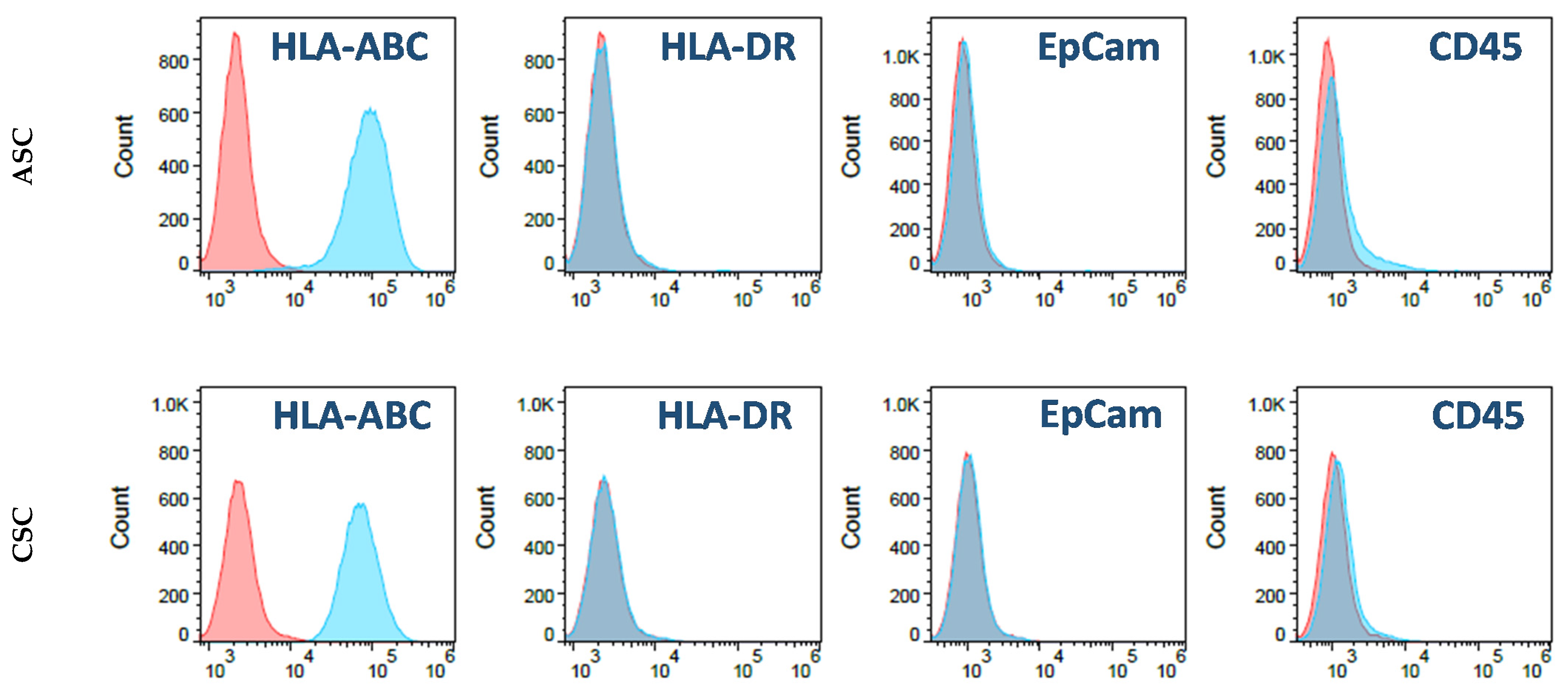
2.1. Characterization of ASCs and CSCs
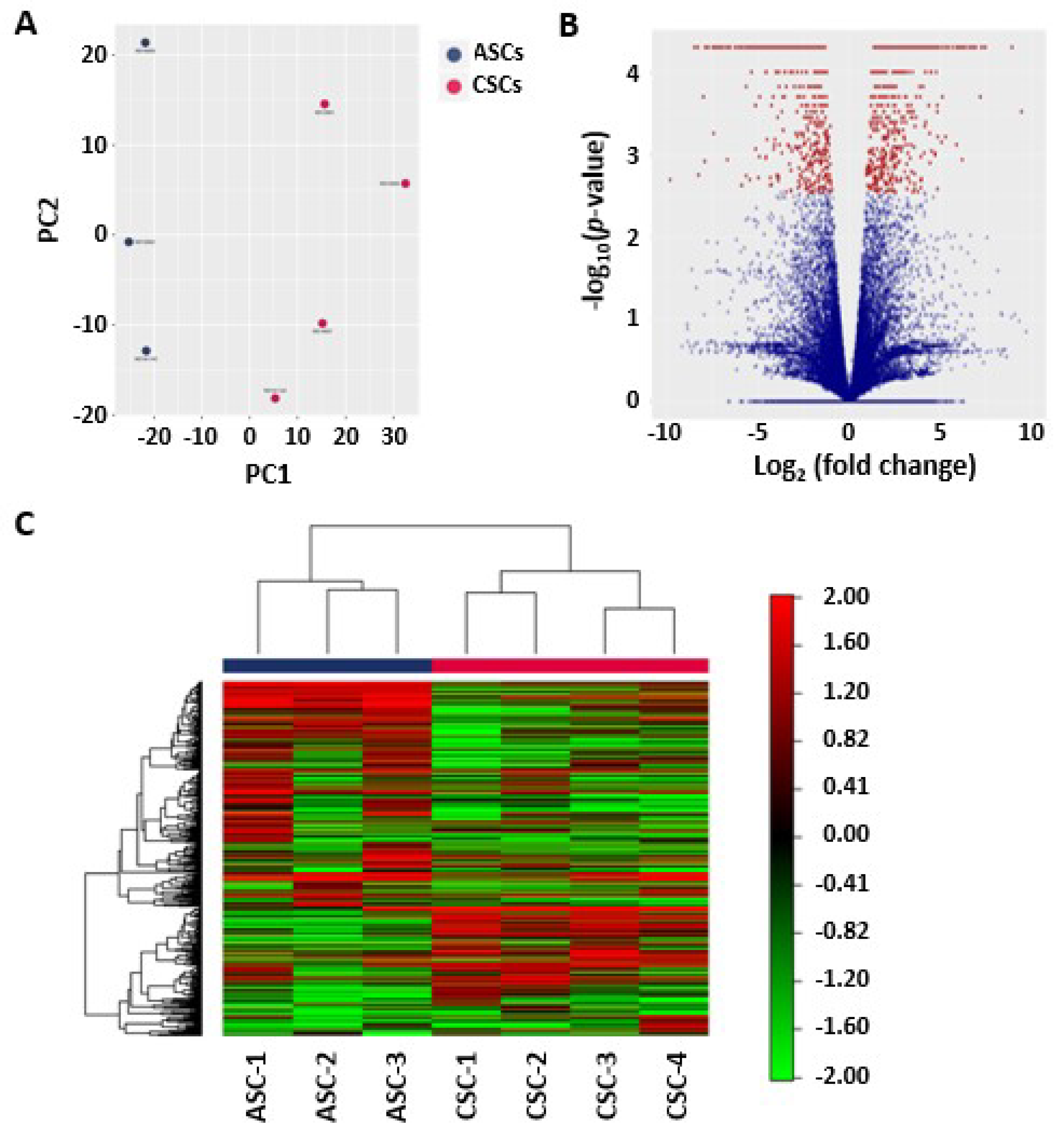
2.2. Principal Component Analysis of the ASCs and CSCs
2.3. Heat Map and Unsupervised Hierarchical Clustering
2.4. Differentially Expressed Genes and Novel Isoforms
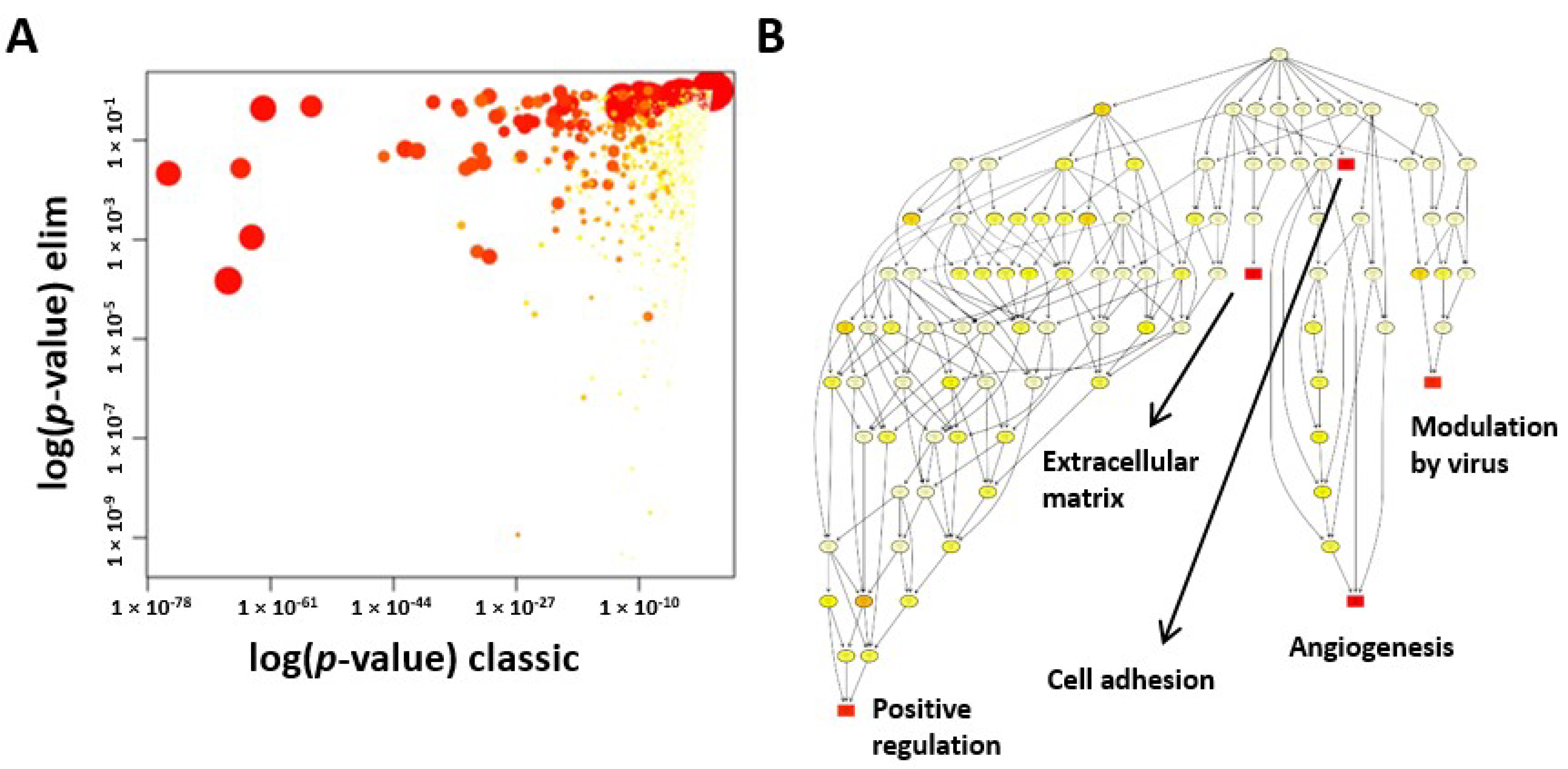
2.5. GO Enrichment Analysis
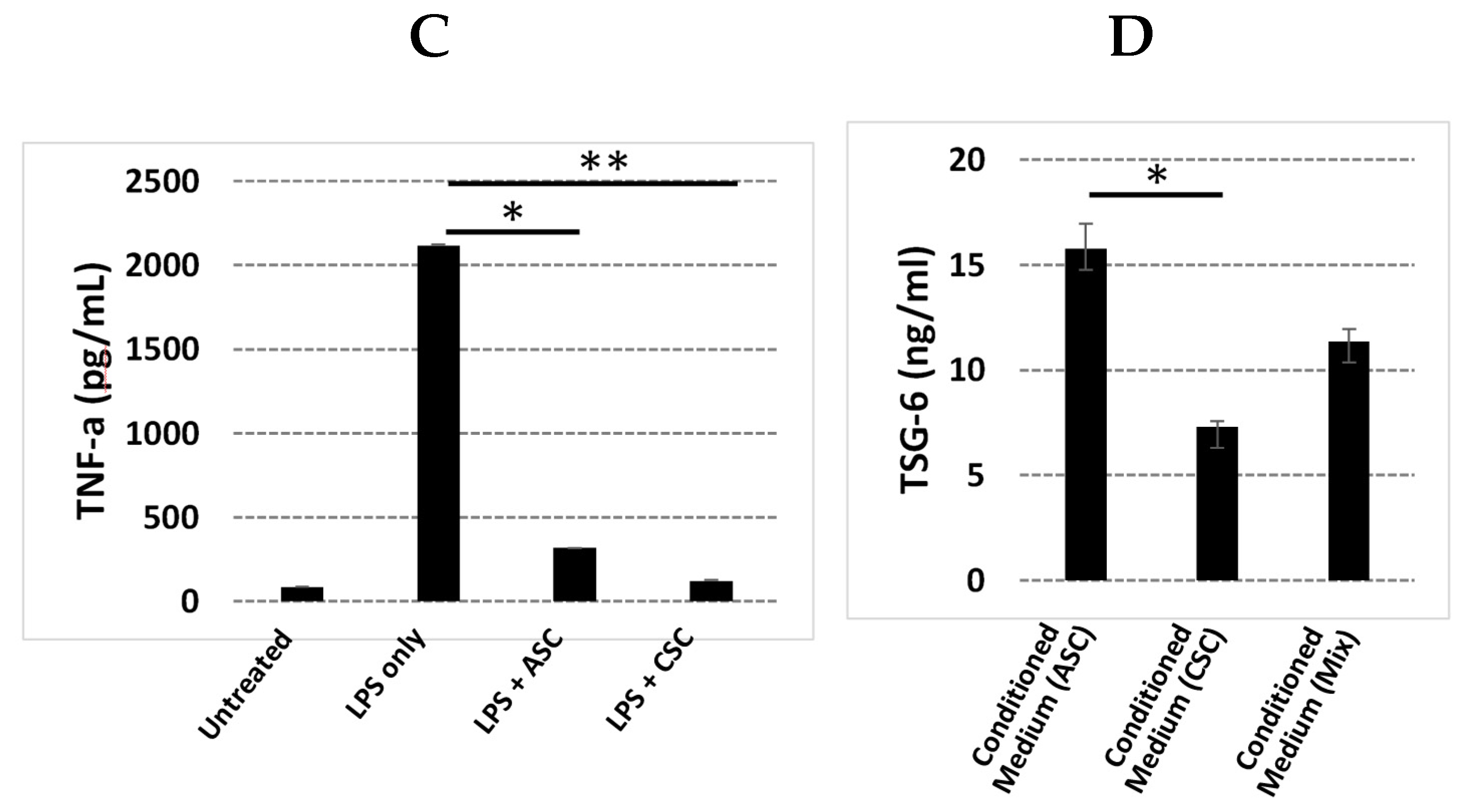
2.6. CSCs Secrete Significant Amounts of Angiogenic and Inflammatory Factors
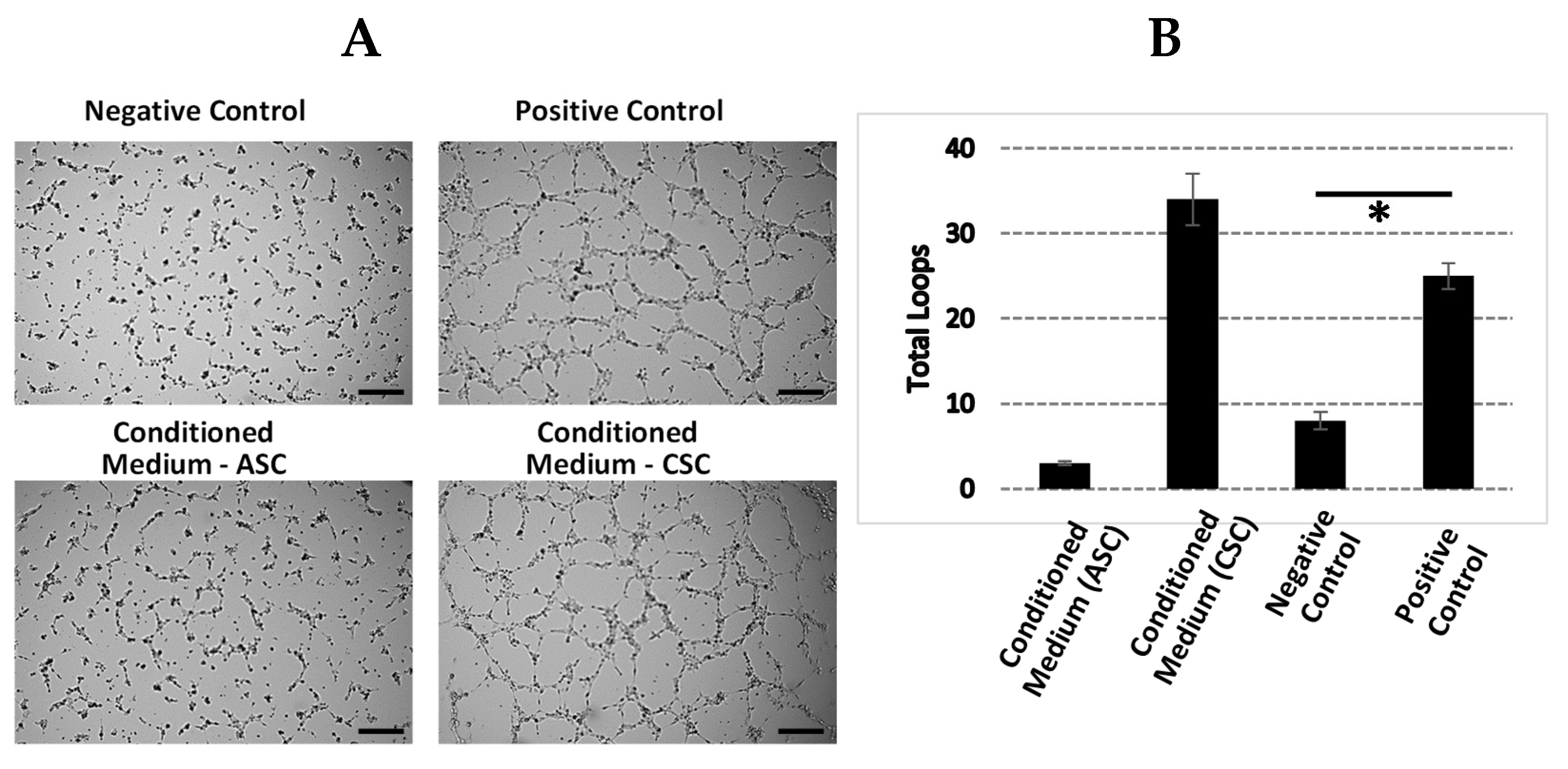
2.7. CSCs Exhibit Increased Angiogenic Function
2.8. ACSs Show Strong Anti-Inflammatory Function
3. Discussion
4. Materials and Methods
4.1. Tissue Procurement and Ethics Statement
4.2. Placental Tissue Processing and Isolation of the Amniotic Membrane and the Chorionic Membrane
4.3. Isolation of ASCs and CSCs from the Amniotic Membrane and Chorionic Membrane
4.4. Cell Culture and Characteristic Analysis through Flow Cytometry
4.5. RNA-Seq and Data Analysis
4.6. Characterization of Cytokine and Growth Factor Profiles in ASCs and CSCs
4.7. Secretion of Inflammatory Factors by Activated THP-1 cells
4.8. HUVEC Tube Formation Induced by ASC and CSC Cell Culture Supernatants
4.9. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bailo, M.; Soncini, M.; Vertua, E.; Signoroni, P.B.; Sanzone, S.; Lombardi, G.; Arienti, D.; Calamani, F.; Zatti, D.; Paul, P.; et al. Engraftment Potential of Human Amnion and Chorion Cells Derived from Term Placenta. Transplantation 2004, 78, 1439–1448. [Google Scholar] [CrossRef]
- Pogozhykh, O.; Prokopyuk, V.; Figueiredo, C.; Pogozhykh, D. Placenta and Placental Derivatives in Regenerative Therapies: Experimental Studies, History, and Prospects. Stem Cells Int. 2018, 2018, 4837930. [Google Scholar] [CrossRef]
- Dhall, S.; Coksaygan, T.; Hoffman, T.; Moorman, M.; Lerch, A.; Kuang, J.-Q.; Sathyamoorthy, M.; Danilkovitch, A. Viable cryopreserved umbilical tissue (vCUT) reduces post-operative adhesions in a rabbit abdominal adhesion model. Bioact. Mater. 2019, 4, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Massee, M.; Chinn, K.; Lei, J.; Lim, J.J.; Young, C.S.; Koob, T.J. Dehydrated human amnion/chorion membrane regulates stem cell activityin vitro. J. Biomed. Mater. Res. Part B Appl. Biomater. 2015, 104, 1495–1503. [Google Scholar] [CrossRef] [PubMed]
- Liang, L.; Li, Z.; Ma, T.; Han, Z.; Du, W.; Geng, J.; Jia, H.; Zhao, M.; Wang, J.; Zhang, B.; et al. Transplantation of Human Placenta-Derived Mesenchymal Stem Cells Alleviates Critical Limb Ischemia in Diabetic Nude Rats. Cell Transplant. 2017, 26, 45–61. [Google Scholar] [CrossRef] [PubMed]
- Mao, Y.; Singh-Varma, A.; Hoffman, T.; Dhall, S.; Danilkovitch, A.; Kohn, J. The Effect of Cryopreserved Human Placental Tissues on Biofilm Formation of Wound-Associated Pathogens. J. Funct. Biomater. 2018, 9, 3. [Google Scholar] [CrossRef] [PubMed]
- Dhall, S.; Hoffman, T.; Sathyamoorthy, M.; Lerch, A.; Jacob, V.; Moorman, M.; Kuang, J.-Q.; Danilkovitch, A. A Viable Lyopreserved Amniotic Membrane Modulates Diabetic Wound Microenvironment and Accelerates Wound Closure. Adv. Wound Care 2019, 8, 355–367. [Google Scholar] [CrossRef] [PubMed]
- Mao, Y.; Hoffman, T.; Dhall, S.; Singal, A.; Sathyamoorthy, M.; Danilkovitch, A.; Kohn, J. Endogenous viable cells in lyopreserved amnion retain differentiation potential and anti-fibrotic activity in vitro. Acta Biomater. 2019, 94, 330–339. [Google Scholar] [CrossRef] [PubMed]
- McQuilling, J.P.; Burnette, M.; Kimmerling, K.A.; Kammer, M.; Mowry, K.C. A mechanistic evaluation of the angiogenic properties of a dehydrated amnion chorion membrane in vitro and in vivo. Wound Repair Regen. 2019, 27, 609–621. [Google Scholar] [CrossRef]
- Mueller, S.M.; Navarini, A.A.; Itin, P.; Schwegler, S.; Laeuchli, S.; Goldust, M.; Ivanova, K. Pain reduction by dehydrated human amnion/chorion membrane allograft in nondiabetic leg ulcers might be an early indicator of good response: A case series. Dermatol. Ther. 2020, 33, e13587. [Google Scholar] [CrossRef] [PubMed]
- Basmaeil, Y.S.; Al Subayyil, A.M.; Khatlani, T.; Bahattab, E.; Al-Alwan, M.; Abomaray, F.M.; Kalionis, B.; AlShabibi, M.A.; Alaskar, A.S.; Abumaree, M.H. Human chorionic villous mesenchymal stem/stromal cells protect endothelial cells from injury induced by high level of glucose. Stem Cell Res. Ther. 2018, 9, 238. [Google Scholar] [CrossRef] [PubMed]
- Maacha, S.; Sidahmed, H.; Jacob, S.; Gentilcore, G.; Calzone, R.; Grivel, J.-C.; Cugno, C. Paracrine Mechanisms of Mesenchymal Stromal Cells in Angiogenesis. Stem Cells Int. 2020, 2020, 1–12. [Google Scholar] [CrossRef]
- Mathew, S.A.; Naik, C.; Cahill, P.A.; Bhonde, R.R. Placental mesenchymal stromal cells as an alternative tool for therapeutic angiogenesis. Cell. Mol. Life Sci. 2020, 77, 253–265. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Xu, Z.; Qu, G.; Wang, H.; Lin, L.; Li, X.; Xie, X.; Lei, Y.; He, X.; Chen, Y.; et al. Hypoxic Preconditioning Enhances the Efficacy of Mesenchymal Stem Cells-Derived Conditioned Medium in Switching Microglia toward Anti-inflammatory Polarization in Ischemia/Reperfusion. Cell. Mol. Neurobiol. 2020, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Zhan, X.-S.; El-Ashram, S.; Luo, D.-Z.; Luo, H.-N.; Wang, B.-Y.; Chen, S.-F.; Bai, Y.-S.; Chen, Z.-S.; Liu, C.-Y.; Ji, H.-Q. A Comparative Study of Biological Characteristics and Transcriptome Profiles of Mesenchymal Stem Cells from Different Canine Tissues. Int. J. Mol. Sci. 2019, 20, 1485. [Google Scholar] [CrossRef] [PubMed]
- Adolfsson, E.; Helenius, G.; Friberg, Ö.; Samano, N.; Frøbert, O.; Johansson, K. Bone marrow- and adipose tissue-derived mesenchymal stem cells from donors with coronary artery disease; growth, yield, gene expression and the effect of oxygen concentration. Scand. J. Clin. Lab. Investig. 2020, 80, 318–326. [Google Scholar] [CrossRef] [PubMed]
- Raynaud, C.M.; Maleki, M.; Lis, R.; Ahmed, B.; Al-Azwani, I.; Malek, J.; Safadi, F.F.; Rafii, A. Comprehensive Characterization of Mesenchymal Stem Cells from Human Placenta and Fetal Membrane and Their Response to Osteoactivin Stimulation. Stem Cells Int. 2012, 2012, 1–13. [Google Scholar] [CrossRef]
- González, P.L.; Carvajal, C.; Cuenca, J.; Alcayaga-Miranda, F.; Figueroa, F.E.; Bartolucci, J.; Salazar-Aravena, L.; Khoury, M. Chorion Mesenchymal Stem Cells Show Superior Differentiation, Immunosuppressive, and Angiogenic Potentials in Comparison with Haploidentical Maternal Placental Cells. Stem Cells Transl. Med. 2015, 4, 1109–1121. [Google Scholar] [CrossRef] [PubMed]
- Du, W.; Li, X.; Chi, Y.; Ma, F.; Li, Z.; Yang, S.; Song, B.; Cui, J.; Ma, T.; Li, J.; et al. VCAM-1+ placenta chorionic villi-derived mesenchymal stem cells display potent pro-angiogenic activity. Stem Cell Res. Ther. 2016, 7, 1–13. [Google Scholar] [CrossRef]
- Ferreira, M.S.V.; Bienert, M.; Müller, K.; Rath, B.; Goecke, T.; Opländer, C.; Braunschweig, T.; Mela, P.; Brümmendorf, T.H.; Beier, F.; et al. Comprehensive characterization of chorionic villi-derived mesenchymal stromal cells from human placenta. Stem Cell Res. Ther. 2018, 9, 28. [Google Scholar] [CrossRef]
- De La Torre, P.; Pérez-Lorenzo, M.J.; Flores, A.I. Human Placenta-Derived Mesenchymal Stromal Cells: A Review from Basic Research to Clinical Applications. Stromal Cells Struct. Funct. Ther. Implic. 2019. [Google Scholar]
- Hao, D.; He, C.; Ma, B.; Lankford, L.; Reynaga, L.; Farmer, D.L.; Guo, F.; Wang, A. Hypoxic Preconditioning Enhances Survival and Proangiogenic Capacity of Human First Trimester Chorionic Villus-Derived Mesenchymal Stem Cells for Fetal Tissue Engineering. Stem Cells Int. 2019, 2019, 9695239-12. [Google Scholar] [CrossRef]
- Heo, J.S.; Choi, Y.; Kim, H.-S.; Kim, H.O. Comparison of molecular profiles of human mesenchymal stem cells derived from bone marrow, umbilical cord blood, placenta and adipose tissue. Int. J. Mol. Med. 2016, 37, 115–125. [Google Scholar] [CrossRef]
- Xu, Y.; Guo, S.; Wei, C.; Li, H.; Chen, L.; Yin, C.; Zhang, C. The Comparison of Adipose Stem Cell and Placental Stem Cell in Secretion Characteristics and in Facial Antiaging. Stem Cells Int. 2016, 2016, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Bačenková, D.; Rosocha, J.; Tóthová, T.; Rosocha, L.; Šarišský, M. Isolation and basic characterization of human term amnion and chorion mesenchymal stromal cells. Cytotherapy 2011, 13, 1047–1056. [Google Scholar] [CrossRef]
- Huang, Q.; Yang, Y.; Luo, C.; Wen, Y.; Liu, R.; Li, S.; Chen, T.; Sun, H.; Tang, L. An efficient protocol to generate placental chorionic plate-derived mesenchymal stem cells with superior proliferative and immunomodulatory properties. Stem Cell Res. Ther. 2019, 10, 301. [Google Scholar] [CrossRef] [PubMed]
- Shakouri-Motlagh, A.; O’Connor, A.J.; Kalionis, B.; Heath, D.E. Improved ex vivo expansion of mesenchymal stem cells on solubilized acellular fetal membranes. J. Biomed. Mater. Res. Part A 2018, 107, 232–242. [Google Scholar] [CrossRef] [PubMed]
- Shilina, M.A.; Silachev, D.N.; Gorunov, K.V.; Kozhukharova, I.V.; Pugovkina, N.A.; Lyublinskaya, O.G.; Ivanova, Y.S.; Nikolsky, N.N.; Grinchuk, T.M. Characterization of Cultured Mesenchymal Stromal Cells Established from Human Chorion. Cell Tissue Biol. 2020, 14, 196–201. [Google Scholar] [CrossRef]
- Soncini, M.; Vertua, E.; Gibelli, L.; Zorzi, F.; Denegri, M.; Albertini, A.; Wengler, G.S.; Parolini, O. Isolation and characterization of mesenchymal cells from human fetal membranes. J. Tissue Eng. Regen. Med. 2007, 1, 296–305. [Google Scholar] [CrossRef]
- Choi, Y.S.; Park, Y.-B.; Ha, C.-W.; A Kim, J.; Heo, J.-C.; Han, W.-J.; Oh, S.-Y.; Choi, S.-J. Different characteristics of mesenchymal stem cells isolated from different layers of full term placenta. PLoS ONE 2017, 12, e0172642. [Google Scholar] [CrossRef] [PubMed]
- Yamahara, K.; Harada, K.; Ohshima, M.; Ishikane, S.; Ohnishi, S.; Tsuda, H.; Otani, K.; Taguchi, A.; Soma, T.; Ogawa, H.; et al. Comparison of Angiogenic, Cytoprotective, and Immunosuppressive Properties of Human Amnion- and Chorion-Derived Mesenchymal Stem Cells. PLoS ONE 2014, 9, e88319. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Zhang, R.; Zou, Q.; Chen, Y.; Zhou, M.; Li, X.; Ran, R.; Chen, Q. Comparison of the Biological Characteristics of Mesenchymal Stem Cells Derived from the Human Placenta and Umbilical Cord. Sci. Rep. 2018, 8, 1–9. [Google Scholar] [CrossRef]
- Kim, M.J.; Shin, K.S.; Jeon, J.H.; Lee, D.R.; Shim, S.H.; Kim, J.K.; Cha, D.-H.; Yoon, T.K.; Kim, G.J. Human chorionic-plate-derived mesenchymal stem cells and Wharton’s jelly-derived mesenchymal stem cells: A comparative analysis of their potential as placenta-derived stem cells. Cell Tissue Res. 2011, 346, 53–64. [Google Scholar] [CrossRef]
- Kannaiyan, J.; Veeramanikandan, V.; Muthukumarasamy, E.; Paulraj, B. An in-vitro study of Amniotic membrane, Villous chorion and Wharton’s jelly-derived Mesenchymal stem cells and their potential for cardiac repair. J. Stem Cell Res. Ther. 2018, 4, 1. [Google Scholar] [CrossRef]
- Ding, L.; Rath, E.; Bai, Y. Comparison of Alternative Splicing Junction Detection Tools Using RNA-Seq Data. Curr. Genom. 2017, 18, 268–277. [Google Scholar] [CrossRef] [PubMed]
- Sood, R.; Zehnder, J.L.; Druzin, M.L.; Brown, P.O. Gene expression patterns in human placenta. Proc. Natl. Acad. Sci. USA 2006, 103, 5478–5483. [Google Scholar] [CrossRef]
- Cheng, Y.; Hou, T.; Ping, J.; Chen, T.; Yin, B. LMO3 promotes hepatocellular carcinoma invasion, metastasis and anoikis inhibition by directly interacting with LATS1 and suppressing Hippo signaling. J. Exp. Clin. Cancer Res. 2018, 37, 228. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.-S.; Jiang, N.-N.; Zhou, Y.; Yu, K.-Y.; Gong, H.-Y.; Liao, G.-J. LMO3 promotes gastric cancer cell invasion and proliferation through Akt-mTOR and Akt-GSK3β signaling. Int. J. Mol. Med. 2018, 41, 2755–2763. [Google Scholar] [CrossRef]
- Dupain, C.; Gracia, C.; Harttrampf, A.C.; Rivière, J.; Geoerger, B.; Massaad-Massade, L. Newly identified LMO3-BORCS5 fusion oncogene in Ewing sarcoma at relapse is a driver of tumor progression. Oncogene 2019, 38, 7200–7215. [Google Scholar] [CrossRef] [PubMed]
- Tran, N.; Broun, A.; Ge, K. Lysine Demethylase KDM6A in Differentiation, Development, and Cancer. Mol. Cell. Biol. 2020, 40. [Google Scholar] [CrossRef]
- Pang, A.L.; Clark, J.; Chan, W.-Y.; Rennert, O.M. Expression of humanNAA11(ARD1B) gene is tissue-specific and is regulated by DNA methylation. Epigenetics 2011, 6, 1391–1399. [Google Scholar] [CrossRef]
- Barry, S.N.; Crow, K.D. The role of HoxA11 and HoxA13 in the evolution of novel fin morphologies in a representative batoid (Leucoraja erinacea). EvoDevo 2017, 8, 24. [Google Scholar] [CrossRef]
- Bradaschia-Correa, V.; Leclerc, K.; Josephson, A.M.; Lee, S.; Palma, L.; Litwa, H.P.; Neibart, S.S.; Huo, J.C.; Leucht, P. Hox gene expression determines cell fate of adult periosteal stem/progenitor cells. Sci. Rep. 2019, 9, 5043. [Google Scholar] [CrossRef] [PubMed]
- Desanlis, I.; Kherdjemil, Y.; Mayran, A.; Bouklouch, Y.; Gentile, C.; Sheth, R.; Zeller, R.; Drouin, J.; Kmita, M. HOX13-dependent chromatin accessibility underlies the transition towards the digit development program. Nat. Commun. 2020, 11, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Mattick, J.S.; Rinn, J.L. Discovery and annotation of long noncoding RNAs. Nat. Struct. Mol. Biol. 2015, 22, 5–7. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Gadad, S.S.; Kim, D.-S.; Kraus, W.L. Discovery, Annotation, and Functional Analysis of Long Noncoding RNAs Controlling Cell-Cycle Gene Expression and Proliferation in Breast Cancer Cells. Mol. Cell 2015, 59, 698–711. [Google Scholar] [CrossRef] [PubMed]
- Mungunsukh, O.; McCart, E.A.; Day, R.M. Hepatocyte Growth Factor Isoforms in Tissue Repair, Cancer, and Fibrotic Remodeling. Biomedicines 2014, 2, 301–326. [Google Scholar] [CrossRef] [PubMed]
- Dhall, S.; Sathyamoorthy, M.; Kuang, J.-Q.; Hoffman, T.; Moorman, M.; Lerch, A.; Jacob, V.; Sinclair, S.M.; Danilkovitch, A. Properties of viable lyopreserved amnion are equivalent to viable cryopreserved amnion with the convenience of ambient storage. PLoS ONE 2018, 13, e0204060. [Google Scholar] [CrossRef] [PubMed]
- Johnson, A.; Gyurdieva, A.; Dhall, S.; Danilkovitch, A.; Duan-Arnold, Y. Understanding the Impact of Preservation Methods on the Integrity and Functionality of Placental Allografts. Ann. Plast. Surg. 2017, 79, 203–213. [Google Scholar] [CrossRef]
- Jacob, V.; Johnson, N.; Lerch, M.A.; Jones, M.B.; Dhall, S.; Sathyamoorthy, M.; Danilkovitch, A. Structural and Functional Equivalency Between Lyopreserved and Cryopreserved Chorions with Viable Cells. Adv. Wound Care 2020, 9, 502–515. [Google Scholar] [CrossRef]
- Robinson, J.T.; Thorvaldsdóttir, H.; Winckler, W.; Guttman, M.; Lander, E.S.; Getz, G.; Mesirov, J.P. Integrative genomics viewer. Nat. Biotechnol. 2011, 29, 24–26. [Google Scholar] [CrossRef] [PubMed]
- Thorvaldsdóttir, H.; Robinson, J.T.; Mesirov, J.P. Integrative Genomics Viewer (IGV): High-performance genomics data visualization and exploration. Brief. Bioinform. 2013, 14, 178–192. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Pachter, L.; Salzberg, S.L. TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; Van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Roberts, A.; A Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef]
- Trapnell, C.; Hendrickson, D.G.; Sauvageau, M.; A Goff, L.; Rinn, J.L.; Pachter, L. Differential analysis of gene regulation at transcript resolution with RNA-seq. Nat. Biotechnol. 2013, 31, 46–53. [Google Scholar] [CrossRef]
- Hansen, K.D.; Brenner, S.E.; Dudoit, S. Biases in Illumina transcriptome sequencing caused by random hexamer priming. Nucleic Acids Res. 2010, 38, e131. [Google Scholar] [CrossRef]
- Roberts, A.; Pimentel, H.; Trapnell, C.; Pachter, L. Identification of novel transcripts in annotated genomes using RNA-Seq. Bioinformatics 2011, 27, 2325–2329. [Google Scholar] [CrossRef]
- Roberts, A.; Trapnell, C.; Donaghey, J.; Rinn, J.L.; Pachter, L. Improving RNA-Seq expression estimates by correcting for fragment bias. Genome Biol. 2011, 12, R22. [Google Scholar] [CrossRef]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | CSCs FPKM | ASCs FPKM | Log2 Fold Change | q-Value |
|---|---|---|---|---|
| NAA11 | 3.47 | 0 | −9.82 | 0.038 |
| LMO3 | 0.02 | 11.54 | 9.44 | 0.009 |
| UTY | 0.01 | 4.46 | 8.93 | 0.002 |
| BIRC7 | 97.61 | 0.28 | −8.47 | 0.002 |
| APOD | 34.72 | 0.11 | −8.28 | 0.002 |
| HOXA13 | 2.87 | 0.01 | −8.24 | 0.034 |
| MKRN4P | 5.69 | 0.02 | −7.98 | 0.006 |
| HOXA11 | 13.17 | 0.06 | −7.89 | 0.025 |
| RGPD1 | 0.77 | 0 | −7.87 | 0.002 |
| NR2F1 | 24.25 | 0.1 | −7.87 | 0.002 |
| Gene | CSCs FPKM | ASCs FPKM | Log2 Fold Change | q-Value |
|---|---|---|---|---|
| CXCL6 | 1.28 | 223.96 | 7.45 | 0.021 |
| XLOC_040015 | 0.08 | 9.7 | 6.95 | 0.021 |
| FPXF1-AS1 | 17.86 | 0.17 | −6.71 | 0.021 |
| HGF | 308.31 | 3.13 | −6.62 | 0.021 |
| MAEL | 17.59 | 0.2 | −6.43 | 0.021 |
| NPTX1 | 39.61 | 0.47 | −6.39 | 0.021 |
| HGF | 1110.15 | 1.7 | −6.02 | 0.021 |
| ATP2A3 | 34.04 | 0.75 | −5.5 | 0.021 |
| DPT | 3.26 | 136.88 | 5.39 | 0.021 |
| POSTN | 3.39 | 125.22 | 5.21 | 0.021 |
| Closest Known Transcript | CSCs FPKM | ASCs FPKM | Log2 Fold Change | q-Value |
|---|---|---|---|---|
| HGF | 110.15 | 1.7 | −6.02 | 0.021 |
| RP11 | 2.11 | 0.09 | −4.52 | 0.036 |
| MICAL2 | 14.4 | 0.95 | −3.93 | 0.021 |
| KRT7 | 54.11 | 7.08 | −2.93 | 0.021 |
| PERP | 17.19 | 116.26 | 2.76 | 0.021 |
| ACRC | 0.74 | 4.95 | 2.75 | 0.021 |
| PLBD1 | 14.14 | 2.39 | −2.57 | 0.021 |
| IL1B | 52.28 | 246.72 | 2.24 | 0.021 |
| MEST | 43.64 | 10.09 | −2.11 | 0.021 |
| CSF1 | 66.89 | 15.8 | −2.08 | 0.021 |
| GO Term | Annoteted | Significant | Exprected | p-Value |
|---|---|---|---|---|
| Extracellular Matrix Organization | 322 | 47 | 17.75 | 3.80 × 10−10 |
| Angiogenesis | 387 | 57 | 21.33 | 4.80 × 10−10 |
| Cell Adhesion | 1199 | 151 | 66.09 | 1.10 × 10−9 |
| Positive Regulation of Transcription | 777 | 76 | 42.83 | 3.20 × 10−9 |
| Modulation by Virus | 336 | 18 | 18.52 | 4.70 × 10−9 |
| Negative Regulation of Cell Proliferation | 563 | 62 | 31.03 | 2.50 × 10−8 |
| Homophilic Cell Adhesion | 105 | 15 | 5.79 | 2.80 × 10−8 |
| Postive Regulation of Protein Kinase B Signaling | 72 | 13 | 3.97 | 1.50 × 10−7 |
| SRP-dependent Cotranslational Protein | 317 | 9 | 17.47 | 2.10 × 10−7 |
| Nuclear-transcribed mRNA Catabolic Process | 520 | 62 | 28.66 | 4.00 × 10−7 |
| Conditioned Medium (pg/μg) | Cell Lysate (pg/105 cells) | |||
|---|---|---|---|---|
| Name | ASCs | CSCs | ASCs | CSCs |
| IFNα2 | 2.26 | 1.64 | 0.04 | 0.02 |
| IFNβ | 143.02 | 92.45 | 2.52 | 1.54 |
| IFNγ | 14.85 | 11.21 | 0.3 | 0.18 |
| IL-2 | 22.92 | 13.66 | 0.51 | 0.27 |
| sIL-6Rα | 1662.59 | 1202.55 | 31.29 | 16.06 |
| IL-8 | --- | 670.22 | 6.02 | 3.4 |
| IL-10 | 9.6 | 8.06 | 0.14 | 0.11 |
| IL-11 | --- | 180.26 | 1.29 | 0.18 |
| IL-12 (p40) | 149.74 | 104.04 | 2.51 | 1.17 |
| IL-19 | 14.23 | 11.22 | 1.11 | 0.6 |
| IL-20 | 9.49 | 5.72 | 0.15 | 0.09 |
| IL-22 | 50.76 | 20.73 | 0.32 | 0.18 |
| IL-26 | 478.9 | 295.39 | 8.41 | 6.07 |
| IL-27 (p28) | 102.45 | 65.27 | 2.03 | 1.13 |
| IL-28A (IFNγ2) | 4.87 | 2.69 | 0.07 | 0.03 |
| IL-29/IFNγ1) | 50.11 | 31.01 | 1.25 | 0.61 |
| IL-32 | 125.38 | 132.85 | 1.89 | 1.32 |
| IL-34 | 299.74 | 145.86 | 5.98 | 3.34 |
| IL-35 | 256.36 | 181.68 | 2.68 | 0.94 |
| LIGHT/TNFSF14 | 22.25 | 8.97 | 0.15 | 0.03 |
| MMP-1 | 67.65 | 56.42 | 0.97 | 0.63 |
| MMP-2 | 731.38 | 443.49 | 46.75 | 30.95 |
| MMP-3 | 926.57 | 674.78 | 17.08 | 11.39 |
| sTNF-R1 | 25.78 | 33.69 | 0.41 | 0.36 |
| sTNF-R2 | 49.91 | 32.98 | 0.74 | 0.55 |
| TSLP | 13.16 | 10.46 | 0.24 | 0.15 |
| TWEAK/TNFSF12 | 3.6 | 2.07 | 0.08 | 0.05 |
| PGE2 | 252.84 | 239.12 | -- | -- |
| Angiogenin | -- | 30.51 | -- | -- |
| FGF-2 | -- | 98.91 | -- | -- |
| VEGFA | -- | 98.91 | -- | -- |
| Protein Name | Amnion Tissue | Chorion Tissue |
|---|---|---|
| IL-1RA | **** | **** |
| IL-4 | * | ** |
| IL-10 | * | ** |
| bFGF | *** | **** |
| Angiogenin | *** | *** |
| Angiopoietin-1 | ** | ** |
| Angiopoietin-2 | ** | *** |
| EGF | ** | ** |
| HGF | **** | **** |
| SDF-1a | ** | *** |
| IGFBP-1 | **** | ***** |
| PDGF-AA | * | ** |
| PDGF-BB | ** | *** |
| PIGF | * | * |
| VEGF-A | * | *** |
| VEGF-D | * | ** |
| TIMP-1 | **** | **** |
| Periostin | *** | *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jones, B.; Li, C.; Park, M.S.; Lerch, A.; Jacob, V.; Johnson, N.; Kuang, J.-Q.; Dhall, S.; Sathyamoorthy, M. Comprehensive Comparison of Amnion Stromal Cells and Chorion Stromal Cells by RNA-Seq. Int. J. Mol. Sci. 2021, 22, 1901. https://doi.org/10.3390/ijms22041901
Jones B, Li C, Park MS, Lerch A, Jacob V, Johnson N, Kuang J-Q, Dhall S, Sathyamoorthy M. Comprehensive Comparison of Amnion Stromal Cells and Chorion Stromal Cells by RNA-Seq. International Journal of Molecular Sciences. 2021; 22(4):1901. https://doi.org/10.3390/ijms22041901
Chicago/Turabian StyleJones, Brielle, Chaoyang Li, Min Sung Park, Anne Lerch, Vimal Jacob, Nicholas Johnson, Jin-Qiang Kuang, Sandeep Dhall, and Malathi Sathyamoorthy. 2021. "Comprehensive Comparison of Amnion Stromal Cells and Chorion Stromal Cells by RNA-Seq" International Journal of Molecular Sciences 22, no. 4: 1901. https://doi.org/10.3390/ijms22041901
APA StyleJones, B., Li, C., Park, M. S., Lerch, A., Jacob, V., Johnson, N., Kuang, J.-Q., Dhall, S., & Sathyamoorthy, M. (2021). Comprehensive Comparison of Amnion Stromal Cells and Chorion Stromal Cells by RNA-Seq. International Journal of Molecular Sciences, 22(4), 1901. https://doi.org/10.3390/ijms22041901