
Systematic Analysis and Biochemical Characterization of the Caffeoyl Shikimate Esterase Gene Family in Poplar

 , ,
, ,
Abstract
:1. Introduction
2. Results
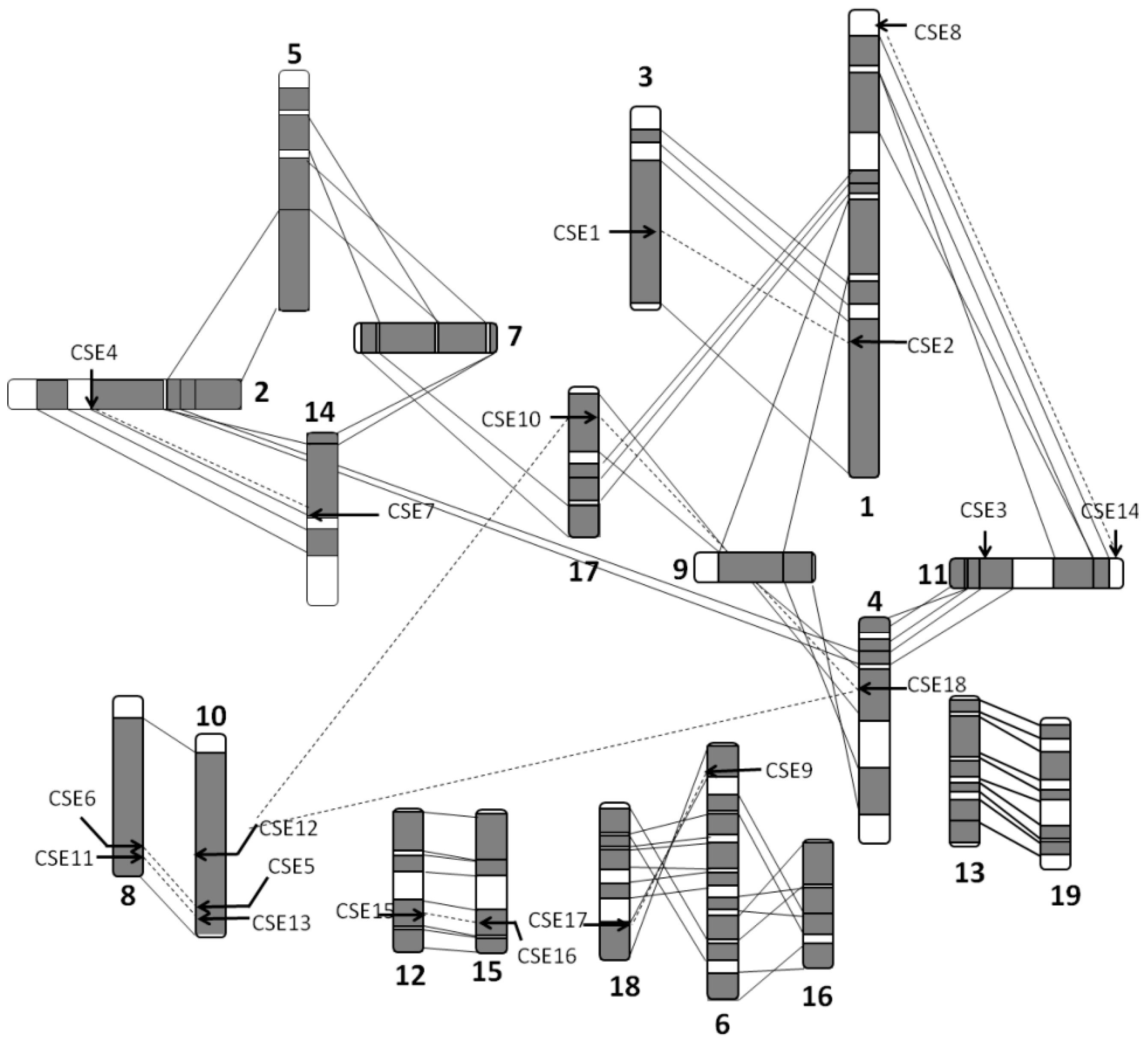
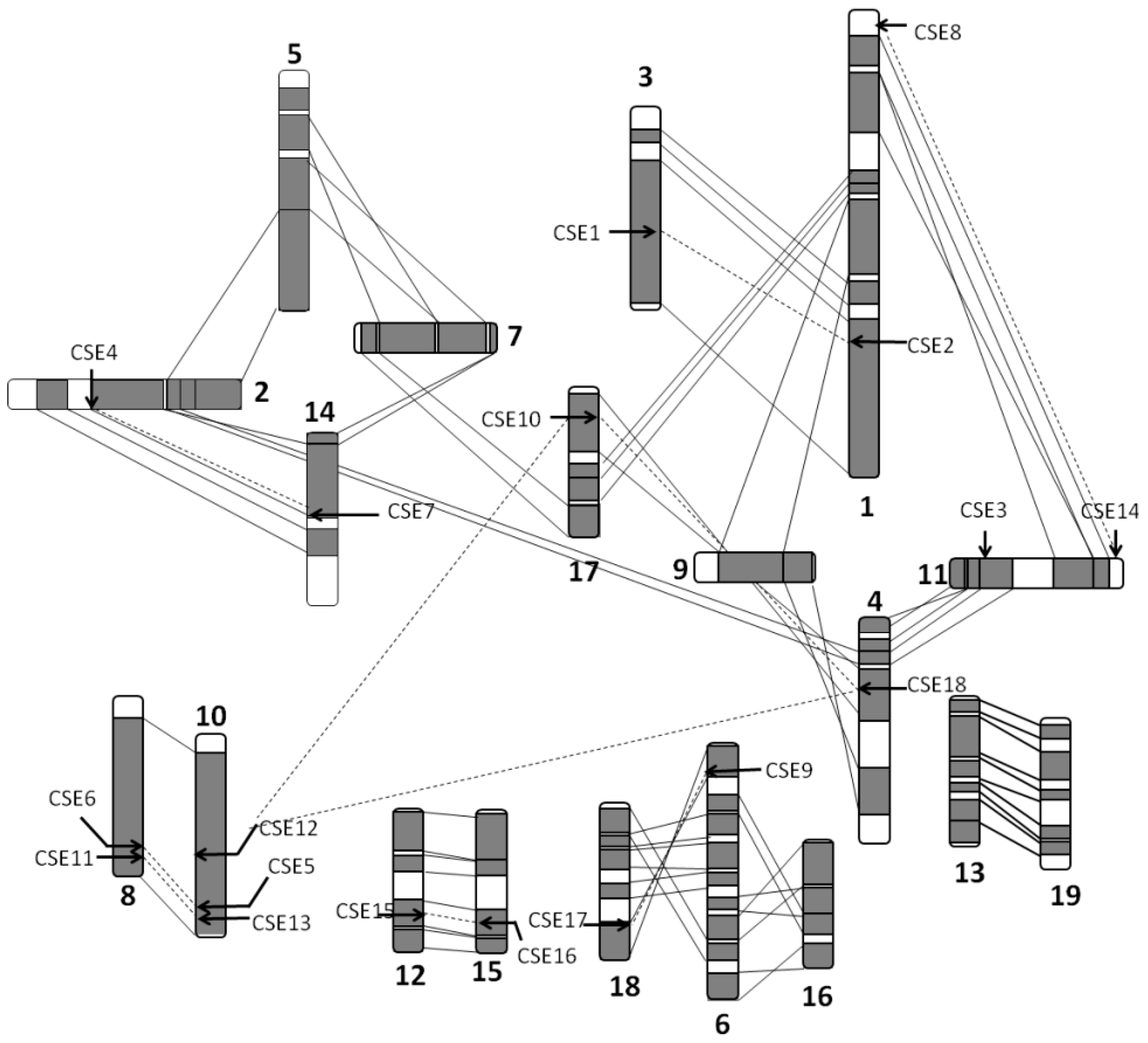
2.1. Organization and Distribution of the CSE Gene Family
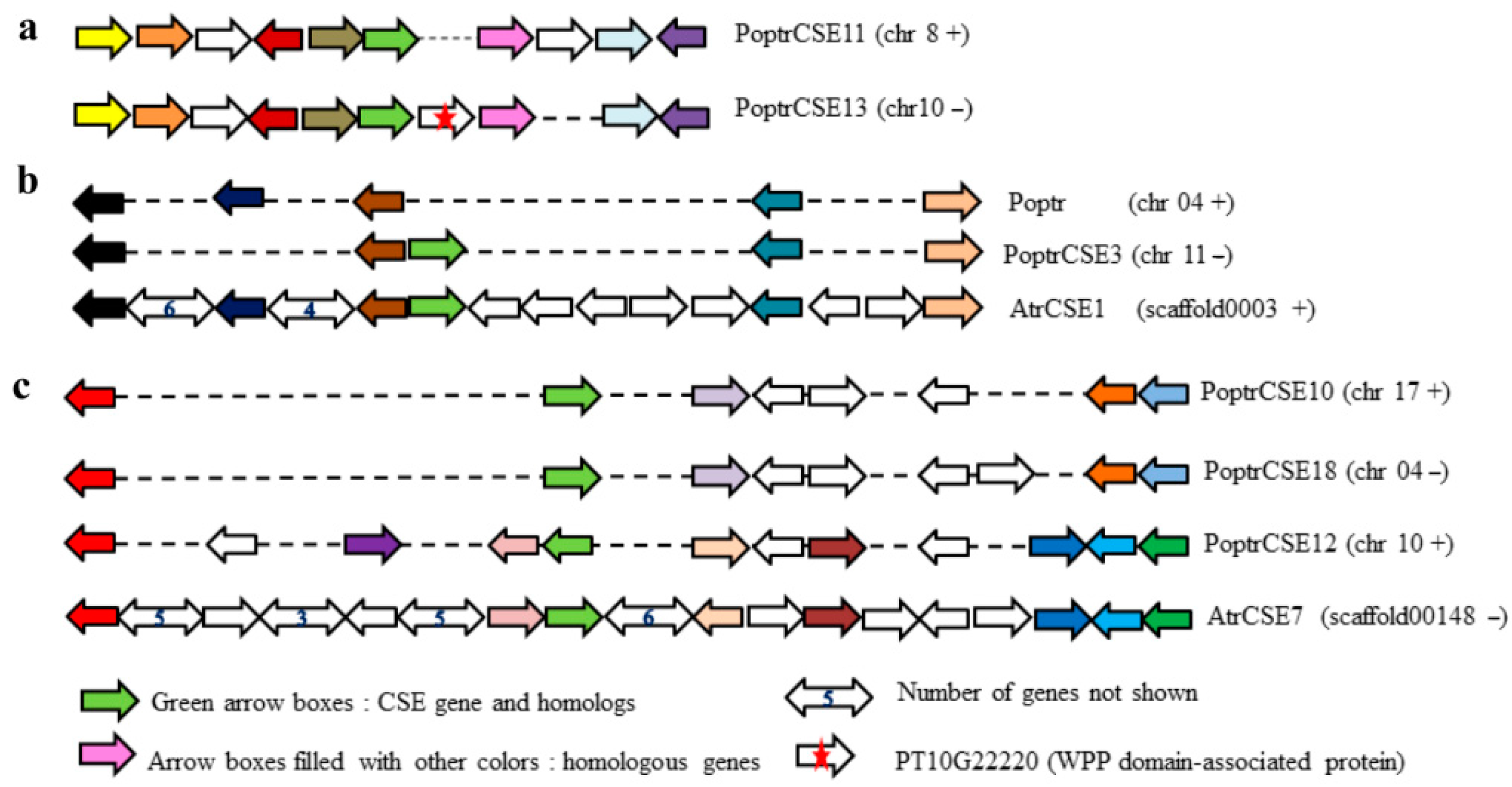
2.2. Promoter Sequence Analysis
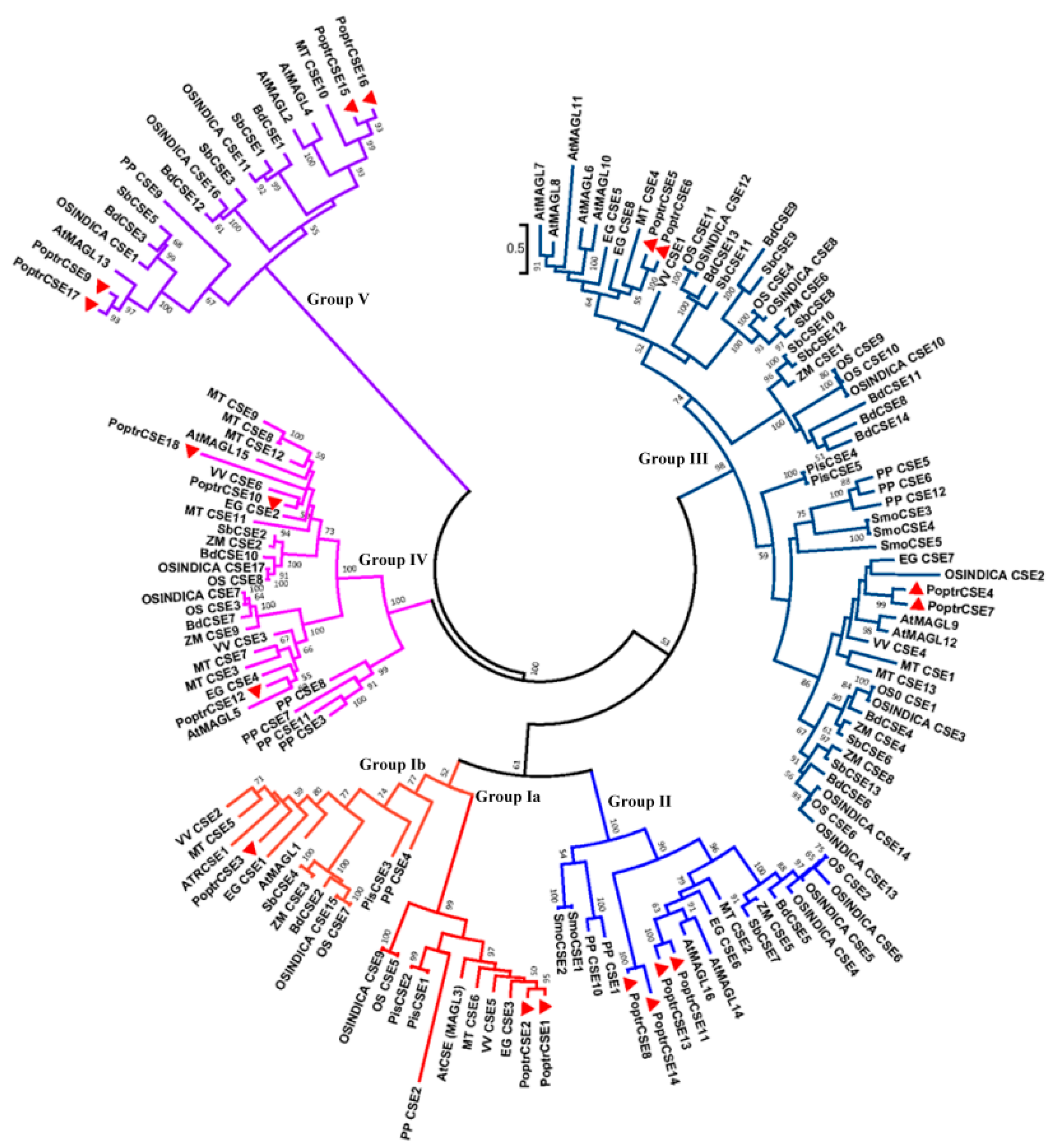
2.3. Alignment and Phylogenetic Analysis of CSE and CSE-like Genes
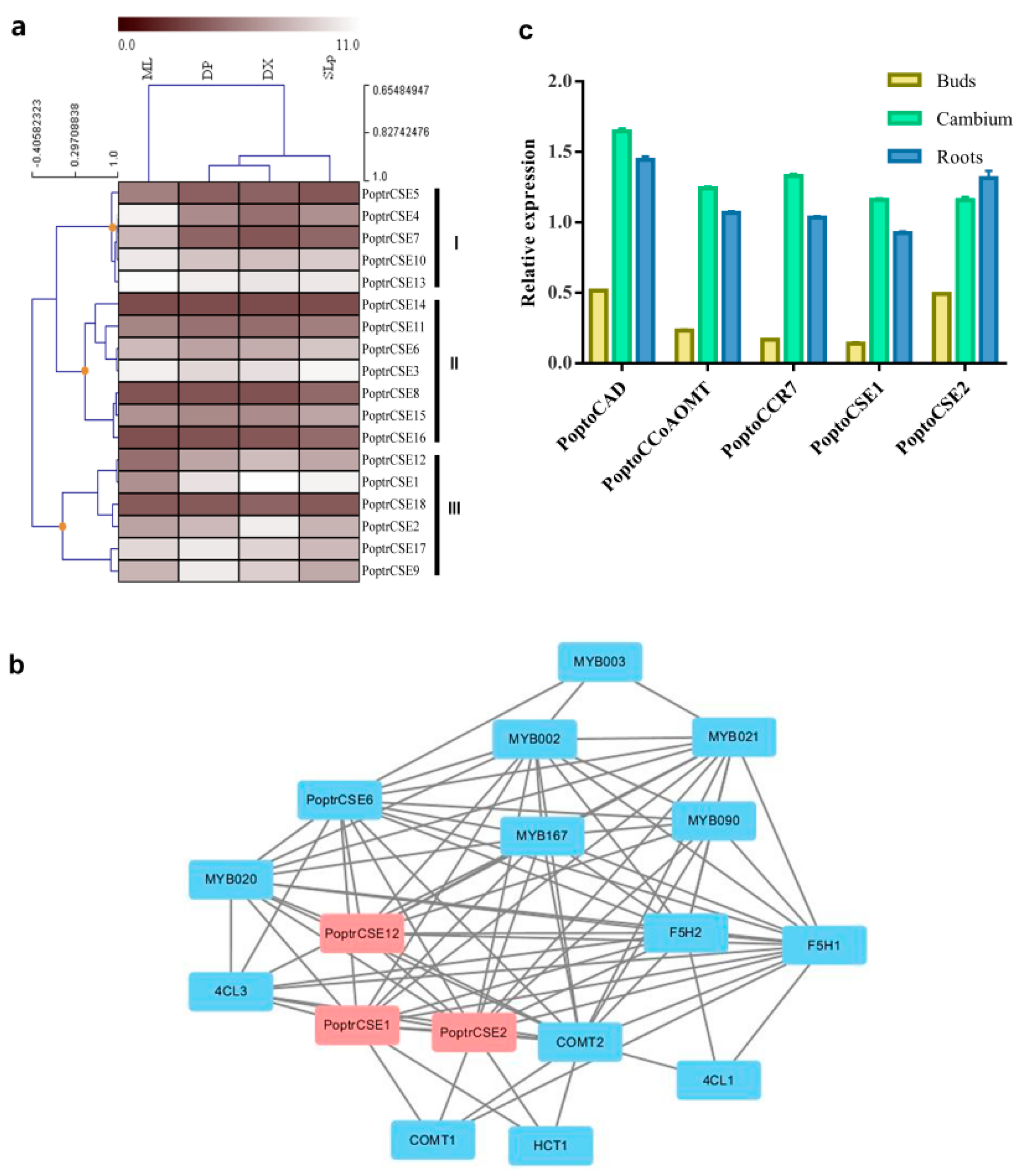
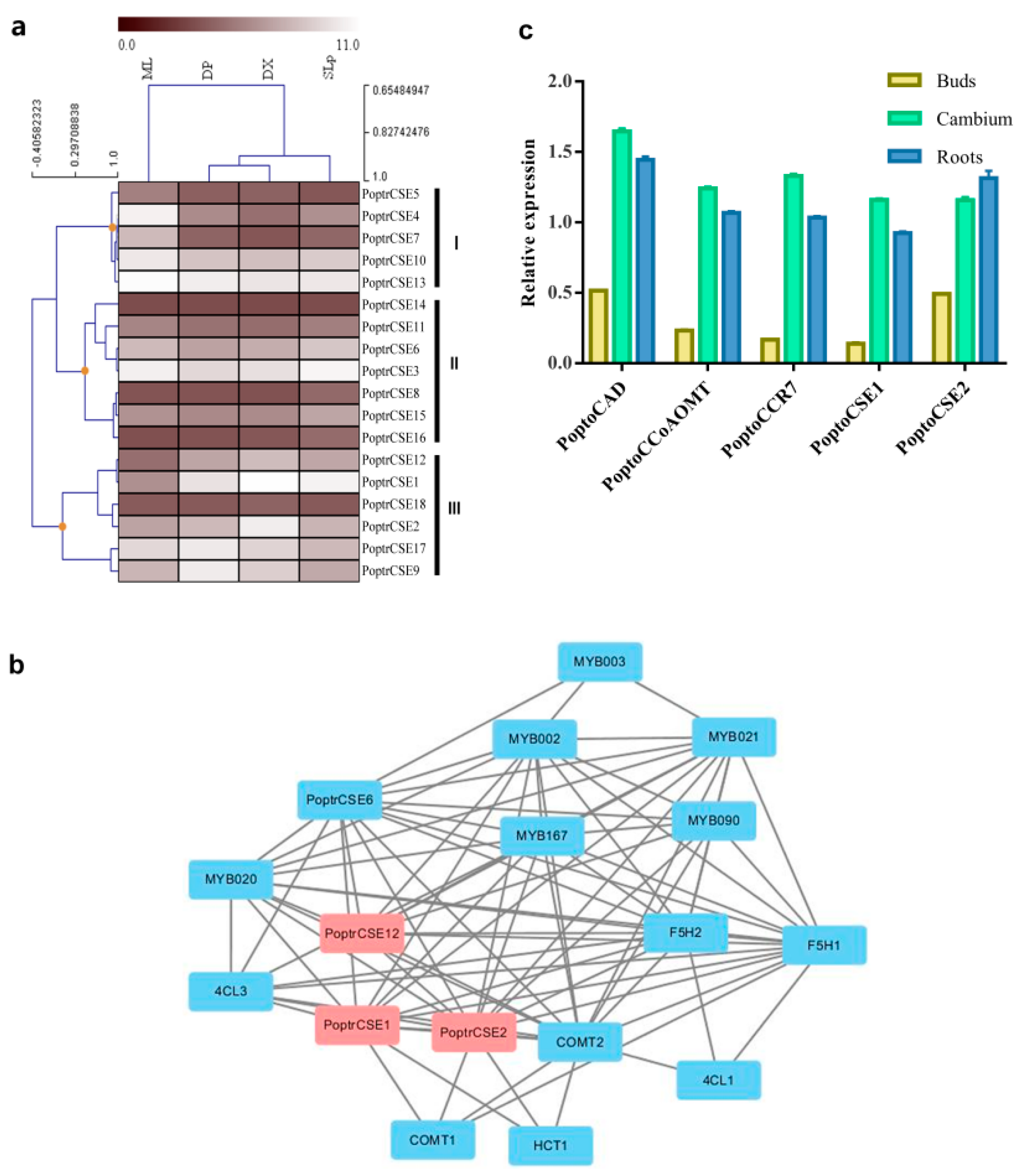
2.4. Expression Analysis of CSE Genes
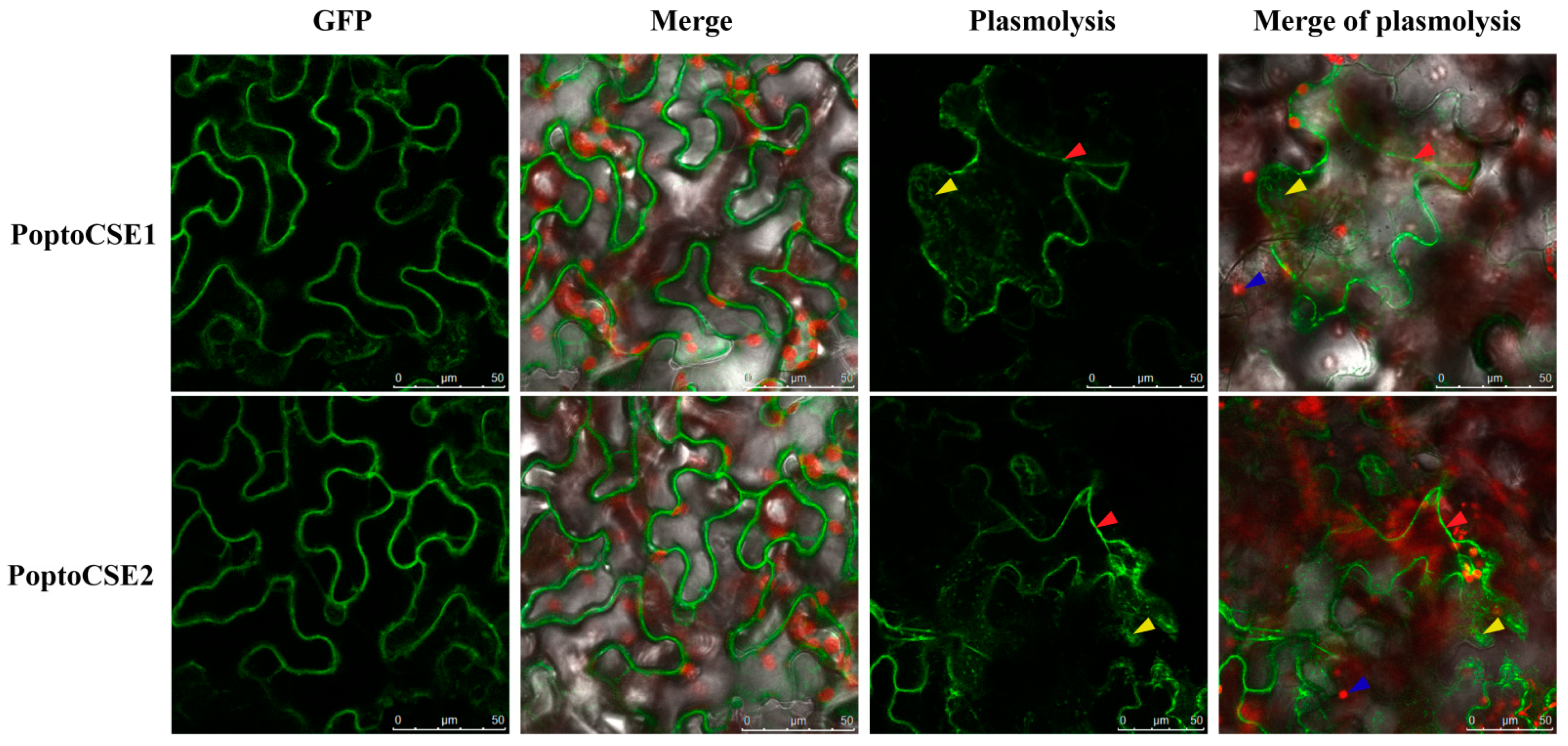
2.5. Characteristics of PoptoCSE1 and PoptoCSE2
2.6. Enzymatic Assay of PoptoCSE1, -2 and -12 of P. tomentosa
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Genome-Wide Identification of CSE Gene Family Members
4.3. Organization and Distribution of CSE Genes and Homologs on Populus Chromosomes
4.4. Promoter Analysis of PoptrCSE Genes
4.5. CSE Sequence Alignment and Phylogenetic Analysis
4.6. RNA Extraction and cDNA Synthesis
4.7. CSE Expression Profile in P. trichocarpa and P. tomentosa
4.8. Subcellular Localization of PoptoCSE
4.9. Purification of Recombinant CSE and HCT
4.10. Enzymatic Assay of Recombinant PoptoCSEs
4.11. GenBank Accession Number
4.12. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 4CL | 4-coumarate: CoA ligase |
| C3′H | P-coumarate 3-hydroxylase |
| C4H | Cinnamate 4-hydroxylase |
| CAD | Cinnamyl alcohol dehydrogenase |
| CCoAOMT | Caffeoyl-CoA 3-O-methyltransferas |
| CCR | Cinnamoyl-CoA reductase |
| DP | Developing phloem |
| DX | Developing xylem |
| ER | Endoplasmic reticulum |
| F5H | Ferulate 5-hydroxylase |
| GFP | Green fluorescent protein |
| HCT | Hydroxylcinnamoyl-CoA shikimate/quinate hydroxycinnamoyl transferase |
| HSE | Heat stress responsive element |
| LTR | Low temperature responsive element |
| LysoPL2 | Lysophospholipase 2 |
| MAGL | Monoacylglycerol lipase |
| MGAT | Monoacylglycerol acyltransferase |
| ML | Mature leaf |
| PAL | Phenylalanine ammonia-lyase |
| Semi-qRT-PCR | Semi-quantitative reverse transcription polymerase chain reaction |
| SLp | Shoot and leaf primordium |
| SMRE | Secondary wall MYB responsive element |
| SNBE | Secondary wall NAC binding element |
References
- Poovaiah, C.R.; Nageswara-Rao, M.; Soneji, J.R.; Baxter, H.L.; Stewart, C.N., Jr. Altered lignin biosynthesis using biotechnology to improve lignocellulosic biofuel feedstocks. Plant Biotechnol. J. 2014, 12, 1163–1173. [Google Scholar] [CrossRef]
- Vanholme, R.; Demedts, B.; Morreel, K.; Ralph, J.; Boerjan, W. Lignin biosynthesis and structure. Plant Physiol. 2010, 153, 895–905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhuiyan, N.H.; Selvaraj, G.; Wei, Y.; King, J. Gene expression profiling and silencing reveal that monolignol biosynthesis plays a critical role in penetration defence in wheat against powdery mildew invasion. J. Exp. Bot. 2009, 60, 509–521. [Google Scholar] [CrossRef]
- Wang, J.P.; Matthews, M.L.; Williams, C.M.; Shi, R.; Yang, C.; Tunlaya-Anukit, S.; Chen, H.-C.; Li, Q.; Liu, J.; Lin, C.-Y.; et al. Improving wood properties for wood utilization through multi-omics integration in lignin biosynthesis. Nat. Commun. 2018, 9, 1579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ragauskas, A.J.; Beckham, G.T.; Biddy, M.J.; Chandra, R.; Chen, F.; Davis, M.F.; Davison, B.H.; Dixon, R.A.; Gilna, P.; Keller, M.; et al. Lignin valorization: Improving lignin processing in the biorefinery. Science 2014, 344, 1246843. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Dixon, R.A. Altering the cell wall and its impact on plant disease: From forage to bioenergy. Annu. Rev. Phytopathol. 2014, 52, 69–91. [Google Scholar] [CrossRef] [Green Version]
- Tang, W.; Tang, A.Y. Transgenic woody plants for biofuel. J. For. Res. 2014, 25, 225–236. [Google Scholar] [CrossRef]
- Vanholme, R.; Storme, V.; Vanholme, B.; Sundin, L.; Christensen, J.H.; Goeminne, G.; Halpin, C.; Rohde, A.; Morreel, K.; Boerjan, W. A systems biology view of responses to lignin biosynthesis perturbations in Arabidopsis. Plant Cell 2012, 24, 3506–3529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Chapple, C. Understanding lignification: Challenges beyond monolignol biosynthesis. Plant Physiol. 2010, 154, 449–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voelker, S.L.; Lachenbruch, B.; Meinzer, F.C.; Strauss, S.H. Reduced wood stiffness and strength, and altered stem form, in young antisense 4CL transgenic poplars with reduced lignin contents. New Phytol. 2011, 189, 1096–1109. [Google Scholar] [CrossRef]
- Stewart, J.; Akiyama, T.; Chapple, C.; Ralph, J.; Mansfield, S. The effects on lignin structure of overexpression of ferulate 5-hydroxylase in hybrid poplar. Plant Physiol. 2009, 150, 621–635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Punith, N.; Wang, J.P.; Ronald, S.; Vincent, C.; Cranos, W.; Ducoste, J.J.; Daniel, C. Assessing the impact of the 4CL enzyme complex on the robustness of monolignol biosynthesis using metabolic pathway analysis. PLoS ONE 2018, 13, e0193896. [Google Scholar] [CrossRef] [Green Version]
- Vanholme, R.; Meester, B.D.; Ralph, J.; Boerjan, W. Lignin biosynthesis and its integration into metabolism. Curr. Opin. Biotechnol. 2019, 56, 230–239. [Google Scholar] [CrossRef]
- Qiao, Z. Lignification: Flexibility, biosynthesis and regulation. Trends Plant Sci. 2016, 21, 713–721. [Google Scholar] [CrossRef]
- Whetten, R.; Sederoff, R. Lignin biosynthesis. Plant Cell 1995, 7, 1001–1013. [Google Scholar] [CrossRef]
- Boerjan, W.; Ralph, J.; Baucher, M. Lignin biosynthesis. Annu. Rev. Plant Biol. 2003, 54, 519–546. [Google Scholar] [CrossRef]
- Weng, J.K.; Chapple, C. The origin and evolution of lignin biosynthesis. New Phytol. 2010, 187, 273–285. [Google Scholar] [CrossRef]
- Vanholme, R.; Cesarino, I.; Rataj, K.; Xiao, Y.; Sundin, L.; Goeminne, G.; Kim, H.; Cross, J.; Morreel, K.; Araujo, P.; et al. Caffeoyl shikimate esterase (CSE) is an enzyme in the lignin biosynthetic pathway in Arabidopsis. Science 2013, 341, 1103–1106. [Google Scholar] [CrossRef]
- Wang, J.; Matthews, M.; Naik, P.; Williams, C.; Ducoste, J.; Sederoff, R.; Chiang, V. Flux modeling for monolignol biosynthesis. Curr. Opin. Biotechnol. 2019, 56, 187–192. [Google Scholar] [CrossRef]
- Guo, L.; Wang, P.; Jaini, R.; Dudareva, N.; Chapple, C.; Morgan, J. Dynamic modeling of subcellular phenylpropanoid metabolism in Arabidopsis lignifying cells. Metab. Eng. 2018, 49, 36–46. [Google Scholar] [CrossRef]
- Faraji, M.; Fonseca, L.; Escamilla-Treviño, L.; Barros-Rios, J.; Engle, N.; Yang, Z.; Tschaplinski, T.; Dixon, R.; Voit, E. Mathematical models of lignin biosynthesis. Biotechnol. Biofuels 2018, 11, 34. [Google Scholar] [CrossRef] [Green Version]
- Faraji, M.; Fonseca, L.; Escamilla-Treviño, L.; Barros-Rios, J.; Engle, N.; Yang, Z.; Tschaplinski, T.; Dixon, R.; Voit, E. A dynamic model of lignin biosynthesis in Brachypodium distachyon. Biotechnol. Biofuels 2018, 11, 253. [Google Scholar] [CrossRef]
- Saleme, M.L.S.; Cesarino, I.; Vargas, L.; Kim, H.; Vanholme, R.; Goeminne, G.; Van Acker, R.; Fonseca, F.C.A.; Pallidis, A.; Voorend, W.; et al. Silencing caffeoyl shikimate esterase affects lignification and improves saccharification in poplar. Plant Physiol. 2017, 175, 1040–1057. [Google Scholar] [CrossRef] [Green Version]
- Gao, W.; Li, H.Y.; Xiao, S.; Chye, M.L. Acyl-CoA-binding protein 2 binds lysophospholipase 2 and lysoPC to promote tolerance to cadmium-induced oxidative stress in transgenic Arabidopsis. Plant J. Cell Mol. Biol. 2010, 62, 989–1003. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.P.; Naik, P.P.; Chen, H.C.; Shi, R.; Lin, C.Y.; Liu, J.; Shuford, C.M.; Li, Q.; Sun, Y.H.; Tunlaya-Anukit, S.; et al. Complete proteomic-based enzyme reaction and inhibition kinetics reveal how monolignol biosynthetic enzyme families affect metabolic flux and lignin in Populus trichocarpa. Plant Cell 2014, 26, 894–914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ha, C.M.; Escamilla-Trevino, L.; Yarce, J.C.; Kim, H.; Ralph, J.; Chen, F.; Dixon, R.A. An essential role of caffeoyl shikimate esterase in monolignol biosynthesis in Medicago truncatula. Plant J. Cell Mol. Biol. 2016, 86, 363–375. [Google Scholar] [CrossRef]
- Wang, X.; Chao, N.; Zhang, M.; Jiang, X.; Gai, Y. Functional characteristics of caffeoyl shikimate esterase in Larix kaempferi and monolignol biosynthesis in gymnosperms. Int. J. Mol. Sci. 2019, 20, 6071. [Google Scholar] [CrossRef] [Green Version]
- de Vries, L.; Brouckaert, M.; Chanoca, A.; Kim, H.; Regner, M.R.; Timokhin, V.I.; Sun, Y.; De Meester, B.; Van Doorsselaere, J.; Goeminne, G.; et al. CRISPR-Cas9 editing of caffeoyl shikimate esterase 1 and 2 shows their importance and partial redundancy in lignification in Populus tremula × P. alba. Plant Biotechnol. J. 2021, 19, 2221–2234. [Google Scholar] [CrossRef]
- Jang, H.A.; Bae, E.K.; Kim, M.H.; Park, S.J.; Choi, N.Y.; Pyo, S.W.; Lee, C.; Jeong, H.Y.; Lee, H.; Choi, Y.I.; et al. CRISPR-Knockout of CSE Gene improves saccharification efficiency by reducing lignin content in hybrid poplar. Int. J. Mol. Sci. 2021, 22, 9750. [Google Scholar] [CrossRef] [PubMed]
- Tuskan, G.A.; Difazio, S.; Jansson, S.; Bohlmann, J.; Grigoriev, I.; Hellsten, U.; Putnam, N.; Ralph, S.; Rombauts, S.; Salamov, A.; et al. The genome of black cottonwood, Populus trichocarpa (Torr. & Gray). Science 2006, 313, 1596–1604. [Google Scholar] [CrossRef] [Green Version]
- Dai, X.; Hu, Q.; Cai, Q.; Feng, K.; Ye, N.; Tuskan, G.; Milne, R.; Chen, Y.; Wan, Z.; Wang, Z.; et al. The willow genome and divergent evolution from poplar after the common genome duplication. Cell Res. 2014, 24, 1274–1277. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.; Ye, N.; Dong, Z.; Lu, M.; Li, L.; Yin, T. Major Chromosomal rearrangements distinguish willow and poplar after the ancestral “Salicoid” genome duplication. Genome Biol. Evol. 2016, 8, 1868–1875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, J.; Wang, Z. Molecular cloning, expression and characterization of a phenylalanine ammonia-lyase gene (SmPAL1) from Salvia miltiorrhiza. Mol. Biol. Rep. 2008, 36, 939–952. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, M.; Hanada, A.; Yamauchi, Y.; Kuwahara, A.; Kamiya, Y.; Yamaguchi, S. Gibberellin biosynthesis and response during Arabidopsis seed germination. Plant Cell 2003, 15, 1591–1604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hobo, T.; Asada, M.; Kowyama, Y.; Hattori, T. ACGT-containing abscisic acid response element (ABRE) and coupling element 3 (CE3) are functionally equivalent. Plant J. Cell Mol. Biol. 1999, 19, 679–689. [Google Scholar] [CrossRef] [PubMed]
- Hao, D.; Ohme-Takagi, M.; Sarai, A. Unique mode of GCC box recognition by the DNA-binding domain of ethylene-responsive element-binding factor (ERF domain) in plant. J. Biol. Chem. 1998, 273, 26857–26861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, J.; Klessig, D. Identification of a salicylic acid-responsive element in the promoter of the tobacco pathogenesis-related β-1,3-glucanase gene, PR-2d. Plant J. Cell Mol. Biol. 1997, 10, 1089–1101. [Google Scholar] [CrossRef]
- Guilfoyle, T.; Hagen, G.; Li, Y.; Ulmasov, T.; Liu, Z.B.; Strabala, T.; Gee, M. Auxin-regulated transcription. Aust. J. Plant Physiol. 1993, 20, 489–502. [Google Scholar] [CrossRef]
- Hartmann, U.; Sagasser, M.; Mehrtens, F.; Stracke, R.; Weisshaar, B. Differential combinatorial interactions of cis-acting elements recognized by R2R3-MYB, BZIP, and BHLH factors control light-responsive and tissue-specific activation of phenylpropanoid biosynthesis genes. Plant Mol. Biol. 2005, 57, 155–171. [Google Scholar] [CrossRef] [Green Version]
- Hatton, D.; Sablowski, R.; Yung, M.H.; Smith, C.; Schuch, W.; Bevan, M. Two classes of cis sequences contribute to tissue-specific expression of a PAL2 promoter in transgenic tobacco. Plant J. Cell Mol. Biol. 1995, 7, 859–876. [Google Scholar] [CrossRef]
- Zhao, Q.; Dixon, R.A. Transcriptional networks for lignin biosynthesis: More complex than we thought? Trends Plant Sci. 2011, 16, 227–233. [Google Scholar] [CrossRef] [Green Version]
- Legay, S.; Sivadon, P.; Blervacq, A.S.; Pavy, N.; Baghdady, A.; Tremblay, L.; Levasseur, C.; Ladouce, N.; Lapierre, C.; Seguin, A.; et al. EgMYB1, an R2R3 MYB transcription factor from eucalyptus negatively regulates secondary cell wall formation in Arabidopsis and poplar. New Phytol. 2010, 188, 774–786. [Google Scholar] [CrossRef]
- Öhman, D.; Demedts, B.; Kumar, M.; Gerber, L.; Gorzsás, A.; Goeminne, G.; Hedenström, M.; Ellis, B.; Boerjan, W.; Sundberg, B. MYB103 is required for ferulate-5-hydroxylase expression and syringyl lignin biosynthesis in Arabidopsis stems. Plant J. Cell Mol. Biol. 2012, 73, 63–76. [Google Scholar] [CrossRef] [PubMed]
- Fornale, S.; Sonbol, F.-M.; Maes, T.; Capellades, M.; Puigdomènech, P.; Rigau, J.; Caparros-Ruiz, D. Down-regulation of the maize and Arabidopsis thaliana caffeic acid o-methyl-transferase genes by two new maize R2R3-MYB transcription factors. Plant Mol. Biol. 2007, 62, 809–823. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, R.; Zhong, R.; Yu, L. Secondary wall NAC binding element (SNBE), a key cis-acting element required for target gene activation by secondary wall NAC master switches. Plant Signal. Behav. 2011, 6, 1282–1285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, R.; Ye, Z.H. Complexity of the transcriptional network controlling secondary wall biosynthesis. Plant Sci. Int. J. Exp. Plant Biol. 2014, 229, 193–207. [Google Scholar] [CrossRef] [Green Version]
- Vijayaraj, P.; Jashal, C.B.; Vijayakumar, A.; Rani, S.H.; Venkata Rao, D.K.; Rajasekharan, R. A bifunctional enzyme that has both monoacylglycerol acyltransferase and acyl hydrolase activities. Plant Physiol. 2012, 160, 667–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, A.J.; Loo, R.; Chen, Z.L.; Dennis, E. Regiospecificity and catalytic triad of lysophospholipase I. J. Biol. Chem. 1997, 272, 22030–22036. [Google Scholar] [CrossRef] [Green Version]
- Raes, J.; Rohde, A.; Christensen, J.H.; Van de Peer, Y.; Boerjan, W. Genome-wide characterization of the lignification toolbox in Arabidopsis. Plant Physiol. 2003, 133, 1051–1071. [Google Scholar] [CrossRef] [Green Version]
- Kim, R.J.; Kim, H.J.; Shim, D.; Suh, M.C. Molecular and biochemical characterizations of the monoacylglycerol lipase gene family of Arabidopsis thaliana. Plant J. 2016, 85, 758–771. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Lee, C.; Zhong, R.; Ye, Z.H. MYB58 and MYB63 are transcriptional activators of the lignin biosynthetic pathway during secondary cell wall formation in Arabidopsis. Plant Cell 2009, 21, 248–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, R.; Ye, Z.H. MYB46 and MYB83 bind to the SMRE sites and directly activate a suite of transcription factors and secondary wall biosynthetic genes. Plant Cell Physiol. 2012, 53, 368–380. [Google Scholar] [CrossRef]
- Zhong, R.; Lee, C.; Ye, Z.H. Global analysis of direct targets of secondary wall NAC master switches in Arabidopsis. Mol. Plant 2010, 3, 1087–1103. [Google Scholar] [CrossRef]
- Tronchet, M.; Balague, C.; Kroj, T.; Jouanin, L.; Roby, D. Cinnamyl alcohol dehydrogenases-C and D, key enzymes in lignin biosynthesis, play an essential role in disease resistance in Arabidopsis. Mol. Plant Pathol. 2010, 11, 83–92. [Google Scholar] [CrossRef]
- Moura, J.C.; Bonine, C.A.; de Oliveira Fernandes Viana, J.; Dornelas, M.C.; Mazzafera, P. Abiotic and biotic stresses and changes in the lignin content and composition in plants. J. Integr. Plant Biol. 2010, 52, 360–376. [Google Scholar] [CrossRef]
- Lauvergeat, V.; Lacomme, C.; Lacombe, E.; Lasserre, E.; Roby, D.; Grima-Pettenati, J. Two cinnamoyl-CoA reductase (CCR) genes from Arabidopsis thaliana are differentially expressed during development and in response to infection with pathogenic bacteria. Phytochemistry 2001, 57, 1187–1195. [Google Scholar] [CrossRef]
- Shah, J. Lipases in signaling plant defense responses. In Signaling and Communication in Plants; Wang, X., Ed.; Phospholipases in plant signaling; Springer: Berlin, Germany, 2014; Volume 20, pp. 207–228. [Google Scholar]
- Coordinators, N.R. Database resources of the national center for biotechnology information. Nucleic Acids Res. 2017, 46, D8–D13. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morita, S.; Yamashita, Y.; Fujiki, M.; Todaka, R.; Nishikawa, Y.; Hosoki, A.; Yabe, C.; Nakamura, J.I.; Kawamura, K.; Suwastika, I.; et al. Expression of a rice glutaredoxin in aleurone layers of developing and mature seeds: Subcellular localization and possible functions in antioxidant defense. Planta 2015, 242, 1195–1206. [Google Scholar] [CrossRef]
- Escamilla-Trevino, L.L.; Shen, H.; Hernandez, T.; Yin, Y.; Xu, Y.; Dixon, R.A. Early lignin pathway enzymes and routes to chlorogenic acid in switchgrass (Panicum virgatum L.). Plant Mol. Biol. 2014, 84, 565–576. [Google Scholar] [CrossRef] [PubMed]
- Chao, N.; Li, N.; Qi, Q.; Li, S.; Lyu, T.; Jiang, X.; Gai, Y. Characterization of the cinnamoyl-CoA reductase (CCR) gene family in Populus tomentosa reveals the enzymatic active sites and evolution of CCR. Planta 2017, 245, 61–75. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of SNBEs | Number of SMREs | Salicylic Acid | MeJA | Gibberellin | Auxin | Abscisic Acid | Ethylene | AC-I Element | AC-II Element | LTR | HSE | Anaerobic Induction | Defense and Stress (TC-Rich Repeats) | MYB Involved in Drought Inducibility (MBS) | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PoptrCSE1 | 4 | 5 | - | - | 1 | - | 1 | - | 2 | 1 | 1 | - | 2 | - | - |
| PoptrCSE2 | 4 | 6 | - | - | - | - | 1 | - | 1 | - | 3 | 3 | - | 4 | - |
| PoptrCSE3 | 1 | 4 | 1 | - | 1 | 1 | - | 1 | - | - | 2 | 2 | 5 | 1 | - |
| PoptrCSE4 | 4 | 1 | - | - | - | - | 1 | - | - | - | - | - | 2 | 1 | 1 |
| PoptrCSE5 | 6 | 2 | - | - | 1 | 1 | - | - | - | 1 | - | - | 2 | 1 | 2 |
| PoptrCSE6 | 4 | 2 | 2 | - | 1 | 1 | 1 | 1 | - | 1 | - | 2 | 3 | 1 | 2 |
| PoptrCSE7 | 3 | 5 | 2 | 1 | - | - | 2 | - | - | - | - | 4 | 1 | 2 | 1 |
| PoptrCSE8 | 5 | 3 | - | 1 | 1 | - | 1 | 2 | - | - | - | - | 2 | - | 1 |
| PoptrCSE9 | 3 | 3 | 1 | - | 1 | 1 | - | 1 | - | - | 2 | 2 | 5 | 1 | - |
| PoptrCSE10 | 2 | 5 | 1 | 2 | - | 1 | 1 | 1 | - | - | - | - | 3 | - | - |
| PoptrCSE11 | 2 | 3 | - | - | - | - | 1 | - | - | - | - | - | 5 | 2 | 1 |
| PoptrCSE12 | 5 | 2 | 1 | 1 | - | - | 1 | 1 | 1 | - | - | 3 | 1 | 1 | 4 |
| PoptrCSE13 | 0 | 0 | - | - | - | - | 1 | - | - | - | 1 | 1 | - | - | - |
| PoptrCSE14 | 7 | 2 | 1 | 2 | 1 | 1 | 1 | - | - | - | - | 3 | 1 | 2 | 3 |
| PoptrCSE15 | 5 | 4 | 1 | 3 | - | 2 | 1 | - | - | - | 1 | 1 | - | 1 | 2 |
| PoptrCSE16 | 5 | 4 | 1 | 3 | - | - | - | - | - | - | - | 2 | 1 | 1 | 2 |
| PoptrCSE17 | 2 | 2 | 1 | 2 | 2 | 1 | 3 | 1 | - | - | 1 | 1 | - | 3 | 4 |
| PoptrCSE18 | 2 | 4 | 1 | 1 | 1 | - | - | - | - | 1 | 3 | 1 | 4 | 2 | 2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Chao, N.; Zhang, A.; Kang, J.; Jiang, X.; Gai, Y. Systematic Analysis and Biochemical Characterization of the Caffeoyl Shikimate Esterase Gene Family in Poplar. Int. J. Mol. Sci. 2021, 22, 13366. https://doi.org/10.3390/ijms222413366
Wang X, Chao N, Zhang A, Kang J, Jiang X, Gai Y. Systematic Analysis and Biochemical Characterization of the Caffeoyl Shikimate Esterase Gene Family in Poplar. International Journal of Molecular Sciences. 2021; 22(24):13366. https://doi.org/10.3390/ijms222413366
Chicago/Turabian StyleWang, Xuechun, Nan Chao, Aijing Zhang, Jiaqi Kang, Xiangning Jiang, and Ying Gai. 2021. "Systematic Analysis and Biochemical Characterization of the Caffeoyl Shikimate Esterase Gene Family in Poplar" International Journal of Molecular Sciences 22, no. 24: 13366. https://doi.org/10.3390/ijms222413366
APA StyleWang, X., Chao, N., Zhang, A., Kang, J., Jiang, X., & Gai, Y. (2021). Systematic Analysis and Biochemical Characterization of the Caffeoyl Shikimate Esterase Gene Family in Poplar. International Journal of Molecular Sciences, 22(24), 13366. https://doi.org/10.3390/ijms222413366