
Long-Chain and Very Long-Chain Ceramides Mediate Doxorubicin-Induced Toxicity and Fibrosis

 , , , ,
, , , ,  and
and
Abstract
:1. Introduction
2. Results
2.1. Determination of Dox Concentration and Lethality Measurement Identified 0.7 µM Dox as Most Suited Concentration
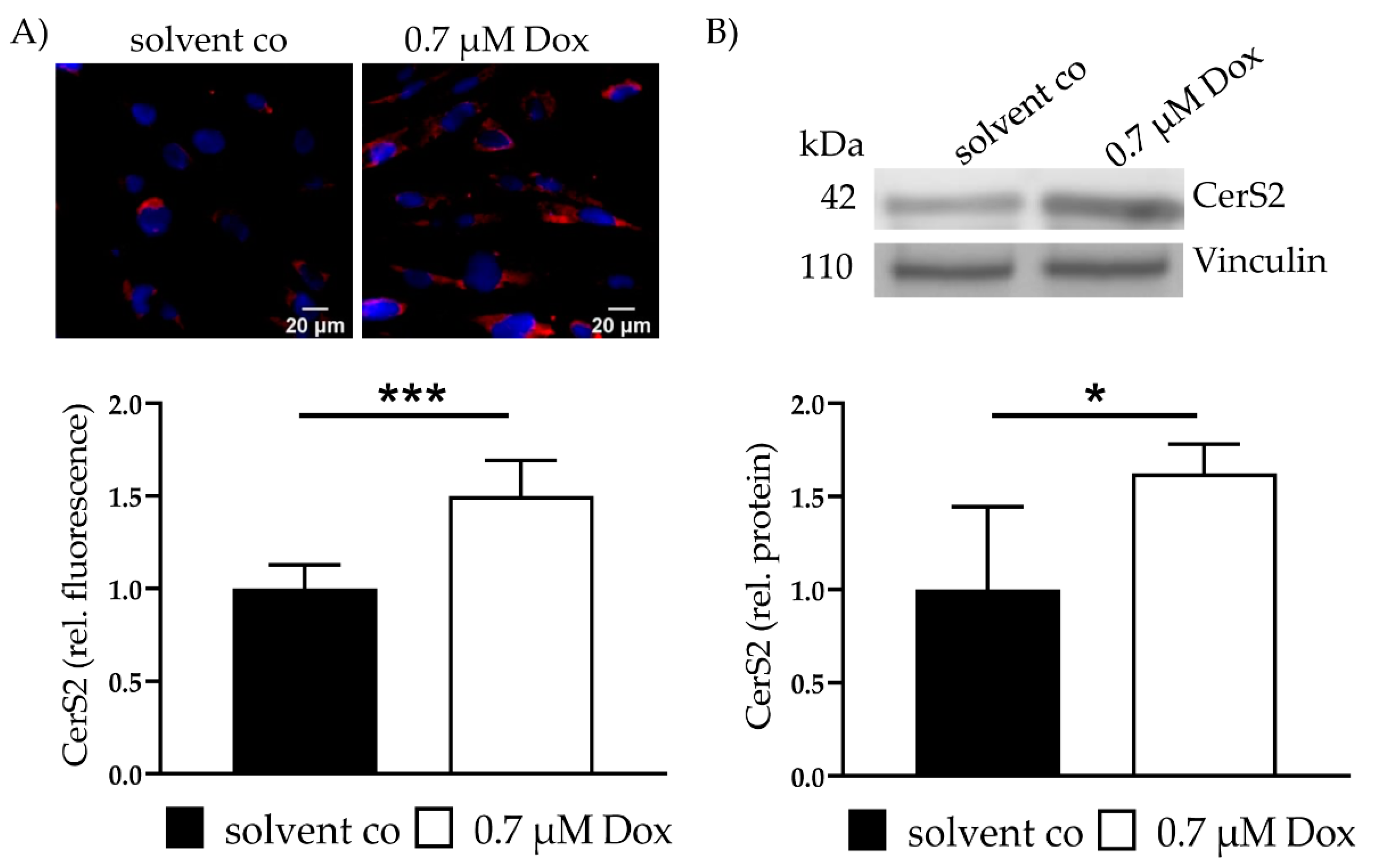
2.2. Dox Increases CerS2 Protein Expression and Ceramide Levels
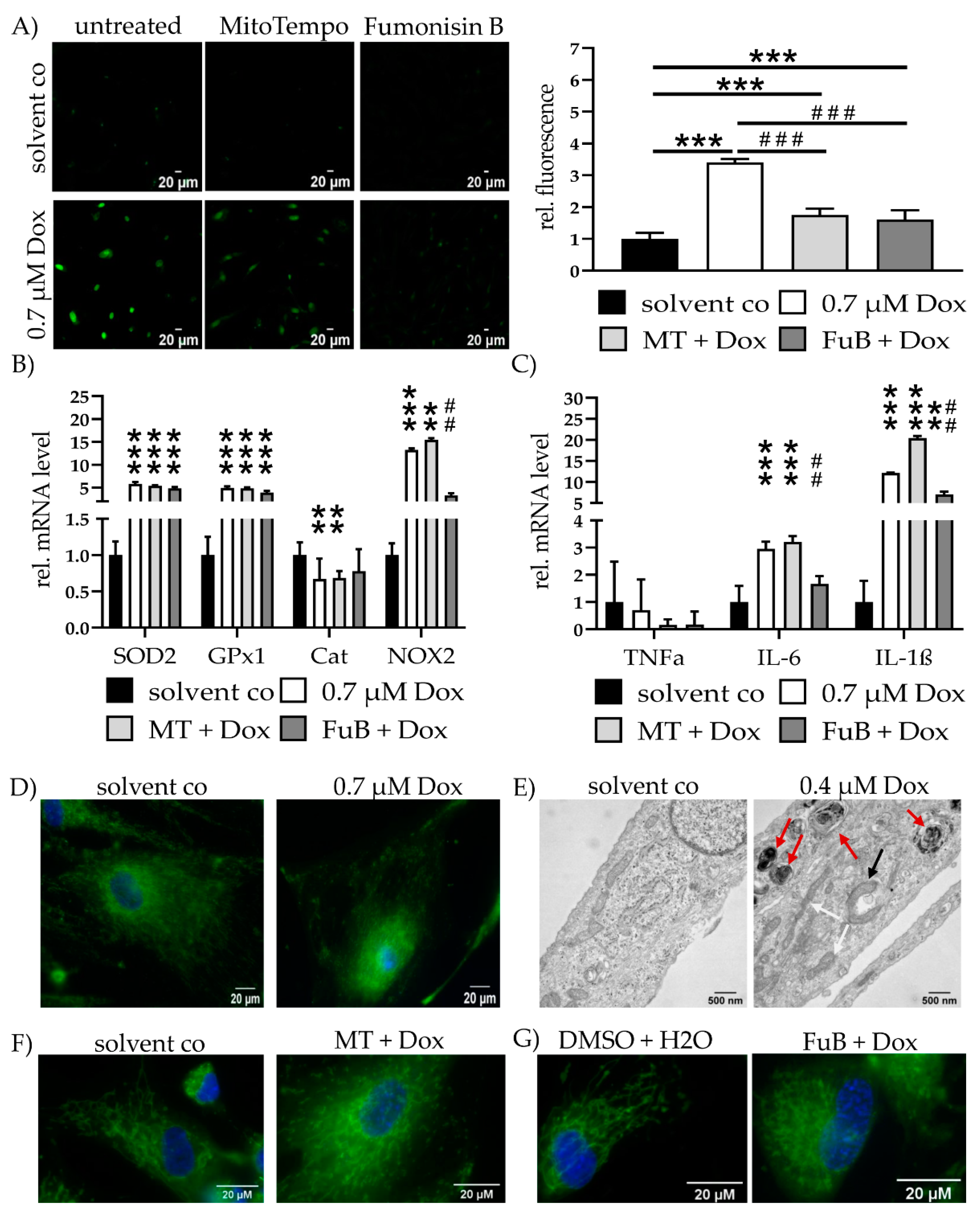
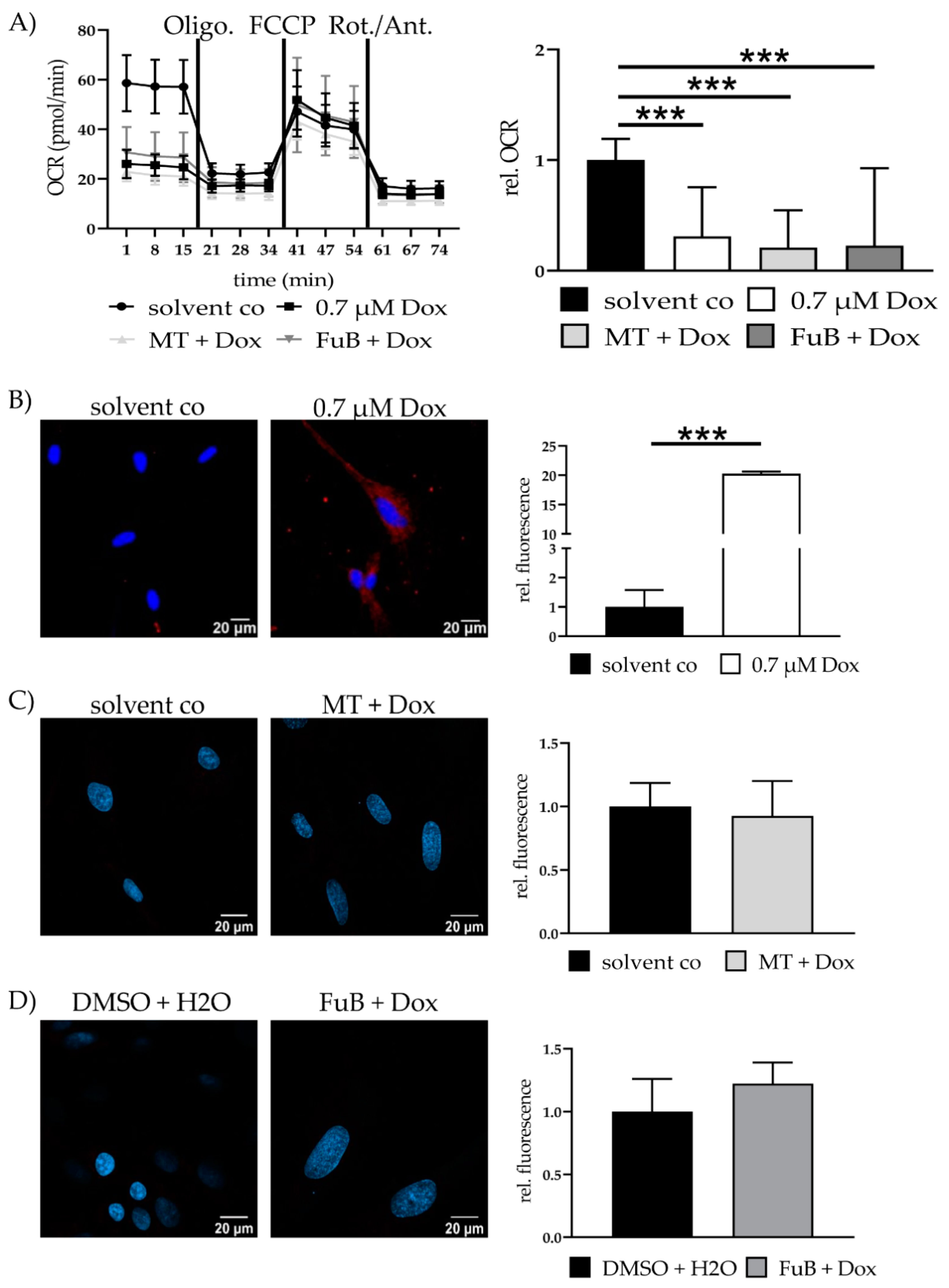
2.3. Dox Damages Mitochondrial Structure by Increasing Mitochondrial ROS Levels
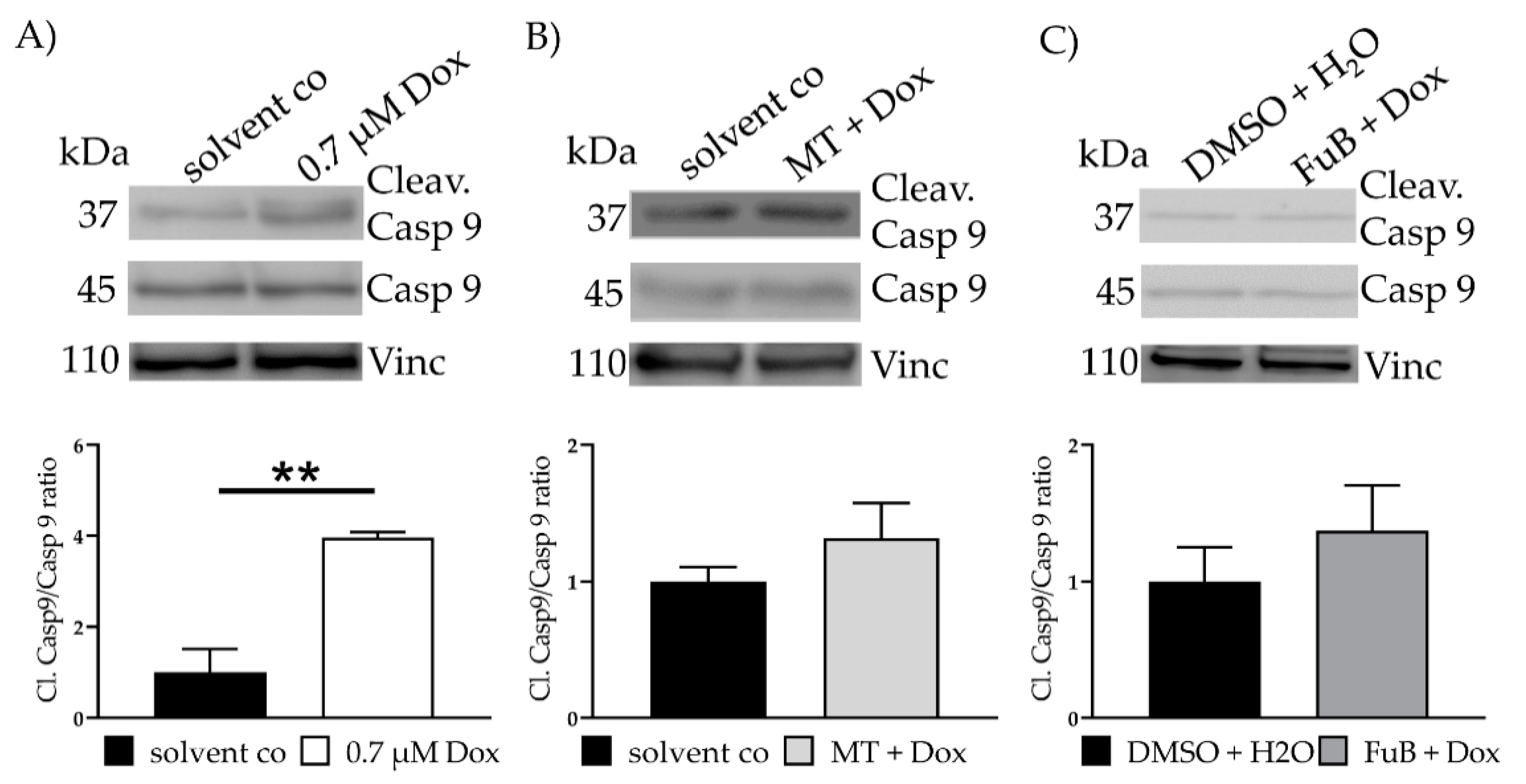
2.4. Dox impairs Mitochondrial Homeostasis and Function and Causes Apoptosis
2.5. Dox Promotes Fibrosis
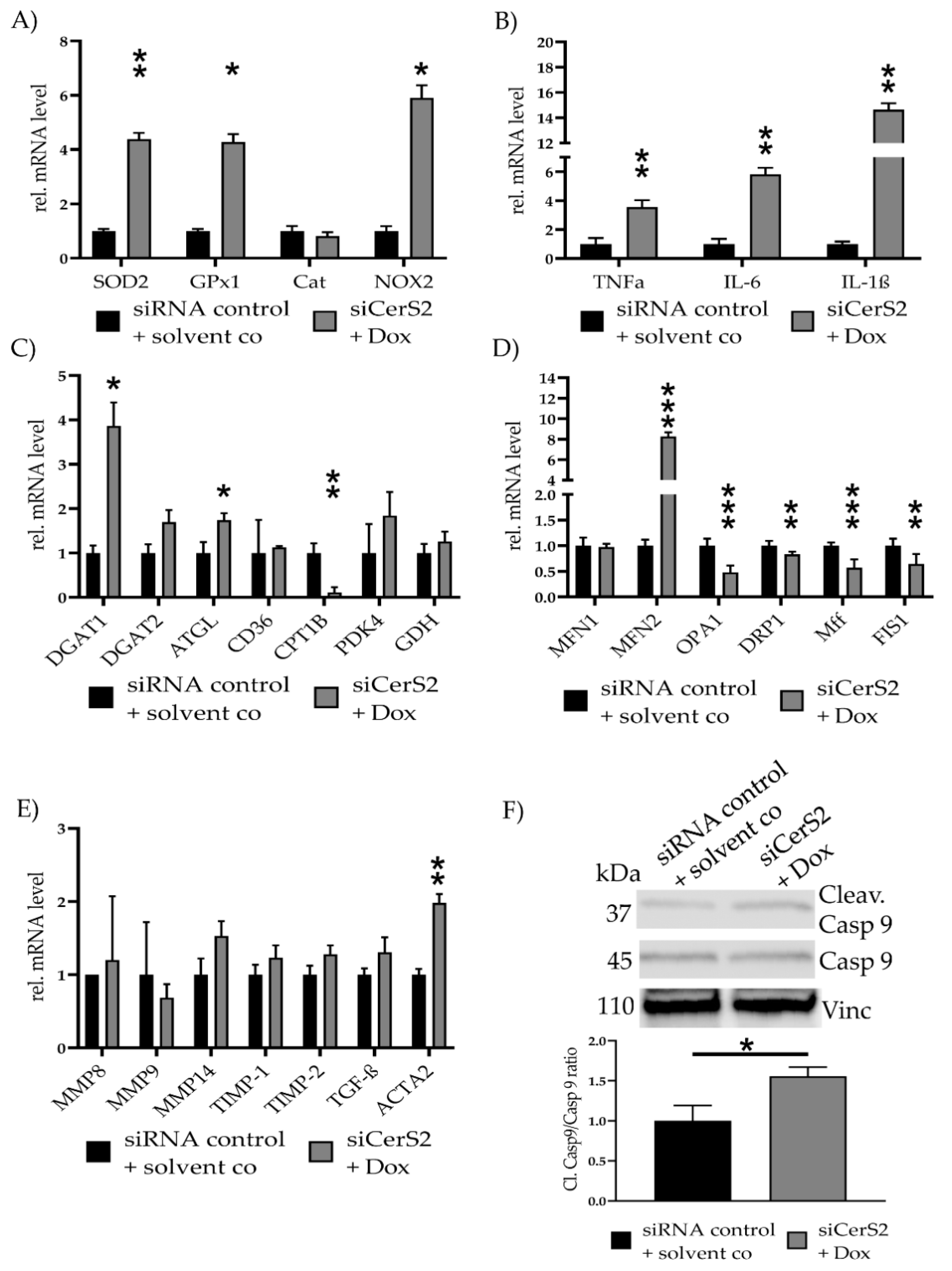
2.6. CerS2 Knockdown Improves Dox-Mediated Mitochondrial Damage and Fibrosis
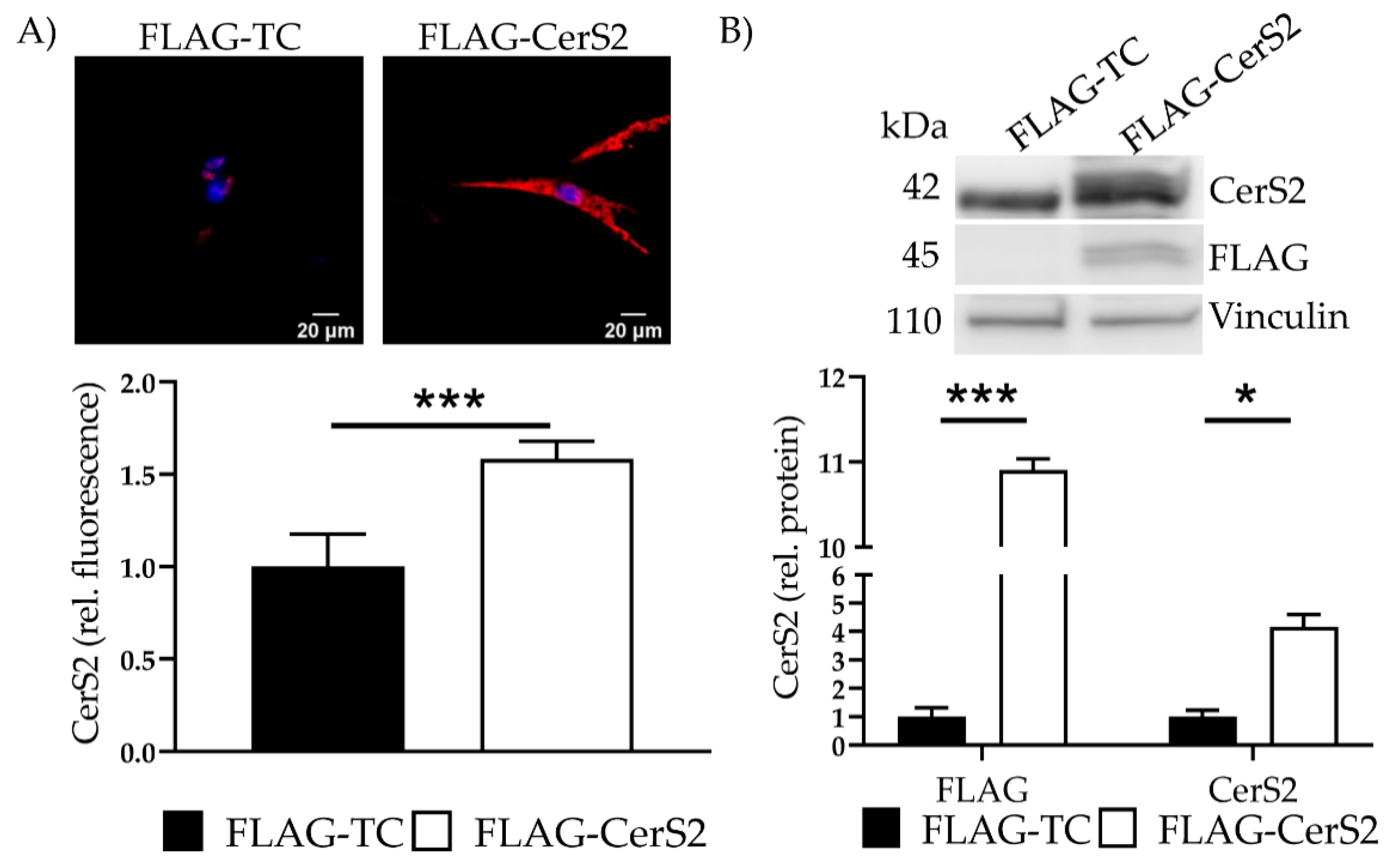
2.7. CerS2 Overexpression Leads to an Increased Ceramide Production
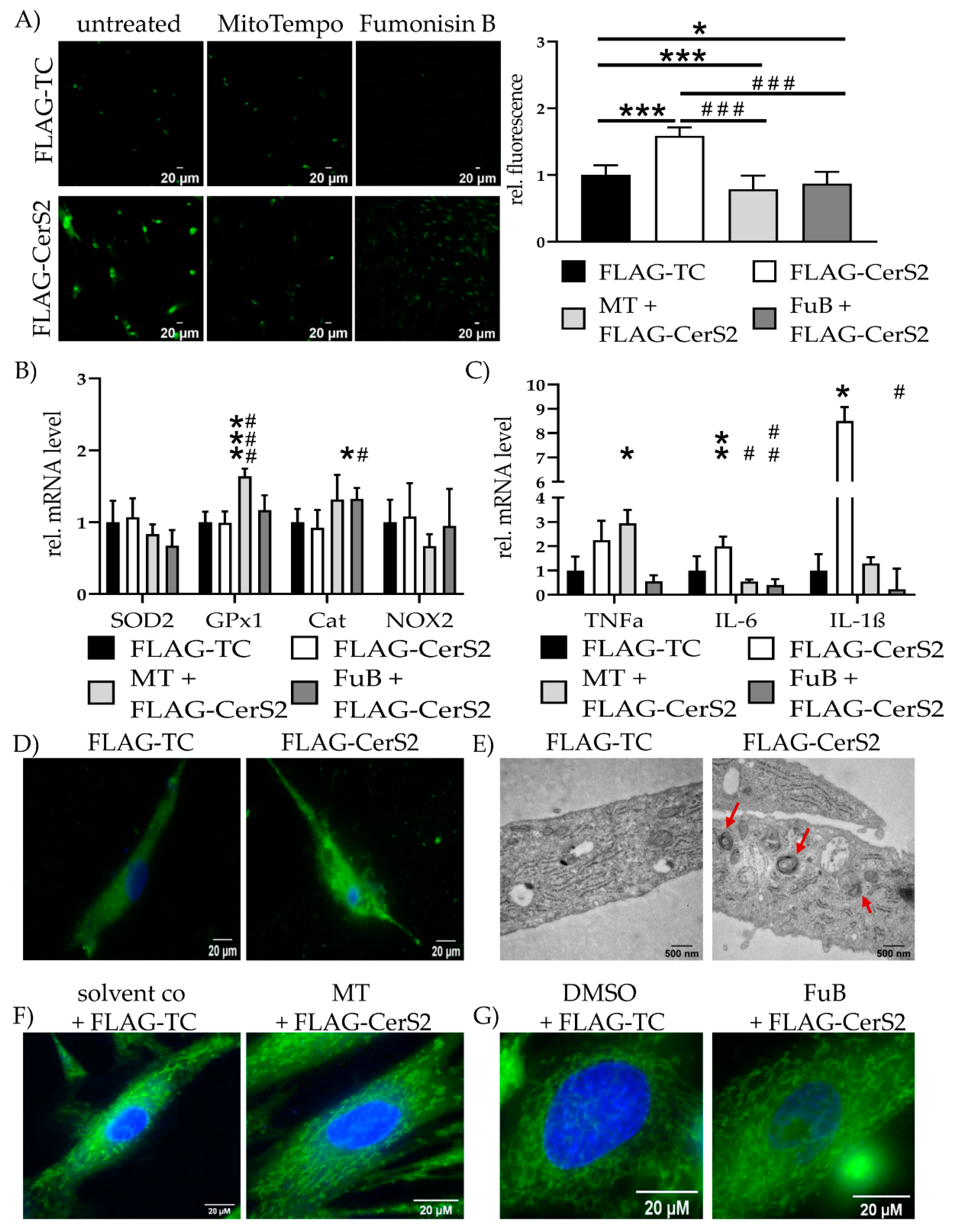
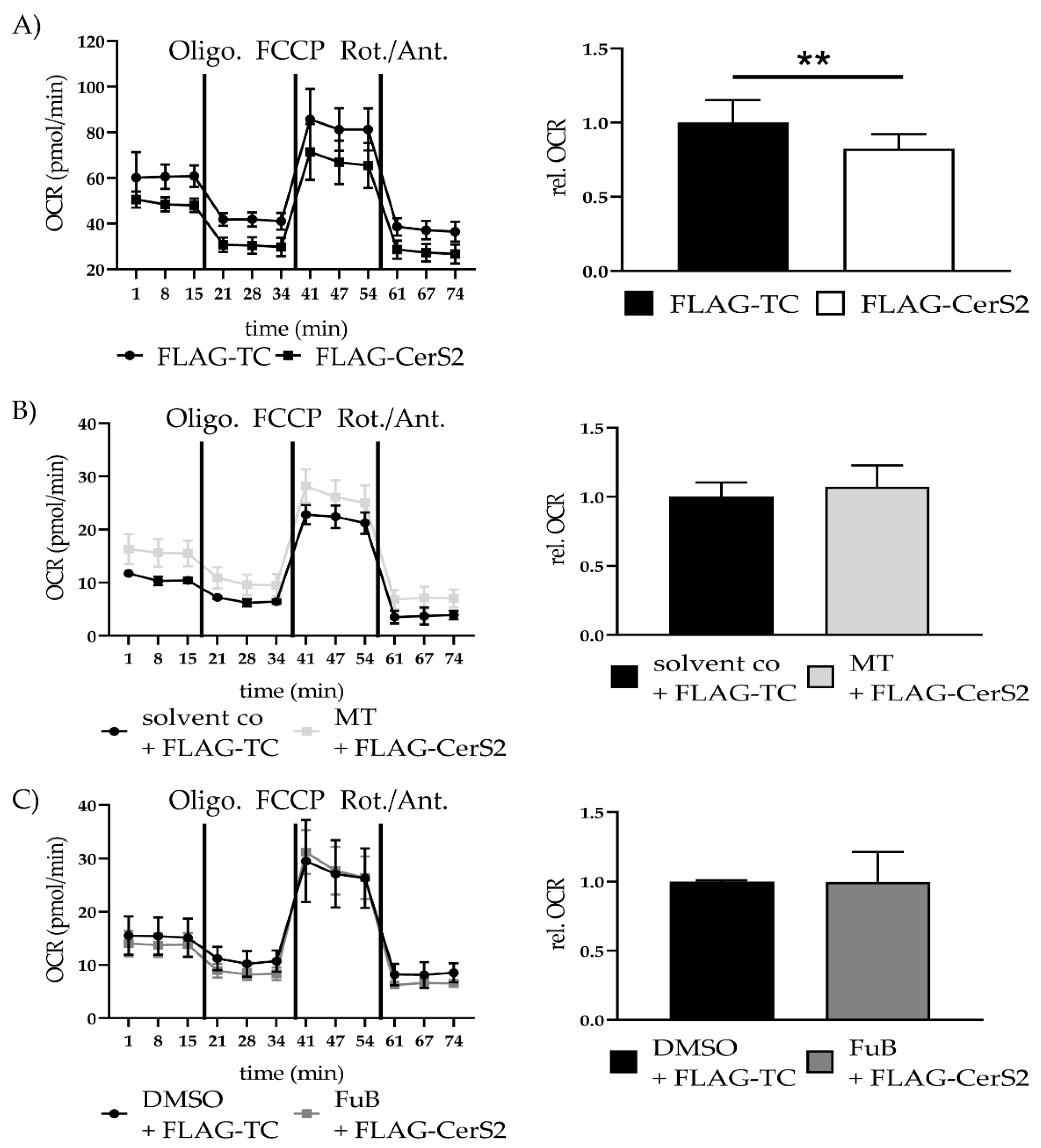
2.8. CerS2 Overexpression Impairs Mitochondrial Structure and Function
2.9. FLAG-CerS2 Leads to Reduced Mitochondrial Structure and Function and Induces Apoptosis
3. Discussion
4. Material and Methods
4.1. CerS2 Overexpression and CerS2 Silencing
4.2. Cell Culture
4.3. RNA Isolation and mRNA Expression Level Validation
4.4. Protein Isolation and Protein Expression Level Detection
4.5. Ceramide Content
4.6. Ceramide Detection with FACS
4.7. Immunohistochemistry
4.8. Electron Microscopy
4.9. Mitochondrial ROS Measurement with MitoSOX
4.10. Seahorse Analysis
4.11. Collagen Measurement in Cell Supernatant
4.12. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tacar, O.; Sriamornsak, P.; Dass, C.R. Doxorubicin: An update on anticancer molecular action, toxicity and novel drug delivery systems. J. Pharm. Pharmacol. 2013, 65, 157–170. [Google Scholar] [CrossRef]
- Chatterjee, K.; Zhang, J.; Honbo, N.; Karliner, J.S. Doxorubicin cardiomyopathy. Cardiology 2010, 115, 155–162. [Google Scholar] [CrossRef]
- Levick, S.P.; Soto-Pantoja, D.R.; Bi, J.; Hundley, W.G.; Widiapradja, A.; Manteufel, E.J.; Bradshaw, T.W.; Melendez, G.C. Doxorubicin-Induced Myocardial Fibrosis Involves the Neurokinin-1 Receptor and Direct Effects on Cardiac Fibroblasts. Heart Lung Circ. 2019, 28, 1598–1605. [Google Scholar] [CrossRef] [PubMed]
- Alonso, A.; Goni, F.M. The Physical Properties of Ceramides in Membranes. Annu. Rev. Biophys. 2018, 47, 633–654. [Google Scholar] [CrossRef] [PubMed]
- Petrache, I.; Berdyshev, E.V. Ceramide Signaling and Metabolism in Pathophysiological States of the Lung. Annu. Rev. Physiol. 2016, 78, 463–480. [Google Scholar] [CrossRef]
- Law, B.A.; Liao, X.; Moore, K.S.; Southard, A.; Roddy, P.; Ji, R.; Szulc, Z.; Bielawska, A.; Schulze, P.C.; Cowart, L.A. Lipotoxic very-long-chain ceramides cause mitochondrial dysfunction, oxidative stress, and cell death in cardiomyocytes. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2018, 32, 1403–1416. [Google Scholar] [CrossRef] [Green Version]
- Jin, J.; Hou, Q.; Mullen, T.D.; Zeidan, Y.H.; Bielawski, J.; Kraveka, J.M.; Bielawska, A.; Obeid, L.M.; Hannun, Y.A.; Hsu, Y.T. Ceramide generated by sphingomyelin hydrolysis and the salvage pathway is involved in hypoxia/reoxygenation-induced Bax redistribution to mitochondria in NT-2 cells. J. Biol. Chem. 2008, 283, 26509–26517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitatani, K.; Idkowiak-Baldys, J.; Hannun, Y.A. The sphingolipid salvage pathway in ceramide metabolism and signaling. Cell. Signal. 2008, 20, 1010–1018. [Google Scholar] [CrossRef] [Green Version]
- Wattenberg, B.W. The long and the short of ceramides. J. Biol. Chem. 2018, 293, 9922–9923. [Google Scholar] [CrossRef] [Green Version]
- Levy, M.; Futerman, A.H. Mammalian ceramide synthases. IUBMB Life 2010, 62, 347–356. [Google Scholar] [CrossRef]
- Bandet, C.L.; Tan-Chen, S.; Bourron, O.; Le Stunff, H.; Hajduch, E. Sphingolipid Metabolism: New Insight into Ceramide-Induced Lipotoxicity in Muscle Cells. Int. J. Mol. Sci. 2019, 20, 479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walls, S.M.; Cammarato, A.; Chatfield, D.A.; Ocorr, K.; Harris, G.L.; Bodmer, R. Ceramide-Protein Interactions Modulate Ceramide-Associated Lipotoxic Cardiomyopathy. Cell Rep. 2018, 22, 2702–2715. [Google Scholar] [CrossRef] [Green Version]
- Hartmann, D.; Lucks, J.; Fuchs, S.; Schiffmann, S.; Schreiber, Y.; Ferreiros, N.; Merkens, J.; Marschalek, R.; Geisslinger, G.; Grosch, S. Long chain ceramides and very long chain ceramides have opposite effects on human breast and colon cancer cell growth. Int. J. Biochem. Cell Biol. 2012, 44, 620–628. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, Q.; Xu, C.; Lu, Y.; Hu, H.; Qin, B.; Wang, Y.; He, D.; Li, C.; Yu, X.; et al. MitoTEMPO Prevents Oxalate Induced Injury in NRK-52E Cells via Inhibiting Mitochondrial Dysfunction and Modulating Oxidative Stress. Oxid Med. Cell. Longev. 2017, 2017, 7528090. [Google Scholar] [CrossRef]
- Bouhet, S.; Hourcade, E.; Loiseau, N.; Fikry, A.; Martinez, S.; Roselli, M.; Galtier, P.; Mengheri, E.; Oswald, I.P. The mycotoxin fumonisin B1 alters the proliferation and the barrier function of porcine intestinal epithelial cells. Toxicol. Sci. 2004, 77, 165–171. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Yang, Z.; Li, J.; Li, E. Dexmedetomidine reduces the inflammation and apoptosis of doxorubicin-induced myocardial cells. Exp. Mol. Pathol. 2020, 113, 104371. [Google Scholar] [CrossRef]
- Du, J.; Hang, P.; Pan, Y.; Feng, B.; Zheng, Y.; Chen, T.; Zhao, L.; Du, Z. Inhibition of miR-23a attenuates doxorubicin-induced mitochondria-dependent cardiomyocyte apoptosis by targeting the PGC-1alpha/Drp1 pathway. Toxicol. Appl. Pharmacol. 2019, 369, 73–81. [Google Scholar] [CrossRef]
- Chitraju, C.; Mejhert, N.; Haas, J.T.; Diaz-Ramirez, L.G.; Grueter, C.A.; Imbriglio, J.E.; Pinto, S.; Koliwad, S.K.; Walther, T.C.; Farese, R.V., Jr. Triglyceride Synthesis by DGAT1 Protects Adipocytes from Lipid-Induced ER Stress during Lipolysis. Cell Metab. 2017, 26, 407–418.e3. [Google Scholar] [CrossRef]
- Chitraju, C.; Walther, T.C.; Farese, R.V., Jr. The triglyceride synthesis enzymes DGAT1 and DGAT2 have distinct and overlapping functions in adipocytes. J. Lipid Res. 2019, 60, 1112–1120. [Google Scholar] [CrossRef] [Green Version]
- Haemmerle, G.; Moustafa, T.; Woelkart, G.; Buttner, S.; Schmidt, A.; van de Weijer, T.; Hesselink, M.; Jaeger, D.; Kienesberger, P.C.; Zierler, K.; et al. ATGL-mediated fat catabolism regulates cardiac mitochondrial function via PPAR-alpha and PGC-1. Nat. Med. 2011, 17, 1076–1085. [Google Scholar] [CrossRef] [Green Version]
- Okamoto, F.; Tanaka, T.; Sohmiya, K.; Kawamura, K. CD36 abnormality and impaired myocardial long-chain fatty acid uptake in patients with hypertrophic cardiomyopathy. Jpn. Circ. J. 1998, 62, 499–504. [Google Scholar] [CrossRef] [Green Version]
- He, L.; Kim, T.; Long, Q.; Liu, J.; Wang, P.; Zhou, Y.; Ding, Y.; Prasain, J.; Wood, P.A.; Yang, Q. Carnitine palmitoyltransferase-1b deficiency aggravates pressure overload-induced cardiac hypertrophy caused by lipotoxicity. Circulation 2012, 126, 1705–1716. [Google Scholar] [CrossRef] [PubMed]
- Jezek, J.; Cooper, K.F.; Strich, R. Reactive Oxygen Species and Mitochondrial Dynamics: The Yin and Yang of Mitochondrial Dysfunction and Cancer Progression. Antioxidants 2018, 7, 13. [Google Scholar] [CrossRef]
- Mullen, T.D.; Hannun, Y.A.; Obeid, L.M. Ceramide synthases at the centre of sphingolipid metabolism and biology. Biochem. J. 2012, 441, 789–802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McLaughlin, D.; Zhao, Y.; O'Neill, K.M.; Edgar, K.S.; Dunne, P.D.; Kearney, A.M.; Grieve, D.J.; McDermott, B.J. Signalling mechanisms underlying doxorubicin and Nox2 NADPH oxidase-induced cardiomyopathy: Involvement of mitofusin-2. Br. J. Pharmacol. 2017, 174, 3677–3695. [Google Scholar] [CrossRef] [Green Version]
- Dorn, G., II. Mitochondrial fission/fusion and cardiomyopathy. Curr. Opin. Genet. Dev. 2016, 38, 38–44. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Detmer, S.A.; Ewald, A.J.; Griffin, E.E.; Fraser, S.E.; Chan, D.C. Mitofusins Mfn1 and Mfn2 coordinately regulate mitochondrial fusion and are essential for embryonic development. J. Cell Biol. 2003, 160, 189–200. [Google Scholar] [CrossRef]
- Tang, H.; Tao, A.; Song, J.; Liu, Q.; Wang, H.; Rui, T. Doxorubicin-induced cardiomyocyte apoptosis: Role of mitofusin 2. Int. J. Biochem. Cell Biol. 2017, 88, 55–59. [Google Scholar] [CrossRef]
- Liu, R.; Chan, D.C. The mitochondrial fission receptor Mff selectively recruits oligomerized Drp1. Mol. Biol. Cell 2015, 26, 4466–4477. [Google Scholar] [CrossRef]
- Shen, Q.; Yamano, K.; Head, B.P.; Kawajiri, S.; Cheung, J.T.; Wang, C.; Cho, J.H.; Hattori, N.; Youle, R.J.; van der Bliek, A.M. Mutations in Fis1 disrupt orderly disposal of defective mitochondria. Mol. Biol. Cell 2014, 25, 145–159. [Google Scholar] [CrossRef] [Green Version]
- Wynn, T.A. Cellular and molecular mechanisms of fibrosis. J. Pathol. 2008, 214, 199–210. [Google Scholar] [CrossRef] [Green Version]
- Fang, C.; Wen, G.; Zhang, L.; Lin, L.; Moore, A.; Wu, S.; Ye, S.; Xiao, Q. An important role of matrix metalloproteinase-8 in angiogenesis in vitro and in vivo. Cardiovasc. Res. 2013, 99, 146–155. [Google Scholar] [CrossRef]
- Appleby, T.C.; Greenstein, A.E.; Hung, M.; Liclican, A.; Velasquez, M.; Villasenor, A.G.; Wang, R.; Wong, M.H.; Liu, X.; Papalia, G.A.; et al. Biochemical characterization and structure determination of a potent, selective antibody inhibitor of human MMP9. J. Biol. Chem. 2017, 292, 6810–6820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Zhao, Y.; Chen, C.; Yang, L.; Lee, H.H.; Wang, Z.; Zhang, N.; Kolonin, M.G.; An, Z.; Ge, X.; et al. The critical role of MMP14 in adipose tissue remodeling during obesity. Mol. Cell. Biol. 2020, 40, e00564-19. [Google Scholar] [CrossRef]
- Khalil, H.; Kanisicak, O.; Prasad, V.; Correll, R.N.; Fu, X.; Schips, T.; Vagnozzi, R.J.; Liu, R.; Huynh, T.; Lee, S.J.; et al. Fibroblast-specific TGF-beta-Smad2/3 signaling underlies cardiac fibrosis. J. Clin. Investig. 2017, 127, 3770–3783. [Google Scholar] [CrossRef] [PubMed]
- Harikrishnan, V.; Titus, A.S.; Cowling, R.T.; Kailasam, S. Collagen receptor cross-talk determines alpha-smooth muscle actin-dependent collagen gene expression in angiotensin II-stimulated cardiac fibroblasts. J. Biol. Chem. 2019, 294, 19723–19739. [Google Scholar] [CrossRef]
- Fan, D.; Takawale, A.; Lee, J.; Kassiri, Z. Cardiac fibroblasts, fibrosis and extracellular matrix remodeling in heart disease. Fibrogenesis Tissue Repair 2012, 5, 15. [Google Scholar] [CrossRef] [Green Version]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ceramide | Solvent co | 0.7 µM Dox | FuB + Dox |
|---|---|---|---|
| Total Ceramides | 1.0 ± 0.07 | 1.39 ± 0.01 | 0.70 ± 0.11 |
| (C14–C24:1) | (p = 0.002 *) | (p < 0.001 *) | |
| (p < 0.001 #) | |||
| C14 | 1.0 ± 0.04 | 1.20 ± 0.01 | 0.21 ± 0.39 |
| (p = 0.003 *) | (p < 0.001 *) | ||
| (p < 0.001 #) | |||
| C16 | 1.0 ± 0.05 | 1.28 ± 0.02 | 1.03 ± 0.09 |
| (p = 0.002 *) | (p = 0.67) | ||
| (p = 0.001 #) | |||
| C18 | 1.0 ± 0.07 | 1.29 ± 0.02 | 1.04 ± 0.09 |
| (p = 0.006 *) | (p < 0.001 *) | ||
| (p < 0.001 #) | |||
| C20 | 1.0 ± 0.03 | 1.24 ± 0.02 | 0.12 ± 0.41 |
| (p < 0.001 *) | (p < 0.001 *) | ||
| (p < 0.001 #) | |||
| C22 | 1.0 ± 0.12 | 1.54 ± 0.03 | 0.72 ± 0.09 |
| (p = 0.004 *) | (p < 0.001 *) | ||
| (p < 0.001 #) | |||
| C22:1 | 1.0 ± 0.08 | 1.45 ± 0.03 | 0.35 ± 0.13 |
| (p = 0.002 *) | (p < 0.001 *) | ||
| (p < 0.001 #) | |||
| C24 | 1.0 ± 0.23 | 1.64 ± 0.02 | 0.69 ± 0.17 |
| (p = 0.02 *) | (p = 0.02 *) | ||
| (p < 0.001 #) | |||
| C24:1 | 1.0 ± 0.03 | 1.33 ± 0.02 | 0.52 ± 0.14 |
| (p < 0.001 *) | (p < 0.001 *) | ||
| (p < 0.001 #) |
| Gene | Solvent Co | 0.7 µM Dox | MT + Dox | FuB + Dox |
|---|---|---|---|---|
| MFN1 | 1.0 ± 0.09 | 0.80 ± 0.14 | 0.77 ± 0.17 | 0.73 ± 0.22 |
| (p = 0.007 *) | (p = 0.004 *) | (p = 0.002 *) | ||
| MFN2 | 1.0 ± 0.28 | 6.79 ± 0.40 | 6.65 ± 0.20 | 5.91 ± 0.23 |
| (p < 0.001 *) | (p < 0.001 *) | (p < 0.001 *) | ||
| OPA1 | 1.0 ± 0.16 | 0.66 ± 0.18 | 0.49 ± 0.10 | 0.51 ± 0.26 |
| (p = 0.002 *) | (p < 0.001 *) | (p < 0.001 *) | ||
| (p = 0.03 #) | ||||
| DRP1 | 1.0 ± 0.19 | 0.74 ± 0.21 | 0.65 ± 0.21 | 0.64 ± 0.12 |
| (p = 0.03 *) | (p = 0.004 *) | (p = 0.002 *) | ||
| Mff | 1.0 ± 0.18 | 0.90 ± 0.16 | 0.77 ± 0.15 | 0.63 ± 0.29 |
| (p = 0.02 *) | (p = 0.002 *) | |||
| (p = 0.03 #) | ||||
| FIS1 | 1.0 ± 0.15 | 0.89 ± 0.14 | 0.77 ± 0.10 | 0.68 ± 0.10 |
| (p = 0.01 *) | (p < 0.001 *) | |||
| (p = 0.02 #) |
| Gene | Solvent Co | 0.7 µM Dox | MT + Dox | FuB + Dox |
|---|---|---|---|---|
| DGAT1 | 1.0 ± 0.30 | 2.65 ± 0.25 | 2.86 ± 0.28 | 3.24 ± 0.47 |
| (p < 0.001 *) | (p < 0.001 *) | (p = 0.006 *) | ||
| DGAT2 | 1.0 ± 0.29 | 1.95 ± 0.22 | 1.44 ± 0.50 | 1.37 ± 0.24 |
| (p = 0.06 *) | ||||
| ATGL | 1.0 ± 0.38 | 3.02 ± 0.18 | 3.64 ± 0.22 | 2.26 ± 0.33 |
| (p < 0.001 *) | (p < 0.001 *) | (p = 0.01 *) | ||
| CD36 | 1.0 ± 0.58 | 4.02 ± 0.53 | 4.83 ± 0.24 | 3.35 ± 0.54 |
| (p < 0.001 *) | (p < 0.001 *) | (p = 0.002 *) | ||
| CPT1B | 1.0 ± 0.40 | 0.28 ± 0.38 | 0.20 ± 0.58 | 0.19 ± 0.82 |
| (p = 0.005 *) | (p = 0.006 *) | (p = 0.002 *) | ||
| PDK4 | 1.0 ± 0.25 | 0.55 ± 0.35 | 0.30 ± 0.25 | 0.46 ± 0.40 |
| (p = 0.01 *) | (p = 0.002 *) | (p = 0.008 *) | ||
| (p = 0.05 #) | ||||
| GDH | 1.0 ± 0.15 | 0.89 ± 0.35 | 1.20 ± 0.20 | 0.84 ± 0.27 |
| Gene | Solvent Co | 0.7 µM Dox | MT + Dox | FuB + Dox |
|---|---|---|---|---|
| MMP8 | 1.0 ± 0.32 | 3.65 ± 0.20 | 2.91 ± 0.49 | 1.46 ± 0.32 |
| (p < 0.001 *) | (p = 0.03 *) | (p = 0.001 #) | ||
| MMP9 | 1.0 ± 0.25 | 3.31 ± 0.52 | 2.72 ± 0.52 | 1.69 ± 0.53 |
| (p = 0.01 *) | (p = 0.03 *) | |||
| MMP14 | 1.0 ± 0.22 | 1.53 ± 0.24 | 1.20 ± 0.30 | 1.14 ± 0.41 |
| (p = 0.01 *) | ||||
| TIMP1 | 1.0 ± 0.18 | 1.63 ± 0.14 | 2.89 ± 0.33 | 1.86 ± 0.21 |
| (p = 0.005 *) | (p = 0.005 *) | (p = 0.006 *) | ||
| TIMP2 | 1.0 ± 0.04 | 1.66 ± 0.20 | 1.38 ± 0.29 | 1.32 ± 0.24 |
| (p = 0.01 *) | ||||
| TGF-β | 1.0 ± 0.14 | 2.26 ± 0.23 | 1.45 ± 0.37 | 1.24 ± 0.50 |
| (p = 0.01 *) | ||||
| ACTA2 | 1.0 ± 0.26 | 2.00 ± 0.32 | 2.12 ± 0.37 | 2.00 ± 0.28 |
| (p = 0.009 *) | (p = 0.02 *) | (p = 0.005 *) |
| Ceramide Species | FLAG-TC | FLAG-CerS2 | p Value |
|---|---|---|---|
| Total Ceramides (C14–C24:1) | 1.0 ± 0.29 | 2.12 ± 0.08 | 0.009 |
| C14 | 1.0 ± 0.36 | 1.32 ± 0.09 | 0.30 |
| C16 | 1.0 ± 0.82 | 2.83 ± 0.09 | 0.04 |
| C18 | 1.0 ± 0.16 | 2.27 ± 0.11 | 0.004 |
| C20 | 1.0 ± 0.11 | 1.28 ± 0.12 | 0.10 |
| C22 | 1.0 ± 0.19 | 2.90 ± 0.11 | 0.002 |
| C22:1 | 1.0 ± 0.18 | 0.92 ± 0.06 | 0.56 |
| C24 | 1.0 ± 0.29 | 2.89 ± 0.15 | 0.006 |
| C24:1 | 1.0 ± 0.24 | 1.51 ± 0.02 | 0.04 |
| Ceramide Species | DMSO+ FLAG-TC | FuB + FLAG-CerS2 | p Value |
|---|---|---|---|
| Total Ceramides (C14–C24:1) | 1.0 ± 0.34 | 0.85 ± 0.23 | 0.42 |
| C14 | 1.0 ± 0.20 | 0.56 ± 0.25 | 0.002 |
| C16 | 1.0 ± 0.32 | 0.90 ± 0.26 | 0.59 |
| C18 | 1.0 ± 0.18 | 0.97 ± 0.16 | 0.77 |
| C20 | 1.0 ± 0.13 | 1.11 ± 0.35 | 0.57 |
| C22 | 1.0 ± 0.31 | 0.90 ± 0.26 | 0.55 |
| C22:1 | 1.0 ± 0.05 | 1.03 ± 0.14 | 0.66 |
| C24 | 1.0 ± 0.47 | 0.64 ± 0.35 | 0.16 |
| C24:1 | 1.0 ± 0.40 | 0.88 ± 0.22 | 0.56 |
| Gene | FLAG-TC | FLAG-CerS2 | MT + FLAG-CerS2 | FuB + FLAG-CerS2 |
|---|---|---|---|---|
| MFN1 | 1.0 ± 0.01 | 0.86 ± 0.07 | 0.72 ± 0.25 | 0.30 ± 0.21 |
| (p = 0.04 *) | (p < 0.001 *) | |||
| (p < 0.001 #) | ||||
| MFN2 | 1.0 ± 0.11 | 0.68 ± 0.10 | 1.03 ± 0.11 | 1.14 ± 0.31 |
| (p = 0.02 *) | (p = 0.02 #) | |||
| OPA1 | 1.0 ± 0.08 | 0.82 ± 0.13 | 0.63 ± 0.14 | 0.51 ± 0.15 |
| (p = 0.01 *) | (p = 0.003 *) | |||
| (p = 0.03 #) | ||||
| DRP1 | 1.0 ± 0.05 | 0.89 ± 0.05 | 0.74 ± 0.18 | 0.44 ± 0.22 |
| (p = 0.002 *) | ||||
| (p = 0.004 #) | ||||
| Mff | 1.0 ± 0.06 | 0.79 ± 0.06 | 0.78 ± 0.16 | 0.72 ± 0.07 |
| (p = 0.02) | (p = 0.006 *) | |||
| FIS1 | 1.0 ± 0.08 | 0.89 ± 0.10 | 0.71 ± 0.13 | 0.63 ± 0.06 |
| (p = 0.03 *) | (p = 0.003 *) | |||
| (p = 0.02 #) |
| Gene | FLAG-TC | FLAG-CerS2 | MT + FLAG-CerS2 | FuB + FLAG-CerS2 |
|---|---|---|---|---|
| DGAT1 | 1.0 ± 0.21 | 1.03 ± 0.19 | 1.23 ± 0.21 | 0.67 ± 0.03 |
| (p = 0.03 #) | ||||
| DGAT2 | 1.0 ± 0.21 | 0.66 ± 0.31 | 0.45 ± 0.41 | 0.40 ± 0.44 |
| (p = 0.03 *) | (p = 0.01 *) | (p = 0.007 *) | ||
| ATGL | 1.0 ± 0.24 | 1.20 ± 0.37 | 0.80 ± 0.16 | 0.79 ± 0.05 |
| CD36 | 1.0 ± 0.85 | 3.61 ± 0.26 | 4.57 ± 0.34 | 1.25 ± 0.53 |
| (p < 0.001 *) | (p = 0.002 *) | (p = 0.01 #) | ||
| CPT1B | 1.0 ± 0.38 | 0.42 ± 0.65 | 0.34 ± 0.35 | 0.24 ± 0.67 |
| (p = 0.02 *) | (p = 0.05 *) | (p = 0.03 *) | ||
| PDK4 | 1.0 ± 0.38 | 0.86 ± 0.23 | 0.64 ± 0.26 | 0.85 ± 0.56 |
| GDH | 1.0 ± 0.24 | 0.92 ± 0.23 | 0.85 ± 0.12 | 0.72 ± 0.16 |
| Gene | FLAG-TC | FLAG-CerS2 | MT + FLAG-CerS2 | FuB + FLAG-CerS2 |
|---|---|---|---|---|
| MMP8 | 1.0 ± 0.17 | 2.55 ± 0.53 | 1.49 ± 0.36 | n.d. |
| (p = 0.03 *) | ||||
| MMP9 | 1.0 ± 0.36 | 1.45 ± 0.69 | 1.25 ± 0.41 | n.d. |
| MMP14 | 1.0 ± 0.33 | 0.91 ± 0.21 | 1.04 ± 0.10 | 7.17 ± 0.16 * 10−5 |
| (p = 0.004 *) | ||||
| (p < 0.001 #) | ||||
| TIMP1 | 1.0 ± 0.40 | 1.09 ± 0.52 | 1.12 ± 0.41 | 1.29 ± 0.07 |
| TIMP2 | 1.0 ± 0.09 | 0.93 ± 0.19 | 1.06 ± 0.07 | 0.96 ± 0.14 |
| TGF-β | 1.0 ± 0.18 | 1.23 ± 0.36 | 1.32 ± 0.39 | 1.72 ± 0.19 |
| (p = 0.03 *) | ||||
| ACTA2 | 1.0 ± 0.23 | 0.72 ± 0.14 | 1.25 ± 0.09 | 1.19 ± 0.13 |
| (p = 0.007 #) | (p = 0.02 #) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kretzschmar, T.; Bekhite, M.M.; Wu, J.M.F.; Haase, D.; Förster, M.; Müller, T.; Nietzsche, S.; Westermann, M.; Franz, M.; Gräler, M.H.; et al. Long-Chain and Very Long-Chain Ceramides Mediate Doxorubicin-Induced Toxicity and Fibrosis. Int. J. Mol. Sci. 2021, 22, 11852. https://doi.org/10.3390/ijms222111852
Kretzschmar T, Bekhite MM, Wu JMF, Haase D, Förster M, Müller T, Nietzsche S, Westermann M, Franz M, Gräler MH, et al. Long-Chain and Very Long-Chain Ceramides Mediate Doxorubicin-Induced Toxicity and Fibrosis. International Journal of Molecular Sciences. 2021; 22(21):11852. https://doi.org/10.3390/ijms222111852
Chicago/Turabian StyleKretzschmar, Tom, Mohamed M. Bekhite, Jasmine M. F. Wu, Daniela Haase, Martin Förster, Tina Müller, Sandor Nietzsche, Martin Westermann, Marcus Franz, Markus H. Gräler, and et al. 2021. "Long-Chain and Very Long-Chain Ceramides Mediate Doxorubicin-Induced Toxicity and Fibrosis" International Journal of Molecular Sciences 22, no. 21: 11852. https://doi.org/10.3390/ijms222111852
APA StyleKretzschmar, T., Bekhite, M. M., Wu, J. M. F., Haase, D., Förster, M., Müller, T., Nietzsche, S., Westermann, M., Franz, M., Gräler, M. H., & Schulze, P. C. (2021). Long-Chain and Very Long-Chain Ceramides Mediate Doxorubicin-Induced Toxicity and Fibrosis. International Journal of Molecular Sciences, 22(21), 11852. https://doi.org/10.3390/ijms222111852