
Application of Ligature-Induced Periodontitis in Mice to Explore the Molecular Mechanism of Periodontal Disease

 , ,
, ,  , and
, and
Abstract
1. Introduction
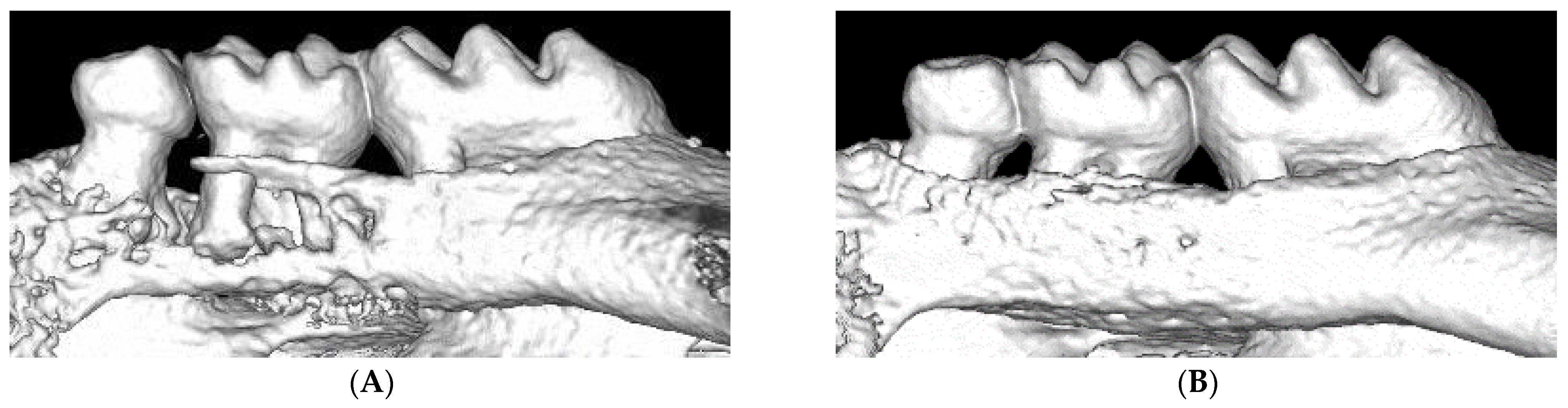
2. Ligature-Induced Periodontitis in Mice
3. Evaluation in Periodontal Tissues
3.1. Evaluation in Gingiva
3.2. Evaluation in Bone
3.2.1. Osteoclasts
3.2.2. Osteocytes
3.2.3. Others
3.3. Evaluation in Periodontal Ligament Cells
3.4. Summary
4. Ligature-Induced Periodontitis in Disease Model Mice
4.1. Diabetes and Obesity Model Mice with Ligature-Induced Periodontitis
4.2. Other Disease Model Mice with Ligature-Induced Periodontitis
4.3. Summary
5. Systemic Effects
5.1. Effect on the Cardiovascular and Blood System
5.2. Effect on the Central Nervous System
5.3. Effect on the Intestinal Flora and Digestive System
5.4. Summary
6. Immunology
6.1. Innate Immunity
6.1.1. Neutrophils
6.1.2. Macrophages
6.1.3. Lymphoid Cells
6.1.4. Solitary Chemosensory Cells
6.1.5. Receptors
6.1.6. Complements
6.2. Acquired Immunity
6.2.1. T Cells and IL-17
6.2.2. B Cells and IL-10
6.3. Cytokines, Molecules, and Genes
6.4. Summary
7. Microbiology
7.1. Oral and Gut Microbiota in Ligature-Induced Periodontitis Model
7.2. Summary
8. Ligature-Induced Periodontitis Models with Bacterial Factors
8.1. Periodontitis Model Induced by Ligature Inoculated with P. gingivalis
8.2. Periodontitis Model Induced by Ligation and Lipopolysaccharide of P. gingivalis
8.3. Periodontitis Model Induced by Ligation and Oral Gavage of P. gingivalis
8.4. Ligature-Induced Periodontitis Model Affected by Smoking Factors
8.5. Summary
9. New Treatment Methods
9.1. Effects of Systemic Antibiotics
9.2. Effects of Peptides or Proteins
9.3. Effects of Natural and Synthetic Compounds
9.4. Effects of Extracts from Plants
9.5. Effects of Treatment Instrumentation
9.6. Effects of Antibody, Cell, Cytokine etc.
9.7. Summary
10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pihlstrom, B.L.; Michalowicz, B.S.; Johnson, N.W. Periodontal diseases. Lancet 2005, 366, 1809–1820. [Google Scholar] [CrossRef]
- Kinane, D.F.; Stathopoulou, P.G.; Papapanou, P.N. Periodontal diseases. Nat. Rev. Dis. Primers 2017, 3, 1–14. [Google Scholar] [CrossRef]
- Lang, N.P.; Lindhe, J. Clinical Periodontology and Implant Dentistry, 6th ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2015; 1480p, ISBN 978-0-470-67248-8. [Google Scholar]
- Graves, D.T.; Fine, D.; Teng, Y.-T.A.; Van Dyke, T.E.; Hajishengallis, G. The use of rodent models to investigate host-bacteria interactions related to periodontal diseases. J. Clin. Periodontol. 2008, 35, 89–105. [Google Scholar] [CrossRef]
- Marchesan, J.; Girnary, M.S.; Jing, L.; Miao, M.Z.; Zhang, S.; Sun, L.; Morelli, T.; Schoenfisch, M.H.; Inohara, N.; Offenbacher, S.; et al. An experimental murine model to study periodontitis. Nat. Protoc. 2018, 13, 2247–2267. [Google Scholar] [CrossRef] [PubMed]
- de Molon, R.S.; de Avila, E.D.; Boas Nogueira, A.V.; Chaves de Souza, J.A.; Avila-Campos, M.J.; de Andrade, C.R.; Cirelli, J.A. Evaluation of the host response in various models of induced periodontal disease in mice. J. Periodontol. 2014, 85, 465–477. [Google Scholar] [CrossRef]
- Wong, R.L.; Hiyari, S.; Yaghsezian, A.; Davar, M.; Lin, Y.-L.; Galvan, M.; Tetradis, S.; Camargo, P.M.; Pirih, F.Q. Comparing the Healing Potential of Late-Stage Periodontitis and Peri-Implantitis. J. Oral Implant. 2017, 43, 437–445. [Google Scholar] [CrossRef] [PubMed]
- Abe, T.; Hajishengallis, G. Optimization of the ligature-induced periodontitis model in mice. J. Immunol. Methods 2013, 394, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Akkaoui, J.; Yamada, C.; Duarte, C.; Ho, A.; Vardar-Sengul, S.; Kawai, T.; Movila, A. Contribution of Porphyromonas gingivalis lipopolysaccharide to experimental periodontitis in relation to aging. GeroScience 2021, 43, 367–376. [Google Scholar] [CrossRef]
- Qi, W.; Yang, X.; Ye, N.; Li, S.; Han, Q.; Huang, J.; Wu, B. TLR4 gene in the regulation of periodontitis and its molecular mechanism. Exp. Ther. Med. 2019, 18, 1961–1966. [Google Scholar] [CrossRef] [PubMed]
- Fine, N.; Chadwick, J.; Sun, C.; Parbhakar, K.; Khoury, N.; Barbour, A.; Goldberg, M.; Tenenbaum, H.; Glogauer, M. Periodontal Inflammation Primes the Systemic Innate Immune Response. J. Dent. Res. 2021, 100, 318–325. [Google Scholar] [CrossRef] [PubMed]
- Ohnishi, T.; Okamoto, A.; Kakimoto, K.; Bandow, K.; Chiba, N.; Matsuguchi, T. Involvement of Cot/Tp12 in Bone Loss during Periodontitis. J. Dent. Res. 2010, 89, 192–197. [Google Scholar] [CrossRef]
- Rojas, C.; García, M.P.; Polanco, A.F.; González-Osuna, L.; Sierra-Cristancho, A.; Melgar-Rodríguez, S.; Cafferata, E.A.; Vernal, R. Humanized Mouse Models for the Study of Periodontitis: An Opportunity to Elucidate Unresolved Aspects of Its Immunopathogenesis and Analyze New Immunotherapeutic Strategies. Front. Immunol. 2021, 12, 663328. [Google Scholar] [CrossRef]
- Sartori, R.; Li, F.; Kirkwood, K. MAP Kinase Phosphatase-1 Protects against Inflammatory Bone Loss. J. Dent. Res. 2009, 88, 1125–1130. [Google Scholar] [CrossRef] [PubMed]
- Hart, G.T.; Shaffer, D.J.; Akilesh, S.; Brown, A.C.; Moran, L.; Roopenian, D.C.; Baker, P.J. Quantitative Gene Expression Profiling Implicates Genes for Susceptibility and Resistance to Alveolar Bone Loss. Infect. Immun. 2004, 72, 4471–4479. [Google Scholar] [CrossRef] [PubMed]
- Cekici, A.; Kantarci, A.; Hasturk, H.; Van Dyke, T.E. Inflammatory and immune pathways in the pathogenesis of periodontal disease. Periodontology 2000 2014, 64, 57–80. [Google Scholar] [CrossRef] [PubMed]
- Tamura, H.; Maekawa, T.; Hiyoshi, T.; Terao, Y. Analysis of Experimental Ligature-Induced Periodontitis Model in Mice. In Periodontal Pathogens: Methods and Protocols; Humana Inc.: New York, NY, USA, 2021; pp. 237–250. [Google Scholar] [CrossRef]
- Fujita, T.; Yoshimoto, T.; Kajiya, M.; Ouhara, K.; Matsuda, S.; Takemura, T.; Akutagawa, K.; Takeda, K.; Mizuno, N.; Kurihara, H. Regulation of defensive function on gingival epithelial cells can prevent periodontal disease. Jpn. Dent. Sci. Rev. 2018, 54, 66–75. [Google Scholar] [CrossRef]
- Boivin, F.J.; Schmidt-Ott, K.M. Transcriptional mechanisms coordinating tight junction assembly during epithelial differentiation. Ann. N. Y. Acad. Sci. 2017, 1397, 80–99. [Google Scholar] [CrossRef]
- Meyle, J.; Guttig, K.; Rascher, G.; Wolburg, H. Transepithelial electrical resistance and tight junctions of human gingival keratinocvtes. J. Periodontal Res. 1999, 34, 214–222. [Google Scholar] [CrossRef]
- Wang, S.; Samakovlis, C. Grainy Head and Its Target Genes in Epithelial Morphogenesis and Wound Healing. Curr. Top. Dev. Biol. 2012, 98, 35–63. [Google Scholar] [CrossRef]
- Mlacki, M.; Darido, C.; Jane, S.M.; Wilanowski, T. Loss of Grainy Head-Like 1 Is Associated with Disruption of the Epidermal Barrier and Squamous Cell Carcinoma of the Skin. PLoS ONE 2014, 9, e89247. [Google Scholar] [CrossRef]
- Aue, A.; Hinze, C.; Walentin, K.; Ruffert, J.; Yurtdas, Y.; Werth, M.; Chen, W.; Rabien, A.; Kilic, E.; Schulzke, J.-D.; et al. A Grainyhead-Like 2/Ovo-Like 2 Pathway Regulates Renal Epithelial Barrier Function and Lumen Expansion. J. Am. Soc. Nephrol. 2015, 26, 2704–2715. [Google Scholar] [CrossRef]
- Sumigray, K.; Lechler, T. Cell Adhesion in Epidermal Development and Barrier Formation. Curr. Top. Dev. Biol. 2015, 112, 383–414. [Google Scholar] [CrossRef]
- Mehrazarin, S.; Chen, W.; Oh, J.-E.; Liu, Z.X.; Kang, K.L.; Yi, J.K.; Kim, R.H.; Shin, K.-H.; Park, N.-H.; Kang, M.K. The p63 Gene Is Regulated by Grainyhead-like 2 (GRHL2) through Reciprocal Feedback and Determines the Epithelial Phenotype in Human Keratinocytes. J. Biol. Chem. 2015, 290, 19999–20008. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Yi, J.K.; Shimane, T.; Mehrazarin, S.; Lin, Y.-L.; Shin, K.-H.; Kim, R.H.; Park, N.-H.; Kang, M.K. Grainyhead-like 2 regulates epithelial plasticity and stemness in oral cancer cells. Carcinogenesis 2016, 37, 500–510. [Google Scholar] [CrossRef]
- Chen, W.; AlShaikh, A.; Kim, S.; Kim, J.; Chun, C.; Mehrazarin, S.; Lee, J.; Lux, R.; Kim, R.; Shin, K.; et al. Porphyromonas gingivalis Impairs Oral Epithelial Barrier through Targeting GRHL2. J. Dent. Res. 2019, 98, 1150–1158. [Google Scholar] [CrossRef]
- Jin, Y.-R.; Stohn, J.P.; Wang, Q.; Nagano, K.; Baron, R.; Bouxsein, M.L.; Rosen, C.J.; Adarichev, V.A.; Lindner, V. Inhibition of osteoclast differentiation and collagen antibody-induced arthritis by CTHRC1. Bone 2017, 97, 153–167. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Yang, Q.; Sun, H. Collagen triple helix repeat containing-1: A novel biomarker associated with disease activity in Systemic lupus erythematosus. Lupus 2018, 27, 2076–2085. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.K.; Li, Y.M.; Li, Y.; Wei, Y.R.; Zhang, J.; Li, B.; You, Z.R.; Chen, Y.; Huang, B.Y.; Miao, Q.; et al. CTHRC1 expression in primary biliary cholangitis. J. Dig. Dis. 2019, 20, 371–376. [Google Scholar] [CrossRef]
- Guo, Y.; Jiang, C.; Yao, S.; Ma, L.; Zhang, H.; Wang, X.; Xu, S.; Cao, Z. CTHRC1 Knockdown Promotes Inflammatory Responses Partially by p38 MAPK Activation in Human Periodontal Ligament Cells. Inflammation 2021, in press. [Google Scholar] [CrossRef] [PubMed]
- Movila, A.; Kajiya, M.; Wisitrasameewong, W.; Stashenko, P.; Vardar-Sengul, S.; Hernandez, M.; Temple, H.T.; Kawai, T. Intravital endoscopic technology for real-time monitoring of inflammation caused in experimental periodontitis. J. Immunol. Methods 2018, 457, 26–29. [Google Scholar] [CrossRef]
- Laukkanen, M.O.; Castellone, M.D. Gastrin-Releasing Peptide Receptor Targeting in Cancer Treatment: Emerging Signaling Networks and Therapeutic Applications. Curr. Drug Targets 2016, 17, 508–514. [Google Scholar] [CrossRef] [PubMed]
- Cornelio, D.; Roesler, R.; Schwartsmann, G. Gastrin-releasing peptide receptor as a molecular target in experimental anticancer therapy. Ann. Oncol. 2007, 18, 1457–1466. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.; Heo, S.C.; Na Kim, Y.; Joo, J.-Y.; Hwang, J.J.; Bae, M.-K.; Kim, H.J. Gastrin-Releasing Peptide (GRP) Stimulates Osteoclastogenesis in Periodontitis. Cells 2020, 10, 50. [Google Scholar] [CrossRef] [PubMed]
- Trackman, P.C.; Kantarci, A. Connective Tissue Metabolism and Gingival Overgrowth. Crit. Rev. Oral Biol. Med. 2004, 15, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Trackman, P.; Kantarci, A. Molecular and clinical aspects of drug-induced gingival overgrowth. J. Dent. Res. 2015, 94, 540–546. [Google Scholar] [CrossRef]
- Okanobu, A.; Matsuda, S.; Kajiya, M.; Fujita, T.; Kittaka, M.; Shiba, H.; Kurihara, H. A novel gingival overgrowth mouse model induced by the combination of CsA and ligature-induced inflammation. J. Immunol. Methods 2017, 445, 31–36. [Google Scholar] [CrossRef]
- Bao, K.; Li, X.; Kajikawa, T.; Toshiharu, A.; Selevsek, N.; Grossmann, J.; Hajishengallis, G.; Bostanci, N. Pressure Cycling Technology Assisted Mass Spectrometric Quantification of Gingival Tissue Reveals Proteome Dynamics during the Initiation and Progression of Inflammatory Periodontal Disease. Proteomics 2019, 20, e1900253. [Google Scholar] [CrossRef]
- Huang, W.; Zhan, Y.; Zheng, Y.; Han, Y.; Hu, W.; Hou, J. Up-regulated ferritin in periodontitis promotes inflammatory cytokine expression in human periodontal ligament cells through transferrin receptor via ERK/P38 MAPK pathways. Clin. Sci. 2019, 133, 135–148. [Google Scholar] [CrossRef]
- Chapple, I.L.C.; Matthews, J.B. The role of reactive oxygen and antioxidant species in periodontal tissue destruction. Periodontology 2000 2007, 43, 160–232. [Google Scholar] [CrossRef]
- Circu, M.L.; Aw, T.Y. Reactive oxygen species, cellular redox systems, and apoptosis. Free Radic. Biol. Med. 2010, 48, 749–762. [Google Scholar] [CrossRef] [PubMed]
- Kataoka, K.; Ekuni, D.; Tomofuji, T.; Irie, K.; Kunitomo, M.; Uchida, Y.; Fukuhara, D.; Morita, M. Visualization of Oxidative Stress Induced by Experimental Periodontitis in Keap1-Dependent Oxidative Stress Detector-Luciferase Mice. Int. J. Mol. Sci. 2016, 17, 1907. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, T.; Hayashi, M.; Takayanagi, H. New insights into osteoclastogenic signaling mechanisms. Trends Endocrinol. Metab. 2012, 23, 582–590. [Google Scholar] [CrossRef]
- Miyamoto, H.; Suzuki, T.; Miyauchi, Y.; Iwasaki, R.; Kobayashi, T.; Sato, Y.; Miyamoto, K.; Hoshi, H.; Hashimoto, K.; Yoshida, S.; et al. Osteoclast stimulatory transmembrane protein and dendritic cell-specific transmembrane protein cooperatively modulate cell-cell fusion to form osteoclasts and foreign body giant cells. J. Bone Miner. Res. 2012, 27, 1289–1297. [Google Scholar] [CrossRef]
- Ishii, T.; Ruiz-Torruella, M.; Ikeda, A.; Shindo, S.; Movila, A.; Mawardi, H.; Albassam, A.; Kayal, R.A.; Al-Dharrab, A.A.; Egashira, K.; et al. OC-STAMP promotes osteoclast fusion for pathogenic bone resorption in periodontitis via up-regulation of permissive fusogen CD9. FASEB J. 2018, 32, 4016–4030. [Google Scholar] [CrossRef] [PubMed]
- Akutagawa, K.; Fujita, T.; Ouhara, K.; Takemura, T.; Tari, M.; Kajiya, M.; Matsuda, S.; Kuramitsu, S.; Mizuno, N.; Shiba, H.; et al. Glycyrrhizic acid suppresses inflammation and reduces the increased glucose levels induced by the combination of Porphyromonas gulae and ligature placement in diabetic model mice. Int. Immunopharmacol. 2019, 68, 30–38. [Google Scholar] [CrossRef]
- Yoshihara-Hirata, C.; Yamashiro, K.; Yamamoto, T.; Aoyagi, H.; Ideguchi, H.; Kawamura, M.; Suzuki, R.; Ono, M.; Wake, H.; Nishibori, M.; et al. Anti-HMGB1 Neutralizing Antibody Attenuates Periodontal Inflammation and Bone Resorption in a Murine Periodontitis Model. Infect. Immun. 2018, 86, e00111-18. [Google Scholar] [CrossRef]
- Coronas-Serna, J.; Valenti, M.; Del Val, E.; Fernández-Acero, T.; Rodríguez-Escudero, I.; Mingo, J.; Luna, S.; Torices, L.; Pulido, R.; Molina, M.; et al. Modeling human disease in yeast: Recreating the PI3K-PTEN-Akt signaling pathway in Saccharomyces cerevisiae. Int. Microbiol. 2020, 23, 75–87. [Google Scholar] [CrossRef]
- Fu, C.; Wei, Z.; Zhang, D. PTEN Inhibits Inflammatory Bone Loss in Ligature-Induced Periodontitis via IL1 and TNF-α. BioMed Res. Int. 2019, 2019, 6712591. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhang, X.; Yang, Q.; Zhu, X.; Chen, F.; Yue, J.; Zhou, R.; Xu, Y.; Qi, S. Knockout of NRAGE promotes autophagy-related gene expression and the periodontitis process in mice. Oral Dis. 2021, 27, 589–599. [Google Scholar] [CrossRef] [PubMed]
- Bouchon, A.; Facchetti, F.; Weigand, M.A.; Colonna, M. TREM-1 amplifies inflammation and is a crucial mediator of septic shock. Nat. Cell Biol. 2001, 410, 1103–1107. [Google Scholar] [CrossRef]
- Bostanci, N.; Abe, T.; Belibasakis, G.N.; Hajishengallis, G. TREM-1 Is Upregulated in Experimental Periodontitis, and Its Blockade Inhibits IL-17A and RANKL Expression and Suppresses Bone loss. J. Clin. Med. 2019, 8, 1579. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.Y.; Feng, L.; Park, H.-T.; Havlioglu, N.; Wen, L.; Tang, H.; Bacon, K.B.; Jiang, Z.-H.; Zhang, X.-C.; Rao, Y. The neuronal repellent Slit inhibits leukocyte chemotaxis induced by chemotactic factors. Nat. Cell Biol. 2001, 410, 948–952. [Google Scholar] [CrossRef]
- Wang, L.; Zheng, J.; Pathak, J.L.; Chen, Y.; Liang, D.; Yang, L.; Sun, H.; Zhong, M.; Wu, L.; Li, L.; et al. SLIT2 Overexpression in Periodontitis Intensifies Inflammation and Alveolar Bone Loss, Possibly via the Activation of MAPK Pathway. Front. Cell Dev. Biol. 2020, 8, 593. [Google Scholar] [CrossRef]
- Mukai, T.; Ishida, S.; Ishikawa, R.; Yoshitaka, T.; Kittaka, M.; Gallant, R.; Lin, Y.-L.; Rottapel, R.; Brotto, M.; Reichenberger, E.J.; et al. SH3BP2 Cherubism Mutation Potentiates TNF-α-Induced Osteoclastogenesis via NFATc1 and TNF-α-Mediated Inflammatory Bone Loss. J. Bone Miner. Res. 2014, 29, 2618–2635. [Google Scholar] [CrossRef]
- Kittaka, M.; Yoshimoto, T.; Schlosser, C.; Rottapel, R.; Kajiya, M.; Kurihara, H.; Reichenberger, E.J.; Ueki, Y. Alveolar Bone Protection by Targeting the SH3BP2-SYK Axis in Osteoclasts. J. Bone Miner. Res. 2020, 35, 382–395. [Google Scholar] [CrossRef]
- Izawa, T.; Arakaki, R.; Mori, H.; Tsunematsu, T.; Kudo, Y.; Tanaka, E.; Ishimaru, N. The Nuclear Receptor AhR Controls Bone Homeostasis by Regulating Osteoclast Differentiation via the RANK/c-Fos Signaling Axis. J. Immunol. 2016, 197, 4639–4650. [Google Scholar] [CrossRef]
- Huang, J.; Cai, X.; Ou, Y.; Fan, L.; Zhou, Y.; Wang, Y. Protective roles of FICZ and aryl hydrocarbon receptor axis on alveolar bone loss and inflammation in experimental periodontitis. J. Clin. Periodontol. 2019, 46, 882–893. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Floyd, D.H.; Hughes, A.; Xiang, J.; Schneider, J.G.; Uluckan, O.; Heller, E.; Deng, H.; Zou, W.; Craft, C.S.; et al. The ADP receptor P2RY12 regulates osteoclast function and pathologic bone remodeling. J. Clin. Investig. 2012, 122, 3579–3592. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Kajikawa, T.; Walsh, M.C.; Takegahara, N.; Jeong, Y.H.; Hajishengallis, G.; Choi, Y. The purinergic receptor P2X5 contributes to bone loss in experimental periodontitis. BMB Rep. 2018, 51, 468–473. [Google Scholar] [CrossRef]
- Chen, Y.; Yang, Q.; Lv, C.; Chen, Y.; Zhao, W.; Li, W.; Chen, H.; Wang, H.; Sun, W.; Yuan, H. NLRP3 regulates alveolar bone loss in ligature-induced periodontitis by promoting osteoclastic differentiation. Cell Prolif. 2021, 54, e12973. [Google Scholar] [CrossRef] [PubMed]
- Tominaga, M.; Tominaga, T. Structure and function of TRPV1. Pflugers Arch. 2005, 451, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, N.; Matsuda, Y.; Sato, K.; de Jong, P.; Bertin, S.; Tabeta, K.; Yamazaki, K. Neuronal TRPV1 activation regulates alveolar bone resorption by suppressing osteoclastogenesis via CGRP. Sci. Rep. 2016, 6, 29294. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Tang, Y.; Zhu, X.; Tu, T.; Sui, L.; Han, Q.; Yu, L.; Meng, S.; Zheng, L.; Valverde, P.; et al. Overexpression of MiR-335-5p Promotes Bone Formation and Regeneration in Mice. J. Bone Miner. Res. 2017, 32, 2466–2475. [Google Scholar] [CrossRef]
- Lian, J.; Wu, X.; Liu, Y.; Qiu, W.; Zhu, X.; Wang, X.; Meng, S.; Valverde, P.; Steffensen, B.; Tu, Q.; et al. Potential roles of miR-335-5p on pathogenesis of experimental periodontitis. J. Periodontal Res. 2020, 55, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Tsukasaki, M.; Takayanagi, H. Osteoimmunology: Evolving concepts in bone–immune interactions in health and disease. Nat. Rev. Immunol. 2019, 19, 626–642. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhang, Y.; Kang, H.; Liu, W.; Liu, P.; Zhang, J.; Harris, S.E.; Wu, D. Sclerostin binds to LRP5/6 and antagonizes canonical Wnt signaling. J. Biol. Chem. 2005, 280, 19883–19887. [Google Scholar] [CrossRef]
- Goes, P.; Dutra, C.; Lösser, L.; Hofbauer, L.C.; Rauner, M.; Thiele, S. Loss of Dkk-1 in Osteocytes Mitigates Alveolar Bone Loss in Mice with Periodontitis. Front. Immunol. 2019, 10, 2924. [Google Scholar] [CrossRef] [PubMed]
- Hermann, P.; Gera, I.; Borbély, J.; Fejérdy, P.; Madléna, M. Periodontal health of an adult population in Hungary: Findings of a national survey. J. Clin. Periodontol. 2009, 36, 449–457. [Google Scholar] [CrossRef]
- Duan, X.; Gleason, R.C.; Hajishengallis, G.; Hosur, K.B.; Huang, D.; Wang, H.; Hajishengallis, G.; Liang, S. Sex dimorphism in periodontitis in animal models. J. Periodontal Res. 2016, 51, 196–202. [Google Scholar] [CrossRef]
- Aung, K.T.; Akiyama, K.; Kunitomo, M.; Mun, A.Y.; Tosa, I.; Nguyen, H.T.T.; Zhang, J.; Kohno, T.; Ono, M.; Hara, E.S.; et al. Aging-Affected MSC Functions and Severity of Periodontal Tissue Destruction in a Ligature-Induced Mouse Periodontitis Model. Int. J. Mol. Sci. 2020, 21, 8103. [Google Scholar] [CrossRef]
- Wong, R.L.; Hiyari, S.; Yaghsezian, A.; Davar, M.; Casarin, M.; Lin, Y.-L.; Tetradis, S.; Camargo, P.M.; Pirih, F.Q. Early intervention of peri-implantitis and periodontitis using a mouse model. J. Periodontol. 2018, 89, 669–679. [Google Scholar] [CrossRef] [PubMed]
- Trombetta-Esilva, J.; A Rosset, E.; Hepfer, R.G.; Wright, G.J.; Baicu, C.; Yao, H.; Bradshaw, A.D. Decreased Mechanical Strength and Collagen Content in SPARC-Null Periodontal Ligament Is Reversed by Inhibition of Transglutaminase Activity. J. Bone Miner. Res. 2015, 30, 1914–1924. [Google Scholar] [CrossRef]
- Rosset, E.M.; Trombetta-Esilva, J.; Hepfer, G.; Chen, P.; Yao, H.; Bradshaw, A.D. Inhibition of transglutaminase activity in periodontitis rescues periodontal ligament collagen content and architecture. J. Periodontal Res. 2020, 55, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Han, D.; Yuan, Z.; Hu, H.; Zhao, Z.; Yang, R.; Jin, Y.; Zou, C.; Chen, Y.; Wang, G.; et al. Long non-coding RNA H19 confers 5-Fu resistance in colorectal cancer by promoting SIRT1-mediated autophagy. Cell Death Dis. 2018, 9, 1–14. [Google Scholar] [CrossRef]
- Yu, T.-X.; Chung, H.K.; Xiao, L.; Piao, J.-J.; Lan, S.; Jaladanki, S.K.; Turner, D.J.; Raufman, J.-P.; Gorospe, M.; Wang, J.-Y. Long Noncoding RNA H19 Impairs the Intestinal Barrier by Suppressing Autophagy and Lowering Paneth and Goblet Cell Function. Cell. Mol. Gastroenterol. Hepatol. 2020, 9, 611–625. [Google Scholar] [CrossRef]
- An, Y.; Liu, W.; Xue, P.; Zhang, Y.; Wang, Q.; Jin, Y. Increased autophagy is required to protect periodontal ligament stem cells from apoptosis in inflammatory microenvironment. J. Clin. Periodontol. 2016, 43, 618–625. [Google Scholar] [CrossRef]
- Jiang, M.; Li, Z.; Zhu, G. The role of autophagy in the pathogenesis of periodontal disease. Oral Dis. 2020, 26, 259–269. [Google Scholar] [CrossRef] [PubMed]
- Guo, R.; Huang, Y.; Liu, H.; Zheng, Y.; Jia, L.; Li, W. Long Non-Coding RNA H19 Participates in Periodontal Inflammation via Activation of Autophagy. J. Inflamm. Res. 2020, 13, 635–646. [Google Scholar] [CrossRef]
- Genco, R.J.; Sanz, M. Clinical and public health implications of periodontal and systemic diseases: An overview. Periodontology 2000 2020, 83, 7–13. [Google Scholar] [CrossRef]
- Morita, I.; Inagaki, K.; Nakamura, F.; Noguchi, T.; Matsubara, T.; Yoshii, S.; Nakagaki, H.; Mizuno, K.; Sheiham, A.; Sabbah, W. Relationship between Periodontal Status and Levels of Glycated Hemoglobin. J. Dent. Res. 2012, 91, 161–166. [Google Scholar] [CrossRef]
- Demmer, R.; Holtfreter, B.; Desvarieux, M.; Jacobs, D.R.; Kerner, W.; Nauck, M.; Völzke, H.; Kocher, T. The Influence of Type 1 and Type 2 Diabetes on Periodontal Disease Progression: Prospective results from the Study of Health in Pomerania (SHIP). Diabetes Care 2012, 35, 2036–2042. [Google Scholar] [CrossRef]
- Saito, T.; Shimazaki, Y.; Sakamoto, M. Obesity and Periodontitis. New Engl. J. Med. 1998, 339, 482–483. [Google Scholar] [CrossRef]
- Katagiri, S.; Nitta, H.; Nagasawa, T.; Izumi, Y.; Kanazawa, M.; Matsuo, A.; Chiba, H.; Miyazaki, S.; Miyauchi, T.; Nakamura, N.; et al. High prevalence of periodontitis in non-elderly obese Japanese adults. Obes. Res. Clin. Pract. 2010, 4, e301–e306. [Google Scholar] [CrossRef]
- Maekawa, S.; Katagiri, S.; Takeuchi, Y.; Komazaki, R.; Ohtsu, A.; Udagawa, S.; Izumi, Y. Bone metabolic microarray analysis of ligature-induced periodontitis in streptozotocin-induced diabetic mice. J. Periodontal Res. 2017, 52, 233–245. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.; Zhao, L.; Huang, X.; Xie, B.; Zhang, J.; Xuan, D. Aberrant Periodontal and Systemic Immune Response of Overweight Rodents to Periodontal Infection. BioMed Res. Int. 2019, 2019, 1–10. [Google Scholar] [CrossRef]
- Huang, X.; Yu, T.; Ma, C.; Wang, Y.; Xie, B.; Xuan, D.; Zhang, J. Macrophages Play a Key Role in the Obesity-Induced Periodontal Innate Immune Dysfunction via Nucleotide-Binding Oligomerization Domain-Like Receptor Protein 3 Pathway. J. Periodontol. 2016, 87, 1195–1205. [Google Scholar] [CrossRef]
- Yu, T.; Zhao, L.; Huang, X.; Xie, M.; Wang, X.; Ma, C.; Xu, Y.; Wang, Y.; Xie, B.; Luo, G.; et al. Postoperative Weight Loss Masks Metabolic Impacts of Periodontitis in Obese Rodents. J. Periodontol. 2017, 88, e97–e108. [Google Scholar] [CrossRef]
- Gyurko, R.; Siqueira, C.C.; Caldon, N.; Gao, L.; Kantarci, A.; Van Dyke, T.E. Chronic Hyperglycemia Predisposes to Exaggerated Inflammatory Response and Leukocyte Dysfunction in Akita Mice. J. Immunol. 2006, 177, 7250–7256. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, Q. Periodontitis aggravated pancreatic β-cell dysfunction in diabetic mice through interleukin-12 regulation on Klotho. J. Diabetes Investig. 2016, 7, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Xu, T.; Liang, Z.; Zhao, L.; Xiong, X.; Xie, K.; Yu, W.; Zeng, X.; Gao, J.; Zhou, Y.; et al. Untargeted and targeted gingival metabolome in rodents reveal metabolic links between high-fat diet-induced obesity and periodontitis. J. Clin. Periodontol. 2021, 48, 1137–1148. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Yamazaki, K.; Kato, T.; Nakanishi, Y.; Tsuzuno, T.; Yokoji-Takeuchi, M.; Yamada-Hara, M.; Miura, N.; Okuda, S.; Ohno, H.; et al. Obesity-Related Gut Microbiota Aggravates Alveolar Bone Destruction in Experimental Periodontitis through Elevation of Uric Acid. mBio 2021, 12, e0077121. [Google Scholar] [CrossRef]
- Kantarci, A.; Tognoni, C.M.; Yaghmoor, W.; Marghalani, A.; Stephens, D.; Ahn, J.-Y.; Carreras, I.; Dedeoglu, A. Microglial response to experimental periodontitis in a murine model of Alzheimer’s disease. Sci. Rep. 2020, 10, 18561. [Google Scholar] [CrossRef]
- Qian, X.; Zhang, S.; Duan, L.; Yang, F.; Zhang, K.; Yan, F.; Ge, S. Periodontitis Deteriorates Cognitive Function and Impairs Neurons and Glia in a Mouse Model of Alzheimer’s Disease. J. Alzheimer’s Dis. 2021, 79, 1785–1800. [Google Scholar] [CrossRef] [PubMed]
- Orlandi, M.; Graziani, F.; D’Aiuto, F. Periodontal therapy and cardiovascular risk. Periodontology 2000 2020, 83, 107–124. [Google Scholar] [CrossRef] [PubMed]
- Schenkein, H.A.; Papapanou, P.N.; Genco, R.; Sanz, M. Mechanisms underlying the association between periodontitis and atherosclerotic disease. Periodontology 2000 2020, 83, 90–106. [Google Scholar] [CrossRef]
- Suh, J.S.; Lee, S.H.; Fouladian, Z.; Lee, J.Y.; Kim, T.; Kang, M.K.; Lusis, A.J.; Boström, K.I.; Kim, R.H.; Park, N.-H. Rosuvastatin Prevents the Exacerbation of Atherosclerosis in Ligature-Induced Periodontal Disease Mouse Model. Sci. Rep. 2020, 10, 6383. [Google Scholar] [CrossRef]
- O’Boyle, C.; Haley, M.J.; LeMarchand, E.; Smith, C.J.; Allan, S.M.; E Konkel, J.; Lawrence, C.B. Ligature-induced periodontitis induces systemic inflammation but does not alter acute outcome after stroke in mice. Int. J. Stroke 2020, 15, 175–187. [Google Scholar] [CrossRef]
- Kim, T.; Kim, S.; Song, M.; Lee, C.; Yagita, H.; Williams, D.W.; Sung, E.C.; Hong, C.; Shin, K.-H.; Kang, M.K.; et al. Removal of Pre-Existing Periodontal Inflammatory Condition before Tooth Extraction Ameliorates Medication-Related Osteonecrosis of the Jaw–Like Lesion in Mice. Am. J. Pathol. 2018, 188, 2318–2327. [Google Scholar] [CrossRef]
- Handa, K.; Abe, S.; Suresh, V.V.; Fujieda, Y.; Ishikawa, M.; Orimoto, A.; Kobayashi, Y.; Yamada, S.; Yamaba, S.; Murakami, S.; et al. Fibrillin-1 insufficiency alters periodontal wound healing failure in a mouse model of Marfan syndrome. Arch. Oral Biol. 2018, 90, 53–60. [Google Scholar] [CrossRef]
- Coyac, B.; Falgayrac, G.; Baroukh, B.; Slimani, L.; Sadoine, J.; Penel, G.; Duplan, M.B.; Schinke, T.; Linglart, A.; McKee, M.D.; et al. Tissue-specific mineralization defects in the periodontium of the Hyp mouse model of X-linked hypophosphatemia. Bone 2017, 103, 334–346. [Google Scholar] [CrossRef] [PubMed]
- Candeo, L.C.; Rigonato-Oliveira, N.C.; Brito, A.; Marcos, R.L.; Franca, C.; Fernandes, K.; Mesquita-Ferrari, R.A.; Bussadori, S.; Vieira, R.P.; Lino-Dos-Santos-Franco, A.; et al. Effects of periodontitis on the development of asthma: The role of photodynamic therapy. PLoS ONE 2017, 12, e0187945. [Google Scholar] [CrossRef]
- Rosa, E.P.; Murakami-Malaquias-Da-Silva, F.; Palma-Cruz, M.; Garcia, G.D.C.; De Brito, A.A.; Andreo, L.; Kamei, S.K.; Negreiros, R.M.; Rodrigues, M.F.D.S.S.D.; Mesquita-Ferrari, R.A.; et al. The impact of periodontitis in the course of chronic obstructive pulmonary disease: Pulmonary and systemic effects. Life Sci. 2020, 261, 118257. [Google Scholar] [CrossRef]
- Ide, M.; Papapanou, P.N. Epidemiology of association between maternal periodontal disease and adverse pregnancy outcomes—Systematic review. J. Clin. Periodontol. 2013, 40, S181–S194. [Google Scholar] [CrossRef] [PubMed]
- Ye, C.; Katagiri, S.; Miyasaka, N.; Bharti, P.; Kobayashi, H.; Takeuchi, Y.; Momohara, Y.; Sekiguchi, M.; Takamine, S.; Nagasawa, T.; et al. The anti-phospholipid antibody-dependent and independent effects of periodontopathic bacteria on threatened preterm labor and preterm birth. Arch. Gynecol. Obstet. 2013, 288, 65–72. [Google Scholar] [CrossRef]
- Figuero, E.; Han, Y.W.; Furuichi, Y. Periodontal diseases and adverse pregnancy outcomes: Mechanisms. Periodontology 2000 2020, 83, 175–188. [Google Scholar] [CrossRef]
- Hays, A.; Duan, X.; Zhu, J.; Zhou, W.; Upadhyayula, S.; Shivde, J.; Song, L.; Wang, H.; Su, L.; Zhou, X.; et al. Down-regulated Treg cells in exacerbated periodontal disease during pregnancy. Int. Immunopharmacol. 2019, 69, 299–306. [Google Scholar] [CrossRef]
- Anbinder, A.L.; Moraes, R.; Lima, G.M.; Oliveira, F.E.; Campos, D.R.; Rossoni, R.; Oliveira, L.; Junqueira, J.; Ma, Y.; Elefteriou, F. Periodontal disease exacerbates systemic ovariectomy-induced bone loss in mice. Bone 2016, 83, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Arjunan, P.; Swaminathan, R.; Yuan, J.; Elashiry, M.; Tawfik, A.; Al-Shabrawey, M.; Martin, P.; Muthusamy, T.; Cutler, C. Exacerbation of AMD Phenotype in Lasered CNV Murine Model by Dysbiotic Oral Pathogens. Antioxidants 2021, 10, 309. [Google Scholar] [CrossRef]
- Bi, C.; Han, X.; Li, X.; Sun, J.; Liu, Y.; Duan, S.; Lu, H. Periodontitis aggravates renal inflammatory response in a mouse model of renal fibrosis. Oral Dis. 2020, in press. [Google Scholar] [CrossRef]
- Beck, J.; Papapanou, P.N.; Philips, K.; Offenbacher, S. Periodontal Medicine: 100 Years of Progress. J. Dent. Res. 2019, 98, 1053–1062. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, A.; Santos-Junior, N.N.; Luiz, J.P.M.; De Oliveira, M.; Kanashiro, A.; Taira, T.M.; Fukada, S.Y.; Alves-Filho, J.C.; Junior, R.F.; Salgado, H.C. Cardiovascular and Autonomic Dysfunction in Murine Ligature-Induced Periodontitis. Sci. Rep. 2020, 10, 6891. [Google Scholar] [CrossRef]
- Zhan, Y.; Lu, R.; Meng, H.; Hou, J.; Huang, W.; Wang, X.; Hu, W. Platelets as inflammatory mediators in a murine model of periodontitis. J. Clin. Periodontol. 2020, 47, 572–582. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Huang, W.; Meng, H.; Zhan, Y.; Hou, J. Pro-inflammatory cytokine interleukin-6-induced hepcidin, a key mediator of periodontitis-related anemia of inflammation. J. Periodontal Res. 2021, 56, 690–701. [Google Scholar] [CrossRef]
- Matsuda, Y.; Kato, T.; Takahashi, N.; Nakajima, M.; Arimatsu, K.; Minagawa, T.; Sato, K.; Ohno, H.; Yamazaki, K. Ligature-induced periodontitis in mice induces elevated levels of circulating interleukin-6 but shows only weak effects on adipose and liver tissues. J. Periodontal Res. 2016, 51, 639–646. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Tong, Y.; Zhang, J.; Khan, N.; Zhang, K.; Bai, H.; Zhang, Q.; Chen, Y. Neuroinflammation changes with periodontal inflammation status during periodontitis in wild-type mice. Oral Dis. 2021, 27, 1001–1011. [Google Scholar] [CrossRef] [PubMed]
- Furutama, D.; Matsuda, S.; Yamawaki, Y.; Hatano, S.; Okanobu, A.; Memida, T.; Oue, H.; Fujita, T.; Ouhara, K.; Kajiya, M.; et al. IL-6 Induced by Periodontal Inflammation Causes Neuroinflammation and Disrupts the Blood–Brain Barrier. Brain Sci. 2020, 10, 679. [Google Scholar] [CrossRef]
- Xue, L.; Zou, X.; Yang, X.-Q.; Peng, F.; Yu, D.-K.; Du, J.-R. Chronic periodontitis induces microbiota-gut-brain axis disorders and cognitive impairment in mice. Exp. Neurol. 2020, 326, 113176. [Google Scholar] [CrossRef]
- Kitamoto, S.; Nagao-Kitamoto, H.; Jiao, Y.; Gillilland, M.G.; Hayashi, A.; Imai, J.; Sugihara, K.; Miyoshi, M.; Brazil, J.C.; Kuffa, P.; et al. The Intermucosal Connection between the Mouth and Gut in Commensal Pathobiont-Driven Colitis. Cell 2020, 182, 447–462.e14. [Google Scholar] [CrossRef]
- Silva, I.L.; Cascales, E. Molecular Strategies Underlying Porphyromonas gingivalis Virulence. J. Mol. Biol. 2021, 433, 166836. [Google Scholar] [CrossRef]
- Katz, J.; Sambandam, V.; Wu, J.H.; Michalek, S.M.; Balkovetz, D.F. Characterization of Porphyromonas gingivalis -Induced Degradation of Epithelial Cell Junctional Complexes. Infect. Immun. 2000, 68, 1441–1449. [Google Scholar] [CrossRef] [PubMed]
- Cugini, C.; Ramasubbu, N.; Tsiagbe, V.K.; Fine, D.H. Dysbiosis from a Microbial and Host Perspective Relative to Oral Health and Disease. Front. Microbiol. 2021, 12, 617485. [Google Scholar] [CrossRef]
- Jia, L.; Han, N.; Du, J.; Guo, L.; Luo, Z.; Liu, Y. Pathogenesis of Important Virulence Factors of Porphyromonas gingivalis via Toll-Like Receptors. Front. Cell. Infect. Microbiol. 2019, 9, 262. [Google Scholar] [CrossRef]
- Meyle, J.; Dommisch, H.; Groeger, S.; Giacaman, R.A.; Costalonga, M.; Herzberg, M. The innate host response in caries and periodontitis. J. Clin. Periodontol. 2017, 44, 1215–1225. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.; Hu, Y.; Wang, Y.; Kawai, T.; Wang, Z.; Han, X. Different engagement of TLR2 and TLR4 in Porphyromonas gingivalis vs. ligature-induced periodontal bone loss. Braz. Oral Res. 2017, 31, e63. [Google Scholar] [CrossRef] [PubMed]
- Beutler, B. Innate immunity: An overview. Mol. Immunol. 2004, 40, 845–859. [Google Scholar] [CrossRef]
- Strydom, N.; Rankin, S.M. Regulation of Circulating Neutrophil Numbers under Homeostasis and in Disease. J. Innate Immun. 2013, 5, 304–314. [Google Scholar] [CrossRef] [PubMed]
- Chadwick, J.W.; Fine, N.; Khoury, W.; Tasevski, N.; Sun, C.; Boroumand, P.; Klip, A.; Glogauer, M. Tissue-specific murine neutrophil activation states in health and inflammation. J. Leukoc. Biol. 2020, 110, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Nauseef, W.M.; Borregaard, N. Neutrophils at work. Nat Immun. 2014, 15, 602–611. [Google Scholar] [CrossRef]
- Kim, A.R.; Kim, J.; Choi, Y.H.; Jeon, Y.; Cha, J.; Bak, E.; Yoo, Y. The presence of neutrophils causes RANKL expression in periodontal tissue, giving rise to osteoclast formation. J. Periodontal Res. 2020, 55, 868–876. [Google Scholar] [CrossRef] [PubMed]
- Pliyev, B.K.; Kalintseva, M.V.; Abdulaeva, S.V.; Yarygin, K.N.; Savchenko, V.G. Neutrophil microparticles modulate cytokine production by natural killer cells. Cytokine 2014, 65, 126–129. [Google Scholar] [CrossRef]
- Sima, C.; Aboodi, G.M.; Lakschevitz, F.S.; Sun, C.; Goldberg, M.B.; Glogauer, M. Nuclear Factor Erythroid 2-Related Factor 2 Down-Regulation in Oral Neutrophils Is Associated with Periodontal Oxidative Damage and Severe Chronic Periodontitis. Am. J. Pathol. 2016, 186, 1417–1426. [Google Scholar] [CrossRef]
- Pathak, J.L.; Fang, Y.; Chen, Y.; Ye, Z.; Guo, X.; Yan, Y.; Zha, J.; Liang, D.; Ke, X.; Yang, L.; et al. Downregulation of Macrophage-Specific Act-1 Intensifies Periodontitis and Alveolar Bone Loss Possibly via TNF/NF-κB Signaling. Front Cell Dev Biol. 2021, 9, 628139. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Z.; Yoshizawa-Smith, S.; Glowacki, A.; Maltos, K.; Pacheco, C.; Shehabeldin, M.; Mulkeen, M.; Myers, N.; Chong, R.; Verdelis, K.; et al. Induction of M2 Macrophages Prevents Bone Loss in Murine Periodontitis Models. J. Dent. Res. 2019, 98, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, T.; Movila, A.; Kataoka, S.; Wisitrasameewong, W.; Torruella, M.R.; Murakoshi, M.; Murakami, S.; Kawai, T. Proinflammatory M1 Macrophages Inhibit RANKL-Induced Osteoclastogenesis. Infect. Immun. 2016, 84, 2802–2812. [Google Scholar] [CrossRef] [PubMed]
- McKenzie, A.N.; Spits, H.; Eberl, G. Innate Lymphoid Cells in Inflammation and Immunity. Immun. 2014, 41, 366–374. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.; Hoda, N.; Susin, C.; Wheeler, J.N.; Marshall, B.; Perry, L.; Saad, N.; Yin, L.; Elsayed, R.; Elsalanty, M.; et al. Increased Innate Lymphoid Cells in Periodontal Tissue of the Murine Model of Periodontitis: The Role of AMP-Activated Protein Kinase and Relevance for the Human Condition. Front. Immunol. 2017, 8, 922. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Tizzano, M.; Redding, K.; He, J.; Peng, X.; Jiang, P.; Xu, X.; Zhou, X.; Margolskee, R.F. Gingival solitary chemosensory cells are immune sentinels for periodontitis. Nat. Commun. 2019, 10, 4496. [Google Scholar] [CrossRef] [PubMed]
- Medzhitov, R. Toll-like receptors and innate immunity. Nat. Rev. Immunol. 2001, 1, 135–145. [Google Scholar] [CrossRef]
- Lin, J.; Bi, L.; Yu, X.; Kawai, T.; Taubman, M.A.; Shen, B.; Han, X. Porphyromonas gingivalis Exacerbates Ligature-Induced, RANKL-Dependent Alveolar Bone Resorption via Differential Regulation of Toll-Like Receptor 2 (TLR2) and TLR4. Infect. Immun. 2014, 82, 4127–4134. [Google Scholar] [CrossRef]
- Crump, K.E.; Oakley, J.C.; Xia-Juan, X.; Madu, T.C.; Devaki, S.; Mooney, E.C.; Sahingur, S.E. Interplay of Toll-Like Receptor 9, Myeloid Cells, and Deubiquitinase A20 in Periodontal Inflammation. Infect. Immun. 2017, 85, e00814-16. [Google Scholar] [CrossRef]
- Catrysse, L.; Vereecke, L.; Beyaert, R.; van Loo, G. A20 in inflammation and autoimmunity. Trends Immunol. 2014, 35, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Mooney, E.C.; Holden, S.E.; Xia, X.J.; Cohen, D.J.; Walsh, S.W.; Ma, A.; Sahingur, S.E. A20 Orchestrates Inflammatory Response in the Oral Mucosa through Restraining NF-κB Activity. J. Immunol. 2019, 202, 2044–2056. [Google Scholar] [CrossRef]
- Gao, L.; Faibish, D.; Fredman, G.; Herrera, B.; Chiang, N.; Serhan, C.N.; Van Dyke, T.E.; Gyurko, R. Resolvin E1 and Chemokine-like Receptor 1 Mediate Bone Preservation. J. Immunol. 2013, 190, 689–694. [Google Scholar] [CrossRef]
- Abe, T.; Hosur, K.B.; Hajishengallis, E.; Reis, E.S.; Ricklin, D.; Lambris, J.D.; Hajishengallis, G. Local Complement-Targeted Intervention in Periodontitis: Proof-of-Concept Using a C5a Receptor (CD88) Antagonist. J. Immunol. 2012, 189, 5442–5448. [Google Scholar] [CrossRef] [PubMed]
- Maekawa, T.; Abe, T.; Hajishengallis, E.; Hosur, K.B.; DeAngelis, R.A.; Ricklin, D.; Lambris, J.D.; Hajishengallis, G. Genetic and Intervention Studies Implicating Complement C3 as a Major Target for the Treatment of Periodontitis. J. Immunol. 2014, 192, 6020–6027. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, J.; Burrows, P.D.; Wang, J.-Y. B Cell Development and Maturation. In B Cells in Immunity and Tolerance; Advances in Experimental Medicine and Biology; Springer: Singapore, 2020; Volume 1254, pp. 1–22. [Google Scholar] [CrossRef]
- Arce-Sillas, A.; Álvarez-Luquín, D.D.; Tamaya-Domínguez, B.; Gomez-Fuentes, S.; Trejo-García, A.; Melo-Salas, M.; Cárdenas, G.; Rodríguez-Ramírez, J.; Adalid-Peralta, L. Regulatory T Cells: Molecular Actions on Effector Cells in Immune Regulation. J. Immunol. Res. 2016, 2016, 1720827. [Google Scholar] [CrossRef]
- Langrish, C.L.; Chen, Y.; Blumenschein, W.M.; Mattson, J.; Basham, B.; Sedgwick, J.D.; McClanahan, T.; Kastelein, R.A.; Cua, D.J. IL-23 drives a pathogenic T cell population that induces autoimmune inflammation. J. Exp. Med. 2005, 201, 233–240. [Google Scholar] [CrossRef]
- Tsukasaki, M.; Komatsu, N.; Nagashima, K.; Nitta, T.; Pluemsakunthai, W.; Shukunami, C.; Iwakura, Y.; Nakashima, T.; Okamoto, K.; Takayanagi, H. Host defense against oral microbiota by bone-damaging T cells. Nat. Commun. 2018, 9, 701. [Google Scholar] [CrossRef] [PubMed]
- Pacheco, C.M.F.; Maltos, K.L.M.; Shehabeldin, M.S.; Thomas, L.L.; Zhuang, Z.; Yoshizawa, S.; Verdelis, K.; Gaffen, S.L.; Garlet, G.P.; Little, S.R.; et al. Local Sustained Delivery of Anti–IL-17A Antibodies Limits Inflammatory Bone Loss in Murine Experimental Periodontitis. J. Immunol. 2021, 206, 2386–2392. [Google Scholar] [CrossRef]
- Sun, L.; Girnary, M.; Wang, L.; Jiao, Y.; Zeng, E.; Mercer, K.; Zhang, J.; Marchesan, J.T.; Yu, N.; Moss, K.; et al. IL-10 Dampens an IL-17–Mediated Periodontitis-Associated Inflammatory Network. J. Immunol. 2020, 204, 2177–2191. [Google Scholar] [CrossRef]
- Gonçalves-Zillo, T.O.; Pugliese, L.S.; Sales, V.M.T.; Mori, M.A.D.S.; Squaiella-Baptistão, C.C.; Longo-Maugeri, I.M.; Lopes, J.D.; De Oliveira, S.M.; Monteiro, A.C.; Pesquero, J.B. Increased bone loss and amount of osteoclasts in kinin B1 receptor knockout mice. J. Clin. Periodontol. 2013, 40, 653–660. [Google Scholar] [CrossRef] [PubMed]
- Abe, T.; AlSarhan, M.; Benakanakere, M.R.; Maekawa, T.; Kinane, D.F.; Cancro, M.; Korostoff, J.M.; Hajishengallis, G. The B Cell–Stimulatory Cytokines BLyS and APRIL Are Elevated in Human Periodontitis and Are Required for B Cell–Dependent Bone Loss in Experimental Murine Periodontitis. J. Immunol. 2015, 195, 1427–1435. [Google Scholar] [CrossRef]
- Yu, P.; Hu, Y.; Liu, Z.; Kawai, T.; Taubman, M.A.; Li, W.; Han, X. Local Induction of B Cell Interleukin-10 Competency Alleviates Inflammation and Bone Loss in Ligature-Induced Experimental Periodontitis in Mice. Infect. Immun. 2017, 85. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Hu, Y.; Deng, S.; Yu, P.; Chen, B.; Wang, Z.; Han, X. Cytidine-phosphate-guanosine oligodeoxynucleotides in combination with CD40 ligand decrease periodontal inflammation and alveolar bone loss in a TLR9-independent manner. J. Appl. Oral Sci. 2018, 26, e20170451. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Yu, P.; Yu, X.; Hu, X.; Kawai, T.; Han, X. IL-21/anti-Tim1/CD40 ligand promotes B10 activity in vitro and alleviates bone loss in experimental periodontitis in vivo. Biochim. Biophys. Acta 2017, 1863, 2149–2157. [Google Scholar] [CrossRef]
- Bi, J.; Dai, J.; Koivisto, L.; Larjava, M.; Bi, L.; Häkkinen, L.; Larjava, H. Inflammasome and cytokine expression profiling in experimental periodontitis in the integrin β6 null mouse. Cytokine 2019, 114, 135–142. [Google Scholar] [CrossRef]
- Hirschfeld, J.; Howait, M.; Movila, A.; Parčina, M.; Bekeredjian-Ding, I.; Deschner, J.; Jepsen, S.; Kawai, T. Assessment of the involvement of the macrophage migration inhibitory factor-glucocorticoid regulatory dyad in the expression of matrix metalloproteinase-2 during periodontitis. Eur. J. Oral Sci. 2017, 125, 345–354. [Google Scholar] [CrossRef]
- Papadakou, P.; Bletsa, A.; Yassin, M.; Karlsen, T.; Wiig, H.; Berggreen, E. Role of Hyperplasia of Gingival Lymphatics in Periodontal Inflammation. J. Dent. Res. 2017, 96, 467–476. [Google Scholar] [CrossRef]
- Zhou, W.; Su, L.; Duan, X.; Chen, X.; Hays, A.; Upadhyayula, S.; Shivde, J.; Wang, H.; Li, Y.; Huang, D.; et al. MicroRNA-21 down-regulates inflammation and inhibits periodontitis. Mol. Immunol. 2018, 101, 608–614. [Google Scholar] [CrossRef]
- Ouhara, K.; Munenaga, S.; Kajiya, M.; Takeda, K.; Matsuda, S.; Sato, Y.; Hamamoto, Y.; Iwata, T.; Yamasaki, S.; Akutagawa, K.; et al. The induced RNA-binding protein, HuR, targets 3′-UTR region of IL-6 mRNA and enhances its stabilization in periodontitis. Clin. Exp. Immunol. 2018, 192, 325–336. [Google Scholar] [CrossRef]
- Seino, Y.; Fukushima, M.; Yabe, D. GIP and GLP-1, the two incretin hormones: Similarities and differences. J. Diabetes Investig. 2010, 1, 8–23. [Google Scholar] [CrossRef] [PubMed]
- Tsukiyama, K.; Yamada, Y.; Yamada, C.; Harada, N.; Kawasaki, Y.; Ogura, M.; Bessho, K.; Li, M.; Amizuka, N.; Sato, M.; et al. Gastric Inhibitory Polypeptide as an Endogenous Factor Promoting New Bone Formation after Food Ingestion. Mol. Endocrinol. 2006, 20, 1644–1651. [Google Scholar] [CrossRef]
- Miyawaki, K.; Yamada, Y.; Ban, N.; Ihara, Y.; Tsukiyama, K.; Zhou, H.; Fujimoto, S.; Oku, A.; Tsuda, K.; Toyokuni, S.; et al. Inhibition of gastric inhibitory polypeptide signaling prevents obesity. Nat. Med. 2002, 8, 738–742. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y.; Nakamura, N.; Miyabe, M.; Nishikawa, T.; Miyajima, S.; Adachi, K.; Mizutani, M.; Kikuchi, T.; Miyazawa, K.; Goto, S.; et al. Anti-inflammatory role of glucose-dependent insulinotropic polypeptide in periodontitis. J. Diabetes Investig. 2016, 7, 497–505. [Google Scholar] [CrossRef]
- Rajshankar, D.; Sima, C.; Wang, Q.; Goldberg, S.R.; Kazembe, M.; Wang, Y.; Glogauer, M.; Downey, G.P.; McCulloch, C.A. Role of PTPα in the Destruction of Periodontal Connective Tissues. PLoS ONE 2013, 8, e70659. [Google Scholar] [CrossRef][Green Version]
- Kittaka, M.; Yoshimoto, T.; Schlosser, C.; Kajiya, M.; Kurihara, H.; Reichenberger, E.J.; Ueki, Y. Microbe-Dependent Exacerbated Alveolar Bone Destruction in Heterozygous Cherubism Mice. JBMR Plus 2020, 4, e10352. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.; Vuong, H.; Kim, S.; Lenon, A.; Ho, K.; Hsiao, E.; Sung, E.C.; Kim, R. Indigenous Microbiota Protects against Inflammation-Induced Osteonecrosis. J. Dent. Res. 2020, 99, 676–684. [Google Scholar] [CrossRef]
- Hiyoshi, T.; Domon, H.; Maekawa, T.; Yonezawa, D.; Kunitomo, E.; Tabeta, K.; Terao, Y. Protective effect of hinokitiol against periodontal bone loss in ligature-induced experimental periodontitis in mice. Arch. Oral Biol. 2020, 112, 104679. [Google Scholar] [CrossRef]
- Kim, J.; Jung, B.H.; Lee, J.H.; Yoo, K.; Lee, H.; Kang, M.; Lee, J. Effect of Weissella cibaria on the reduction of periodontal tissue destruction in mice. J. Periodontol. 2020, 91, 1367–1374. [Google Scholar] [CrossRef]
- Huang, Y.; Liao, Y.; Luo, B.; Li, L.; Zhang, Y.; Yan, F. Non-surgical Periodontal Treatment Restored the Gut Microbiota and Intestinal Barrier in Apolipoprotein E−/− Mice with Periodontitis. Front. Cell. Infect. Microbiol. 2020, 10, 498. [Google Scholar] [CrossRef]
- Li, L.; Bao, J.; Chang, Y.; Wang, M.; Chen, B.; Yan, F. Gut Microbiota May Mediate the Influence of Periodontitis on Prediabetes. J. Dent. Res. 2021. [Google Scholar] [CrossRef] [PubMed]
- An, T.; Chen, Y.; Li, M.; Liu, Y.; Zhang, Z.; Yang, Q. Inhibition of experimental periodontitis by a monoclonal antibody against Porphyromonas gingivalis HA2. Microb. Pathog. 2021, 154, 104633. [Google Scholar] [CrossRef] [PubMed]
- Bhattarai, G.; Poudel, S.B.; Kook, S.-H.; Lee, J.-C. Anti-inflammatory, anti-osteoclastic, and antioxidant activities of genistein protect against alveolar bone loss and periodontal tissue degradation in a mouse model of periodontitis. J. Biomed. Mater. Res. Part A 2017, 105, 2510–2521. [Google Scholar] [CrossRef]
- Cao, G.; Zhang, X.; Song, Y.; Sun, Y.; Ling, H.; Han, X.; Lin, J. Local promotion of B10 function alleviates experimental periodontitis bone loss through antagonizing RANKL-expressing neutrophils. J. Periodontol. 2021, 92, 907–920. [Google Scholar] [CrossRef]
- Clark, D.; Halpern, B.; Miclau, T.; Nakamura, M.; Kapila, Y.; Marcucio, R. The Contribution of Macrophages in Old Mice to Periodontal Disease. J. Dent. Res. 2021. [Google Scholar] [CrossRef]
- Francis, N.; Ayodele, B.; O’Brien-Simpson, N.M.; Birchmeier, W.; Pike, R.N.; Pagel, C.N.; Mackie, E.J. Keratinocyte-specific ablation of protease-activated receptor 2 prevents gingival inflammation and bone loss in a mouse model of periodontal disease. Cell. Microbiol. 2018, 20, e12891. [Google Scholar] [CrossRef]
- Ideguchi, H.; Yamashiro, K.; Yamamoto, T.; Shimoe, M.; Hongo, S.; Kochi, S.; Yoshihara-Hirata, C.; Aoyagi, H.; Kawamura, M.; Takashiba, S. Molecular imaging assessment of periodontitis lesions in an experimental mouse model. Clin. Oral Investig. 2019, 23, 821–827. [Google Scholar] [CrossRef]
- Lapérine, O.; Cloitre, A.; Caillon, J.; Huck, O.; Bugueno, I.M.; Pilet, P.; Sourice, S.; Le Tilly, E.; Palmer, G.; Davideau, J.-L.; et al. Interleukin-33 and RANK-L Interplay in the Alveolar Bone Loss Associated to Periodontitis. PLoS ONE 2016, 11, e0168080. [Google Scholar] [CrossRef]
- Nagashima, H.; Shinoda, M.; Honda, K.; Kamio, N.; Hasuike, A.; Sugano, N.; Arai, Y.; Sato, S.; Iwata, K. CXCR4 signaling contributes to alveolar bone resorption in Porphyromonas gingivalis—Induced periodontitis in mice. J. Oral Sci. 2017, 59, 571–577. [Google Scholar] [CrossRef]
- Wang, J.; Wang, B.; Lv, X.; Wang, L. NIK inhibitor impairs chronic periodontitis via suppressing non-canonical NF-κB and osteoclastogenesis. Pathog. Dis. 2020, 78, ftaa045. [Google Scholar] [CrossRef]
- Wang, J.; Wang, B.; Lv, X.; Wang, Y. Halofuginone functions as a therapeutic drug for chronic periodontitis in a mouse model. Int. J. Immunopathol. Pharmacol. 2020, 34, 1–9. [Google Scholar] [CrossRef]
- Wang, Y.; Hu, Y.; Pan, K.; Li, H.; Shang, S.; Wang, Y.; Tang, G.; Han, X. In-vivo imaging revealed antigen-directed gingival B10 infiltration in experimental periodontitis. Biochim. Biophys. Acta BBA Mol. Basis Dis. 2021, 1867, 165991. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yu, X.; Lin, J.; Hu, Y.; Zhao, Q.; Kawai, T.; Taubman, M.A.; Han, X. B10 Cells Alleviate Periodontal Bone Loss in Experimental Periodontitis. Infect. Immun. 2017, 85, e00335-17. [Google Scholar] [CrossRef]
- Korah, L.; Amri, N.; Bugueno, I.M.; Hotton, D.; Tenenbaum, H.; Huck, O.; Berdal, A.; Davideau, J.; Valdebenito, I.M.B. Experimental periodontitis in Msx2 mutant mice induces alveolar bone necrosis. J. Periodontol. 2020, 91, 693–704. [Google Scholar] [CrossRef] [PubMed]
- Li, C.H.; Amar, S. Morphometric, Histomorphometric, and Microcomputed Tomographic Analysis of Periodontal Inflammatory Lesions in a Murine Model. J. Periodontol. 2007, 78, 1120–1128. [Google Scholar] [CrossRef]
- Palioto, D.B.; Finoti, L.S.; Kinane, D.F.; Benakanakere, M. Epigenetic and inflammatory events in experimental periodontitis following systemic microbial challenge. J. Clin. Periodontol. 2019, 46, 819–829. [Google Scholar] [CrossRef]
- Wang, B.; Bai, S.; Wang, J.; Ren, N.; Xie, R.; Cheng, G.; Yu, Y. TPCA-1 negatively regulates inflammation mediated by NF-κB pathway in mouse chronic periodontitis model. Mol. Oral Microbiol. 2021, 36, 192–201. [Google Scholar] [CrossRef]
- Yang, X.; Han, X.; Shu, R.; Jiang, F.; Xu, L.; Xue, C.; Chen, T.; Bai, D. Effect of sclerostin removal in vivo on experimental periodontitis in mice. J. Oral Sci. 2016, 58, 271–276. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Sulijaya, B.; Yamada-Hara, M.; Yokoji-Takeuchi, M.; Matsuda-Matsukawa, Y.; Yamazaki, K.; Matsugishi, A.; Tsuzuno, T.; Sato, K.; Aoki-Nonaka, Y.; Takahashi, N.; et al. Antimicrobial function of the polyunsaturated fatty acid KetoC in an experimental model of periodontitis. J. Periodontol. 2019, 90, 1470–1480. [Google Scholar] [CrossRef] [PubMed]
- Duan, X.; Hays, A.; Zhou, W.; Sileewa, N.; Upadhyayula, S.; Wang, H.; Liang, S. Porphyromonas gingivalis induces exacerbated periodontal disease during pregnancy. Microb. Pathog. 2018, 124, 145–151. [Google Scholar] [CrossRef]
- Kimura, S.; Nagai, A.; Onitsuka, T.; Koga, T.; Fujiwara, T.; Kaya, H.; Hamada, S. Induction of Experimental Periodontitis in Mice with Porphyromonas gingivalis—Adhered Ligatures. J. Periodontol. 2000, 71, 1167–1173. [Google Scholar] [CrossRef]
- Bugueno, I.M.; Batool, F.; Korah, L.; Benkirane-Jessel, N.; Huck, O. Porphyromonas gingivalis Differentially Modulates Apoptosome Apoptotic Peptidase Activating Factor 1 in Epithelial Cells and Fibroblasts. Am. J. Pathol. 2018, 188, 404–416. [Google Scholar] [CrossRef] [PubMed]
- Baker, P.J.; Evans, R.; Roopenian, D.C. Oral infection with Porphyromonas gingivalis and induced alveolar bone loss in immunocompetent and severe combined immunodeficient mice. Arch. Oral Biol. 1994, 39, 1035–1040. [Google Scholar] [CrossRef]
- Saadi-Thiers, K.; Huck, O.; Simonis, P.; Tilly, P.; Fabre, J.-E.; Tenenbaum, H.; Davideau, J.-L. Periodontal and Systemic Responses in Various Mice Models of Experimental Periodontitis: Respective Roles of Inflammation Duration and Porphyromonas gingivalis Infection. J. Periodontol. 2013, 84, 396–406. [Google Scholar] [CrossRef]
- Barros, S.P.; Arce, R.M.; Galloway, P.; Lawter, R.; Offenbacher, S. Therapeutic effect of a topical CCR2 antagonist on induced alveolar bone loss in mice. J. Periodontal Res. 2011, 46, 246–251. [Google Scholar] [CrossRef]
- de Molon, R.S.; Mascarenhas, V.I.; de Avila, E.; Finoti, L.S.; Toffoli, G.B.; Spolidorio, D.M.P.; Scarel-Caminaga, R.; Tetradis, S.; Cirelli, J.A. Long-term evaluation of oral gavage with periodontopathogens or ligature induction of experimental periodontal disease in mice. Clin. Oral Investig. 2016, 20, 1203–1216. [Google Scholar] [CrossRef]
- Kubota, M.; Yanagita, M.; Mori, K.; Hasegawa, S.; Yamashita, M.; Yamada, S.; Kitamura, M.; Murakami, S. The Effects of Cigarette Smoke Condensate and Nicotine on Periodontal Tissue in a Periodontitis Model Mouse. PLoS ONE 2016, 11, e0155594. [Google Scholar] [CrossRef]
- Slots, J. Systemic antibiotics in periodontics. J. Periodontol. 2004, 75, 1553–1565. [Google Scholar] [CrossRef]
- Feres, M.; Figueiredo, L.C.; Soares, G.M.S.; Faveri, M. Systemic antibiotics in the treatment of periodontitis. Periodontology 2000 2015, 67, 131–186. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Liu, Z.; Kawai, T.; Zhou, Y.; Han, X. Antibiotic administration alleviates the aggravating effect of orthodontic force on ligature-induced experimental periodontitis bone loss in mice. J. Periodontal Res. 2017, 52, 725–733. [Google Scholar] [CrossRef] [PubMed]
- Maekawa, T.; Tamura, H.; Domon, H.; Hiyoshi, T.; Isono, T.; Yonezawa, D.; Hayashi, N.; Takahashi, N.; Tabeta, K.; Maeda, T.; et al. Erythromycin inhibits neutrophilic inflammation and mucosal disease by upregulating DEL-1. JCI Insight 2020, 5, e136706. [Google Scholar] [CrossRef]
- Tamura, H.; Maekawa, T.; Domon, H.; Hiyoshi, T.; Hirayama, S.; Isono, T.; Sasagawa, K.; Yonezawa, D.; Takahashi, N.; Oda, M.; et al. Effects of Erythromycin on Osteoclasts and Bone Resorption via DEL-1 Induction in Mice. Antibiotics 2021, 10, 312. [Google Scholar] [CrossRef]
- Zimmermann, P.; Ziesenitz, V.; Curtis, N.; Ritz, N. The Immunomodulatory Effects of Macrolides—A Systematic Review of the Underlying Mechanisms. Front. Immunol. 2018, 9, 302. [Google Scholar] [CrossRef]
- Choi, E.Y.; Chavakis, E.; Czabanka, M.A.; Langer, H.F.; Fraemohs, L.; Economopoulou, M.; Kundu, R.K.; Orlandi, A.; Zheng, Y.Y.; Prieto, D.A.; et al. Del-1, an Endogenous Leukocyte-Endothelial Adhesion Inhibitor, Limits Inflammatory Cell Recruitment. Science 2008, 322, 1101–1104. [Google Scholar] [CrossRef]
- Kourtzelis, I.; Li, X.; Mitroulis, I.; Grosser, D.; Kajikawa, T.; Wang, B.; Grzybek, M.; Von Renesse, J.; Czogalla, A.; Troullinaki, M.; et al. DEL-1 promotes macrophage efferocytosis and clearance of inflammation. Nat. Immunol. 2019, 20, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Van Dyke, T.E.; Gyurko, R. Resolvin E1 regulates osteoclast fusion via DC-STAMP and NFATc1. FASEB J. 2013, 27, 3344–3353. [Google Scholar] [CrossRef]
- Belibasakis, G.N.; Emingil, G.; Saygan, B.; Turkoglu, O.; Atilla, G.; Bostanci, N. Gene expression of transcription factor NFATc1 in periodontal diseases. APMIS 2011, 119, 167–172. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Wisitrasameewong, W.; Kajiya, M.; Movila, A.; Rittling, S.; Ishii, T.; Suzuki, M.; Matsuda, S.; Mazda, Y.; Torruella, M.; Azuma, M.; et al. DC-STAMP Is an Osteoclast Fusogen Engaged in Periodontal Bone Resorption. J. Dent. Res. 2017, 96, 685–693. [Google Scholar] [CrossRef] [PubMed]
- Kumawat, K.; Gosens, R. WNT-5A: Signaling and functions in health and disease. Cell. Mol. Life Sci. 2016, 73, 567–587. [Google Scholar] [CrossRef] [PubMed]
- Nanbara, H.; Wara-Aswapati, N.; Nagasawa, T.; Yoshida, Y.; Yashiro, R.; Bando, Y.; Kobayashi, H.; Khongcharoensuk, J.; Hormdee, D.; Pitiphat, W.; et al. Modulation of Wnt5a Expression by Periodontopathic Bacteria. PLoS ONE 2012, 7, e34434. [Google Scholar] [CrossRef]
- Maekawa, T.; Kulwattanaporn, P.; Hosur, K.; Domon, H.; Oda, M.; Terao, Y.; Maeda, T.; Hajishengallis, G. Differential Expression and Roles of Secreted Frizzled-Related Protein 5 and the Wingless Homolog Wnt5a in Periodontitis. J. Dent. Res. 2017, 96, 571–577. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, R.; Meisel, H. Food-derived peptides with biological activity: From research to food applications. Curr. Opin. Biotechnol. 2007, 18, 163–169. [Google Scholar] [CrossRef]
- Lin, M.-C.; Lin, S.-B.; Lee, S.-C.; Lin, C.-C.; Hui, C.-F.; Chen, J.-Y. Antimicrobial peptide of an anti-lipopolysaccharide factor modulates of the inflammatory response in RAW264.7 cells. Peptides 2010, 31, 1262–1272. [Google Scholar] [CrossRef]
- Tamura, H.; Maekawa, T.; Domon, H.; Hiyoshi, T.; Yonezawa, D.; Nagai, K.; Ochiai, A.; Taniguchi, M.; Tabeta, K.; Maeda, T.; et al. Peptides from rice endosperm protein restrain periodontal bone loss in mouse model of periodontitis. Arch. Oral Biol. 2019, 98, 132–139. [Google Scholar] [CrossRef]
- Aoki-Nonaka, Y.; Tabeta, K.; Yokoji, M.; Matsugishi, A.; Matsuda, Y.; Takahashi, N.; Sulijaya, B.; Domon, H.; Terao, Y.; Taniguchi, M.; et al. A peptide derived from rice inhibits alveolar bone resorption via suppression of inflammatory cytokine production. J. Periodontol. 2019, 90, 1160–1169. [Google Scholar] [CrossRef]
- Mizuno, M.; Miyazawa, K.; Tabuchi, M.; Tanaka, M.; Yoshizako, M.; Minamoto, C.; Torii, Y.; Tamaoka, Y.; Kawatani, M.; Osada, H.; et al. Reveromycin A Administration Prevents Alveolar Bone Loss in Osteoprotegerin Knockout Mice with Periodontal Disease. Sci. Rep. 2015, 5, 16510. [Google Scholar] [CrossRef] [PubMed]
- Woo, J.-T.; Kawatani, M.; Kato, M.; Shinki, T.; Yonezawa, T.; Kanoh, N.; Nakagawa, H.; Takami, M.; Lee, K.H.; Stern, P.H.; et al. Reveromycin A, an agent for osteoporosis, inhibits bone resorption by inducing apoptosis specifically in osteoclasts. Proc. Natl. Acad. Sci. USA 2006, 103, 4729–4734. [Google Scholar] [CrossRef]
- Ong, K.C.; Khoo, H.E. Biological effects of myricetin. Gen. Pharmacol. Vasc. System 1997, 29, 121–126. [Google Scholar] [CrossRef]
- Kuo, P.L. Myricetin inhibits the induction of anti-Fas IgM-, tumor necrosis factor-alpha- and interleukin-1beta-mediated apoptosis by Fas pathway inhibition in human osteoblastic cell line MG-63. Life Sci. 2005, 77, 2964–2976. [Google Scholar] [CrossRef] [PubMed]
- Hsu, Y.-L.; Chang, J.-K.; Tsai, C.-H.; Chien, T.-T.C.; Kuo, P.-L. Myricetin induces human osteoblast differentiation through bone morphogenetic protein-2/p38 mitogen-activated protein kinase pathway. Biochem. Pharmacol. 2007, 73, 504–514. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, M.; Hamamoto, R.; Uchiyama, S.; Ishiyama, K. Effects of flavonoid on calcium content in femoral tissue culture and parathyroid hormone-stimulated osteoclastogenesis in bone marrow culture in vitro. Mol. Cell. Biochem. 2007, 303, 83–88. [Google Scholar] [CrossRef]
- Wu, C.; Wang, W.; Tian, B.; Liu, X.; Qu, X.; Zhai, Z.; Li, H.; Liu, F.; Fan, Q.; Tang, T.; et al. Myricetin prevents titanium particle-induced osteolysis in vivo and inhibits RANKL-induced osteoclastogenesis in vitro. Biochem. Pharmacol. 2015, 93, 59–71. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Wu, C.; Tian, B.; Zhou, X.; Ma, N.; Qian, Y. Myricetin Prevents Alveolar Bone Loss in an Experimental Ovariectomized Mouse Model of Periodontitis. Int. J. Mol. Sci. 2016, 17, 422. [Google Scholar] [CrossRef]
- Kalogeropoulos, N.; Chiou, A.; Ioannou, M.; Karathanos, V.T.; Hassapidou, M.; Andrikopoulos, N.K. Nutritional evaluation and bioactive microconstituents (phytosterols, tocopherols, polyphenols, triterpenic acids) in cooked dry legumes usually consumed in the Mediterranean countries. Food Chem. 2010, 121, 682–690. [Google Scholar] [CrossRef]
- Lee, W.; Yang, E.-J.; Ku, S.-K.; Song, K.-S.; Bae, J.-S. Anti-inflammatory Effects of Oleanolic Acid on LPS-Induced Inflammation In Vitro and In Vivo. Inflammation 2013, 36, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Žiberna, L.; Šamec, D.; Mocan, A.; Nabavi, S.; Bishayee, A.; Farooqi, A.A.; Sureda, A. Oleanolic Acid Alters Multiple Cell Signaling Pathways: Implication in Cancer Prevention and Therapy. Int. J. Mol. Sci. 2017, 18, 643. [Google Scholar] [CrossRef]
- Adhikari, N.; Neupane, S.; Aryal, Y.P.; Choi, M.; Sohn, W.; Lee, Y.; Jung, J.; Ha, J.; Choi, S.; Suh, J.; et al. Effects of oleanolic acid acetate on bone formation in an experimental periodontitis model in mice. J. Periodontal Res. 2019, 54, 533–545. [Google Scholar] [CrossRef]
- Baliga, M.S.; Haniadka, R.; Pereira, M.M.; D’Souza, J.J.; Pallaty, P.L.; Bhat, H.P.; Popuri, S. Update on the Chemopreventive Effects of Ginger and its Phytochemicals. Crit. Rev. Food Sci. Nutr. 2011, 51, 499–523. [Google Scholar] [CrossRef]
- Kou, X.; Wang, X.; Ji, R.; Liu, L.; Qiao, Y.; Lou, Z.; Ma, C.; Li, S.; Wang, H.; Ho, C.-T. Occurrence, biological activity and metabolism of 6-shogaol. Food Funct. 2018, 9, 1310–1327. [Google Scholar] [CrossRef]
- Kim, Y.G.; Kim, M.O.; Kim, S.H.; Kim, H.J.; Pokhrel, N.K.; Lee, J.H.; Lee, H.J.; Kim, J.Y.; Lee, Y. 6-Shogaol, an active ingredient of ginger, inhibits osteoclastogenesis and alveolar bone resorption in ligature-induced periodontitis in mice. J. Periodontol. 2020, 91, 809–818. [Google Scholar] [CrossRef] [PubMed]
- Ihn, H.J.; Lee, T.; Kim, J.A.; Lee, D.; Kim, N.D.; Shin, H.-I.; Bae, Y.C.; Park, E.K. OCLI-023, a Novel Pyrimidine Compound, Suppresses Osteoclastogenesis In Vitro and Alveolar Bone Resorption In Vivo. PLoS ONE 2017, 12, e0170159. [Google Scholar] [CrossRef]
- Ihn, H.J.; Lee, T.; Lee, D.; Kim, J.A.; Kim, K.; Lim, S.; Kim, J.-Y.; Lee, Y.; Kim, S.-H.; Lee, H.-S.; et al. A novel benzamide derivative protects ligature-induced alveolar bone erosion by inhibiting NFATc1-mediated osteoclastogenesis. Toxicol. Appl. Pharmacol. 2018, 355, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Yao, W.; Guan, M.; Jia, J.; Dai, W.; Lay, Y.-A.E.; Amugongo, S.; Liu, R.; Olivos, D.; Saunders, M.; Lam, K.S.; et al. Reversing bone loss by directing mesenchymal stem cells to bone. Stem Cells 2013, 31, 2003–2014. [Google Scholar] [CrossRef] [PubMed]
- Guan, M.; Yao, W.; Liu, R.; Lam, K.S.; Nolta, J.; Jia, J.; Panganiban, B.; Meng, L.; Zhou, P.; Shahnazari, M.; et al. Directing mesenchymal stem cells to bone to augment bone formation and increase bone mass. Nat. Med. 2012, 18, 456–462. [Google Scholar] [CrossRef]
- Jiang, M.; Liu, L.; Liu, R.; Lam, K.S.; Lane, N.E.; Yao, W. A new anabolic compound, LLP2A-Ale, reserves periodontal bone loss in mice through augmentation of bone formation. BMC Pharmacol. Toxicol. 2020, 21, 76. [Google Scholar] [CrossRef]
- Cafferata, E.A.; Terraza-Aguirre, C.; Barrera, R.; Faúndez, N.; González, N.; Rojas, C.; Melgar-Rodríguez, S.; Hernández, M.; Carvajal, P.; Cortez, C.; et al. Interleukin-35 inhibits alveolar bone resorption by modulating the Th17/Treg imbalance during periodontitis. J. Clin. Periodontol. 2020, 47, 676–688. [Google Scholar] [CrossRef]
- Zhen, L.; Fan, D.-S.; Zhang, Y.; Cao, X.-M.; Wang, L.-M. Resveratrol ameliorates experimental periodontitis in diabetic mice through negative regulation of TLR4 signaling. Acta Pharmacol. Sin. 2015, 36, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, E.; Ikeda, Y.; Wang, Y.; Fine, N.; Sheikh, Z.; Viniegra, A.; Barzilay, O.; Ganss, B.; Tenenbaum, H.C.; Glogauer, M. Resveratrol derivative-rich melinjo seed extract induces healing in a murine model of established periodontitis. J. Periodontol. 2018, 89, 586–595. [Google Scholar] [CrossRef]
- Kaboosaya, B.; Wulansari, L.K.; Nguyen, V.T.; Kasugai, S. Drinking green tea alleviates alveolar bone resorption in ligature-induced periodontitis in mice. J. Oral Biosci. 2020, 62, 162–168. [Google Scholar] [CrossRef]
- Cui, D.; Li, H.; Lei, L.; Chen, C.; Yan, F. Nonsurgical periodontal treatment reduced aortic inflammation in ApoE−/− mice with periodontitis. SpringerPlus 2016, 5, 1–13. [Google Scholar] [CrossRef]
- Ying, S.; Tan, M.; Feng, G.; Kuang, Y.; Chen, D.; Li, J.; Song, J. Low-intensity Pulsed Ultrasound regulates alveolar bone homeostasis in experimental Periodontitis by diminishing Oxidative Stress. Theranostics 2020, 10, 9789–9807. [Google Scholar] [CrossRef]
- Snipes, M.; Sun, C.; Yu, H. Inhibition of sphingosine-1-phosphate receptor 2 attenuated ligature-induced periodontitis in mice. Oral Dis. 2021, 27, 1283–1291. [Google Scholar] [CrossRef]
- Kure, K.; Sato, H.; Suzuki, J.-I.; Itai, A.; Aoyama, N.; Izumi, Y. A novel IkB kinase inhibitor attenuates ligature-induced periodontal disease in mice. J. Periodontal Res. 2019, 54, 164–173. [Google Scholar] [CrossRef]
- Ihn, H.J.; Kim, Y.S.; Lim, S.; Bae, J.S.; Jung, J.C.; Kim, Y.H.; Park, J.W.; Wang, Z.; Koh, J.T.; Bae, Y.C.; et al. PF-3845, a Fatty Acid Amide Hydrolase Inhibitor, Directly Suppresses Osteoclastogenesis through ERK and NF-κB Pathways In Vitro and Alveolar Bone Loss In Vivo. Int. J. Mol. Sci. 2021, 22, 1915. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, U.; Kajioka, S.; Finoti, L.; Palioto, D.; Kinane, D.; Benakanakere, M. Decitabine Inhibits Bone Resorption in Periodontitis by Upregulating Anti-Inflammatory Cytokines and Suppressing Osteoclastogenesis. Biomedicines 2021, 9, 199. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, S.H.P.; Brito, V.G.B.; Frasnelli, S.C.T.; Ribeiro, B.D.S.; Ferreira, M.N.; Queiroz, D.P.; Beltan, C.T.; Lara, V.S.; dos Santos, C.F. Aliskiren Attenuates the Inflammatory Response and Wound Healing Process in Diabetic Mice with Periodontal Disease. Front. Pharmacol. 2019, 10, 708. [Google Scholar] [CrossRef]
- Iwamatsu-Kobayashi, Y.; Abe, S.; Fujieda, Y.; Orimoto, A.; Kanehira, M.; Handa, K.; Venkataiah, V.S.; Zou, W.; Ishikawa, M.; Saito, M. Metal ions from S-PRG filler have the potential to prevent periodontal disease. Clin. Exp. Dent. Res. 2017, 3, 126–133. [Google Scholar] [CrossRef]
- Yu, M.-C.; Chang, C.-Y.; Chao, Y.-C.; Jheng, Y.-H.; Yang, C.; Lee, N.; Yu, S.-H.; Yu, X.-H.; Liu, D.-M.; Chang, P.-C.; et al. pH-Responsive Hydrogel with an Anti-Glycation Agent for Modulating Experimental Periodontitis. J. Periodontol. 2016, 87, 742–748. [Google Scholar] [CrossRef] [PubMed]
- Iguchi, S.; Suzuki, D.; Kawano, E.; Mashimo, T.; Kajiya, M.; Toriumi, T.; Kawai, T.; Kurihara, H.; Isokawa, K.; Sato, S.; et al. Effect of local bone marrow stromal cell administration on ligature-induced periodontitis in mice. J. Oral Sci. 2017, 59, 629–637. [Google Scholar] [CrossRef]
- Nakao, Y.; Fukuda, T.; Zhang, Q.; Sanui, T.; Shinjo, T.; Kou, X.; Chen, C.; Liu, D.; Watanabe, Y.; Hayashi, C.; et al. Exosomes from TNF-α-treated human gingiva-derived MSCs enhance M2 macrophage polarization and inhibit periodontal bone loss. Acta Biomater. 2021, 122, 306–324. [Google Scholar] [CrossRef]
- Nagai, K.; Ideguchi, H.; Kajikawa, T.; Li, X.; Chavakis, T.; Cheng, J.; Messersmith, P.B.; Heber-Katz, E.; Hajishengallis, G. An injectable hydrogel-formulated inhibitor of prolyl-4-hydroxylase promotes T regulatory cell recruitment and enhances alveolar bone regeneration during resolution of experimental periodontitis. FASEB J. 2020, 34, 13726–13740. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Reference No. | Year Author | Periodontal Tissue | Main Findings |
|---|---|---|---|
| [27] | 2019 Chen et al. | Gingiva | Grhl2 KO impaired the expression of the junction proteins at the junctional epithelium and increased the alveolar bone loss in the ligature-induced periodontitis model. |
| [31] | 2021 Guo et al. | Gingiva | CTHRC1 was highly expressed in GCF and gingival tissues of periodontitis patients and animal models. |
| [32] | 2018 Movila et al. | Gingiva | Induced inflammation was observed in the gingiva, using a novel intravital endoscopic technology. |
| [35] | 2020 Choi et al. | Gingiva | GRP-positive cells were mostly located at the oral epithelium of samples from experimental periodontitis model |
| [38] | 2017 Okanobu et al. | Gingiva | A significant increase in the degree of gingival overgrowth and expansion of the connective tissue area was observed in cyclosporine A (CsA) and ligature-induced periodontal model mice, whereas cessation of CsA and antibiotic administration reduced gingival overgrowth. |
| [39] | 2019 Bao et al. | Gingiva | The gingival tissue protein abundance was shown to be mainly dependent on the progression of periodontitis by unsupervised clustering analysis. Additionally, over-representation of innate immune regulation, signal transduction, and homeostasis processes was revealed by gene ontology enrichment analysis. |
| [40] | 2019 Huang et al. | Gingiva | Ferritin was detected in the gingival epithelium and gingival connective tissue by immunochemical staining; the intensity of positive staining became significantly stronger along with the extent of inflammatory infiltration. |
| [43] | 2016 Kataoka et al. | Gingiva | The ligature model was used to visualize the oxidative stress induced by experimental periodontitis. The Nrf2/antioxidant defense pathway was activated and could be visualized in Keap1-dependent oxidative stress detector-luciferase mouse model. |
| [46] | 2018 Ishii et al. | Bone | Systemic administration of anti-OC-STAMP-mAb suppressed the expression of CD9 mRNA, but not DC-STAMP mRNA, in periodontal tissue. OC-STAMP partnered CD9 to promote periodontal destruction by upregulation of fusion during osteoclastogenesis. |
| [48] | 2018 Yoshihara-Hirata et al. | Bone | Administration of an anti-HMGB1 neutralizing antibody in an experimental periodontitis model attenuated alveolar bone resorption and inflammatory cytokines. |
| [50] | 2019 Fu et al. | Bone | A significant decrease in PTEN was observed in ligature-induced periodontitis model. PTEN could directly inhibit inflammation and bone loss via inhibiting the IL-1 and TNF-alpha pathway in periodontitis. |
| [51] | 2021 Liu et al. | Bone | Knockout of NRAGE induced autophagy-related gene expression and accelerated bone destruction by increasing the activity and differentiation of osteoclasts. |
| [53] | 2019 Bostanci et al. | Bone | TREM-1 regulates the IL-17A-RANKL/OPG axis and bone loss in experimental periodontitis. |
| [55] | 2020 Wang et al. | Bone | SLIT2 expression was upregulated in periodontitis in both humans and mice, and a higher expression of SLIT2 accelerated the progression of periodontitis. |
| [57] | 2020 Kittaka et al. | Bone | Ligature-induced alveolar bone loss in Sh3bp2-deficient mice was reduced compared to WT mice. SH3BP2/SYK signaling axis regulates inflammatory response, osteoclastogenesis, and osteoclast functions in periodontitis. |
| [59] | 2019 Huang et al. | Bone | In vitro experiments revealed that FICZ pre-treatment reduced LPS-induced inflammation in periodontal ligament cells via increased phosphorylation of STAT3. FICZ promoted the mineralization of PDLCs via activation of the Wnt/β-catenin signaling pathway. |
| [61] | 2018 Kim et al. | Bone | P2rx5-deficient mice had decreased ligature-induced bone loss compared to WT littermates. Gene expression analysis of gingival tissue of ligated mice showed that Il1b, Il6, Il17a, and Tnfsf11 expression levels were significantly reduced in P2rx5-deficient mice. |
| [62] | 2021 Chen et al. | Bone | NLRP3 deficiency reduced the number of osteoclast precursors and bone loss in ligature-induced periodontitis. |
| [64] | 2016 Takahashi et al. | Bone | Genetic and chemical deletion of TRPV1 exacerbated ligature-induced bone loss. Neuronal TRPV1 signaling in periodontal tissue is crucial for the regulation of osteoclastogenesis via the neuropeptide CGRP. |
| [65] | 2017 Zhang et al. | Bone | The expression of miR-335-5p decreased significantly in the periodontal tissue of EP. Overexpression of miR-335-5p in vivo weakened the periodontal bone destruction and inflammation compared to WT-EP group. |
| [69] | 2019 Goes et al. | Bone | Osteocyte specific Dkk1-deficient mice significantly prevented ligature-induced bone loss and mitigated inflammation. Dkk-1 derived from osteocytes played a crucial role in alveolar bone loss in periodontitis. |
| [71] | 2016 Duan et al. | Bone | Female mice displayed significantly increased periodontal bone loss, accompanied by elevated expression of proinflammatory cytokines and higher numbers of oral bacteria. |
| [72] | 2020 Aung et al. | Bone | Aged mice showed severe bone loss associated with increased osteoclast activity compared to young mice. Aging-induced impairment of MSC function is potentially correlated with progressive periodontal tissue deterioration. |
| [73] | 2018 Wong et al. | Bone | Micro-CT analysis revealed ligature-induced bone loss in peri-implant sites, such as periodontitis. The ligature was removed after one week, and the periodontitis group experienced significant bone gain, whereas the peri-implantitis group did not. |
| [75] | 2020 Rosset et al. | PDL | Inhibition of transglutaminase activity increased total collagen and thick collagen fiber content in the group with ligation at five days. |
| [80] | 2020 Guo et al. | PDL | Autophagy was significantly increased, and H19 expression was also significantly upregulated during inflammation in PDLCs of ligated mice. |
| Reference No. | Year Author | Disease Model | Main Findings |
|---|---|---|---|
| [86] | 2017 Maekawa et al. | Streptozotocin-induced diabetic model | Significantly severe alveolar bone loss was observed in STZ mice compared to WT mice at 7 days post ligation. An increased number of TRAP-positive multinucleated cells were observed at the ligated sites in STZ mice. |
| [87] | 2019 Yu et al. | High-fat diet-induced obesity model | ABL and periodontal osteoclast numbers were not affected by diet regardless of ligation or sham-ligation. Leukocyte and macrophage numbers and protein level of TNF-α in the periodontium and serum IL-6 level were downregulated in periodontitis mice fed a high-fat diet. |
| [88] | 2016 Huang et al. | High-fat diet-induced obesity model | ABL was significantly increased with periodontitis and obesity. F4/80 and MCP1 expression was significantly upregulated in gingival tissues with periodontitis but significantly downregulated in the context of obesity. |
| [89] | 2017 Yu et al. | High-fat diet-induced obesity model | In diet-induced obesity mice, periodontitis increased TG levels. After adjusting confounding effects of postoperative weight loss, periodontitis increased levels of not only TG, but also fasting insulin, HOMA-IR, and HDL in the diet-induced obesity mice. |
| [90] | 2006 Gyurko et al. | Akita mice | Ligature-induced periodontal bone loss was significantly greater in Akita mice compared with WT. Chronic hyperglycemia predisposed to an exaggerated inflammatory response and primed leukocytes for marginalization and superoxide production. |
| [91] | 2016 Liu et al. | db/db mice | Pancreatic β-cell failure, with insulin resistance, was observed in db/db mice, while periodontitis could aggravate β-cell dysfunction-related features. |
| [92] | 2021 Chen et al. | High-fat diet-induced obesity model | Compared with healthy conditions, periodontitis and a high-fat diet had distinct effects on the gingival metabolome. The metabolomic impact of periodontitis was generally greater in high-fat diet mice than in lean controls. |
| [93] | 2021 Sato et al. | High-fat diet-induced obesity model | Gut dysbiosis-associated metabolites from high-fat diet-fed mice worsen alveolar bone destruction. Obesity increases the risk of periodontal disease by increasing the production of uric acid mediated by gut dysbiosis. |
| [94] | 2020 Kantarci et al. | 5xFAD mice | Periodontitis increased neuroinflammation in WT mice and disrupted the neuroinflammatory response in 5xFAD mice, suggesting that microglia are central to the association between periodontal disease and Alzheimer’s disease. |
| [95] | 2021 Qian et al. | Amyloid-β protein precursor (AβPP)/presenilin (PS1) transgenic mice | Ligature-induced periodontitis with P. gingivalis LPS injection into the periodontal tissue caused cognitive impairment and a significant reduction in the number of neurons. |
| [98] | 2020 Suh et al. | ApoE knockout mice with a high-fat diet | Mice received ligation with/without P. gingivalis LPS showed severe periodontitis, systemic inflammation, and aortic plaque formation. The magnitude of systemic inflammation and aortic plaque formation was notably greater in the ligated mice injected with P. gingivalis LPS. |
| [99] | 2020 O’Boyle et al. | C57BL/6 mice with focal cerebral ischemia induced by transient occlusion of the middle cerebral artery | Ligature-induced periodontitis with intravenous injection of LPS from P. gingivalis did not affect acute stroke pathology in terms of severity, determined primarily by infarct volume, despite the observation of elevated systemic inflammation. |
| [100] | 2018 Kim et al. | BRONJ and DRONJ models under the administration of zoledronic acid or anti- RANKL Ab | Pre-existing pathologic inflammatory condition exacerbated ONJ development after tooth extraction. |
| [101] | 2018 Handa et al. | Fbn-1C1039G/+ mice (C1039G/+) with Marfan syndrome | Fbn-1c1039G/+ mice exhibited slower wound healing compared to WT mice, but periodontal tissue destruction did not differ between these mice. |
| [102] | 2017 Coyac et al. | Phex gene-null mutant phenotype mice | Bone and cementum mineralization appeared to be similarly disturbed, where the hypomineralized pericellular matrix surrounded cells, and the protein osteopontin accumulated in a tissue-specific manner, most notably in the perilacunar matrix surrounding osteocytes. |
| [103] | 2017 Candeo et al. | BALB/c male mice with Asthma induced by ovalbumin injection | Ligature-induced periodontitis reduced the total number of cells in the bronchoalveolar lavage in a mouse model of asthma. |
| [104] | 2020 Rosa et al. | C57BL/6J mice with chronic obstructive pulmonary disease induced by cigarette extract | Ligature induced periodontitis decreased macrophages, TNF-α, and INF-γ expression in bronchoalveolar lavage. |
| [108] | 2019 Hays et al. | Pregnant C57BL/6 mice | The pregnant mice with ligation and P. gingivalis administration developed more severe alveolar bone loss. |
| [109] | 2016 Anbinder et al. | BALB/c with ovariectomy | Combination of periodontitis and ovariectomy induced a significantly higher femoral and mandibular bone loss than periodontitis or ovariectomy alone. |
| [110] | 2021 Arjunan et al. | C57BL/6J mice with Laser-induced choroidal neovascularization | An increase in mRNA expression related to oxidative stress, angiogenesis, and pro-inflammatory mediators in the retinae was observed, whereas antioxidant and anti-inflammatory-related gene expression was notably decreased. |
| [111] | 2020 Bi et al. | C57BL/6 male mice with unilateral ureteral ligation | Periodontitis increased mRNA expression of TNFα and IL-1β in the kidneys. Fibrotic areas in the kidneys in the mice with periodontitis were slightly, but not significantly, greater than those without periodontitis. |
| Reference No. | Year Author | Type of System | Main Findings |
|---|---|---|---|
| [113] | 2020 Ribeiro et al. | Cardiovascular system | Heart rate and arterial pressure variability were higher in mice with periodontitis. The mice with periodontitis showed decreased cardiac output and ejection fraction associated with increased myocardial cytokines. |
| [114] | 2020 Zhan et al. | Blood system | Experimental periodontitis induced platelet activation and platelet–leucocyte interaction. |
| [115] | 2021 Han et al. | Blood system | Lower red blood cell counts, hemoglobin, and hematocrit were detected in ligature-induced mice, whereas the levels of hepcidin mRNA expression and serum hepcidin concentrations increased. |
| [116] | 2016 Matsuda et al. | Digestive system | Ligation did not change the insulin sensitivity; it only affected the expression of Pck1 and Acaca in the liver. Ligature placement weakly affected the composition of gut microbiota and gene expression in the intestines. |
| [117] | 2021 Wang et al. | Central nervous system | Microgliosis and astrogliosis were notable in the ligated mice, and the number of glial cells displayed a positive correlation between the degree of periodontal inflammation. TNF-α was highly expressed in the hippocampus of ligated mice. |
| [118] | 2020 Furutama et al. | Central nervous system | In the hippocampus, the IL-1β expression levels were significantly increased by ligature-induced periodontitis. The ligation decreased the Cldn5 expression levels in the hippocampus, and the neutralization of IL-6 restored its levels. The blood-brain barrier was disrupted by ligature treatment. |
| [119] | 2020 Xue et al. | Central nervous system Digestive system | Long-term ligature placement caused progressive cognitive deficits, cerebral neuronal injury, synaptic injury, and glial activation. Ligation disrupted the intestinal barrier and blood-brain barrier and increased the brain LPS levels, TLR4 expression, NF-κB nuclear translocation, and proinflammatory cytokine mRNA levels. Ligated mice exhibited significant dysbiosis of the oral and gut microbiota. |
| [120] | 2020 Kitamoto et al. | Digestive system | Ligation in mice leads to expansion of Klebsiella and Enterobacter species in the oral cavity and translocation to the gut. The inflammation was triggered by the immigration of oral pathobionts and enhanced by the immigration of oral pathobiont-reactive Th17 cells in parallel. |
| Reference No. | Year Author | Cells and Receptors and Complements | Main Findings |
|---|---|---|---|
| [129] | 2020 Chadwick et al. | Neutrophils | Neutrophils in healthy oral cavity had the highest expression of CD11a and CD66a. The expression of CD11b was further upregulated by experimental periodontitis. |
| [11] | 2021 Fine et al. | Polymorphonuclear neutrophils (PMNs) | In periodontitis model in mice, PMNs were found to be elevated in the gingiva and bone marrow. The number of PMNs in the blood and colon of mice with induced periodontitis and peritonitis increased more than that in mice with acute peritonitis alone. |
| [131] | 2020 Kim et al. | Neutrophils | Neutrophils in gingival tissue of periodontitis induced RNAKL expression, leading to the early formation of osteoclasts. |
| [133] | 2016 Sima et al. | Polymorphonuclear neutrophils (PMNs) | In the Nrf2−/− mouse model of ligation-induced periodontitis, an increase in 8-OHdG–positive cells and more severe alveolar bone loss and breakdown of periodontal tissue were found. |
| [134] | 2021 Pathak et al. | Macrophages | Ligation-induced periodontitis in anti-Act1 mice exhibited a higher level of infiltration of macrophages in periodontal tissue and polarization of M1 macrophages. A higher degree of bone and tissue destruction with an increase in osteoclasts were also found. |
| [135] | 2019 Zhuang et al. | Macrophages | After treatment with CCL2, the loss of alveolar bone and the formation of osteoclasts in the ligation area were significantly reduced. M1/M2 ratio was also reduced. |
| [136] | 2016 Yamaguchi et al. | Macrophages | Transfer of M1 significantly inhibited bone loss and induction of TRAP-positive cells in mice with ligature-induced periodontitis. However, the transfer of M0 or M2 macrophages failed in achieving this efficacy. |
| [138] | 2017 Qin et al. | Lymphoid cells | All subtypes of innate lymphocytes were present in gingival tissues that were healthy or affected by periodontal disease. In periodontitis, the expression of IL-33, as well as ILC2s, were significantly increased through the regulation of AMPK. |
| [139] | 2019 Zheng et al. | Solitary chemosensory cells (SCC) | Induction of periodontitis by ligation in mice lacking SCC function showed more severe alveolar bone loss and gingival inflammatory infiltration. Meanwhile, colonization of the ligation sites was characterized by high bacterial load, low diversity, and high pathogen levels. Local activation of SCC function enhanced expression of β-defensin-3 (Defb3) in gingiva of mice to reduce bacterial load. |
| [141] | 2014 Lin et al. | TLR (TLR2 & TLR) | Alveolar bone loss produced by ligation-induced periodontitis is independent of TLR2 or TLR4. Elevated IL-1β and TNF-α with reduced IL-10 were also observed in all ligated models in mice (WT, Tlr2 KO, Tlr4 KO). |
| [126] | 2017 Lin et al. | TLR (TLR2 & TLR) | RNAKL protein expression and significant alveolar bone loss were observed in four mouse models of ligature-induced periodontitis (WT, Tlr2 KO, Tlr4 KO, Tlr2, and Tlr4 KO). |
| [142] | 2017 Crump et al. | TLR (TLR9) | Loss of alveolar bone and gingival inflammation were suppressed in Tlr9−/− mice, while the expression of A20 mRNA in the gingiva was increased. |
| [144] | 2019 Li et al. | A20 | Partial absence of A20 led to more severe alveolar bone loss and inflammatory cell infiltration in a ligature-induced periodontitis mouse model. |
| [145] | 2013 Gao et al. | ChemR23 (chemokine-like receptor 1) | In chemR23tg mice, ligation-induced periodontitis-related alveolar bone resorption was reduced. |
| [146] | 2012 Abe et al. | Complement C5a receptor (C5aR; CD88) | Mice with periodontitis treated with C5aRA at the ligation site showed approximately 50% reduction in bone loss and reduced levels of pro-inflammatory factors compared to the PBS-treated group. |
| [147] | 2014 Maekawa et al. | The third component of complement (C3) | In C3-deficient mice, the loss of alveolar bone caused by periodontitis was suppressed. |
| Reference No. | Year Author | Cell and ILs | Main Findings |
|---|---|---|---|
| [151] | 2018 Tsukasaki et al. | Th17 cells | Pathogenic Th17 cells prevented periodontitis progression by inducing mucosal immune response, alveolar bone damage, and tooth loss. |
| [152] | 2021 Pacheco et al. | IL-17 | In periodontitis model in mice, Il17a expression correlates with the time point of ligation induction. IL-17A antibody reduced IL-6 expression, osteoclastic activity, and alveolar bone loss in murine models. |
| [153] | 2020 Sun et al. | IL-17 | Higher levels of Il17 and IL-17-associated chemokines and cytokines were observed in IL-10-deficient mice with periodontitis. Moreover, the polarization of M1 macrophages and osteoclast differentiation activity in the gingiva were increased. |
| [154] | 2013 Gonçalves-Zillo et al. | IL-17 | On day 21, increased IL-17 secretion was observed in all periodontitis mouse models. However, no significant changes in IL-10 and TNF- α levels were observed in Bdkrb1−/− mice. |
| [155] | 2015 Abe et al. | B cells | Alveolar bone loss was significantly ameliorated in B-cell-deficient mice. After using APRIL and BLyS antibodies, significant inhibition of B cells in gingival tissue and alveolar bone resorption was observed. |
| [156] | 2017 Yu et al. | B10 cells | Local injection of CpG with CD40L into the gingiva induced B10 cell activity, which increased IL-10 mRNA expression and significantly inhibited alveolar bone loss and expression of pro-inflammatory factors in the gingiva. |
| [157] | 2018 Zhao et al. | B10 cells | In ligation-induced periodontitis in Tlr9-deficient mice, treatment by CpG ODNs in combination with CD40L still significantly improved periodontal destruction. |
| [158] | 2017 Hu et al. | B10 cells | Using co-stimulatory molecules (CD40L, IL-21, and anti-Tim-1 mAb) for B10 cells in ligation-induced periodontitis mice could also upregulate the mRNA expression of gingival IL-10 while decreasing the expression of RANKL. |
| Reference No. | Year Author | Cytokines and Molecules and Genes | Main Findings |
|---|---|---|---|
| [159] | 2019 Bi et al. | β6 integrin | The absence of integrin αvβ6 in the junctional epithelium could lead to the downregulation of the Aim2 inflammasome and anti-inflammatory IL-10. |
| [160] | 2017 Hirschfeld et al. | Macrophage migration inhibitory factor (MIF) | In Mif-deficient mice, the compensatory upregulation of IL-6 and the downregulation of corticosterone levels were observed. However, the release of MMP2 in the periodontal tissue of mice did not seem to be affected by MIF. |
| [161] | 2017 Papadakou et al. | Vascular endothelial growth factor C (VEGFC) | In K14-VEGFC mice, lymphatic vessel hyperplasia occurred, but the severity of bone loss and inflammation did not decrease in the experimental periodontitis. |
| [12] | 2010 Ohnishi et al. | Cot/Tp12 | Compared to WT mice, less alveolar bone loss, osteoclast formation, and expression of TNF-α were observed in ligation-induced periodontitis in cot/tp12-deficient (cot/tp12−/−) mice. |
| [162] | 2018 Zhou et al. | MicroRNA-21 | In the MicroRNA21 knockout mice with periodontitis, alveolar bone resorption and inflammatory infiltration in gingival were both elevated. |
| [163] | 2018 Ouhara et al. | Human antigen-R (HuR) | HuR stabilizes IL-6 in gingival tissue. Quercetin, as a HuR inhibitor, can lower the level of bone resorption in ligature-induced periodontitis mice model. |
| [167] | 2016 Suzuki et al. | Glucose-dependent insulinotropic polypeptide (GIP) | Gipr-knockout mice with ligature-induced periodontitis also showed a significant increase in the gene expression of gingival inflammatory cytokines, TNF-α, and inducible nitric oxide synthase (iNOS), as compared to WT mice with experimental periodontitis. |
| [168] | 2013 Rajshankar et al. | Protein tyrosine phosphatase-α (PTPα) | Loss of alveolar bone, reduction in gingival lamina propria thickness, and collagen fibril number were suppressed in PTPα−/− mice. |
| Reference No. | Year Author | Target | Method | Main Findings |
|---|---|---|---|---|
| [169] | 2020 Kittaka et al. | Oral microbiota | 16S rDNA amplicon sequencing | The most predominant bacteria in ligatures in a mouse model of cherubism were Pasteurellales. The most predominant bacteria could be a trigger for the initiation of jawbone destruction in human cherubism. |
| [170] | 2020 Williams et al. | Oral microbiota Gut microbiota | 16S rDNA amplicon sequencing | The normal oral microbiota in mice protects against inflammation-induced osteonecrosis. |
| [119] | 2020 Xue et al. | Oral microbiota Gut microbiota | 16S rDNA amplicon sequencing | Proteobacteria were the most abundant phylum detected in both ligature-induced periodontitis mice and mice without periodontitis, although Proteobacteria were significantly less abundant in ligature-induced periodontitis mice compared with mice without periodontitis. |
| [171] | 2020 Hiyoshi et al. | Oral microbiota | Counts of colony-forming units | Hinokitiol treatment significantly inhibited the alveolar bone loss and osteoclast differentiation induced by tooth ligation. |
| [172] | 2020 Kim et al. | Oral microbiota | Real-time PCR | An oral care probiotic, Weissella cibaria CMU, reduced periodontal tissue destruction by regulating the inflammatory cytokines and by reducing oral bacteria in ligature-induced periodontitis mice. |
| [173] | 2020 Huang et al. | Gut microbiota | 16S rDNA amplicon sequencing | Periodontitis led to gut microbiota dysbiosis in mice with hyperlipidemia. Non-surgical periodontal treatment normalized the gut microbiota. |
| [174] | 2021 Li et al. | Gut microbiota | 16S rDNA amplicon sequencing | Parabacteroides and Desulfovibrionaceae increased, and several butyrate-producing bacteria decreased significantly in the gut microbiota of ligature-induced periodontitis mice compared to control mice. |
| Reference No. | Year Author | Periodontal Enhance Factor | Main Findings |
|---|---|---|---|
| [9] | 2021 Akkaoui et al. | Ligation with injection of LPS from P. gingivalis | P. gingivalis-LPS mice elevated the pro-inflammatory cytokines compared to control mice. |
| [175] | 2021 An et al. | Ligation with oral feeding of P. gingivalis | A monoclonal antibody that targeted the DHYAVMISK peptide might protect against a P. gingivalis infection. |
| [176] | 2017 Bhattarai et al. | Ligation with injection of LPS from P. gingivalis | Genistein protected against alveolar bone loss and periodontal tissue destruction in LPS/ligature-induced periodontitis mice. |
| [177] | 2021 Cao et al. | A ligature inoculated with P. gingivalis | Local transfer of CD19+ CD1dhi CD5+ B cells could inhibit attenuate alveolar bone loss in periodontitis mice by ligatures inoculated with P. gingivalis. |
| [178] | 2021 Clark et al. | A ligature inoculated with P. gingivalis | The age-related changes to the macrophage contributed to the pathogenesis of periodontal disease. |
| [179] | 2018 Francis et al. | A ligature inoculated with P. gingivalis | Keratinocyte-specific ablation of protease-activated receptor 2 prevented the increase in the number of osteoclasts and the up-regulation of the inflammatory markers in ligature-induced periodontitis mice. |
| [180] | 2019 Ideguchi et al. | A ligature inoculated with P. gingivalis | Levels of serum IgG antibody against P. gingivalis were significantly higher in the ligature + P. gingivalis group than in the P. gingivalis + glycyrrhizic group. |
| [181] | 2016 Lapérine et al. | A ligature inoculated with P. gingivalis | IL-33 expression of periodontitis induced by a P. gingivalis-soaked ligature was increased in gingival epithelial cells similarly as in human CP. |
| [182] | 2017 Nagashima et al. | A ligature inoculated with P. gingivalis | Periodontal CXC-chemokine receptor 4 signaling in several cell types such as fibroblasts, macrophages, osteoblasts, and osteoclasts in P. gingivalis-induced periodontitis depresses alveolar bone resorption in periodontitis. |
| [183] | 2020 Wang et al. | A ligature inoculated with P. gingivalis | NIK-SMI1 treatment resulted in attenuated periodontitis progression and pro-inflammatory cytokines expression in periodontitis mouse model. |
| [184] | 2020 Wang et al. | A ligature inoculated with P. gingivalis | Halofuginone significantly reduced the expression levels of pro-inflammatory cytokines, and markedly suppressed immune cell infiltration into the infected sites. |
| [185] | 2021 Wang et al. | A ligature inoculated with P. gingivalis | The local infiltration of B10 cells into periodontal tissue promoted anti-inflammatory responses in ligature-induced periodontitis mice. |
| [186] | 2017 Wang et al. | A ligature inoculated with P. gingivalis | The adoptive transfer of B10 cells alleviated periodontal inflammation and bone loss in experimental periodontitis mice. |
| [187] | 2020 Korah et al. | A ligature inoculated with P. gingivalis | Osteonecrosis area and osteoclast number were significantly elevated in Msx2 knock-in periodontitis mice compared with wild-type periodontitis mice. |
| [188] | 2007 Li et al. | A ligature inoculated with P. gingivalis | The ligature inoculated with P. gingivalis group showed significantly increased epithelial downgrowth, inflammation, alveolar bone loss, and osteoclast activity throughout the experimental period compared to the controls. |
| [189] | 2019 Palioto et al. | Ligation with P. gingivalis gavage | Ligature, P. gingivalis gavage, and ligature with P. gingivalis gavage groups induced significant bone loss compared to the sham group. In terms of the inflammatory markers and epigenetic changes, the alteration of P. gingivalis gavage and ligature with P. gingivalis gavage groups were significantly higher than that of local ligature-induced periodontitis. |
| [10] | 2019 Qi et al. | A ligature inoculated with P. gingivalis | Inhibition TLR4 in vivo significantly improved the alveolar bone resorption in periodontitis model mice. |
| [190] | 2021 Wang et al. | A ligature inoculated with P. gingivalis | Treatment of TPCA-1 inhibited the osteoclastogenesis through the inactivation of NF-κB pathway in mouse chronic periodontitis model. |
| [191] | 2016 Yang et al. | A ligature inoculated with P. gingivalis | Periodontitis mice induced by ligation with P. gingivalis showed significantly more severe alveolar bone loss than control mice. |
| [192] | 2019 Sulijaya et al. | Ligation with oral feeding of P. gingivalis | KetoC attenuated alveolar bone destruction and suppressed the abundance level of P. gingivalis in the ligature-induced periodontitis mice. |
| [193] | 2018 Duan et al. | Ligation combined with P. gingivalis infection | Ligation itself did not cause higher gingival inflammation and bone loss in pregnant mice than non-pregnant mice, while ligation combined with P. gingivalis infection led to increased gingival inflammation and periodontal bone loss. |
| [194] | 2000 Kimura et al. | A ligature inoculated with P. gingivalis | The ligature + P. gingivalis mice showed alveolar bone loss in the infected sites which was found to be greater than that in the periodontitis mice induced by ligation alone. |
| [195] | 2018 Bugueno et al. | A ligature inoculated with P. gingivalis | An increased expression of apoptotic peptidase activating factor 1 was observed in a murine experimental periodontitis model induced by P. gingivalis soaked ligatures. |
| [200] | 2016 Kubota et al. | Ligation with injection of nicotine and cigarette smoke condensate | Nicotine and cigarette smoke condensate promoted periodontal destruction in mice with a ligature, accompanied by increased osteoclastogenesis and RANKL expression. |
| Reference No. | Year Author | Treatment | Main Findings |
|---|---|---|---|
| [203] | 2017 Shi et al. | Systemic antibiotics | Systemic antibiotic administration reduced bone loss. Gingival mRNA and protein RANKL/OPG expression ratio was decreased. |
| [204] | 2020 Maekawa et al. | Erythromycin | ERM suppressed inflammation and inhibited bone loss via activation of DEL-1 expression. |
| [205] | 2021 Tamura et al. | Erythromycin | ERM inhibited alveolar bone loss by increasing Del1 expression and decreasing the expression of osteoclast-related factors. |
| [211] | 2017 Wisitrasameewong et al. | Anti-DC-STAMP-mAb | Local injection and systemic administration of anti-DC-STAMP-mAb significantly downregulated the alveolar bone loss. DC-STAMP promotes local osteoclast cell fusion without affecting adaptive immune responses to oral bacteria. |
| [214] | 2017 Maekawa et al. | Secreted frizzled-related protein 5 (sFRP5) | Local sFRP5 administration inhibited inflammation and bone absorption and decreased the number of osteoclasts in bone tissue. |
| [217] | 2019 Tamura et al. | Rice endosperm protein (REP) 9 and 11 | Local treatment with REP 9 and 11 inhibited the activity of inflammatory and osteoclast-related molecules and significantly decreased bone resorption. |
| [218] | 2019 Aoki-Nonaka et al. | Amyl-1-18 | Amyl-1-18 prevented alveolar bone destruction via suppression of LPS-induced inflammatory cytokine production. |
| [219] | 2015 Mizuno et al. | Reveromycin A (RMA) | RMA treatment decreased osteoclasts, alveolar bone loss, attachment loss, and inflammatory cytokine expression. |
| [226] | 2016 Huang et al. | Myricetin | Myricetin inhibited osteoclast formation and prevented alveolar bone loss in an OVX mouse model. |
| [230] | 2019 Adhikari et al. | Oleanolic acid acetate (OAA) | OAA induced bone formation and remodeling via proper modulation of osteoblasts, osteoclasts, and inflammation by regulating TGF-β and Wnt signaling. |
| [233] | 2020 Kim et al. | 6-Shogaol | Administration of 6-shogaol prevented osteoclastogenesis and alveolar bone resorption, decreased the number of macrophages and neutrophils, and downregulated the expression of IL-1β and TNF-α in periodontal tissue. |
| [234] | 2017 Ihn et al. | A novel pyrimidine compound (OCLI-023) | OCLI-023 inhibited ligature-induced bone loss and reduced the number of osteoclasts induced by periodontitis. |
| [235] | 2018 Ihn et al. | A novel benzamide-linked derivative (OCLI-070) | OCLI-070 inhibited osteoclast formation and differentiation, reduced Nfatc1 and the expression of osteoclast-specific genes, and prevented bone resorption via the suppression of RANKL-mediated ERK and NF-κB signaling pathways. |
| [238] | 2020 Jiang et al. | LLP2A-alendronate (LLP2A-Ale) | LLP2A-Ale stimulated alveolar bone formation, reversed bone loss, and reduced the levels of periodontitis-induced circulating inflammatory cytokines. |
| [239] | 2020 Cafferata et al. | Boldine | Boldine decreased osteoclast numbers and the RANKL/OPG ratio in periodontal sites. The Th17-lymphocyte detection and response were reduced, and the Treg-lymphocyte detection and response were increased by boldine. |
| [240] | 2015 Zhen et al. | Resveratrol | Blood glucose levels were decreased, alveolar bone loss was ameliorated, and high levels of IL-1β, IL-6, IL-8, TNF-α, and TLR4 were suppressed by resveratrol in the gingival tissue. |
| [241] | 2018 Ikeda et al. | Resveratrol derivative-rich merinjo (MSE) | MSE decreased oxidative stress, inhibited M-CSF/sRANKL-mediated osteoclast formation, and downregulated osteoclast activity on the periodontal side. |
| [47] | 2019 Akutagawa et al. | Glycyrrhizic acid | Glycyrrhizic acid suppressed the release of inflammatory cytokines, high mobility group box 1, and receptor for AGE, which were increased by ligature/P. gulae at the mRNA level in gingival tissue and the protein level in the serum of diabetic mice. |
| [242] | 2020 Kaboosaya et al. | Green tea | Systemic administration of green tea relieved alveolar bone loss and reduced the number of inflammatory cells and osteoclasts. |
| [243] | 2016 Cui et al. | Scaling and root planing | Prevention of alveolar bone loss, improvement of lipid profile, and inhibition of systemic inflammation with reduced plasma IL-6 levels were observed. |
| [244] | 2020 Ying et al. | Low-intensity pulsed ultrasound (LIPUS) | LIPUS treatment alleviated alveolar bone homeostasis in periodontitis by downregulating oxidative stress via PI3K-Akt/nuclear factor erythroid 2-related factor (NRF2) signaling. |
| [245] | 2021 Snipes et al. | Inhibitor of sphingosine-1-phosphate receptor 2 (JTE013) | JTE013 attenuated ligature-induced alveolar bone loss and reduced inflammation-related gene expression levels. |
| [246] | 2019 Kure et al. | IkB kinase inhibitor (IMD-0354) | IMD-0354 suppressed bone loss and RANKL gene expression levels in gingival tissue by downregulating NF-κB. |
| [247] | 2021 Ihn et al. | Fatty acid amide hydrolase inhibitor (PF-3845) | PF-3845 demonstrated anti-osteoclastogenic and anti-resorptive activities by suppressing the phosphorylation of rapidly accelerated fibrosarcoma (RAF), mitogen-activated protein kinase (MEK), extracellular signal-regulated kinase (ERK), and NF-κB inhibitor (IκBα). |
| [248] | 2021 Tanaka et al. | DNA methyltransferase inhibitor (Decitabine) | Decitabine inhibited osteoclastogenesis through the upregulation of anti-inflammatory cytokines via Krüppel-like factor-2 (KLF2)-dependent mechanisms. |
| [249] | 2019 Oliveira et al. | Renin inhibitor (Aliskiren) | Aliskiren treatment reduced gene expression of renin-angiotensin system (RAS) components in the gingival tissue of normal and diabetic mice. Administration of aliskiren also downregulated the gene expression of COL1A1, COL1A2, and fibronectin that was increased by periodontitis. |
| [98] | 2020 Suh et al. | Treatment of atherosclerosis (Rosuvastatin) | Rosuvastatin inhibited the TNF-α-induced osteoclast formation, endothelial cell phenotypic changes, foam cell formation, and the expression of CD47 and other oncogenes in arterial smooth muscle cells related to atherosclerosis. |
| [250] | 2017 Iwamatsu-Kobayashi et al. | Metal ions from S-PRG filler | The S-PRG metal ion suppressed the destruction of the collagen bundle in the periodontal ligament and inhibited the infiltration of inflammatory cells. The number of infiltrating neutrophils and macrophages was significantly suppressed. |
| [251] | 2016 Yu et al. | pH-responsive hydrogel with an anti-glycation agent (PTB) | PTB significantly reduced periodontal bone levels (PBL) and inflammatory cell infiltration in experimental periodontitis plus hydrogel with PTB during the induction phase. PTB significantly decreased PBL and inflammatory cell infiltration, and significantly greater collagen deposition was observed in both hydrogel groups with and without PTB at 4 and 14 days after removing ligature. |
| [252] | 2017 Iguchi et al. | Local bone marrow stromal cell (BMSC) | Alveolar bone loss around the ligated molars was significantly reduced in BMSC-treated mice. Inflammatory infiltration was also decreased, and the number of TRAP-positive osteoclasts was notably increased by BMSC treatment. |
| [253] | 2021 Nakao et al. | Exosomes from TNF-alpha-treated human gingiva-derived MSCs (GMSCs) | Local injection of GMSC-derived exosomes significantly decreased bone loss and the number of TRAP-positive osteoclasts, and these effects were further enhanced by preconditioning of GMSCs with TNF-α. The exosome with TNF-α reduced RANKL mRNA expression and significantly elevated OPG mRNA expression. |
| [254] | 2020 Nagai et al. | Hydrogel-formulated inhibitor of prolyl-4-hydroxylase (1,4-DPCA/hydrogel) | Injection of 1,4-DPCA/hydrogel increased gingival hypoxia-inducible factor 1α (HIF-1α) protein levels and alveolar bone regeneration. The 1,4-DPCA/hydrogel-induced regeneration of alveolar bone was associated with enhanced expression of osteogenic genes, decreased expression of inflammation-related genes, and increased abundance of Tregs in the periodontal tissue. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, P.; Niimi, H.; Ohsugi, Y.; Tsuchiya, Y.; Shimohira, T.; Komatsu, K.; Liu, A.; Shiba, T.; Aoki, A.; Iwata, T.; et al. Application of Ligature-Induced Periodontitis in Mice to Explore the Molecular Mechanism of Periodontal Disease. Int. J. Mol. Sci. 2021, 22, 8900. https://doi.org/10.3390/ijms22168900
Lin P, Niimi H, Ohsugi Y, Tsuchiya Y, Shimohira T, Komatsu K, Liu A, Shiba T, Aoki A, Iwata T, et al. Application of Ligature-Induced Periodontitis in Mice to Explore the Molecular Mechanism of Periodontal Disease. International Journal of Molecular Sciences. 2021; 22(16):8900. https://doi.org/10.3390/ijms22168900
Chicago/Turabian StyleLin, Peiya, Hiromi Niimi, Yujin Ohsugi, Yosuke Tsuchiya, Tsuyoshi Shimohira, Keiji Komatsu, Anhao Liu, Takahiko Shiba, Akira Aoki, Takanori Iwata, and et al. 2021. "Application of Ligature-Induced Periodontitis in Mice to Explore the Molecular Mechanism of Periodontal Disease" International Journal of Molecular Sciences 22, no. 16: 8900. https://doi.org/10.3390/ijms22168900
APA StyleLin, P., Niimi, H., Ohsugi, Y., Tsuchiya, Y., Shimohira, T., Komatsu, K., Liu, A., Shiba, T., Aoki, A., Iwata, T., & Katagiri, S. (2021). Application of Ligature-Induced Periodontitis in Mice to Explore the Molecular Mechanism of Periodontal Disease. International Journal of Molecular Sciences, 22(16), 8900. https://doi.org/10.3390/ijms22168900