
Analysis and Fine Specificity of the HCMV-Specific Cell-Free and Cell-Associated Antibody-Dependent Cellular Phagocytosis (ADCP) Responses in Lung Transplant Recipients

 , , and
, , and {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Study Cohort and Antibody Profile
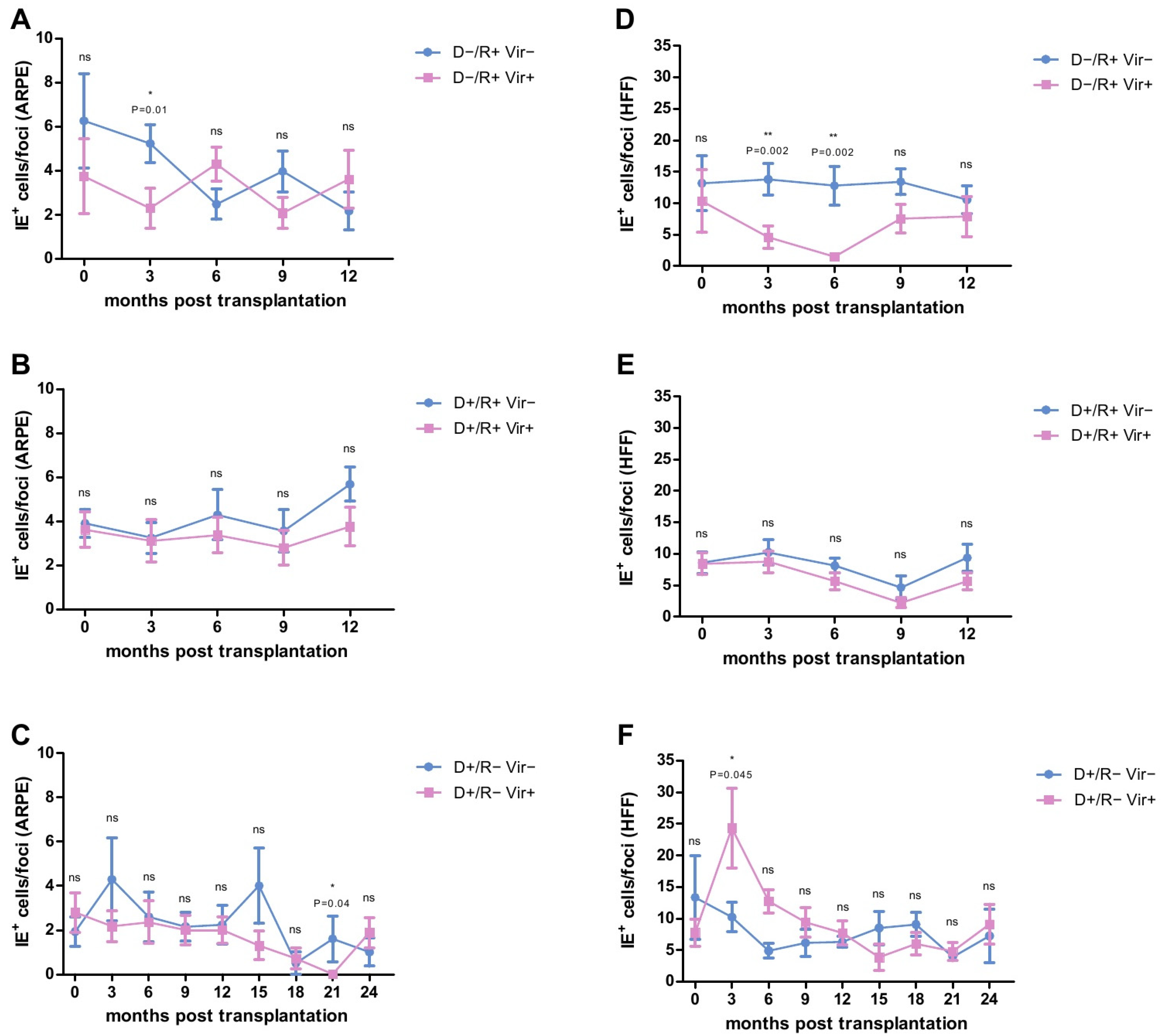
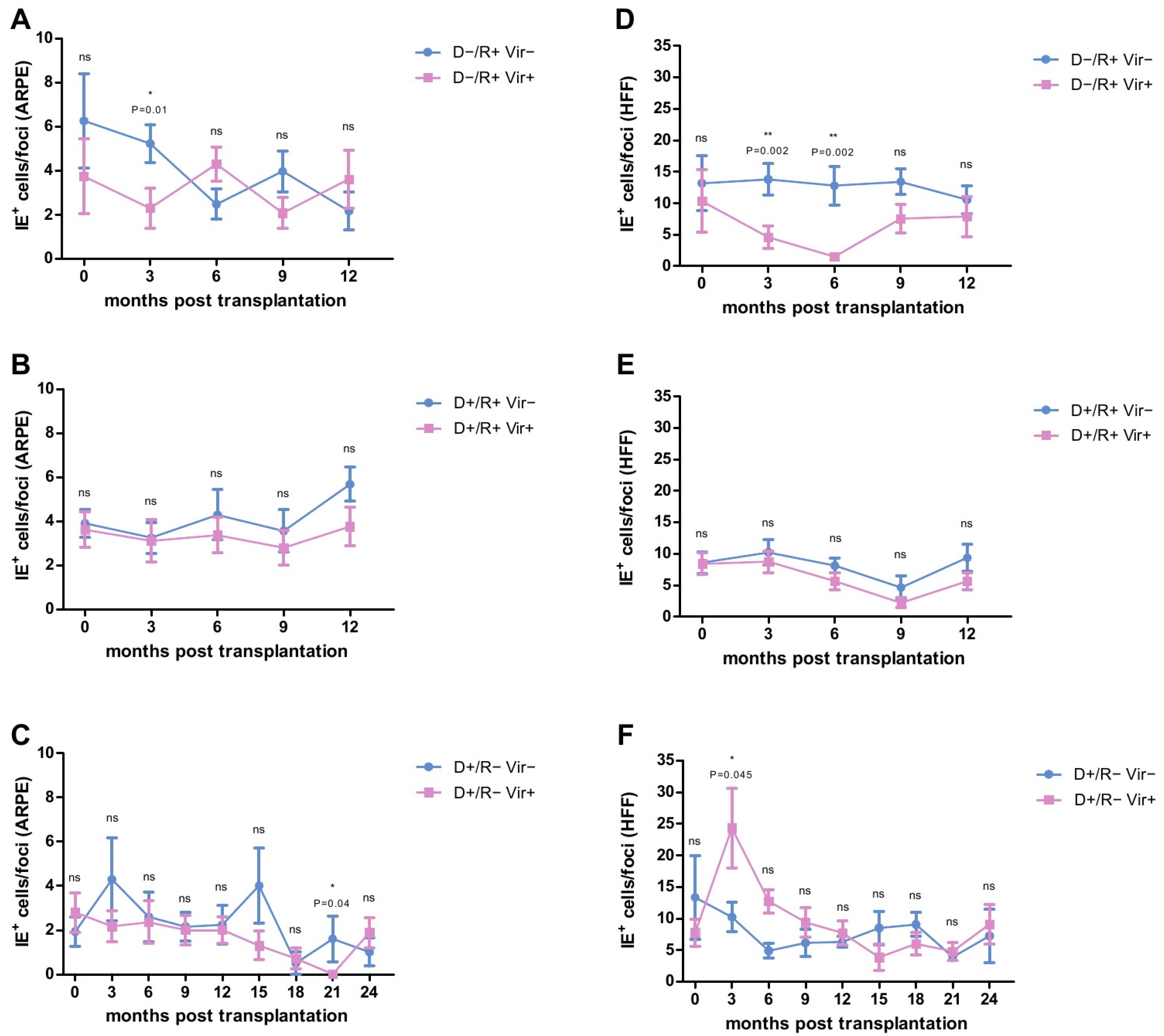
2.2. Analysis of the HCMV-Specific ADCP Response against Infected Cells
2.3. HCMV-Specific ADCP Response in Viremic- and No/Low Viremic R+ LTRs
2.4. HCMV-Specific ADCP Response in Viremic and No/Low Viremic D+/R− LTRs
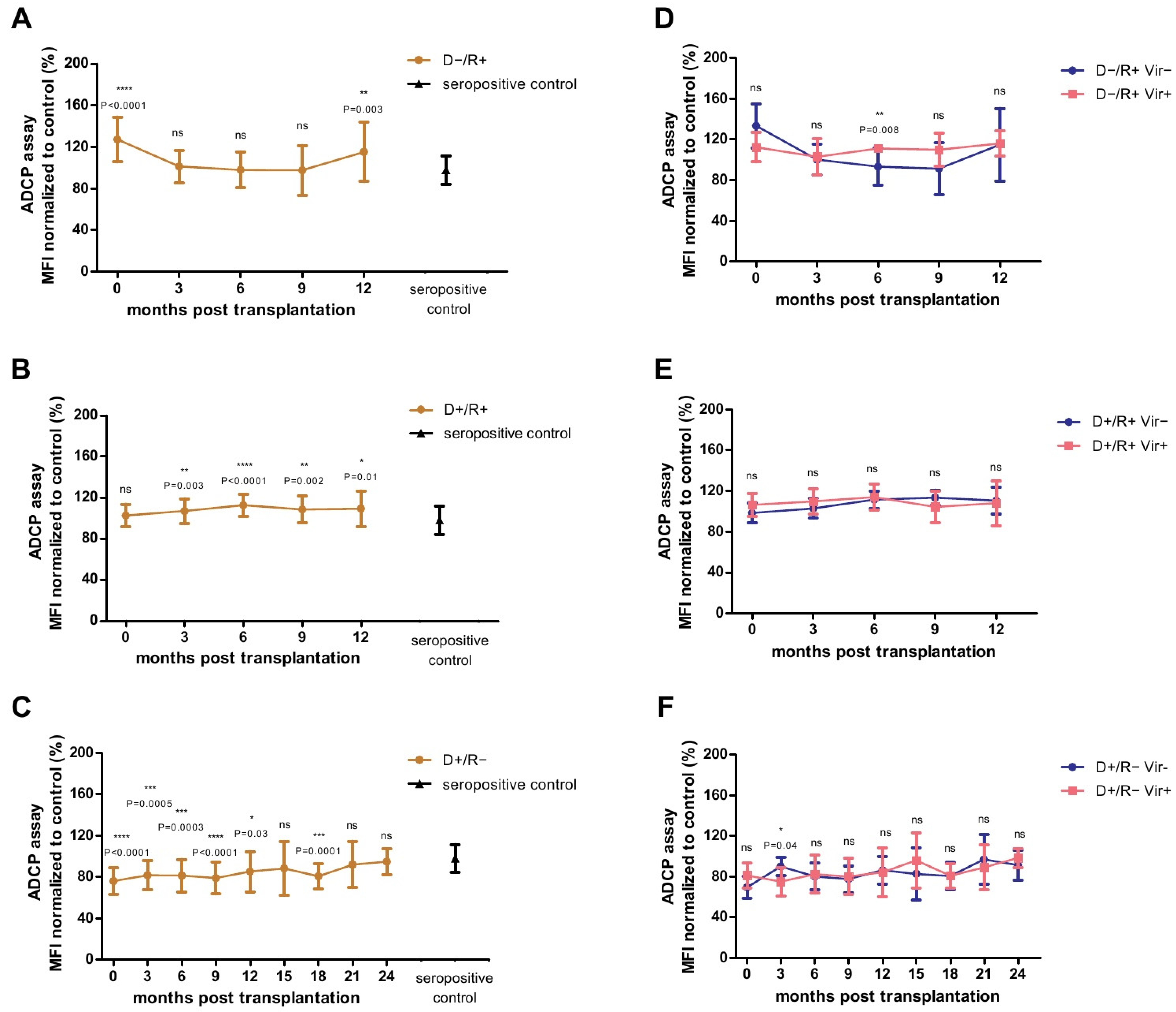
2.5. Analysis of the HCMV-Specific ADCP Response against Cell-Free Virions
2.6. Association between HCMV-Specific Adcp Response against Cell-Free Virions and Viremia
2.7. Correlation of the ADCP Response and the HCMV-Specific Antibody Titers
2.8. Analysis of the ADCP Response over Time in Relation to Primary Infection
3. Discussion
4. Material and Methods
4.1. Patients and Samples
4.2. Cells and Virus
4.3. Focus Expansion Assay (FEA)
4.4. Whole HCMV Virion Phagocytosis Assay
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ADCC | antibody-dependent cellular cytotoxicity |
| ADCP | antibody-dependent cellular phagocytosis |
| D− | donor HCMV-seronegative |
| D+ | donor HCMV-seropositive |
| FcγR | Fcγ receptors |
| FEA | focal expansion assay |
| gB | glycoprotein B |
| HCMV | human cytomegalovirus |
| HFF | human foreskin fibroblasts |
| IE antigen | immediate early antigen |
| LTX | lung transplantation |
| LTR | lung transplant recipients |
| MFI | mean fluorescent intensity |
| MOI | multiplicity of infection |
| NK cell | natural killer cell |
| NPV | positive predictive values |
| OR | odds ratio |
| PC | pentameric complex |
| PPV | negative predictive values |
| R− | recipient HCMV-seronegative |
| R+ | recipient HCMV-seropositive |
| SOT | solid organ transplantation |
| TX | transplantation |
References
- Razonable, R.R.; Humar, A. Cytomegalovirus in solid organ transplant recipients—guidelines of the American Society of Transplantation Infectious Diseases Community of Practice. Clin. Transplant. 2019, 33, e13512. [Google Scholar] [CrossRef] [PubMed]
- Gomes, A.C.; Griffiths, P.D.; Reeves, M.B. The humoral immune response against the gB vaccine: Lessons learnt from protection in solid organ transplantation. Vaccines 2019, 7, 67. [Google Scholar] [CrossRef] [Green Version]
- Jacob, C.L.; Lamorte, L.; Sepulveda, E.; Lorenz, I.C.; Gauthier, A.; Franti, M. Neutralizing antibodies are unable to inhibit direct viral cell-to-cell spread of human cytomegalovirus. Virology 2013, 444, 140–147. [Google Scholar] [CrossRef] [Green Version]
- Scrivano, L.; Sinzger, C.; Nitschko, H.; Koszinowski, U.H.; Adler, B. HCMV spread and cell tropism are determined by distinct virus populations. PLoS Pathog. 2011, 7, e1001256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, C.S.; Huffman, T.; Jenks, J.A.; de la Rosa, E.C.; Xie, G.; Vandergrift, N.; Permar, S.R. HCMV glycoprotein B subunit vaccine efficacy mediated by nonneutralizing antibody effector functions. Proc. Natl. Acad. Sci. USA 2018, 115, 6267–6272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffiths, P.D.; Stanton, A.; McCarrell, E.; Smith, C.; Osman, M.; Harber, M.; Burroughs, A.K. Cytomegalovirus glycoprotein-B vaccine with MF59 adjuvant in transplant recipients: A phase 2 randomised placebo-controlled trial. Lancet 2011, 377, 1256–1263. [Google Scholar] [CrossRef] [Green Version]
- Baraniak, I.; Kropff, B.; Ambrose, L.; McIntosh, M.; McLean, G.R.; Pichon, S.; Reeves, M.B. Protection from cytomegalovirus viremia following glycoprotein B vaccination is not dependent on neutralizing antibodies. Proc. Natl. Acad. Sci. USA 2018, 115, 6273–6278. [Google Scholar] [CrossRef] [Green Version]
- Tay, M.Z.; Wiehe, K.; Pollara, J. Antibody-dependent cellular phagocytosis in antiviral immune responses. Front. Immunol. 2019, 10, 332. [Google Scholar] [CrossRef]
- Rosales, C.; Uribe-Querol, E. Phagocytosis: A fundamental process in immunity. BioMed Res. Int. 2017, 2017, 9042851. [Google Scholar] [CrossRef] [Green Version]
- Nimmerjahn, F.; Ravetch, J.V. Fcγ receptors as regulators of immune responses. Nat. Rev. Immunol. 2008, 8, 34–47. [Google Scholar] [CrossRef]
- Uribe-Querol, E.; Rosales, C. Phagocytosis: Our current understanding of a universal biological process. Front. Immunol. 2020, 11, 1066. [Google Scholar] [CrossRef] [PubMed]
- Nelson, C.S.; Herold, B.C.; Permar, S.R. A new era in cytomegalovirus vaccinology: Considerations for rational design of next-generation vaccines to prevent congenital cytomegalovirus infection. Npj Vaccines 2018, 3, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Vietzen, H.; Görzer, I.; Honsig, C.; Jaksch, P.; Puchhammer-Stöckl, E. Human Cytomegalovirus (HCMV)-Specific Antibody Response and Development of Antibody-Dependent Cellular Cytotoxicity Against HCMV After Lung Transplantation. J. Infect. Dis. 2020, 222, 417–427. [Google Scholar] [CrossRef]
- Brait, N.; Stögerer, T.; Kalser, J.; Adler, B.; Kunz, I.; Benesch, M.; Görzer, I. Influence of human cytomegalovirus glycoprotein O polymorphism on the inhibitory effect of soluble forms of trimer-and pentamer-specific entry receptors. J. Virol. 2020, 94, e00107-20. [Google Scholar] [CrossRef] [PubMed]
- Britt, W.J.; Vugler, L.G. Processing of the gp55-116 envelope glycoprotein complex (gB) of human cytomegalovirus. J. Virol. 1989, 63, 403–410. [Google Scholar] [CrossRef] [Green Version]
- Corrales-Aguilar, E.; Hoffmann, K.; Hengel, H. CMV-encoded Fcγ receptors: Modulators at the interface of innate and adaptive immunity. In Seminars in Immunopathology; Springer: Berlin/Heidelberg, Germany, 2014; Volume 36, pp. 627–640. [Google Scholar]
- Corrales-Aguilar, E.; Trilling, M.; Hunold, K.; Fiedler, M.; Le VT, K.; Reinhard, H.; Hengel, H. Human cytomegalovirus Fcγ binding proteins gp34 and gp68 antagonize Fcγ receptors I, II and III. PLoS Pathog. 2014, 10, e1004131. [Google Scholar] [CrossRef] [Green Version]
- Kolb, P.; Hoffmann, K.; Sievert, A.; Reinhard, H.; Merce-Maldonado, E.; Le-Trilling VT, K.; Hengel, H. Human cytomegalovirus antagonizes activation of Fcγ receptors by distinct and synergizing modes of IgG manipulation. Elife 2021, 10, e63877. [Google Scholar] [CrossRef]
- Baraniak, I.; Kern, F.; Holenya, P.; Griffiths, P.; Reeves, M. Original antigenic sin shapes the immunological repertoire evoked by human cytomegalovirus glycoprotein B/MF59 vaccine in seropositive recipients. J. Infect. Dis. 2019, 220, 228–232. [Google Scholar] [CrossRef]
- Sinzger, C.; Digel, M.; Jahn, G. Cytomegalovirus cell tropism. Hum. Cytomegal. 2008, 325, 63–83. [Google Scholar]
- Schampera, M.S.; Schweinzer, K.; Abele, H.; Kagan, K.O.; Klein, R.; Rettig, I.; Hamprecht, K. Comparison of cytomegalovirus (CMV)-specific neutralization capacity of hyperimmunoglobulin (HIG) versus standard intravenous immunoglobulin (IVIG) preparations: Impact of CMV IgG normalization. J. Clin. Virol. 2017, 90, 40–45. [Google Scholar] [CrossRef]
- Jaksch, P.; Ankersmit, J.; Scheed, A.; Kocher, A.; Muraközy, G.; Klepetko, W.; Lang, G. Alemtuzumab in lung transplantation: An open-label, randomized, prospective single center study. Am. J. Transplant. 2014, 14, 1839–1845. [Google Scholar] [CrossRef]
- Freedman, M.S.; Kaplan, J.M.; Markovic-Plese, S. Insights into the mechanisms of the therapeutic efficacy of alemtuzumab in multiple sclerosis. J. Clin. Cell. Immunol. 2013, 4, 1000152. [Google Scholar] [CrossRef] [PubMed]
- Kannegieter, N.M.; Hesselink, D.A.; Dieterich, M.; Kraaijeveld, R.; Rowshani, A.T.; Leenen, P.J.; Baan, C.C. The effect of tacrolimus and mycophenolic acid on CD14+ monocyte activation and function. PLoS ONE 2017, 12, e0170806. [Google Scholar]
- Ehrchen, J.M.; Roth, J.; Barczyk-Kahlert, K. More than suppression: Glucocorticoid action on monocytes and macrophages. Front. Immunol. 2019, 10, 2028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foglierini, M.; Marcandalli, J.; Perez, L. HCMV envelope glycoprotein diversity demystified. Front. Microbiol. 2019, 10, 1005. [Google Scholar] [CrossRef] [PubMed]
- Frey, A.; Di Canzio, J.; Zurakowski, D. A statistically defined endpoint titer determination method for immunoassays. J. Immunol. Methods 1998, 221, 35–41. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eberhard, S.; Vietzen, H.; Görzer, I.; Jaksch, P.; Puchhammer-Stöckl, E. Analysis and Fine Specificity of the HCMV-Specific Cell-Free and Cell-Associated Antibody-Dependent Cellular Phagocytosis (ADCP) Responses in Lung Transplant Recipients. Int. J. Mol. Sci. 2021, 22, 8206. https://doi.org/10.3390/ijms22158206
Eberhard S, Vietzen H, Görzer I, Jaksch P, Puchhammer-Stöckl E. Analysis and Fine Specificity of the HCMV-Specific Cell-Free and Cell-Associated Antibody-Dependent Cellular Phagocytosis (ADCP) Responses in Lung Transplant Recipients. International Journal of Molecular Sciences. 2021; 22(15):8206. https://doi.org/10.3390/ijms22158206
Chicago/Turabian StyleEberhard, Simone, Hannes Vietzen, Irene Görzer, Peter Jaksch, and Elisabeth Puchhammer-Stöckl. 2021. "Analysis and Fine Specificity of the HCMV-Specific Cell-Free and Cell-Associated Antibody-Dependent Cellular Phagocytosis (ADCP) Responses in Lung Transplant Recipients" International Journal of Molecular Sciences 22, no. 15: 8206. https://doi.org/10.3390/ijms22158206
APA StyleEberhard, S., Vietzen, H., Görzer, I., Jaksch, P., & Puchhammer-Stöckl, E. (2021). Analysis and Fine Specificity of the HCMV-Specific Cell-Free and Cell-Associated Antibody-Dependent Cellular Phagocytosis (ADCP) Responses in Lung Transplant Recipients. International Journal of Molecular Sciences, 22(15), 8206. https://doi.org/10.3390/ijms22158206