
Role of Insulin in Health and Disease: An Update

 ,
,  ,
,  ,
,  ,
,  ,
,  and
and 
Abstract

1. Introduction
2. Regulation of Insulin Secretion
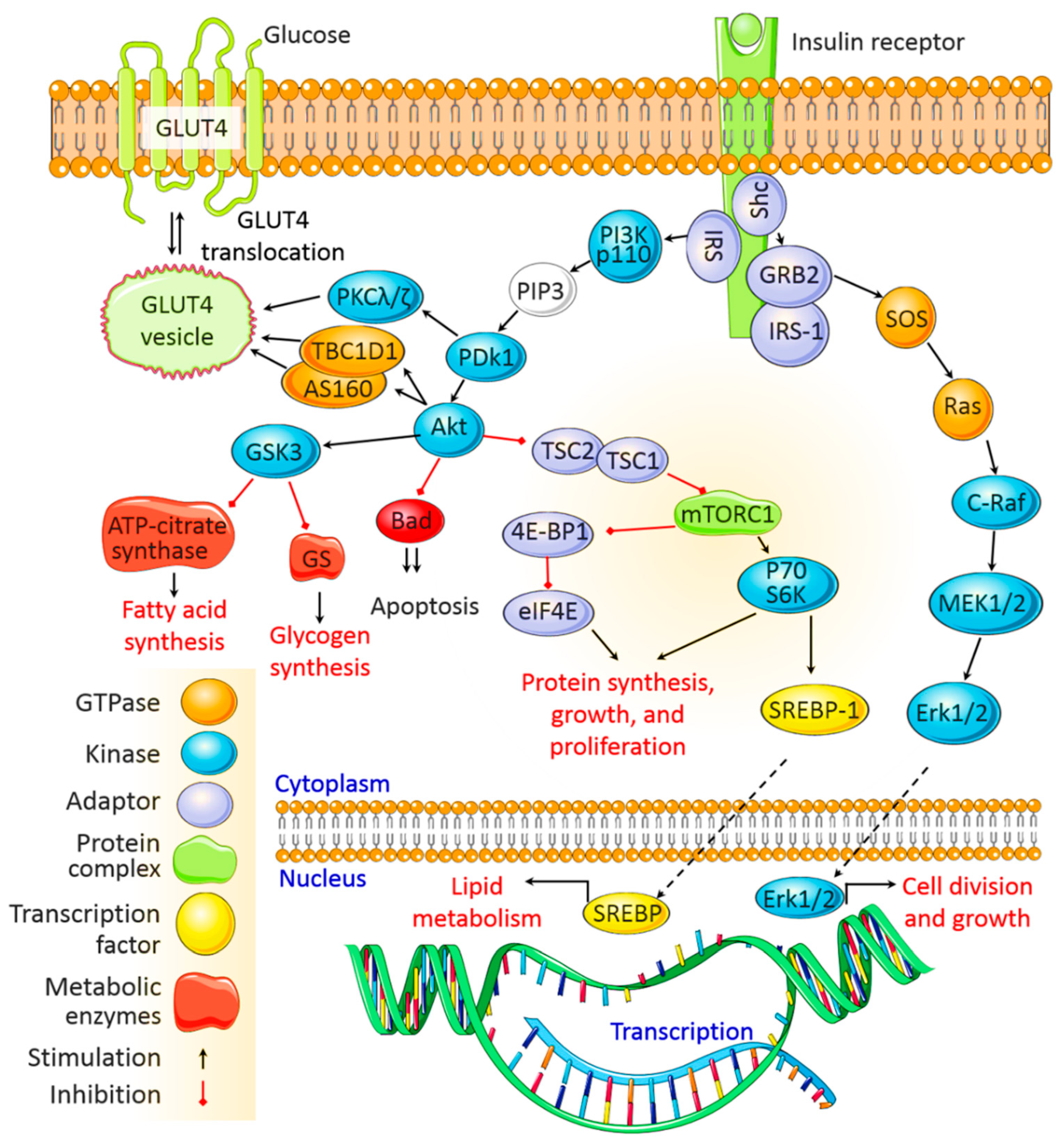
3. Insulin Signaling Pathways
3.1. The PI3K/Akt Signaling Pathway
3.2. The MAPK Signaling Pathway
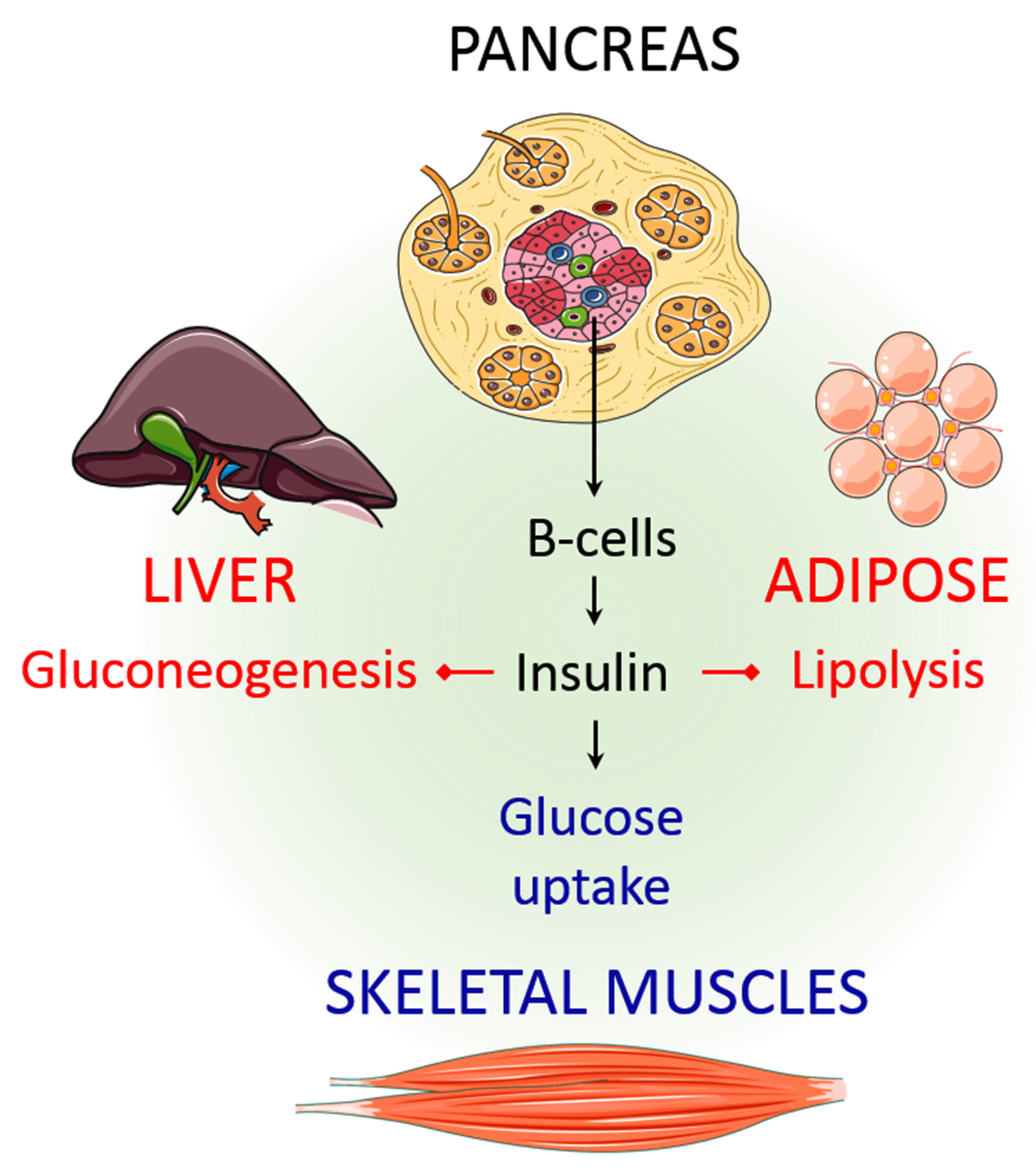
4. Physiological Roles of Insulin
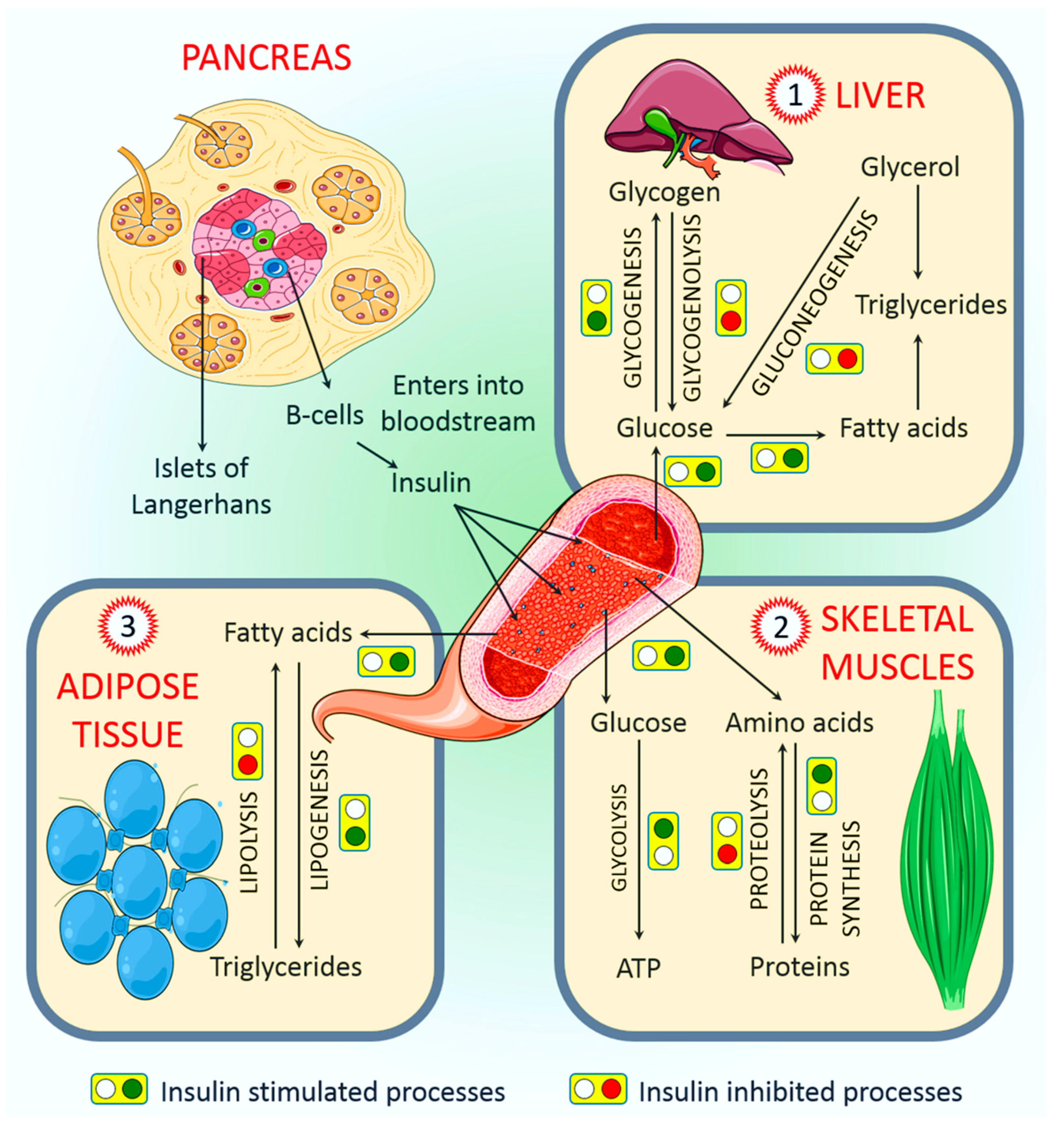
4.1. Role of Insulin in the Regulation of Liver Function
4.2. Role of Insulin in the Regulation of Skeletal Muscle Function
4.3. Role of Insulin in the Regulation of Adipose Tissue Function
4.4. Other Major Physiological Roles
4.4.1. Endothelium and Vasculature
4.4.2. Brain
4.4.3. Kidney
4.4.4. Bone
4.4.5. Skin and Hair Follicles
5. Role of Insulin in Pathology
5.1. Insulin Deficiency
5.2. Hyperinsulinemia
5.3. Hyperglycemia
5.4. Hyperlipidemia
6. Recent Advances in Insulin-Signaling Targeted Therapy
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lewis, G.F.; Brubaker, P.L. The discovery of insulin revisited: Lessons for the modern era. J. Clin. Investig. 2021, 131, e142239. [Google Scholar] [CrossRef] [PubMed]
- Vecchio, I.; Tornali, C.; Bragazzi, N.L.; Martini, M. The Discovery of Insulin: An Important Milestone in the History of Medicine. Front. Endocrinol. 2018, 9, 613. [Google Scholar] [CrossRef] [PubMed]
- Csajbok, E.A.; Tamas, G. Cerebral cortex: A target and source of insulin? Diabetologia 2016, 59, 1609–1615. [Google Scholar] [CrossRef] [PubMed]
- Alarcon, C.; Lincoln, B.; Rhodes, C.J. The biosynthesis of the subtilisin-related proprotein convertase PC3, but no that of the PC2 convertase, is regulated by glucose in parallel to proinsulin biosynthesis in rat pancreatic islets. J. Biol. Chem. 1993, 268, 4276–4280. [Google Scholar] [CrossRef]
- Kaufman, B.A.; Li, C.; Soleimanpour, S.A. Mitochondrial regulation of beta-cell function: Maintaining the momentum for insulin release. Mol. Asp. Med. 2015, 42, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Vasiljevic, J.; Torkko, J.M.; Knoch, K.P.; Solimena, M. The making of insulin in health and disease. Diabetologia 2020, 63, 1981–1989. [Google Scholar] [CrossRef]
- Suckale, J.; Solimena, M. The insulin secretory granule as a signaling hub. Trends Endocrinol. Metab. 2010, 21, 599–609. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.Y.; Zhai, G.; Gong, Y.L.; Su, J.Z.; Peng, X.Y.; Shang, G.H.; Han, D.; Jin, J.Y.; Liu, H.K.; Du, Z.Y.; et al. Different physiological roles of insulin receptors in mediating nutrient metabolism in zebrafish. Am. J. Physiol. Endocrinol. Metab. 2018, 315, E38–E51. [Google Scholar] [CrossRef]
- Avruch, J. Insulin signal transduction through protein kinase cascades. Mol. Cell. Biochem. 1998, 182, 31–48. [Google Scholar] [CrossRef]
- Taniguchi, C.M.; Emanuelli, B.; Kahn, C.R. Critical nodes in signalling pathways: Insights into insulin action. Nat. Rev. Mol. Cell Biol. 2006, 7, 85–96. [Google Scholar] [CrossRef]
- Accili, D. Insulin Action Research and the Future of Diabetes Treatment: The 2017 Banting Medal for Scientific Achievement Lecture. Diabetes 2018, 67, 1701–1709. [Google Scholar] [CrossRef]
- Davidovici, B.B.; Sattar, N.; Prinz, J.; Puig, L.; Emery, P.; Barker, J.N.; van de Kerkhof, P.; Stahle, M.; Nestle, F.O.; Girolomoni, G.; et al. Psoriasis and systemic inflammatory diseases: Potential mechanistic links between skin disease and co-morbid conditions. J. Investig. Dermatol. 2010, 130, 1785–1796. [Google Scholar] [CrossRef]
- Plikus, M.V.; Van Spyk, E.N.; Pham, K.; Geyfman, M.; Kumar, V.; Takahashi, J.S.; Andersen, B. The circadian clock in skin: Implications for adult stem cells, tissue regeneration, cancer, aging, and immunity. J. Biol. Rhythm. 2015, 30, 163–182. [Google Scholar] [CrossRef]
- Baselga Torres, E.; Torres-Pradilla, M. Cutaneous manifestations in children with diabetes mellitus and obesity. Actas Dermosifiliogr. 2014, 105, 546–557. [Google Scholar] [CrossRef] [PubMed]
- Hough, F.S.; Pierroz, D.D.; Cooper, C.; Ferrari, S.L.; Bone, I.C.; Diabetes Working Group. Mechanisms in endocrinology: Mechanisms and evaluation of bone fragility in type 1 diabetes mellitus. Eur. J. Endocrinol. 2016, 174, R127–R138. [Google Scholar] [CrossRef] [PubMed]
- Hirsch, I.B.; Juneja, R.; Beals, J.M.; Antalis, C.J.; Wright, E.E. The Evolution of Insulin and How it Informs Therapy and Treatment Choices. Endocr. Rev. 2020, 41, 733–755. [Google Scholar] [CrossRef]
- Wendt, A.; Eliasson, L. Pancreatic alpha-cells—The unsung heroes in islet function. Semin. Cell Dev. Biol 2020, 103, 41–50. [Google Scholar] [CrossRef]
- Cabrera, O.; Berman, D.M.; Kenyon, N.S.; Ricordi, C.; Berggren, P.O.; Caicedo, A. The unique cytoarchitecture of human pancreatic islets has implications for islet cell function. Proc. Natl. Acad. Sci. USA 2006, 103, 2334–2339. [Google Scholar] [CrossRef] [PubMed]
- Rorsman, P.; Braun, M. Regulation of insulin secretion in human pancreatic islets. Annu. Rev. Physiol. 2013, 75, 155–179. [Google Scholar] [CrossRef]
- Jansson, L.; Hellerstrom, C. Glucose-induced changes in pancreatic islet blood flow mediated by central nervous system. Am. J. Physiol. 1986, 251, E644–E647. [Google Scholar] [CrossRef]
- Fu, Z.; Gilbert, E.R.; Liu, D. Regulation of insulin synthesis and secretion and pancreatic Beta-cell dysfunction in diabetes. Curr. Diabetes Rev. 2013, 9, 25–53. [Google Scholar] [CrossRef]
- De Vos, A.; Heimberg, H.; Quartier, E.; Huypens, P.; Bouwens, L.; Pipeleers, D.; Schuit, F. Human and rat beta cells differ in glucose transporter but not in glucokinase gene expression. J. Clin. Investig. 1995, 96, 2489–2495. [Google Scholar] [CrossRef]
- McCulloch, L.J.; van de Bunt, M.; Braun, M.; Frayn, K.N.; Clark, A.; Gloyn, A.L. GLUT2 (SLC2A2) is not the principal glucose transporter in human pancreatic beta cells: Implications for understanding genetic association signals at this locus. Mol. Genet. Metab. 2011, 104, 648–653. [Google Scholar] [CrossRef] [PubMed]
- Gloyn, A.L.; Odili, S.; Zelent, D.; Buettger, C.; Castleden, H.A.; Steele, A.M.; Stride, A.; Shiota, C.; Magnuson, M.A.; Lorini, R.; et al. Insights into the structure and regulation of glucokinase from a novel mutation (V62M), which causes maturity-onset diabetes of the young. J. Biol. Chem. 2005, 280, 14105–14113. [Google Scholar] [CrossRef]
- Doliba, N.M.; Qin, W.; Najafi, H.; Liu, C.; Buettger, C.W.; Sotiris, J.; Collins, H.W.; Li, C.; Stanley, C.A.; Wilson, D.F.; et al. Glucokinase activation repairs defective bioenergetics of islets of Langerhans isolated from type 2 diabetics. Am. J. Physiol. Endocrinol. Metab. 2012, 302, E87–E102. [Google Scholar] [CrossRef]
- Yajima, H.; Komatsu, M.; Schermerhorn, T.; Aizawa, T.; Kaneko, T.; Nagai, M.; Sharp, G.W.; Hashizume, K. cAMP enhances insulin secretion by an action on the ATP-sensitive K+ channel-independent pathway of glucose signaling in rat pancreatic islets. Diabetes 1999, 48, 1006–1012. [Google Scholar] [CrossRef] [PubMed]
- Yamada, S.; Komatsu, M.; Sato, Y.; Yamauchi, K.; Kojima, I.; Aizawa, T.; Hashizume, K. Time-dependent stimulation of insulin exocytosis by 3’,5’-cyclic adenosine monophosphate in the rat islet beta-cell. Endocrinology 2002, 143, 4203–4209. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Haeusler, R.A.; McGraw, T.E.; Accili, D. Biochemical and cellular properties of insulin receptor signalling. Nat. Rev. Mol. Cell Biol. 2018, 19, 31–44. [Google Scholar] [CrossRef] [PubMed]
- Saltiel, A.R. Insulin signaling in health and disease. J. Clin. Investig. 2021, 131, e142241. [Google Scholar] [CrossRef] [PubMed]
- Burks, D.J.; White, M.F. IRS proteins and beta-cell function. Diabetes 2001, 50 (Suppl. 1), S140–S145. [Google Scholar] [CrossRef]
- Alves-Bezerra, M.; Cohen, D.E. Triglyceride Metabolism in the Liver. Compr. Physiol. 2017, 8, 1–8. [Google Scholar]
- Hatting, M.; Tavares, C.D.J.; Sharabi, K.; Rines, A.K.; Puigserver, P. Insulin regulation of gluconeogenesis. Ann. NY Acad. Sci. 2018, 1411, 21–35. [Google Scholar] [CrossRef]
- Adeyinka, A.; Kondamudi, N.P. Hyperosmolar Hyperglycemic Nonketotic Coma. In StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Gastaldelli, A.; Toschi, E.; Pettiti, M.; Frascerra, S.; Quinones-Galvan, A.; Sironi, A.M.; Natali, A.; Ferrannini, E. Effect of physiological hyperinsulinemia on gluconeogenesis in nondiabetic subjects and in type 2 diabetic patients. Diabetes 2001, 50, 1807–1812. [Google Scholar] [CrossRef]
- Edgerton, D.S.; Lautz, M.; Scott, M.; Everett, C.A.; Stettler, K.M.; Neal, D.W.; Chu, C.A.; Cherrington, A.D. Insulin’s direct effects on the liver dominate the control of hepatic glucose production. J. Clin. Investig. 2006, 116, 521–527. [Google Scholar] [CrossRef] [PubMed]
- Girard, J. Insulin’s effect on the liver: “direct or indirect?” continues to be the question. J. Clin. Investig. 2006, 116, 302–304. [Google Scholar] [CrossRef]
- Sharabi, K.; Tavares, C.D.; Rines, A.K.; Puigserver, P. Molecular pathophysiology of hepatic glucose production. Mol. Asp. Med. 2015, 46, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Claus, T.H.; Pilkis, S.J. Regulation by insulin of gluconeogenesis in isolated rat hepatocytes. Biochim. Biophys. Acta 1976, 421, 246–262. [Google Scholar] [CrossRef]
- Marks, J.S.; Botelho, L.H. Synergistic inhibition of glucagon-induced effects on hepatic glucose metabolism in the presence of insulin and a cAMP antagonist. J. Biol. Chem. 1986, 261, 15895–15899. [Google Scholar] [CrossRef]
- Sindelar, D.K.; Balcom, J.H.; Chu, C.A.; Neal, D.W.; Cherrington, A.D. A comparison of the effects of selective increases in peripheral or portal insulin on hepatic glucose production in the conscious dog. Diabetes 1996, 45, 1594–1604. [Google Scholar] [CrossRef]
- Sindelar, D.K.; Chu, C.A.; Venson, P.; Donahue, E.P.; Neal, D.W.; Cherrington, A.D. Basal hepatic glucose production is regulated by the portal vein insulin concentration. Diabetes 1998, 47, 523–529. [Google Scholar] [CrossRef]
- Ito, K.; Maruyama, H.; Hirose, H.; Kido, K.; Koyama, K.; Kataoka, K.; Saruta, T. Exogenous insulin dose-dependently suppresses glucopenia-induced glucagon secretion from perfused rat pancreas. Metabolism 1995, 44, 358–362. [Google Scholar] [CrossRef]
- Sindelar, D.K.; Chu, C.A.; Rohlie, M.; Neal, D.W.; Swift, L.L.; Cherrington, A.D. The role of fatty acids in mediating the effects of peripheral insulin on hepatic glucose production in the conscious dog. Diabetes 1997, 46, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Obici, S.; Zhang, B.B.; Karkanias, G.; Rossetti, L. Hypothalamic insulin signaling is required for inhibition of glucose production. Nat. Med. 2002, 8, 1376–1382. [Google Scholar] [CrossRef] [PubMed]
- Bergman, R.N. Non-esterified fatty acids and the liver: Why is insulin secreted into the portal vein? Diabetologia 2000, 43, 946–952. [Google Scholar] [CrossRef] [PubMed]
- Frontera, W.R.; Ochala, J. Skeletal muscle: A brief review of structure and function. Calcif. Tissue Int. 2015, 96, 183–195. [Google Scholar] [CrossRef] [PubMed]
- Baron, A.D.; Brechtel, G.; Wallace, P.; Edelman, S.V. Rates and tissue sites of non-insulin- and insulin-mediated glucose uptake in humans. Am. J. Physiol. 1988, 255 Pt 1, E769–E774. [Google Scholar] [CrossRef]
- DeFronzo, R.A. Pathogenesis of type 2 diabetes mellitus. Med. Clin. N. Am. 2004, 88, 787–835. [Google Scholar] [CrossRef]
- DeFronzo, R.A. Lilly lecture 1987. The triumvirate: Beta-cell, muscle, liver. A collusion responsible for NIDDM. Diabetes 1988, 37, 667–687. [Google Scholar] [CrossRef]
- Defronzo, R.A. Banting Lecture. From the triumvirate to the ominous octet: A new paradigm for the treatment of type 2 diabetes mellitus. Diabetes 2009, 58, 773–795. [Google Scholar] [CrossRef]
- Park, S.W.; Goodpaster, B.H.; Lee, J.S.; Kuller, L.H.; Boudreau, R.; de Rekeneire, N.; Harris, T.B.; Kritchevsky, S.; Tylavsky, F.A.; Nevitt, M.; et al. Excessive loss of skeletal muscle mass in older adults with type 2 diabetes. Diabetes Care 2009, 32, 1993–1997. [Google Scholar] [CrossRef]
- Abdulla, H.; Smith, K.; Atherton, P.J.; Idris, I. Role of insulin in the regulation of human skeletal muscle protein synthesis and breakdown: A systematic review and meta-analysis. Diabetologia 2016, 59, 44–55. [Google Scholar] [CrossRef]
- Romijn, J.A.; Coyle, E.F.; Sidossis, L.S.; Gastaldelli, A.; Horowitz, J.F.; Endert, E.; Wolfe, R.R. Regulation of endogenous fat and carbohydrate metabolism in relation to exercise intensity and duration. Am. J. Physiol. 1993, 265 Pt 1, E380–E391. [Google Scholar] [CrossRef]
- Taniguchi, M.; Yoshida, H. Endoplasmic reticulum stress in kidney function and disease. Curr. Opin. Nephrol. Hypertens. 2015, 24, 345–350. [Google Scholar] [CrossRef] [PubMed]
- Cheng, A.; Dube, N.; Gu, F.; Tremblay, M.L. Coordinated action of protein tyrosine phosphatases in insulin signal transduction. Eur. J. Biochem. 2002, 269, 1050–1059. [Google Scholar] [CrossRef] [PubMed]
- Karakelides, H.; Asmann, Y.W.; Bigelow, M.L.; Short, K.R.; Dhatariya, K.; Coenen-Schimke, J.; Kahl, J.; Mukhopadhyay, D.; Nair, K.S. Effect of insulin deprivation on muscle mitochondrial ATP production and gene transcript levels in type 1 diabetic subjects. Diabetes 2007, 56, 2683–2689. [Google Scholar] [CrossRef] [PubMed]
- Birsoy, K.; Festuccia, W.T.; Laplante, M. A comparative perspective on lipid storage in animals. J. Cell Sci. 2013, 126 Pt 7, 1541–1552. [Google Scholar] [CrossRef]
- Gesta, S.; Tseng, Y.H.; Kahn, C.R. Developmental origin of fat: Tracking obesity to its source. Cell 2007, 131, 242–256. [Google Scholar] [CrossRef]
- Smith, U. Impaired (‘diabetic’) insulin signaling and action occur in fat cells long before glucose intolerance--is insulin resistance initiated in the adipose tissue? Int. J. Obes. Relat. Metab. Disord. 2002, 26, 897–904. [Google Scholar] [CrossRef] [PubMed]
- Cignarelli, A.; Genchi, V.A.; Perrini, S.; Natalicchio, A.; Laviola, L.; Giorgino, F. Insulin and Insulin Receptors in Adipose Tissue Development. Int. J. Mol. Sci. 2019, 20, 759. [Google Scholar] [CrossRef]
- Koyama, K.; Chen, G.; Lee, Y.; Unger, R.H. Tissue triglycerides, insulin resistance, and insulin production: Implications for hyperinsulinemia of obesity. Am. J. Physiol. 1997, 273, E708–E713. [Google Scholar] [CrossRef]
- Favero, G.; Paganelli, C.; Buffoli, B.; Rodella, L.F.; Rezzani, R. Endothelium and its alterations in cardiovascular diseases: Life style intervention. BioMed Res. Int. 2014, 2014, 801896. [Google Scholar] [CrossRef]
- Barac, A.; Campia, U.; Panza, J.A. Methods for evaluating endothelial function in humans. Hypertension 2007, 49, 748–760. [Google Scholar] [CrossRef] [PubMed]
- Muniyappa, R.; Quon, M.J. Insulin action and insulin resistance in vascular endothelium. Curr. Opin. Clin. Nutr. Metab. Care 2007, 10, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Vicent, D.; Ilany, J.; Kondo, T.; Naruse, K.; Fisher, S.J.; Kisanuki, Y.Y.; Bursell, S.; Yanagisawa, M.; King, G.L.; Kahn, C.R. The role of endothelial insulin signaling in the regulation of vascular tone and insulin resistance. J. Clin. Investig. 2003, 111, 1373–1380. [Google Scholar] [CrossRef] [PubMed]
- Potenza, M.A.; Addabbo, F.; Montagnani, M. Vascular actions of insulin with implications for endothelial dysfunction. Am. J. Physiol. Endocrinol. Metab. 2009, 297, E568–E577. [Google Scholar] [CrossRef]
- Hom, F.G.; Goodner, C.J.; Berrie, M.A. A [3H]2-deoxyglucose method for comparing rates of glucose metabolism and insulin responses among rat tissues in vivo. Validation of the model and the absence of an insulin effect on brain. Diabetes 1984, 33, 141–152. [Google Scholar] [CrossRef]
- Hasselbalch, S.G.; Knudsen, G.M.; Videbaek, C.; Pinborg, L.H.; Schmidt, J.F.; Holm, S.; Paulson, O.B. No effect of insulin on glucose blood-brain barrier transport and cerebral metabolism in humans. Diabetes 1999, 48, 1915–1921. [Google Scholar] [CrossRef]
- Havrankova, J.; Schmechel, D.; Roth, J.; Brownstein, M. Identification of insulin in rat brain. Proc. Natl. Acad. Sci. USA 1978, 75, 5737–5741. [Google Scholar] [CrossRef]
- Ramnanan, C.J.; Edgerton, D.S.; Cherrington, A.D. Evidence against a physiologic role for acute changes in CNS insulin action in the rapid regulation of hepatic glucose production. Cell Metab. 2012, 15, 656–664. [Google Scholar] [CrossRef]
- Ramnanan, C.J.; Kraft, G.; Smith, M.S.; Farmer, B.; Neal, D.; Williams, P.E.; Lautz, M.; Farmer, T.; Donahue, E.P.; Cherrington, A.D.; et al. Interaction between the central and peripheral effects of insulin in controlling hepatic glucose metabolism in the conscious dog. Diabetes 2013, 62, 74–84. [Google Scholar] [CrossRef]
- Benedict, C.; Hallschmid, M.; Schultes, B.; Born, J.; Kern, W. Intranasal insulin to improve memory function in humans. Neuroendocrinology 2007, 86, 136–142. [Google Scholar] [CrossRef]
- Craft, S.; Baker, L.D.; Montine, T.J.; Minoshima, S.; Watson, G.S.; Claxton, A.; Arbuckle, M.; Callaghan, M.; Tsai, E.; Plymate, S.R.; et al. Intranasal insulin therapy for Alzheimer disease and amnestic mild cognitive impairment: A pilot clinical trial. Arch. Neurol. 2012, 69, 29–38. [Google Scholar] [CrossRef]
- Taouis, M.; Torres-Aleman, I. Editorial: Insulin and The Brain. Front. Endocrinol. 2019, 10, 299. [Google Scholar] [CrossRef] [PubMed]
- Benedict, C.; Kern, W.; Schultes, B.; Born, J.; Hallschmid, M. Differential sensitivity of men and women to anorexigenic and memory-improving effects of intranasal insulin. J. Clin. Endocrinol. Metab. 2008, 93, 1339–1344. [Google Scholar] [CrossRef] [PubMed]
- Kern, W.; Benedict, C.; Schultes, B.; Plohr, F.; Moser, A.; Born, J.; Fehm, H.L.; Hallschmid, M. Low cerebrospinal fluid insulin levels in obese humans. Diabetologia 2006, 49, 2790–2792. [Google Scholar] [CrossRef] [PubMed]
- Whitmer, R.A.; Gunderson, E.P.; Barrett-Connor, E.; Quesenberry, C.P., Jr.; Yaffe, K. Obesity in middle age and future risk of dementia: A 27 year longitudinal population based study. BMJ 2005, 330, 1360. [Google Scholar] [CrossRef] [PubMed]
- Benedict, C.; Hallschmid, M.; Hatke, A.; Schultes, B.; Fehm, H.L.; Born, J.; Kern, W. Intranasal insulin improves memory in humans. Psychoneuroendocrinology 2004, 29, 1326–1334. [Google Scholar] [CrossRef]
- Brunner, Y.F.; Kofoet, A.; Benedict, C.; Freiherr, J. Central insulin administration improves odor-cued reactivation of spatial memory in young men. J. Clin. Endocrinol. Metab. 2015, 100, 212–219. [Google Scholar] [CrossRef]
- Reger, M.A.; Watson, G.S.; Frey, W.H., 2nd; Baker, L.D.; Cholerton, B.; Keeling, M.L.; Belongia, D.A.; Fishel, M.A.; Plymate, S.R.; Schellenberg, G.D.; et al. Effects of intranasal insulin on cognition in memory-impaired older adults: Modulation by APOE genotype. Neurobiol. Aging 2006, 27, 451–458. [Google Scholar] [CrossRef]
- Reger, M.A.; Watson, G.S.; Green, P.S.; Wilkinson, C.W.; Baker, L.D.; Cholerton, B.; Fishel, M.A.; Plymate, S.R.; Breitner, J.C.; DeGroodt, W.; et al. Intranasal insulin improves cognition and modulates beta-amyloid in early AD. Neurology 2008, 70, 440–448. [Google Scholar] [CrossRef]
- Hallschmid, M.; Benedict, C.; Born, J.; Fehm, H.L.; Kern, W. Manipulating central nervous mechanisms of food intake and body weight regulation by intranasal administration of neuropeptides in man. Physiol. Behav. 2004, 83, 55–64. [Google Scholar] [CrossRef]
- Farber, S.J.; Berger, E.Y.; Earle, D.P. Effect of diabetes and insulin of the maximum capacity of the renal tubules to reabsorb glucose. J. Clin. Investig. 1951, 30, 125–129. [Google Scholar] [CrossRef]
- Singh, S.; Sharma, R.; Kumari, M.; Tiwari, S. Insulin receptors in the kidneys in health and disease. World J. Nephrol. 2019, 8, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Gatica, R.; Bertinat, R.; Silva, P.; Carpio, D.; Ramirez, M.J.; Slebe, J.C.; San Martin, R.; Nualart, F.; Campistol, J.M.; Caelles, C.; et al. Altered expression and localization of insulin receptor in proximal tubule cells from human and rat diabetic kidney. J. Cell. Biochem. 2013, 114, 639–649. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, S.; Halagappa, V.K.; Riazi, S.; Hu, X.; Ecelbarger, C.A. Reduced expression of insulin receptors in the kidneys of insulin-resistant rats. J. Am. Soc. Nephrol. 2007, 18, 2661–2671. [Google Scholar] [CrossRef] [PubMed]
- Winzell, M.S.; Ahren, B. The high-fat diet-fed mouse: A model for studying mechanisms and treatment of impaired glucose tolerance and type 2 diabetes. Diabetes 2004, 53 (Suppl. 3), S215–S219. [Google Scholar] [CrossRef]
- Underwood, P.C.; Adler, G.K. The renin angiotensin aldosterone system and insulin resistance in humans. Curr. Hypertens. Rep. 2013, 15, 59–70. [Google Scholar] [CrossRef]
- Csibi, A.; Communi, D.; Muller, N.; Bottari, S.P. Angiotensin II inhibits insulin-stimulated GLUT4 translocation and Akt activation through tyrosine nitration-dependent mechanisms. PLoS ONE 2010, 5, e10070. [Google Scholar] [CrossRef]
- Akhtar, M.; Taha, N.M.; Nauman, A.; Mujeeb, I.B.; Al-Nabet, A. Diabetic Kidney Disease: Past and Present. Adv. Anat. Pathol. 2020, 27, 87–97. [Google Scholar] [CrossRef]
- Malekzadeh, B.O.; Erlandsson, M.C.; Tengvall, P.; Palmquist, A.; Ransjo, M.; Bokarewa, M.I.; Westerlund, A. Effects of implant-delivered insulin on bone formation in osteoporotic rats. J. Biomed. Mater. Res. A 2018, 106, 2472–2480. [Google Scholar] [CrossRef]
- Fulzele, K.; Riddle, R.C.; DiGirolamo, D.J.; Cao, X.; Wan, C.; Chen, D.; Faugere, M.C.; Aja, S.; Hussain, M.A.; Bruning, J.C.; et al. Insulin receptor signaling in osteoblasts regulates postnatal bone acquisition and body composition. Cell 2010, 142, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Thrailkill, K.M.; Lumpkin, C.K., Jr.; Bunn, R.C.; Kemp, S.F.; Fowlkes, J.L. Is insulin an anabolic agent in bone? Dissecting the diabetic bone for clues. Am. J. Physiol. Endocrinol. Metab. 2005, 289, E735–E745. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Woo, W.M.; Nagao, K.; Li, W.; Terunuma, A.; Mukouyama, Y.S.; Oro, A.E.; Vogel, J.C.; Brownell, I. Perivascular hair follicle stem cells associate with a venule annulus. J. Investig. Dermatol. 2013, 133, 2324–2331. [Google Scholar] [CrossRef] [PubMed]
- Correll, C.U.; Robinson, D.G.; Schooler, N.R.; Brunette, M.F.; Mueser, K.T.; Rosenheck, R.A.; Marcy, P.; Addington, J.; Estroff, S.E.; Robinson, J.; et al. Cardiometabolic risk in patients with first-episode schizophrenia spectrum disorders: Baseline results from the RAISE-ETP study. JAMA Psychiatry 2014, 71, 1350–1363. [Google Scholar] [CrossRef] [PubMed]
- Pierard, G.E.; Seite, S.; Hermanns-Le, T.; Delvenne, P.; Scheen, A.; Pierard-Franchimont, C. The skin landscape in diabetes mellitus. Focus on dermocosmetic management. Clin. Cosmet. Investig. Dermatol. 2013, 6, 127–135. [Google Scholar] [CrossRef]
- Napolitano, M.; Megna, M.; Monfrecola, G. Insulin resistance and skin diseases. Sci. World J. 2015, 2015, 479354. [Google Scholar] [CrossRef]
- Kahana, M.; Grossman, E.; Feinstein, A.; Ronnen, M.; Cohen, M.; Millet, M.S. Skin tags: A cutaneous marker for diabetes mellitus. Acta Derm. Venereol. 1987, 67, 175–177. [Google Scholar]
- Cordain, L.; Lindeberg, S.; Hurtado, M.; Hill, K.; Eaton, S.B.; Brand-Miller, J. Acne vulgaris: A disease of Western civilization. Arch. Dermatol. 2002, 138, 1584–1590. [Google Scholar] [CrossRef]
- Lindeberg, S.; Eliasson, M.; Lindahl, B.; Ahren, B. Low serum insulin in traditional Pacific Islanders--the Kitava Study. Metabolism 1999, 48, 1216–1219. [Google Scholar] [CrossRef]
- Melnik, B.C.; John, S.M.; Schmitz, G. Over-stimulation of insulin/IGF-1 signaling by western diet may promote diseases of civilization: Lessons learnt from laron syndrome. Nutr. Metab. 2011, 8, 41. [Google Scholar] [CrossRef]
- Toussirot, E.; Aubin, F.; Dumoulin, G. Relationships between Adipose Tissue and Psoriasis, with or without Arthritis. Front. Immunol. 2014, 5, 368. [Google Scholar] [CrossRef] [PubMed]
- Yadav, A.; Kataria, M.A.; Saini, V.; Yadav, A. Role of leptin and adiponectin in insulin resistance. Clin. Chim. Acta 2013, 417, 80–84. [Google Scholar] [CrossRef] [PubMed]
- Abdel Hay, R.M.; Rashed, L.A. Association between the leptin gene 2548G/A polymorphism, the plasma leptin and the metabolic syndrome with psoriasis. Exp. Dermatol. 2011, 20, 715–719. [Google Scholar] [CrossRef] [PubMed]
- Coimbra, S.; Oliveira, H.; Reis, F.; Belo, L.; Rocha, S.; Quintanilha, A.; Figueiredo, A.; Teixeira, F.; Castro, E.; Rocha-Pereira, P.; et al. Circulating adipokine levels in Portuguese patients with psoriasis vulgaris according to body mass index, severity and therapy. J. Eur. Acad. Dermatol. Venereol. 2010, 24, 1386–1394. [Google Scholar] [CrossRef]
- Ruegsegger, G.N.; Creo, A.L.; Cortes, T.M.; Dasari, S.; Nair, K.S. Altered mitochondrial function in insulin-deficient and insulin-resistant states. J. Clin. Investig. 2018, 128, 3671–3681. [Google Scholar] [CrossRef]
- Thomas, D.D.; Corkey, B.E.; Istfan, N.W.; Apovian, C.M. Hyperinsulinemia: An Early Indicator of Metabolic Dysfunction. J. Endocr. Soc. 2019, 3, 1727–1747. [Google Scholar] [CrossRef]
- Bazotte, R.B.; Silva, L.G.; Schiavon, F.P. Insulin resistance in the liver: Deficiency or excess of insulin? Cell Cycle 2014, 13, 2494–2500. [Google Scholar] [CrossRef]
- Santoleri, D.; Titchenell, P.M. Resolving the Paradox of Hepatic Insulin Resistance. Cell. Mol. Gastroenterol. Hepatol. 2019, 7, 447–456. [Google Scholar] [CrossRef]
- Titchenell, P.M.; Quinn, W.J.; Lu, M.; Chu, Q.; Lu, W.; Li, C.; Chen, H.; Monks, B.R.; Chen, J.; Rabinowitz, J.D.; et al. Direct Hepatocyte Insulin Signaling Is Required for Lipogenesis but Is Dispensable for the Suppression of Glucose Production. Cell Metab. 2016, 23, 1154–1166. [Google Scholar] [CrossRef]
- Ferrannini, E. Insulin resistance versus insulin deficiency in non-insulin-dependent diabetes mellitus: Problems and prospects. Endocr. Rev. 1998, 19, 477–490. [Google Scholar] [CrossRef]
- Woo, V.C. New Insulins and New Aspects in Insulin Delivery. Can. J. Diabetes 2015, 39, 335–343. [Google Scholar] [CrossRef]
- Majeed, W.; Thabit, H. Closed-loop insulin delivery: Current status of diabetes technologies and future prospects. Expert Rev. Med. Devices 2018, 15, 579–590. [Google Scholar] [CrossRef]
- Ramzy, A.; Mojibian, M.; Kieffer, T.J. Insulin-Deficient Mouse beta-Cells Do Not Fully Mature but Can Be Remedied Through Insulin Replacement by Islet Transplantation. Endocrinology 2018, 159, 83–102. [Google Scholar] [CrossRef] [PubMed]
- Maneschi, F.; Mashiter, K.; Kohner, E.M. Insulin resistance and insulin deficiency in diabetic retinopathy of non-insulin-dependent diabetes. Diabetes 1983, 32, 82–87. [Google Scholar] [CrossRef] [PubMed]
- Cerasi, E. Insulin deficiency and insulin resistance in the pathogenesis of NIDDM: Is a divorce possible? Diabetologia 1995, 38, 992–997. [Google Scholar] [CrossRef] [PubMed]
- Groop, L.C.; Widen, E.; Ferrannini, E. Insulin resistance and insulin deficiency in the pathogenesis of type 2 (non-insulin-dependent) diabetes mellitus: Errors of metabolism or of methods? Diabetologia 1993, 36, 1326–1331. [Google Scholar] [CrossRef]
- Kuzuya, T.; Matsuda, A. Classification of diabetes on the basis of etiologies versus degree of insulin deficiency. Diabetes Care 1997, 20, 219–220. [Google Scholar] [CrossRef]
- Moghetti, P.; Tosi, F. Insulin resistance and PCOS: Chicken or egg? J. Endocrinol. Investig. 2021, 44, 233–244. [Google Scholar] [CrossRef]
- Thomas, N.J.; Lynam, A.L.; Hill, A.V.; Weedon, M.N.; Shields, B.M.; Oram, R.A.; McDonald, T.J.; Hattersley, A.T.; Jones, A.G. Type 1 diabetes defined by severe insulin deficiency occurs after 30 years of age and is commonly treated as type 2 diabetes. Diabetologia 2019, 62, 1167–1172. [Google Scholar] [CrossRef]
- Kopp, W. How Western Diet and Lifestyle Drive the Pandemic of Obesity and Civilization Diseases. Diabetes Metab. Syndr. Obes. 2019, 12, 2221–2236. [Google Scholar] [CrossRef] [PubMed]
- Jezek, P.; Jaburek, M.; Holendova, B.; Plecita-Hlavata, L. Fatty Acid-Stimulated Insulin Secretion vs. Lipotoxicity. Molecules 2018, 23. [Google Scholar]
- Jia, G.; Whaley-Connell, A.; Sowers, J.R. Diabetic cardiomyopathy: A hyperglycaemia- and insulin-resistance-induced heart disease. Diabetologia 2018, 61, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Otto-Buczkowska, E.; Grzyb, K.; Jainta, N. Polycystic ovary syndrome (PCOS) and the accompanying disorders of glucose homeostasis among girls at the time of puberty. Pediatr. Endocrinol. Diabetes Metab. 2018, 24, 40–44. [Google Scholar] [CrossRef] [PubMed]
- Gerich, J.E. Control of glycaemia. Baillieres Clin. Endocrinol. Metab. 1993, 7, 551–586. [Google Scholar] [CrossRef]
- Villegas-Valverde, C.C.; Kokuina, E.; Breff-Fonseca, M.C. Strengthening National Health Priorities for Diabetes Prevention and Management. MEDICC Rev. 2018, 20, 5. [Google Scholar]
- Hammer, M.; Storey, S.; Hershey, D.S.; Brady, V.J.; Davis, E.; Mandolfo, N.; Bryant, A.L.; Olausson, J. Hyperglycemia and Cancer: A State-of-the-Science Review. Oncol. Nurs. Forum 2019, 46, 459–472. [Google Scholar]
- Giugliano, D.; Ceriello, A.; Esposito, K. Glucose metabolism and hyperglycemia. Am. J. Clin. Nutr. 2008, 87, 217S–222S. [Google Scholar] [CrossRef]
- Rawlings, A.M.; Sharrett, A.R.; Albert, M.S.; Coresh, J.; Windham, B.G.; Power, M.C.; Knopman, D.S.; Walker, K.; Burgard, S.; Mosley, T.H.; et al. The Association of Late-Life Diabetes Status and Hyperglycemia with Incident Mild Cognitive Impairment and Dementia: The ARIC Study. Diabetes Care 2019, 42, 1248–1254. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, J.J.; Black, M.H.; Li, B.H.; Reynolds, K.; Lawrence, J.M. Race/ethnicity and measures of glycaemia in the year after diagnosis among youth with type 1 and type 2 diabetes mellitus. J. Diabetes Complicat. 2014, 28, 279–285. [Google Scholar] [CrossRef]
- Yari, Z.; Behrouz, V.; Zand, H.; Pourvali, K. New Insight into Diabetes Management: From Glycemic Index to Dietary Insulin Index. Curr. Diabetes Rev. 2020, 16, 293–300. [Google Scholar] [CrossRef]
- Simon, K.; Wittmann, I. Can blood glucose value really be referred to as a metabolic parameter? Rev. Endocr. Metab. Disord. 2019, 20, 151–160. [Google Scholar] [CrossRef]
- Garber, A.J.; Cryer, P.E.; Santiago, J.V.; Haymond, M.W.; Pagliara, A.S.; Kipnis, D.M. The role of adrenergic mechanisms in the substrate and hormonal response to insulin-induced hypoglycemia in man. J. Clin. Investig. 1976, 58, 7–15. [Google Scholar] [CrossRef]
- Coutinho, M.; Gerstein, H.C.; Wang, Y.; Yusuf, S. The relationship between glucose and incident cardiovascular events. A metaregression analysis of published data from 20 studies of 95,783 individuals followed for 12.4 years. Diabetes Care 1999, 22, 233–240. [Google Scholar] [CrossRef]
- Decode Study Group; European Diabetes Epidemiology Group. Is the current definition for diabetes relevant to mortality risk from all causes and cardiovascular and noncardiovascular diseases? Diabetes Care 2003, 26, 688–696. [Google Scholar] [CrossRef]
- Gosmanov, A.R.; Gosmanova, E.O.; Kitabchi, A.E. Hyperglycemic Crises: Diabetic Ketoacidosis and Hyperglycemic Hyperosmolar State. In Endotext; Feingold, K.R., Anawalt, B., Boyce, A., Chrousos, G., de Herder, W.W., Dhatariya, K., Dungan, K., Grossman, A., Hershman, J.M., Hofland, J., et al., Eds.; MDText.com, Inc.: South Dartmouth, MA, USA, 2000. [Google Scholar]
- Rahman, M.S.; Adegoke, E.O.; Pang, M.G. Drivers of owning more BPA. J. Hazard. Mater. 2021, 417, 126076. [Google Scholar] [CrossRef] [PubMed]
- Esposito, K.; Giugliano, D. Diet and inflammation: A link to metabolic and cardiovascular diseases. Eur. Heart J. 2006, 27, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Gorska, E.; Popko, K.; Stelmaszczyk-Emmel, A.; Ciepiela, O.; Kucharska, A.; Wasik, M. Leptin receptors. Eur. J. Med. Res. 2010, 15 (Suppl. 2), 50–54. [Google Scholar] [CrossRef]
- Marroqui, L.; Gonzalez, A.; Neco, P.; Caballero-Garrido, E.; Vieira, E.; Ripoll, C.; Nadal, A.; Quesada, I. Role of leptin in the pancreatic beta-cell: Effects and signaling pathways. J. Mol. Endocrinol. 2012, 49, R9–R17. [Google Scholar] [CrossRef] [PubMed]
- Lumeng, C.N.; Saltiel, A.R. Inflammatory links between obesity and metabolic disease. J. Clin. Investig. 2011, 121, 2111–2117. [Google Scholar] [CrossRef]
- Khodabandehloo, H.; Gorgani-Firuzjaee, S.; Panahi, G.; Meshkani, R. Molecular and cellular mechanisms linking inflammation to insulin resistance and beta-cell dysfunction. Transl. Res. 2016, 167, 228–256. [Google Scholar] [CrossRef]
- Boni-Schnetzler, M.; Boller, S.; Debray, S.; Bouzakri, K.; Meier, D.T.; Prazak, R.; Kerr-Conte, J.; Pattou, F.; Ehses, J.A.; Schuit, F.C.; et al. Free fatty acids induce a proinflammatory response in islets via the abundantly expressed interleukin-1 receptor I. Endocrinology 2009, 150, 5218–5229. [Google Scholar] [CrossRef]
- Tang, C.; Naassan, A.E.; Chamson-Reig, A.; Koulajian, K.; Goh, T.T.; Yoon, F.; Oprescu, A.I.; Ghanim, H.; Lewis, G.F.; Dandona, P.; et al. Susceptibility to fatty acid-induced beta-cell dysfunction is enhanced in prediabetic diabetes-prone biobreeding rats: A potential link between beta-cell lipotoxicity and islet inflammation. Endocrinology 2013, 154, 89–101. [Google Scholar] [CrossRef]
- McGarry, J.D.; Dobbins, R.L. Fatty acids, lipotoxicity and insulin secretion. Diabetologia 1999, 42, 128–138. [Google Scholar] [CrossRef] [PubMed]
- Boden, G.; Shulman, G.I. Free fatty acids in obesity and type 2 diabetes: Defining their role in the development of insulin resistance and beta-cell dysfunction. Eur. J. Clin. Investig. 2002, 32 (Suppl. 3), 14–23. [Google Scholar] [CrossRef] [PubMed]
- Shimabukuro, M.; Wang, M.Y.; Zhou, Y.T.; Newgard, C.B.; Unger, R.H. Protection against lipoapoptosis of beta cells through leptin-dependent maintenance of Bcl-2 expression. Proc. Natl. Acad. Sci. USA 1998, 95, 9558–9561. [Google Scholar] [CrossRef]
- Gravena, C.; Mathias, P.C.; Ashcroft, S.J. Acute effects of fatty acids on insulin secretion from rat and human islets of Langerhans. J. Endocrinol. 2002, 173, 73–80. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Yamashita, T.; Eto, K.; Okazaki, Y.; Yamashita, S.; Yamauchi, T.; Sekine, N.; Nagai, R.; Noda, M.; Kadowaki, T. Role of uncoupling protein-2 up-regulation and triglyceride accumulation in impaired glucose-stimulated insulin secretion in a beta-cell lipotoxicity model overexpressing sterol regulatory element-binding protein-1c. Endocrinology 2004, 145, 3566–3577. [Google Scholar] [CrossRef]
- Joseph, J.W.; Koshkin, V.; Saleh, M.C.; Sivitz, W.I.; Zhang, C.Y.; Lowell, B.B.; Chan, C.B.; Wheeler, M.B. Free fatty acid-induced beta-cell defects are dependent on uncoupling protein 2 expression. J. Biol. Chem. 2004, 279, 51049–51056. [Google Scholar] [CrossRef]
- Lupi, R.; Dotta, F.; Marselli, L.; Del Guerra, S.; Masini, M.; Santangelo, C.; Patane, G.; Boggi, U.; Piro, S.; Anello, M.; et al. Prolonged exposure to free fatty acids has cytostatic and pro-apoptotic effects on human pancreatic islets: Evidence that beta-cell death is caspase mediated, partially dependent on ceramide pathway, and Bcl-2 regulated. Diabetes 2002, 51, 1437–1442. [Google Scholar] [CrossRef] [PubMed]
- Poitout, V.; Robertson, R.P. Glucolipotoxicity: Fuel excess and beta-cell dysfunction. Endocr. Rev. 2008, 29, 351–366. [Google Scholar] [CrossRef]
- Pan, Q.; Lu, X.; Zhao, C.; Liao, S.; Chen, X.; Guo, F.; Yang, C.; Liu, H.F. Metformin: The updated protective property in kidney disease. Aging 2020, 12, 8742–8759. [Google Scholar] [CrossRef]
- Zhou, T.; Xu, X.; Du, M.; Zhao, T.; Wang, J. A preclinical overview of metformin for the treatment of type 2 diabetes. Biomed. Pharmacother. 2018, 106, 1227–1235. [Google Scholar] [CrossRef]
- Nathan, D.M.; Buse, J.B.; Davidson, M.B.; Ferrannini, E.; Holman, R.R.; Sherwin, R.; Zinman, B. Medical management of hyperglycemia in type 2 diabetes: A consensus algorithm for the initiation and adjustment of therapy: A consensus statement of the American Diabetes Association and the European Association for the Study of Diabetes. Diabetes Care 2009, 32, 193–203. [Google Scholar] [CrossRef]
- Whalen, K.; Miller, S.; Onge, E.S. The Role of Sodium-Glucose Co-Transporter 2 Inhibitors in the Treatment of Type 2 Diabetes. Clin. Ther. 2015, 37, 1150–1166. [Google Scholar] [CrossRef]
- Soccio, R.E.; Chen, E.R.; Lazar, M.A. Thiazolidinediones and the Promise of Insulin Sensitization in Type 2 Diabetes. Cell Metab. 2014, 20, 573–591. [Google Scholar] [CrossRef] [PubMed]
- Usman, B.; Sharma, N.; Satija, S.; Mehta, M.; Vyas, M.; Khatik, G.L.; Khurana, N.; Hansbro, P.M.; Williams, K.; Dua, K. Recent Developments in Alpha-Glucosidase Inhibitors for Management of Type-2 Diabetes: An Update. Curr. Pharm. Des. 2019, 25, 2510–2525. [Google Scholar] [CrossRef] [PubMed]
- Den Hartogh, D.J.; Tsiani, E. Health Benefits of Resveratrol in Kidney Disease: Evidence from In Vitro and In Vivo Studies. Nutrients 2019, 11, 1624. [Google Scholar] [CrossRef] [PubMed]
- Ji, H.; Wu, L.; Ma, X.; Ma, X.; Qin, G. The effect of resveratrol on the expression of AdipoR1 in kidneys of diabetic nephropathy. Mol. Biol. Rep. 2014, 41, 2151–2159. [Google Scholar] [CrossRef]
- Yang, S.C.; Hsu, C.Y.; Chou, W.L.; Fang, J.Y.; Chuang, S.Y. Bioactive Agent Discovery from the Natural Compounds for the Treatment of Type 2 Diabetes Rat Model. Molecules 2020, 25, 5713. [Google Scholar] [CrossRef]
- Garud, M.S.; Kulkarni, Y.A. Gallic acid attenuates type I diabetic nephropathy in rats. Chem. Biol. Interact. 2018, 282, 69–76. [Google Scholar] [CrossRef]
- Du, S.Y.; Liu, H.R.; Lei, T.T.; Xie, X.F.; Wang, H.L.; He, X.; Tong, R.S.; Wang, Y. Mangiferin: An effective therapeutic agent against several disorders. Mol. Med. Rep. 2018, 18, 4775–4786. [Google Scholar] [CrossRef]
- Yen, F.S.; Wei, J.C.; Lin, M.C.; Hsu, C.C.; Hwu, C.M. Long-term outcomes of adding alpha-glucosidase inhibitors in insulin-treated patients with type 2 diabetes. BMC Endocr. Disord. 2021, 21, 25. [Google Scholar] [CrossRef] [PubMed]
- Kanaujia, A.; Duggar, R.; Pannakal, S.T.; Yadav, S.S.; Katiyar, C.K.; Bansal, V.; Anand, S.; Sujatha, S.; Lakshmi, B.S. Insulinomimetic activity of two new gallotannins from the fruits of Capparis moonii. Bioorg. Med. Chem. 2010, 18, 3940–3945. [Google Scholar] [CrossRef]
- Jose, T.; Inzucchi, S.E. Cardiovascular effects of the DPP-4 inhibitors. Diabetes Vasc. Dis. Res. 2012, 9, 109–116. [Google Scholar] [CrossRef] [PubMed]
- El-Shafey, E.S.; Elsherbiny, E.S. The role of apoptosis and autophagy in the insulin-enhancing activity of oxovanadium(IV) bipyridine complex in streptozotocin-induced diabetic mice. Biometals 2020, 33, 123–135. [Google Scholar] [CrossRef] [PubMed]
- Rasouli, H.; Yarani, R.; Pociot, F.; Popovic-Djordjevic, J. Anti-diabetic potential of plant alkaloids: Revisiting current findings and future perspectives. Pharmacol. Res. 2020, 155, 104723. [Google Scholar] [CrossRef]
- Kumar, Y.; Thakur, A.K.; Goyal, R.K. Evaluation of Alpha Glucosidase Inhibitor (Miglitol) for its Efficacy in Constipation Associated with Diabetes. Indian J. Pharm. Educ. 2019, 53, S387–S391. [Google Scholar] [CrossRef]
- Feingold, K.R. Oral and Injectable (Non-Insulin) Pharmacological Agents for Type 2 Diabetes. In Endotext; Feingold, K.R., Anawalt, B., Boyce, A., Chrousos, G., de Herder, W.W., Dhatariya, K., Dungan, K., Grossman, A., Hershman, J.M., Hofland, J., et al., Eds.; MDText.com, Inc.: South Dartmouth, MA, USA, 2000. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
| Categories | Drug | Doses | Experimental Model (Human/Animal) | Disease Model | Mode of Action | References |
|---|---|---|---|---|---|---|
| Natural products | Resveratrol | 20 µM | Human, mouse | Kidney mesangial cells | Induced AdipoR1 mRNA and protein levels Improved FOXO1 activity | Ji et al. [160] |
| Gallic acid | 20–40 mg | Rat | Type I diabetic nephropathy | Decreased TGF-β1 levels and creatinine clearance | Garud, and Kulkarni, [162] | |
| Mangiferin | 40 mg | Human | Type 2 diabetes | Reduced blood glucose levels, AST, ALT, and ALP Activated β-catenin | Du et al. [163] | |
| Saxagliptin | 2.5–5 mg | Human, mouse | Type 2 diabetes | Decreased glucagon production Increased insulin production Inhibited DPP-4 activation | Rasouli et al. [168] | |
| Alpha-glucosidase inhibitors | 25 mg | Type 2 diabetes | Reduced renal function Produced hypoglycemic effect | Kumar et al. [169]; Usman et al. [158] | ||
| Secondary alcohol | 3β-Taraxerol | 200 mg | Human | Type 2 diabetes | Improved pancreatic function Increased insulin secretion | Rasouli et al. [161] |
| Organic compounds | Sulfonylureas | 5 mg | Human | Type 2 diabetes | Increased insulin secretion | Nathan et al. [155] |
| FDA/Commercial Drugs | Metformin | 5 mg | Human, mouse, rat | Type 2 diabetes | Improved glucose uptake, lipotoxicity, and antioxidant activities | Pan et al. [153] |
| Miglitol | 25 mg | Human | Type 2 diabetes | Inhibited α-glucosidase Induced antihyperglycemic activities | Rasouli et al. [168] | |
| SGLT2 inhibitors | 5–10 mg | Human | Type 2 diabetes | Decreased glucose absorption Increased anti-inflammatory and antioxidative activities | Whalen et al. [156] | |
| Thiazolidinediones | 8 mg | Human | Type 2 diabetes | Acted as PPARγ agonist | Soccio et al. [157] | |
| Colesevelam | 3.75 g | Human | Type 2 diabetes | Increased triglycerides Decreased LDLc | Feingold, [170] | |
| Gallotannins | 10–100 ng/mL | Human, rat | Type 2 diabetes | Increased PI3K mRNA and GLUT4 expressions | Kanaujia et al. [165] | |
| DPP-IV inhibitors | 50 mg | Human | Type 2 diabetes | Stimulated incretin system and SGLT-2 | Jose and Inzucchi, [166] | |
| Oxovanadium complex | 200 mg | Mouse | Diabetic mice | Increased blood SOD, GSH, TP, and LC3 levels | El-Shafey and Elsherbiny, [167] | |
| Acarbose | 50 mg | human | Type 2 diabetes | Inhibited α-glucosidase and α-amylase | Rasouli et al. [168] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rahman, M.S.; Hossain, K.S.; Das, S.; Kundu, S.; Adegoke, E.O.; Rahman, M.A.; Hannan, M.A.; Uddin, M.J.; Pang, M.-G. Role of Insulin in Health and Disease: An Update. Int. J. Mol. Sci. 2021, 22, 6403. https://doi.org/10.3390/ijms22126403
Rahman MS, Hossain KS, Das S, Kundu S, Adegoke EO, Rahman MA, Hannan MA, Uddin MJ, Pang M-G. Role of Insulin in Health and Disease: An Update. International Journal of Molecular Sciences. 2021; 22(12):6403. https://doi.org/10.3390/ijms22126403
Chicago/Turabian StyleRahman, Md Saidur, Khandkar Shaharina Hossain, Sharnali Das, Sushmita Kundu, Elikanah Olusayo Adegoke, Md. Ataur Rahman, Md. Abdul Hannan, Md Jamal Uddin, and Myung-Geol Pang. 2021. "Role of Insulin in Health and Disease: An Update" International Journal of Molecular Sciences 22, no. 12: 6403. https://doi.org/10.3390/ijms22126403
APA StyleRahman, M. S., Hossain, K. S., Das, S., Kundu, S., Adegoke, E. O., Rahman, M. A., Hannan, M. A., Uddin, M. J., & Pang, M.-G. (2021). Role of Insulin in Health and Disease: An Update. International Journal of Molecular Sciences, 22(12), 6403. https://doi.org/10.3390/ijms22126403