
Advances and Perspectives in Tissue Culture and Genetic Engineering of Cannabis

 ,
,  and
and
Abstract
1. Introduction
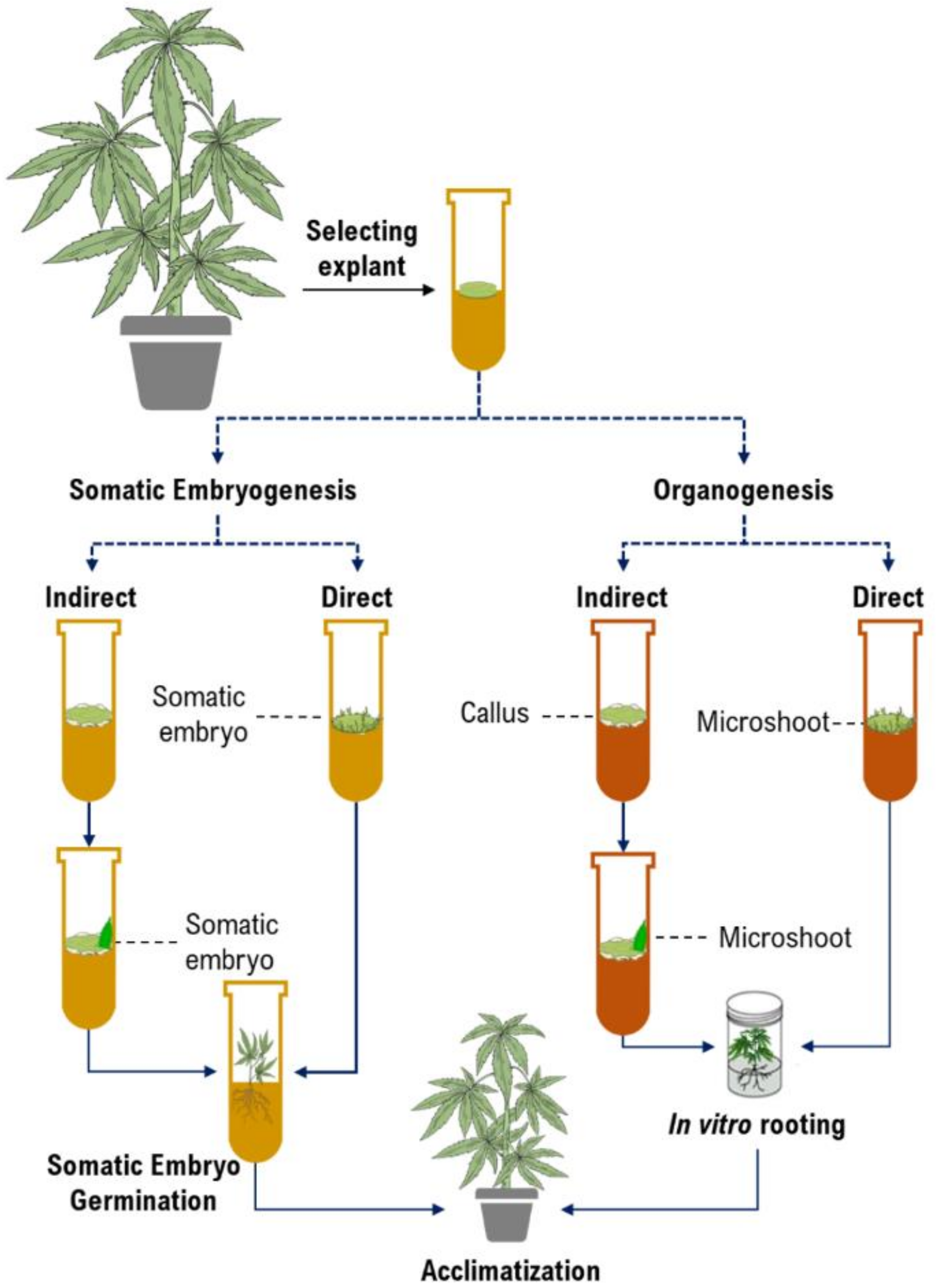
2. In vitro Culture in Cannabis
2.1. Strategies to Improve In Vitro Culture Procedures
2.2. Somaclonal Variation
3. Ploidy Engineering in Cannabis
3.1. Types of Polyploids
3.2. Advantages to Polyploidy in Breeding Programs
3.3. Disadvantages to Polyploid Breeding
3.4. Effects of Polyploidy
3.5. Secondary Metabolites
3.6. Limitations of Existing Polyploidy Literature and Future Potential
4. Genetic Engineering Approaches in Cannabis
4.1. Agrobacterium-Mediated Gene Transformation
4.1.1. Agrobacterium Strains
4.1.2. Infection of Explant
4.1.3. Selection Markers
4.1.4. Eliminating Chimerism
4.1.5. Promoters and Translational Enhancer
4.2. Strategies to Improve Gene Transformation Efficiency
4.2.1. Morphogenic Genes, Key Factors in Plant Regeneration
4.2.2. Strategies to Overcome Pleiotropic Effects
4.3. Strategies to Prevent Transgene Escape
4.4. CRISPR/Cas-Mediated Genome Editing
5. Conclusions and Future Objectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kovalchuk, I.; Pellino, M.; Rigault, P.; van Velzen, R.; Ebersbach, J.; Ashnest, J.; Mau, M.; Schranz, M.; Alcorn, J.; Laprairie, R. The Genomics of Cannabis and Its Close Relatives. Annu. Rev. plant Biol 2020, 71, 713–739. [Google Scholar] [CrossRef] [PubMed]
- Hesami, M.; Pepe, M.; Alizadeh, M.; Rakei, A.; Baiton, A.; Jones, A.M.P. Recent advances in cannabis biotechnology. Ind. Crop. Prod. 2020, 158, 113026. [Google Scholar] [CrossRef]
- Aliferis, K.A.; Bernard-Perron, D. Cannabinomics: Application of Metabolomics in Cannabis (Cannabis sativa L.) Research and Development. Front. plant Sci. 2020, 11, 554. [Google Scholar] [CrossRef] [PubMed]
- Romero, P.; Peris, A.; Vergara, K.; Matus, J.T. Comprehending and improving cannabis specialized metabolism in the systems biology era. plant Sci. 2020, 298, 110571. [Google Scholar] [CrossRef] [PubMed]
- Schultz, C.J.; Lim, W.L.; Khor, S.F.; Neumann, K.A.; Schulz, J.M.; Ansari, O.; Skewes, M.A.; Burton, R.A. Consumer and health-related traits of seed from selected commercial and breeding lines of industrial hemp, Cannabis sativa L. J. Agric. Food Res. 2020, 2, 100025. [Google Scholar] [CrossRef]
- Salami, S.A.; Martinelli, F.; Giovino, A.; Bachari, A.; Arad, N.; Mantri, N. It Is Our Turn to Get Cannabis High: Put Cannabinoids in Food and Health Baskets. Molecules 2020, 25, 4036. [Google Scholar] [CrossRef]
- Schilling, S.; Dowling, C.A.; Shi, J.; Ryan, L.; Hunt, D.; Oreilly, E.; Perry, A.S.; Kinnane, O.; McCabe, P.F.; Melzer, R. The Cream of the Crop: Biology, Breeding and Applications of Cannabis sativa. Authorea Prepr. 2020. [Google Scholar] [CrossRef]
- Devsi, A.; Kiyota, B.; Ouellette, T.; Hegle, A.P.; Rivera-Acevedo, R.E.; Wong, J.; Dong, Y.; Pugsley, M.K.; Fung, T. A pharmacological characterization of Cannabis sativa chemovar extracts. J. Cannabis Res. 2020, 2, 17. [Google Scholar] [CrossRef]
- Valleriani, J. Identity narratives in the face of market competition: The emerging legal medical cannabis market in Canada. Drugs Educ. Prev. Policy 2020, 27, 37–48. [Google Scholar] [CrossRef]
- Wróbel, T.; Dreger, M.; Wielgus, K.; Słomski, R. The application of plant in vitro cultures in cannabinoid production. Biotechnol. Lett. 2018, 40, 445–454. [Google Scholar] [CrossRef]
- Farag, S.; Kayser, O. Cultivation and breeding of Cannabis sativa L. for preparation of standardized extracts for medicinal purposes. In Medicinal and Aromatic Plants of the World; Máthé, Á., Ed.; Springer: Berlin/Heidelberg, Germany, 2015; Volume 1, pp. 165–186. [Google Scholar]
- Maayah, Z.H.; Takahara, S.; Ferdaoussi, M.; Dyck, J.R.B. The molecular mechanisms that underpin the biological benefits of full-spectrum cannabis extract in the treatment of neuropathic pain and inflammation. Biochim. Biophys. Acta (Bba) Mol. Basis Dis. 2020, 1866, 165771. [Google Scholar] [CrossRef] [PubMed]
- Workman, C.D.; Kindred, J.H.; Boles Ponto, L.L.; Kamholz, J.; Rudroff, T. The effects of chronic Δ-9-tetrahydrocannabinol (THC) and cannabidiol (CBD) use on cerebral glucose metabolism in multiple sclerosis: A pilot study. Appl. Physiol. Nutr. Metab. 2019, 45, 450–452. [Google Scholar] [CrossRef] [PubMed]
- Cassano, T.; Villani, R.; Pace, L.; Carbone, A.; Bukke, V.N.; Orkisz, S.; Avolio, C.; Serviddio, G. From Cannabis sativa to Cannabidiol: Promising Therapeutic Candidate for the Treatment of Neurodegenerative Diseases. Front. plant Sci. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Alves, P.; Amaral, C.; Teixeira, N.; Correia-da-Silva, G. Cannabis sativa: Much more beyond Δ9-tetrahydrocannabinol. Pharmacol. Res. 2020, 157, 104822. [Google Scholar] [CrossRef]
- Baron, E.P. Medicinal Properties of Cannabinoids, Terpenes, and Flavonoids in Cannabis, and Benefits in Migraine, Headache, and Pain: An Update on Current Evidence and Cannabis Science. J. Head Face Pain 2018, 58, 1139–1186. [Google Scholar] [CrossRef]
- Nuutinen, T. Medicinal properties of terpenes found in Cannabis sativa and Humulus lupulus. Eur. J. Med. Chem. 2018, 157, 198–228. [Google Scholar] [CrossRef]
- Hwang, E.; Ngo, H.T.T.; Park, B.; Seo, S.-A.; Yang, J.-E.; Yi, T.-H. Myrcene, an Aromatic Volatile Compound, Ameliorates Human Skin Extrinsic Aging via Regulation of MMPs Production. Am. J. Chin. Med. 2017, 45, 1113–1124. [Google Scholar] [CrossRef]
- Aly, E.; Khajah, M.A.; Masocha, W. β-Caryophyllene, a CB2-Receptor-Selective Phytocannabinoid, Suppresses Mechanical Allodynia in a Mouse Model of Antiretroviral-Induced Neuropathic Pain. Molecules 2020, 25, 106. [Google Scholar] [CrossRef] [PubMed]
- Ciftci, O.; Ozdemir, I.; Tanyildizi, S.; Yildiz, S.; Oguzturk, H. Antioxidative effects of curcumin, β-myrcene and 1,8-cineole against 2,3,7,8-tetrachlorodibenzo-p-dioxin-induced oxidative stress in rats liver. Toxicol. Ind. Health 2011, 27, 447–453. [Google Scholar] [CrossRef]
- Shah, S.; Dhanani, T.; Sharma, S.; Singh, R.; Kumar, S.; Kumar, B.; Srivastava, S.; Ghosh, S.; Kumar, R.; Juliet, S. Development and Validation of a Reversed Phase High Performance Liquid Chromatography-Photodiode Array Detection Method for Simultaneous Identification and Quantification of Coumarin, Precocene-I, β-Caryophyllene Oxide, α-Humulene, and β-Caryophyllene in Ageratum Conyzoides Extracts and Essential Oils from Plants. J. AOAC Int. 2020, 103, 857–864. [Google Scholar] [CrossRef]
- Kołodziejczyk, A.; Pyrcz, P.; Błaziak, K.; Pobudkowska, A.; Sarang, K.; Szmigielski, R. Physicochemical Properties of Terebic Acid, MBTCA, Diaterpenylic Acid Acetate, and Pinanediol as Relevant α-Pinene Oxidation Products. ACS Omega 2020, 5, 7919–7927. [Google Scholar] [CrossRef]
- Guzmán-Gutiérrez, S.L.; Bonilla-Jaime, H.; Gómez-Cansino, R.; Reyes-Chilpa, R. Linalool and β-pinene exert their antidepressant-like activity through the monoaminergic pathway. Life Sci. 2015, 128, 24–29. [Google Scholar] [CrossRef]
- Jana, S.; Patra, K.; Sarkar, S.; Jana, J.; Mukherjee, G.; Bhattacharjee, S.; Mandal, D.P. Antitumorigenic Potential of Linalool Is Accompanied by Modulation of Oxidative Stress: An In Vivo Study in Sarcoma-180 Solid Tumor Model. Nutr. Cancer 2014, 66, 835–848. [Google Scholar] [CrossRef] [PubMed]
- Srividya, N.; Lange, I.; Lange, B.M. Determinants of Enantiospecificity in Limonene Synthases. Biochemistry 2020, 59, 1661–1664. [Google Scholar] [CrossRef]
- Faria, G.M.; Soares, I.D.P.; D’Alincourt Salazar, M.; Amorim, M.R.; Pessoa, B.L.; da Fonseca, C.O.; Quirico-Santos, T. Intranasal perillyl alcohol therapy improves survival of patients with recurrent glioblastoma harboring mutant variant for MTHFR rs1801133 polymorphism. BMC Cancer 2020, 20, 294. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Wang, Y.; Huang, W.; He, L.; Lin, Z.; Zhou, J.; He, Q. Toxic effects of terpinolene on Microcystis aeruginosa: Physiological, metabolism, gene transcription, and growth effects. Sci. Total Environ. 2020, 719, 137376. [Google Scholar] [CrossRef] [PubMed]
- Castro, J.A.M.d.; Monteiro, O.S.; Coutinho, D.F.; Rodrigues, A.A.C.; Silva, J.K.R.d.; Maia, J. Seasonal and Circadian Study of a Thymol/3-Terpinene/p-Cymene Type Oil of Ocimum gratissimum L. and Its Antioxidant and Antifungal Effects. J. Braz. Chem. Soc. 2019, 30, 930–938. [Google Scholar] [CrossRef]
- de Oliveira, M.G.B.; Marques, R.B.; de Santana, M.F.; Santos, A.B.D.; Brito, F.A.; Barreto, E.O.; De Sousa, D.P.; Almeida, F.R.C.; Badauê-Passos, D., Jr.; Antoniolli, Â.R.; et al. α-Terpineol Reduces Mechanical Hypernociception and Inflammatory Response. Basic Clin. Pharmacol. Toxicol. 2012, 111, 120–125. [Google Scholar] [CrossRef] [PubMed]
- Vieira, G.; Cavalli, J.; Gonçalves, E.C.D.; Braga, S.F.P.; Ferreira, R.S.; Santos, A.R.S.; Cola, M.; Raposo, N.R.B.; Capasso, R.; Dutra, R.C. Antidepressant-Like Effect of Terpineol in an Inflammatory Model of Depression: Involvement of the Cannabinoid System and D2 Dopamine Receptor. Biomolecules 2020, 10, 792. [Google Scholar] [CrossRef] [PubMed]
- Lira, M.H.P.d.; Andrade Júnior, F.P.d.; Moraes, G.F.Q.; Macena, G.d.S.; Pereira, F.d.O.; Lima, I.O. Antimicrobial activity of geraniol: An integrative review. J. Essent. Oil Res. 2020, 32, 187–197. [Google Scholar] [CrossRef]
- Barros Silva Soares de Souza, E.P.; Trindade, G.d.G.G.; Lins Dantas Gomes, M.V.; Santos Silva, L.A.; Grespan, R.; Quintans Junior, L.J.; Cavalcanti de Albuquerque Júnior, R.L.; Shanmugan, S.; Antunes de Souza Araújo, A. Anti-inflammatory effect of nano-encapsulated nerolidol on zymosan-induced arthritis in mice. Food Chem. Toxicol. 2020, 135, 110958. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Liu, Q.; Lv, H.; Wang, F.; Liu, R.; Zeng, N. Effect of pulegone on the NLPR3 inflammasome during inflammatory activation of THP-1 cells. Exp. Ther. Med. 2020, 19, 1304–1312. [Google Scholar] [CrossRef]
- Xu, C.; Sheng, S.; Dou, H.; Chen, J.; Zhou, K.; Lin, Y.; Yang, H. α-Bisabolol suppresses the inflammatory response and ECM catabolism in advanced glycation end products-treated chondrocytes and attenuates murine osteoarthritis. Int. Immunopharmacol. 2020, 84, 106530. [Google Scholar] [CrossRef] [PubMed]
- Gogineni, V.; Nael, M.A.; León, F.; Núñez, M.J.; Cutler, S.J. Computationally aided stereochemical assignment of undescribed bisabolenes from Calea urticifolia. Phytochemistry 2019, 157, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Tong, H.; Liu, Y.; Jiang, L.; Wang, J. Multi-Targeting by β-Elemene and Its Anticancer Properties: A Good Choice for Oncotherapy and Radiochemotherapy Sensitization. Nutr. Cancer 2020, 72, 554–567. [Google Scholar] [CrossRef]
- Müller, A.D.; Kutscher, E.; Artemyev, A.N.; Demekhin, P.V. Photoelectron circular dichroism in the multiphoton ionization by short laser pulses. III. Photoionization of fenchone in different regimes. J. Chem. Phys. 2020, 152, 044302. [Google Scholar] [CrossRef]
- de Christo Scherer, M.M.; Marques, F.M.; Figueira, M.M.; Peisino, M.C.O.; Schmitt, E.F.P.; Kondratyuk, T.P.; Endringer, D.C.; Scherer, R.; Fronza, M. Wound healing activity of terpinolene and α-phellandrene by attenuating inflammation and oxidative stress in vitro. J. Tissue Viability 2019, 28, 94–99. [Google Scholar] [CrossRef]
- Kotawong, K.; Chaijaroenkul, W.; Roytrakul, S.; Phaonakrop, N.; Na-Bangchang, K. Proteomics Analysis for Identification of Potential Cell Signaling Pathways and Protein Targets of Actions of Atractylodin and β-Eudesmol Against Cholangiocarcinoma. Asian Pac. J. Cancer Prev. 2020, 21, 621–628. [Google Scholar] [CrossRef]
- Lubell, J.D.; Brand, M.H. Foliar sprays of silver thiosulfate produce male flowers on female hemp plants. HortTechnology 2018, 28, 743–747. [Google Scholar] [CrossRef]
- Barnett, P. Transcription Activator Like Effector Nucleases (TALENs): A New, Important, and Versatile Gene Editing Technique with a Growing Literature. Sci. Technol. Libr. 2018, 37, 100–112. [Google Scholar] [CrossRef]
- Novak, S. Plant biotechnology applications of zinc finger technology. In Transgenic Plants. Methods in Molecular Biology; Kumar, S., Barone, P., Smith, M., Eds.; Humana Press: New York, NY, USA, 2019; Volume 1864, pp. 295–310. [Google Scholar]
- Zhu, H.; Li, C.; Gao, C. Applications of CRISPR–Cas in agriculture and plant biotechnology. Nat. Rev. Mol. Cell Biol. 2020, 21, 661–677. [Google Scholar] [CrossRef] [PubMed]
- Barcaccia, G.; Palumbo, F.; Scariolo, F.; Vannozzi, A.; Borin, M.; Bona, S. Potentials and Challenges of Genomics for Breeding Cannabis Cultivars. Front. plant Sci. 2020, 11, 1472. [Google Scholar] [CrossRef] [PubMed]
- Torkamaneh, D.; Boyle, B.; Belzile, F. Efficient genome-wide genotyping strategies and data integration in crop plants. Theor. Appl. Genet. 2018, 131, 499–511. [Google Scholar] [CrossRef] [PubMed]
- Niazian, M. Application of genetics and biotechnology for improving medicinal plants. Planta 2019, 249, 953–973. [Google Scholar] [CrossRef] [PubMed]
- Schachtsiek, J.; Warzecha, H.; Kayser, O.; Stehle, F. Current perspectives on biotechnological cannabinoid production in plants. Planta Med. 2018, 84, 214–220. [Google Scholar] [CrossRef]
- Salentijn, E.M.J.; Petit, J.; Trindade, L.M. The Complex Interactions Between Flowering Behavior and Fiber Quality in Hemp. Front. plant Sci. 2019, 10, 614. [Google Scholar] [CrossRef]
- Hemphill, J.K.; JK, H.; JC, T.; PG, M. Studies on growth and cannabinoid composition of callus derived from different strains of Cannabis sativa. Lloydia 1978, 41, 453–462. [Google Scholar]
- Loh, W.H.T.; Hartsel, S.C.; Robertson, L.W. Tissue Culture of Cannabis sativa L. and in vitro Biotransformation of Phenolics. Z. Pflanzenphysiol. 1983, 111, 395–400. [Google Scholar] [CrossRef]
- Braemer, R.; Paris, M. Biotransformation of cannabinoids by a cell suspension culture of Cannabis sativa L. plant Cell Rep. 1987, 6, 150–152. [Google Scholar] [CrossRef]
- Sirikantaramas, S.; Taura, F.; Tanaka, Y.; Ishikawa, Y.; Morimoto, S.; Shoyama, Y. Tetrahydrocannabinolic Acid Synthase, the Enzyme Controlling Marijuana Psychoactivity, is Secreted into the Storage Cavity of the Glandular Trichomes. plant Cell Physiol. 2005, 46, 1578–1582. [Google Scholar] [CrossRef]
- Staginnus, C.; Zörntlein, S.; de Meijer, E. A PCR marker Linked to a THCA synthase Polymorphism is a Reliable Tool to Discriminate Potentially THC-Rich Plants of Cannabis sativa L. J. Forensic Sci. 2014, 59, 919–926. [Google Scholar] [CrossRef] [PubMed]
- Flores-Sanchez, I.J.; Peč, J.; Fei, J.; Choi, Y.H.; Dušek, J.; Verpoorte, R. Elicitation studies in cell suspension cultures of Cannabis sativa L. J. Biotechnol. 2009, 143, 157–168. [Google Scholar] [CrossRef] [PubMed]
- Gabotti, D.; Locatelli, F.; Cusano, E.; Baldoni, E.; Genga, A.; Pucci, L.; Consonni, R.; Mattana, M. Cell Suspensions of Cannabis sativa (var. Futura): Effect of Elicitation on Metabolite Content and Antioxidant Activity. Molecules 2019, 24, 4056. [Google Scholar] [CrossRef]
- Baek, S.; Ho, T.-T.; Lee, H.; Jung, G.; Kim, Y.E.; Jeong, C.-S.; Park, S.-Y. Enhanced biosynthesis of triterpenoids in Centella asiatica hairy root culture by precursor feeding and elicitation. plant Biotechnol. Rep. 2020, 14, 45–53. [Google Scholar] [CrossRef]
- Sirikantaramas, S.; Morimoto, S.; Shoyama, Y.; Ishikawa, Y.; Wada, Y.; Shoyama, Y.; Taura, F. The gene controlling marijuana psychoactivity molecular cloning and heterologous expression of Δ1-tetrahydrocannabinolic acid synthase from Cannabis sativa L. J. Biol. Chem. 2004, 279, 39767–39774. [Google Scholar] [CrossRef]
- Farag, S.; Kayser, O. Cannabinoids production by hairy root cultures of Cannabis sativa L. Am. J. plant Sci. 2015, 6, 1874. [Google Scholar] [CrossRef]
- Moher, M.; Jones, M.; Zheng, Y. Photoperiodic Response of in vitro Cannabis sativa Plants. HortScience 2021, 56, 108–113. [Google Scholar] [CrossRef]
- Monthony, A.S.; Page, S.R.G.; Hesami, M.; Jones, A.M.P. The Past, Present and Future of Cannabis sativa Tissue Culture. Plants 2021, 10, 185. [Google Scholar] [CrossRef] [PubMed]
- Adhikary, D.; Kulkarni, M.; El-Mezawy, A.; Mobini, S.; Elhiti, M.; Gjuric, R.; Ray, A.; Polowick, P.; Slaski, J.J.; Jones, M.P.; et al. Medical Cannabis and Industrial Hemp Tissue Culture: Present Status and Future Potential. Front. Plant Sci. 2021, 12, 627240. [Google Scholar] [CrossRef]
- Veliky, I.; Genest, K. Growth and metabolites of Cannabis sativa cell suspension cultures. Lloydia 1972, 35, 450–456. [Google Scholar] [PubMed]
- Itokawa, H.; Takeya, K.; Akasu, M. Studies on the constituents isolated from the callus of Cannabis sativa. Shoyakugaku Zasshi 1975, 29, 106–112. [Google Scholar]
- Hartsel, S.; Loh, W.-T.; Robertson, L. Biotransformation of cannabidiol to cannabielsoin by suspension cultures of Cannabis sativa and Saccharum officinarum. Planta Med. 1983, 48, 17–19. [Google Scholar] [CrossRef] [PubMed]
- Richez-Dumanois, C.; Braut-Boucher, F.; Cosson, L.; Paris, M. Multiplication végétative in vitro du chanvre (Cannabis sativa L.). Application à la conserva-tion des clones sélectionnés. Agronomie 1986, 6, 487–495. [Google Scholar] [CrossRef]
- Mandolino, G.; Ranalli, P. Advances in biotechnological approaches for hemp breeding and industry. In Advances in Hemp Research; Ranalli, P., Ed.; Haworth Press: Binghamton, NY, USA, 1999; pp. 185–212. [Google Scholar]
- Slusarkiewicz-Jarzina, A.; Ponitka, A.; Kaczmarek, Z. Influence of cultivar, explant source and plant growth regulator on callus induction and plant regeneration of Cannabis sativa L. Acta Biol. Crac. Ser. Bot. 2005, 47, 145–151. [Google Scholar]
- Plawuszewski, M.; Lassocinski, W.; Wielgus, K. Regeneration of Polish cultivars of monoecious hemp (Cannabis sativa L.) grown in vitro. In Renewable Resources and Plant Biotechnology; Kozłowski, R., Zaikov, G., Pudel, F., Eds.; Nova Publishers: Poznan, Poland, 2006; pp. 149–154. [Google Scholar]
- Bing, X.; Ning, L.; Jinfeng, T.; Nan, G. Rapid Tissue Culture Method of Cannabis sativa for Industrial Uses. CN Patent 1887043, 2007. [Google Scholar]
- Wielgus, K.; Luwanska, A.; Lassocinski, W.; Kaczmarek, Z. Estimation of Cannabis sativa L. tissue culture conditions essential for callus induction and plant regeneration. J. Nat. Fibers 2008, 5, 199–207. [Google Scholar] [CrossRef]
- Wang, R.; He, L.; Xia, B.; Tong, J.; Li, N.; Peng, F. A micropropagation system for cloning of hemp (Cannabis sativa L.) by shoot tip culture. Pak. J. Bot. 2009, 41, 603–608. [Google Scholar]
- Lata, H.; Chandra, S.; Khan, I.; ElSohly, M.A. Thidiazuron-induced high-frequency direct shoot organogenesis of Cannabis sativa L. Vitr. Cell. Dev. Biol. Plant 2009, 45, 12–19. [Google Scholar] [CrossRef]
- Lata, H.; Chandra, S.; Khan, I.A.; ElSohly, M.A. Propagation through alginate encapsulation of axillary buds of Cannabis sativa L.—an important medicinal plant. Physiol. Mol. Biol. Plants 2009, 15, 79–86. [Google Scholar] [CrossRef]
- Lata, H.; Chandra, S.; Khan, I.A.; ElSohly, M.A. High Frequency Plant Regeneration from Leaf Derived Callus of High Δ9-Tetrahydrocannabinol Yielding Cannabis sativa L. Planta Med. 2010, 76, 1629–1633. [Google Scholar] [CrossRef]
- Movahedi, M.; Ghasemi-Omran, V.; Torabi, S. The effect of different concentrations of TDZ and BA on in vitro regeneration of Iranian cannabis (Cannabis sativa) using cotyledon and epicotyl explants. J. plant Mol. Breed. 2015, 3, 20–27. [Google Scholar] [CrossRef]
- Movahedi, M.; Ghasemiomran, V.; Torabi, S. In vitro callus induction and regeneration of medicinal plant Cannabis sativa L. Iran. J. Med. Aromat. Plants 2016, 32, 758–768. [Google Scholar] [CrossRef]
- Movahedi, M.; Ghasemiomran, V.; Torabi, S. Effect of explants type and plant growth regulators on in vitro callus induction and shoot regeneration of Cannabis sativa L. Iran. J. Med. Aromat. Plants 2016, 32, 83–96. [Google Scholar] [CrossRef]
- Lata, H.; Chandra, S.; Techen, N.; Khan, I.A.; ElSohly, M.A. In vitro mass propagation of Cannabis sativa L.: A protocol refinement using novel aromatic cytokinin meta-topolin and the assessment of eco-physiological, biochemical and genetic fidelity of micropropagated plants. J. Appl. Res. Med. Aromat. Plants 2016, 3, 18–26. [Google Scholar] [CrossRef]
- Chaohua, C.; Gonggu, Z.; Lining, Z.; Chunsheng, G.; Qing, T.; Jianhua, C.; Xinbo, G.; Dingxiang, P.; Jianguang, S. A rapid shoot regeneration protocol from the cotyledons of hemp (Cannabis sativa L.). Ind. Crop. Prod. 2016, 83, 61–65. [Google Scholar] [CrossRef]
- Piunno, K.F.; Golenia, G.; Boudko, E.A.; Downey, C.; Jones, A.M.P. Regeneration of shoots from immature and mature inflorescences of Cannabis sativa. Can. J. plant Sci. 2019, 99, 556–559. [Google Scholar] [CrossRef]
- Smýkalová, I.; Vrbová, M.; Cvečková, M.; Plačková, L.; Žukauskaitė, A.; Zatloukal, M.; Hrdlička, J.; Plíhalová, L.; Doležal, K.; Griga, M. The effects of novel synthetic cytokinin derivatives and endogenous cytokinins on the in vitro growth responses of hemp (Cannabis sativa L.) explants. plant Cell Tissue Organ. Cult. 2019, 139, 381–394. [Google Scholar] [CrossRef]
- Monthony, A.S.; Kyne, S.T.; Grainger, C.M.; Jones, A.M.P. Recalcitrance of Cannabis sativa to de novo regeneration; a multi-genotype replication study. bioRxiv 2020. [Google Scholar] [CrossRef]
- Wróbel, T.; Dreger, M.; Wielgus, K.; Słomski, R. Modified Nodal Cuttings and Shoot Tips Protocol for Rapid Regeneration of Cannabis sativa L. J. Nat. Fibers 2020, 8, 1–10. [Google Scholar] [CrossRef]
- Galán-Ávila, A.; García-Fortea, E.; Prohens, J.; Herraiz, F.J. Development of a Direct in vitro Plant Regeneration Protocol From Cannabis sativa L. Seedling Explants: Developmental Morphology of Shoot Regeneration and Ploidy Level of Regenerated Plants. Front. plant Sci. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Monthony, A.S.; Bagheri, S.; Zheng, Y.; Jones, A.M.P. Flower Power: Floral reversion as a viable alternative to nodal micropropagation in Cannabis sativa. bioRxiv 2020. [Google Scholar] [CrossRef]
- Codesido, V.; Meyer, S.; Casano, S. Influence of media composition and genotype for successful Cannabis sativa L. in vitro introduction. Acta Hortic. 2020, 1285, 75–80. [Google Scholar] [CrossRef]
- Mubi, Š.M.; Svetik, S.; FlajŠMan, M.; Murovec, J.; Mestinšek Mubi, Š.; Svetik, S.; Flajšman, M.; Murovec, J. In vitro tissue culture and genetic analysis of two high-CBD medical cannabis (Cannabis sativa L.) breeding lines. GENETIKA 2020, 52, 925–941. [Google Scholar] [CrossRef]
- Ioannidis, K.; Dadiotis, E.; Mitsis, V.; Melliou, E.; Magiatis, P. Biotechnological Approaches on Two High CBD and CBG Cannabis sativa L. (Cannabaceae) Varieties: In vitro Regeneration and Phytochemical Consistency Evaluation of Micropropagated Plants Using Quantitative 1H-NMR. Molecules 2020, 25, 5928. [Google Scholar] [CrossRef]
- Jessica, D.L.-B.; Lauren, E.K.; Mark, H.B. An In vitro–ex vitro Micropropagation System for Hemp. Horttechnol. Hortte 2021, 31, 199–207. [Google Scholar] [CrossRef]
- Murphy, R.; Adelberg, J. Physical factors increased quantity and quality of micropropagated shoots of Cannabis sativa L. in a repeated harvest system with ex vitro rooting. Vitr. Cell. Dev. Biol. Plant 2021, 57, 1–9. [Google Scholar] [CrossRef]
- Zarei, A.; Behdarvandi, B.; Tavakouli Dinani, E.; Maccarone, J. Cannabis sativa L. photoautotrophic micropropagation: A powerful tool for industrial scale in vitro propagation. Vitr. Cell. Dev. Biol. Plant 2021, 57, 1–10. [Google Scholar] [CrossRef]
- Page, S.R.G.; Monthony, A.S.; Jones, A.M.P. DKW basal salts improve micropropagation and callogenesis compared to MS basal salts in multiple commercial cultivars of Cannabis sativa. Botany 2021, 99, 269–279. [Google Scholar] [CrossRef]
- Zhang, X.; Xu, G.; Cheng, C.; Lei, L.; Sun, J.; Xu, Y.; Deng, C.; Dai, Z.; Yang, Z.; Chen, X.; et al. Establishment of an Agrobacterium-mediated genetic transformation and CRISPR/Cas9-mediated targeted mutagenesis in Hemp (Cannabis Sativa L.). plant Biotechnol. J. 2021, 19, 1–25. [Google Scholar] [CrossRef]
- Boonsnongcheep, P.; Pongkitwitoon, B. Factors affecting micropropagation of Cannabis sativa L.: A review. Pharm. Sci. Asia 2020, 47, 21–29. [Google Scholar] [CrossRef]
- Hesami, M.; Jones, A.M.P. Application of artificial intelligence models and optimization algorithms in plant cell and tissue culture. Appl. Microbiol. Biotechnol. 2020, 104, 9449–9485. [Google Scholar] [CrossRef]
- Hesami, M.; Naderi, R.; Tohidfar, M. Introducing a hybrid artificial intelligence method for high-throughput modeling and optimizing plant tissue culture processes: The establishment of a new embryogenesis medium for chrysanthemum, as a case study. Appl. Microbiol. Biotechnol. 2020, 104, 10249–10263. [Google Scholar] [CrossRef]
- Mira, M.M.; Wally, O.S.; Elhiti, M.; El-Shanshory, A.; Reddy, D.S.; Hill, R.D.; Stasolla, C. Jasmonic acid is a downstream component in the modulation of somatic embryogenesis by Arabidopsis Class 2 phytoglobin. J. Exp. Bot. 2016, 67, 2231–2246. [Google Scholar] [CrossRef] [PubMed]
- Sarropoulou, V.; Maloupa, E. Effect of the NO donor “sodium nitroprusside”(SNP), the ethylene inhibitor “cobalt chloride”(CoCl 2) and the antioxidant vitamin E “α-tocopherol” on in vitro shoot proliferation of Sideritis raeseri Boiss. & Heldr. subsp. raeseri. plant Cell Tissue Organ. Cult. 2017, 128, 619–629. [Google Scholar] [CrossRef]
- Ötvös, K.; Pasternak, T.P.; Miskolczi, P.; Domoki, M.; Dorjgotov, D.; Sz˝ cs, A.; Bottka, S.; Dudits, D.; Fehér, A. Nitric oxide is required for, and promotes auxin-mediated activation of, cell division and embryogenic cell formation but does not influence cell cycle progression in alfalfa cell cultures. Plant J. 2005, 43, 849–860. [Google Scholar] [CrossRef] [PubMed]
- Tariq, A.; Ilyas, S.; Naz, S. Nanotechnology and Plant Tissue Culture. In Nanoagronomy; Javad, S., Ed.; Springer: Cham, Switzerland, 2020; pp. 23–35. [Google Scholar]
- Ahmad, M.A.; Javed, R.; Adeel, M.; Rizwan, M.; Ao, Q.; Yang, Y. Engineered ZnO and CuO Nanoparticles Ameliorate Morphological and Biochemical Response in Tissue Culture Regenerants of Candyleaf (Stevia rebaudiana). Molecules 2020, 25, 1356. [Google Scholar] [CrossRef]
- Zaeem, A.; Drouet, S.; Anjum, S.; Khurshid, R.; Younas, M.; Blondeau, J.P.; Tungmunnithum, D.; Giglioli-Guivarc’h, N.; Hano, C.; Abbasi, B.H. Effects of Biogenic Zinc Oxide Nanoparticles on Growth and Oxidative Stress Response in Flax Seedlings vs. in vitro Cultures: A Comparative Analysis. Biomolecules 2020, 10, 918. [Google Scholar] [CrossRef]
- Nalci, O.B.; Nadaroglu, H.; Pour, A.H.; Gungor, A.A.; Haliloglu, K. Effects of ZnO, CuO and γ-Fe3O4 nanoparticles on mature embryo culture of wheat (Triticum aestivum L.). plant Cell Tissue Organ. Cult. 2019, 136, 269–277. [Google Scholar] [CrossRef]
- Hesami, M.; Daneshvar, M.H.; Yoosefzadeh-Najafabadi, M. An efficient in vitro shoot regeneration through direct organogenesis from seedling-derived petiole and leaf segments and acclimatization of Ficus religiosa. J. For. Res. 2019, 30, 807–815. [Google Scholar] [CrossRef]
- Hesami, M.; Tohidfar, M.; Alizadeh, M.; Daneshvar, M.H. Effects of sodium nitroprusside on callus browning of Ficus religiosa: An important medicinal plant. J. For. Res. 2020, 31, 789–796. [Google Scholar] [CrossRef]
- Yaseen, M.; Ahmad, T.; Sablok, G.; Standardi, A.; Hafiz, I.A. Review: Role of carbon sources for in vitro plant growth and development. Mol. Biol. Rep. 2013, 40, 2837–2849. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Díaz, J.I.; Arzate-Fernández, A.M.; Piña-Escutia, J.L.; Vázquez-García, L.M. Media culture factors affecting somatic embryogenesis in Agave angustifolia Haw. Ind. Crop. Prod. 2017, 108, 81–85. [Google Scholar] [CrossRef]
- Asthana, P.; Rai, M.K.; Jaiswal, U. Somatic embryogenesis from sepal explants in Sapindus trifoliatus, a plant valuable in herbal soap industry. Ind. Crop. Prod. 2017, 100, 228–235. [Google Scholar] [CrossRef]
- Peng, C.; Gao, F.; Wang, H.; Shen, H.; Yang, L. Optimization of maturation process for somatic embryo production and cryopreservation of embryogenic tissue in Pinus koraiensis. plant Cell Tissue Organ. Cult. 2020. [Google Scholar] [CrossRef]
- Yancheva, S.D.; Roichev, V. Carbohydrate Source can Influence the Efficiency of Somatic Embryogenesis in Seedless Grapes (Vitis Vinifera L.). Biotechnol. Biotechnol. Equip. 2005, 19, 62–66. [Google Scholar] [CrossRef]
- Yadollahi, A.; Abdollahi, M.R.; Moieni, A.; Danaee, M. Effects of carbon source, polyethylene glycol and abscisic acid on secondary embryo induction and maturation in rapeseed (Brassica napus L.) microspore-derived embryos. Acta Physiol. plant 2011, 33, 1905–1912. [Google Scholar] [CrossRef]
- Hesami, M.; Naderi, R.; Tohidfar, M.; Yoosefzadeh-Najafabadi, M. Application of Adaptive Neuro-Fuzzy Inference System-Non-dominated Sorting Genetic Algorithm-II (ANFIS-NSGAII) for Modeling and Optimizing Somatic Embryogenesis of Chrysanthemum. Front. plant Sci. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Shukla, M.R.; Singh, A.S.; Piunno, K.; Saxena, P.K.; Jones, A.M.P. Application of 3D printing to prototype and develop novel plant tissue culture systems. plant Methods 2017, 13, 6. [Google Scholar] [CrossRef]
- Chen, J.-R.; Wu, L.; Hu, B.-W.; Yi, X.; Liu, R.; Deng, Z.-N.; Xiong, X.-Y. The Influence of Plant Growth Regulators and Light Quality on Somatic Embryogenesis in China Rose (Rosa chinensis Jacq.). J. plant Growth Regul. 2014, 33, 295–304. [Google Scholar] [CrossRef]
- Li, H.; Xu, Z.; Tang, C. Effect of light-emitting diodes on growth and morphogenesis of upland cotton (Gossypium hirsutum L.) plantlets in vitro. plant Cell Tissue Organ. Cult. 2010, 103, 155–163. [Google Scholar] [CrossRef]
- Miler, N.; Kulus, D.; Woźny, A.; Rymarz, D.; Hajzer, M.; Wierzbowski, K.; Nelke, R.; Szeffs, L. Application of wide-spectrum light-emitting diodes in micropropagation of popular ornamental plant species: A study on plant quality and cost reduction. Vitr. Cell. Dev. Biol. Plant 2019, 55, 99–108. [Google Scholar] [CrossRef]
- D’Onofrio, C.; Morini, S.; Bellocchi, G. Effect of light quality on somatic embryogenesis of quince leaves. plant Cell Tissue Organ. Cult. 1998, 53, 91–98. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. plant 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Hesami, M.; Jones, A.M.P. Modeling and optimizing callus growth and development in Cannabis sativa using random forest and support vector machine in combination with a genetic algorithm. Appl. Microbiol. Biotechnol. 2021, 105, in press. [Google Scholar]
- Driver, J.A.; Kuniyuki, A.H. In vitro propagation of Paradox walnut rootstock. HortScience 1984, 19, 507–509. [Google Scholar]
- Hesami, M.; Naderi, R.; Tohidfar, M. Modeling and Optimizing in vitro Sterilization of Chrysanthemum via Multilayer Perceptron-Non-dominated Sorting Genetic Algorithm-II (MLP-NSGAII). Front. plant Sci. 2019, 10, 282. [Google Scholar] [CrossRef]
- Akin, M.; Eyduran, S.P.; Eyduran, E.; Reed, B.M. Analysis of macro nutrient related growth responses using multivariate adaptive regression splines. plant Cell Tissue Organ. Cult. 2020, 140, 661–670. [Google Scholar] [CrossRef]
- García-Pérez, P.; Lozano-Milo, E.; Landín, M.; Gallego, P.P. Machine Learning Technology Reveals the Concealed Interactions of Phytohormones on Medicinal Plant In Vitro Organogenesis. Biomolecules 2020, 10, 746. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Deng, D.; Dai, W.; Li, J.; Jin, X. Optimization of culture conditions for differentiation of melon based on artificial neural network and genetic algorithm. Sci. Rep. 2020, 10, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Hesami, M.; Naderi, R.; Tohidfar, M. Modeling and Optimizing Medium Composition for Shoot Regeneration of Chrysanthemum via Radial Basis Function-Non-dominated Sorting Genetic Algorithm-II (RBF-NSGAII). Sci. Rep. 2019, 9, 18237. [Google Scholar] [CrossRef]
- Hesami, M.; Condori-Apfata, J.A.; Valderrama Valencia, M.; Mohammadi, M. Application of Artificial Neural Network for Modeling and Studying in vitro Genotype-Independent Shoot Regeneration in Wheat. Appl. Sci. 2020, 10, 5370. [Google Scholar] [CrossRef]
- Munasinghe, S.P.; Somaratne, S.; Weerakoon, S.R.; Ranasinghe, C. Prediction of chemical composition for callus production in Gyrinops walla Gaetner through machine learning. Inf. Process. Agric. 2020, 7, 1–12. [Google Scholar] [CrossRef]
- Niazian, M.; Shariatpanahi, M.E.; Abdipour, M.; Oroojloo, M. Modeling callus induction and regeneration in an anther culture of tomato (Lycopersicon esculentum L.) using image processing and artificial neural network method. Protoplasma 2019, 256, 1317–1332. [Google Scholar] [CrossRef]
- Hesami, M.; Naderi, R.; Tohidfar, M.; Yoosefzadeh-Najafabadi, M. Development of support vector machine-based model and comparative analysis with artificial neural network for modeling the plant tissue culture procedures: Effect of plant growth regulators on somatic embryogenesis of chrysanthemum, as a case study. plant Methods 2020, 16, 112. [Google Scholar] [CrossRef] [PubMed]
- Salehi, M.; Farhadi, S.; Moieni, A.; Safaie, N.; Hesami, M. A hybrid model based on general regression neural network and fruit fly optimization algorithm for forecasting and optimizing paclitaxel biosynthesis in Corylus avellana cell culture. plant Methods 2021, 17, 13. [Google Scholar] [CrossRef] [PubMed]
- Farhadi, S.; Salehi, M.; Moieni, A.; Safaie, N.; Sabet, M.S. Modeling of paclitaxel biosynthesis elicitation in Corylus avellana cell culture using adaptive neuro-fuzzy inference system-genetic algorithm (ANFIS-GA) and multiple regression methods. PLoS ONE 2020, 15, e0237478. [Google Scholar] [CrossRef]
- Salehi, M.; Farhadi, S.; Moieni, A.; Safaie, N.; Ahmadi, H. Mathematical Modeling of Growth and Paclitaxel Biosynthesis in Corylus avellana Cell Culture Responding to Fungal Elicitors Using Multilayer Perceptron-Genetic Algorithm. Front. plant Sci 2020, 11, 1148. [Google Scholar] [CrossRef] [PubMed]
- Hesami, M.; Alizadeh, M.; Naderi, R.; Tohidfar, M. Forecasting and optimizing Agrobacterium-mediated genetic transformation via ensemble model- fruit fly optimization algorithm: A data mining approach using chrysanthemum databases. PLoS ONE 2020, 15, e0239901. [Google Scholar] [CrossRef] [PubMed]
- Bidabadi, S.S.; Jain, S.M. Cellular, Molecular, and Physiological Aspects of in vitro Plant Regeneration. Plants 2020, 9, 702. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, D.; Rai, K.K.; Rai, S.K.; Rai, S.P. An improved thin cell layer culture system for efficient clonal propagation and in vitro withanolide production in a medicinal plant Withania coagulans Dunal. Ind. Crop. Prod. 2018, 119, 172–182. [Google Scholar] [CrossRef]
- Tu, H.-Y.; Zhang, A.-L.; Xiao, W.; Lin, Y.-R.; Shi, J.-H.; Wu, Y.-W.; Wu, S.-T.; Zhong, C.-H.; Mo, S.-X. Induction and identification of tetraploid Hedychium coronarium through thin cell layer culture. plant CellTissue Organ. Cult. 2018, 135, 395–406. [Google Scholar] [CrossRef]
- Monja-Mio, K.M.; Robert, M.L. Direct somatic embryogenesis of Agave fourcroydes Lem. through thin cell layer culture. Vitr. Cell. Dev. Biol. Plant 2013, 49, 541–549. [Google Scholar] [CrossRef]
- Vidal, N.; Sánchez, C. Use of bioreactor systems in the propagation of forest trees. Eng. Life Sci. 2019, 19, 896–915. [Google Scholar] [CrossRef] [PubMed]
- Wen, F.; Su, W.-P.; Zheng, H.; Yu, B.-C.; Ma, Z.-F.; Zhang, P.; Guo, W.-W. Plant regeneration via protoplast electrofusion in cassava. J. Integr. Agric. 2020, 19, 632–642. [Google Scholar] [CrossRef]
- Jones, A.M.P.; Shukla, M.R.; Biswas, G.C.G.; Saxena, P.K. Protoplast-to-plant regeneration of American elm (Ulmus americana). Protoplasma 2015, 252, 925–931. [Google Scholar] [CrossRef] [PubMed]
- Beard, K.M.; Boling, A.W.H.; Bargmann, B.O.R. Protoplast isolation, transient transformation, and flow-cytometric analysis of reporter-gene activation in Cannabis sativa L. Ind. Crop. Prod. 2021, 164, 113360. [Google Scholar] [CrossRef]
- Lazič, S. Izolacija Protoplastov Navadne Konoplje (Cannabis sativa L.). Ph.D. Thesis, University of Ljubljana, Ljubljana, Slovenia, 2020. [Google Scholar]
- Morimoto, S.; Tanaka, Y.; Sasaki, K.; Tanaka, H.; Fukamizu, T.; Shoyama, Y.; Shoyama, Y.; Taura, F. Identification and characterization of cannabinoids that induce cell death through mitochondrial permeability transition in Cannabis leaf cells. J. Biol. Chem. 2007, 282, 20739–20751. [Google Scholar] [CrossRef]
- Niazian, M.; Shariatpanahi, M.E. In vitro-based doubled haploid production: Recent improvements. Euphytica 2020, 216, 69. [Google Scholar] [CrossRef]
- Wang, B.; Zhu, L.; Zhao, B.; Zhao, Y.; Xie, Y.; Zheng, Z.; Li, Y.; Sun, J.; Wang, H. Development of a Haploid-Inducer Mediated Genome Editing System for Accelerating Maize Breeding. Mol. plant 2019, 12, 597–602. [Google Scholar] [CrossRef]
- Kelliher, T.; Starr, D.; Su, X.; Tang, G.; Chen, Z.; Carter, J.; Wittich, P.E.; Dong, S.; Green, J.; Burch, E.; et al. One-step genome editing of elite crop germplasm during haploid induction. Nat. Biotechnol. 2019, 37, 287–292. [Google Scholar] [CrossRef]
- Hesami, M.; Yoosefzadeh Najafabadi, M.; Adamek, K.; Torkamaneh, D.; Jones, A.M. Synergizing off-target predictions for in silico insights of CENH3 knockout in cannabis through CRISPR/CAS. Molecules 2021, 26, 2053. [Google Scholar] [CrossRef]
- Li, B.; Cui, G.; Shen, G.; Zhan, Z.; Huang, L.; Chen, J.; Qi, X. Targeted mutagenesis in the medicinal plant Salvia miltiorrhiza. Sci. Rep. 2017, 7, 43320. [Google Scholar] [CrossRef]
- Zhou, Z.; Tan, H.; Li, Q.; Chen, J.; Gao, S.; Wang, Y.; Chen, W.; Zhang, L. CRISPR/Cas9-mediated efficient targeted mutagenesis of RAS in Salvia miltiorrhiza. Phytochemistry 2018, 148, 63–70. [Google Scholar] [CrossRef]
- Feng, S.; Song, W.; Fu, R.; Zhang, H.; Xu, A.; Li, J. Application of the CRISPR/Cas9 system in Dioscorea zingiberensis. plant Cell Tissue Organ Cult. 2018, 135, 133–141. [Google Scholar] [CrossRef]
- Azizi, P.; Hanafi, M.M.; Sahebi, M.; Harikrishna, J.A.; Taheri, S.; Yassoralipour, A.; Nasehi, A. Epigenetic changes and their relationship to somaclonal variation: A need to monitor the micropropagation of plantation crops. Funct. plant Biol. 2020, 47, 508–523. [Google Scholar] [CrossRef]
- Rodriguez-Enriquez, J.; Dickinson, H.G.; Grant-Downton, R.T. MicroRNA misregulation: An overlooked factor generating somaclonal variation. Trends plant Sci. 2011, 16, 242–248. [Google Scholar] [CrossRef]
- Lata, H.; Chandra, S.; Techen, N.; Khan, I.A.; ElSohly, M.A. Assessment of the Genetic Stability of Micropropagated Plants of Cannabis sativa by ISSR Markers. Planta Med. 2010, 76, 97–100. [Google Scholar] [CrossRef]
- Chandra, S.; Lata, H.; Mehmedic, Z.; Khan, I.A.; ElSohly, M.A. Assessment of Cannabinoids Content in Micropropagated Plants of Cannabis sativa and Their Comparison with Conventionally Propagated Plants and Mother Plant during Developmental Stages of Growth. Planta Med. 2010, 76, 743–750. [Google Scholar] [CrossRef] [PubMed]
- Lata, H.; Chandra, S.; Techen, N.; Khan, I.A.; ElSohly, M.A. Molecular analysis of genetic fidelity in Cannabis sativa L. plants grown from synthetic (encapsulated) seeds following in vitro storage. Biotechnol. Lett. 2011, 33, 2503–2508. [Google Scholar] [CrossRef] [PubMed]
- Adamek, K.; Jones, A.M.P.; Torkamaneh, D. Accumulation of somatic mutations leads to genetic mosaicism in Cannabis. bioRxiv 2021. [Google Scholar] [CrossRef]
- Niazian, M.; Nalousi, A.M. Artificial polyploidy induction for improvement of ornamental and medicinal plants. plant Cell Tissue Organ. Cult. 2020, 142, 11–23. [Google Scholar] [CrossRef]
- Iannicelli, J.; Guariniello, J.; Tossi, V.E.; Regalado, J.J.; Di Ciaccio, L.; van Baren, C.M.; Pitta Álvarez, S.I.; Escandón, A.S. The “polyploid effect” in the breeding of aromatic and medicinal species. Sci. Hortic. 2020, 260, 108854. [Google Scholar] [CrossRef]
- Bagheri, M.; Mansouri, H. Effect of Induced Polyploidy on Some Biochemical Parameters in Cannabis sativa L. Appl. Biochem. Biotechnol. 2015, 175, 2366–2375. [Google Scholar] [CrossRef]
- Kurtz, L.E.; Brand, M.H.; Lubell-Brand, J.D. Production of Tetraploid and Triploid Hemp. HortScience 2020, 55, 1703–1707. [Google Scholar] [CrossRef]
- Parsons, J.L.; Martin, S.L.; James, T.; Golenia, G.; Boudko, E.A.; Hepworth, S.R. Polyploidization for the genetic improvement of Cannabis sativa. Front. plant Sci. 2019, 10, 476. [Google Scholar] [CrossRef] [PubMed]
- Mansouri, H.; Bagheri, M. Induction of Polyploidy and Its Effect on Cannabis sativa L. In Cannabis sativa L.-Botany and Biotechnology; Chandra, S., Lata, H., ElSohly, M., Eds.; Springer: Cham, Switzerland, 2017; pp. 365–383. [Google Scholar]
- Sattler, M.C.; Carvalho, C.R.; Clarindo, W.R. The polyploidy and its key role in plant breeding. Planta 2016, 243, 281–296. [Google Scholar] [CrossRef]
- Corneillie, S.; De Storme, N.; Van Acker, R.; Fangel, J.U.; De Bruyne, M.; De Rycke, R.; Geelen, D.; Willats, W.G.; Vanholme, B.; Boerjan, W. Polyploidy affects plant growth and alters cell wall composition. plant Physiol. 2019, 179, 74–87. [Google Scholar] [CrossRef] [PubMed]
- McLaren, J.; Swift, W.; Dillon, P.; Allsop, S. Cannabis potency and contamination: A review of the literature. Addiction 2008, 103, 1100–1109. [Google Scholar] [CrossRef]
- Lavania, U.C. Enhanced productivity of the essential oil in the artificial autopolyploid of vetiver (Vetiveria zizanioides L. Nash). Euphytica 1988, 38, 271–276. [Google Scholar] [CrossRef]
- Tabatabaei, I.; Dal Bosco, C.; Bednarska, M.; Ruf, S.; Meurer, J.; Bock, R. A highly efficient sulfadiazine selection system for the generation of transgenic plants and algae. plant Biotechnol. J. 2019, 17, 638–649. [Google Scholar] [CrossRef] [PubMed]
- Bielecka, M.; Kaminski, F.; Adams, I.; Poulson, H.; Sloan, R.; Li, Y.; Larson, T.R.; Winzer, T.; Graham, I.A. Targeted mutation of Δ12 and Δ15 desaturase genes in hemp produce major alterations in seed fatty acid composition including a high oleic hemp oil. plant Biotechnol. J. 2014, 12, 613–623. [Google Scholar] [CrossRef] [PubMed]
- Schachtsiek, J.; Hussain, T.; Azzouhri, K.; Kayser, O.; Stehle, F. Virus-induced gene silencing (VIGS) in Cannabis sativa L. plant Methods 2019, 15, 157. [Google Scholar] [CrossRef] [PubMed]
- Debernardi, J.M.; Tricoli, D.M.; Ercoli, M.F.; Hayta, S.; Ronald, P.; Palatnik, J.F.; Dubcovsky, J. A GRF–GIF chimeric protein improves the regeneration efficiency of transgenic plants. Nat. Biotechnol. 2020. [Google Scholar] [CrossRef]
- MacKinnon, L.; McDougall, G.; Aziz, N.; Millam, S. Progress towards transformation of fibre hemp. Annu. Rep. Scott. Crop. Res. Inst. 2000, 18, 84–86. [Google Scholar]
- Feeney, M.; Punja, Z.K. Tissue culture and Agrobacterium-mediated transformation of hemp (Cannabis sativa L.). Vitr. Cell. Dev. Biol. Plant 2003, 39, 578–585. [Google Scholar] [CrossRef]
- Wahby, I.; Caba, J.M.; Ligero, F. Agrobacterium infection of hemp (Cannabis sativa L.): Establishment of hairy root cultures. J. plant Interact. 2013, 8, 312–320. [Google Scholar] [CrossRef]
- Feeney, M.; Punja, Z.K. Hemp (Cannabis sativa L.). In Agrobacterium Protocols. Methods in Molecular Biology; Wang, K., Ed.; Springer: New York, NY, USA, 2015; Volume 1224, pp. 319–329. [Google Scholar]
- Deguchi, M.; Bogush, D.; Weeden, H.; Spuhler, Z.; Potlakayala, S.; Kondo, T.; Zhang, Z.J.; Rudrabhatla, S. Establishment and optimization of a hemp (Cannabis sativa L.) agroinfiltration system for gene expression and silencing studies. Sci. Rep. 2020, 10, 3504. [Google Scholar] [CrossRef] [PubMed]
- Sorokin, A.; Yadav, N.S.; Gaudet, D.; Kovalchuk, I. Transient expression of the β-glucuronidase gene in Cannabis sativa varieties. plant Signal. Behav. 2020, 15, 1780037. [Google Scholar] [CrossRef]
- Anand, A.; Wu, E.; Li, Z.; TeRonde, S.; Arling, M.; Lenderts, B.; Mutti, J.S.; Gordon-Kamm, W.; Jones, T.J.; Chilcoat, N.D. High efficiency Agrobacterium-mediated site-specific gene integration in maize utilizing the FLP-FRT recombination system. plant Biotechnol. J. 2019, 17, 1636–1645. [Google Scholar] [CrossRef]
- Karthik, S.; Pavan, G.; Manickavasagam, M. Nitric oxide donor regulates Agrobacterium-mediated genetic transformation efficiency in soybean [Glycine max (L.) Merrill]. plant Cell Tissue Organ. Cult. 2020, 141, 655–660. [Google Scholar] [CrossRef]
- Das Bhowmik, S.S.; Cheng, A.Y.; Long, H.; Tan, G.Z.H.; Hoang, T.M.L.; Karbaschi, M.R.; Williams, B.; Higgins, T.J.V.; Mundree, S.G. Robust Genetic Transformation System to Obtain Non-chimeric Transgenic Chickpea. Front. plant Sci. 2019, 10, 524. [Google Scholar] [CrossRef]
- Jha, P.; Ochatt, S.J.; Kumar, V. WUSCHEL: A master regulator in plant growth signaling. plant Cell Rep. 2020, 39, 431–444. [Google Scholar] [CrossRef]
- Jha, P.; Kumar, V. BABY BOOM (BBM): A candidate transcription factor gene in plant biotechnology. Biotechnol. Lett. 2018, 40, 1467–1475. [Google Scholar] [CrossRef]
- Luo, G.; Palmgren, M. GRF-GIF Chimeras Boost Plant Regeneration. Trends plant Sci. 2020. [Google Scholar] [CrossRef]
- Gordon-Kamm, B.; Sardesai, N.; Arling, M.; Lowe, K.; Hoerster, G.; Betts, S.; Jones, T. Using morphogenic genes to improve recovery and regeneration of transgenic plants. Plants 2019, 8, 38. [Google Scholar] [CrossRef] [PubMed]
- Tian, R.; Paul, P.; Joshi, S.; Perry, S.E. Genetic activity during early plant embryogenesis. Biochem. J. 2020, 477, 3743–3767. [Google Scholar] [CrossRef] [PubMed]
- Hoerster, G.; Wang, N.; Ryan, L.; Wu, E.; Anand, A.; McBride, K.; Lowe, K.; Jones, T.; Gordon-Kamm, B. Use of non-integrating Zm-Wus2 vectors to enhance maize transformation. Vitr. Cell. Dev. Biol. Plant 2020, 56, 265–279. [Google Scholar] [CrossRef]
- Szczygieł-Sommer, A.; Gaj, M.D. The miR396–GRF regulatory module controls the embryogenic response in Arabidopsis via an auxin-related pathway. Int. J. Mol. Sci. 2019, 20, 5221. [Google Scholar] [CrossRef] [PubMed]
- Hecht, V.; Vielle-Calzada, J.-P.; Hartog, M.V.; Schmidt, E.D.; Boutilier, K.; Grossniklaus, U.; de Vries, S.C. The Arabidopsis SOMATIC EMBRYOGENESIS RECEPTOR KINASE 1 gene is expressed in developing ovules and embryos and enhances embryogenic competence in culture. plant Physiol. 2001, 127, 803–816. [Google Scholar] [CrossRef]
- Pérez-Pascual, D.; Jiménez-Guillen, D.; Villanueva-Alonzo, H.; Souza-Perera, R.; Godoy-Hernández, G.; Zúñiga-Aguilar, J.J. Ectopic expression of the Coffea canephora SERK1 homolog-induced differential transcription of genes involved in auxin metabolism and in the developmental control of embryogenesis. Physiol. plant 2018, 163, 530–551. [Google Scholar] [CrossRef] [PubMed]
- Harding, E.W.; Tang, W.; Nichols, K.W.; Fernandez, D.E.; Perry, S.E. Expression and maintenance of embryogenic potential is enhanced through constitutive expression of AGAMOUS-Like 15. plant Physiol. 2003, 133, 653–663. [Google Scholar] [CrossRef]
- Yang, Z.; Li, C.; Wang, Y.; Zhang, C.; Wu, Z.; Zhang, X.; Liu, C.; Li, F. GhAGL15s, preferentially expressed during somatic embryogenesis, promote embryogenic callus formation in cotton (Gossypium hirsutum L.). Mol. Genet. Genom. 2014, 289, 873–883. [Google Scholar] [CrossRef]
- Thakare, D.; Tang, W.; Hill, K.; Perry, S.E. The MADS-domain transcriptional regulator AGAMOUS-LIKE15 promotes somatic embryo development in Arabidopsis and soybean. plant Physiol. 2008, 146, 1663–1672. [Google Scholar] [CrossRef]
- Karlova, R.; Boeren, S.; Russinova, E.; Aker, J.; Vervoort, J.; de Vries, S. The Arabidopsis somatic embryogenesis receptor-like kinase1 protein complex includes brassinosteroid-insensitive1. plant Cell 2006, 18, 626–638. [Google Scholar] [CrossRef] [PubMed]
- Hohmann, U.; Lau, K.; Hothorn, M. The structural basis of ligand perception and signal activation by receptor kinases. Annu. Rev. plant Biol. 2017, 68, 109–137. [Google Scholar] [CrossRef] [PubMed]
- Long, J.A.; Moan, E.I.; Medford, J.I.; Barton, M.K. A member of the KNOTTED class of homeodomain proteins encoded by the STM gene of Arabidopsis. Nature 1996, 379, 66–69. [Google Scholar] [CrossRef]
- Laux, T.; Mayer, K.; Berger, J.; Jurgens, G. The WUSCHEL gene is required for shoot and floral meristem integrity in Arabidopsis. Development 1996, 122, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Arroyo-Herrera, A.; Ku Gonzalez, A.; Canche Moo, R.; Quiroz-Figueroa, F.R.; Loyola-Vargas, V.M.; Rodriguez-Zapata, L.C.; Burgeff D’Hondt, C.; Suárez-Solís, V.M.; Castaño, E. Expression of WUSCHEL in Coffea canephora causes ectopic morphogenesis and increases somatic embryogenesis. plant Cell Tissue Organ. Cult. 2008, 94, 171–180. [Google Scholar] [CrossRef]
- Bouchabké-Coussa, O.; Obellianne, M.; Linderme, D.; Montes, E.; Maia-Grondard, A.; Vilaine, F.; Pannetier, C. Wuschel overexpression promotes somatic embryogenesis and induces organogenesis in cotton (Gossypium hirsutum L.) tissues cultured in vitro. plant Cell Rep. 2013, 32, 675–686. [Google Scholar] [CrossRef] [PubMed]
- Elhiti, M.; Tahir, M.; Gulden, R.H.; Khamiss, K.; Stasolla, C. Modulation of embryo-forming capacity in culture through the expression of Brassica genes involved in the regulation of the shoot apical meristem. J. Exp. Bot. 2010, 61, 4069–4085. [Google Scholar] [CrossRef]
- Zuo, J.; Niu, Q.W.; Frugis, G.; Chua, N.H. The WUSCHEL gene promotes vegetative-to-embryonic transition in Arabidopsis. plant J. 2002, 30, 349–359. [Google Scholar] [CrossRef]
- Boutilier, K.; Offringa, R.; Sharma, V.K.; Kieft, H.; Ouellet, T.; Zhang, L.; Hattori, J.; Liu, C.-M.; van Lammeren, A.A.; Miki, B.L. Ectopic expression of BABY BOOM triggers a conversion from vegetative to embryonic growth. plant Cell 2002, 14, 1737–1749. [Google Scholar] [CrossRef]
- Srinivasan, C.; Liu, Z.; Heidmann, I.; Supena, E.D.J.; Fukuoka, H.; Joosen, R.; Lambalk, J.; Angenent, G.; Scorza, R.; Custers, J.B.M.; et al. Heterologous expression of the BABY BOOM AP2/ERF transcription factor enhances the regeneration capacity of tobacco (Nicotiana tabacum L.). Planta 2006, 225, 341. [Google Scholar] [CrossRef] [PubMed]
- El Ouakfaoui, S.; Schnell, J.; Abdeen, A.; Colville, A.; Labbé, H.; Han, S.; Baum, B.; Laberge, S.; Miki, B. Control of somatic embryogenesis and embryo development by AP2 transcription factors. plant Mol. Biol. 2010, 74, 313–326. [Google Scholar] [CrossRef]
- Morcillo, F.; Gallard, A.; Pillot, M.; Jouannic, S.; Aberlenc-Bertossi, F.; Collin, M.; Verdeil, J.L.; Tregear, J.W. EgAP2-1, an AINTEGUMENTA-like (AIL) gene expressed in meristematic and proliferating tissues of embryos in oil palm. Planta 2007, 226, 1353–1362. [Google Scholar] [CrossRef] [PubMed]
- Florez, S.L.; Erwin, R.L.; Maximova, S.N.; Guiltinan, M.J.; Curtis, W.R. Enhanced somatic embryogenesis in Theobroma cacao using the homologous BABY BOOM transcription factor. BMC plant Biol. 2015, 15, 121. [Google Scholar] [CrossRef] [PubMed]
- Tsuwamoto, R.; Yokoi, S.; Takahata, Y. Arabidopsis EMBRYOMAKER encoding an AP2 domain transcription factor plays a key role in developmental change from vegetative to embryonic phase. plant Mol. Biol. 2010, 73, 481–492. [Google Scholar] [CrossRef]
- Waki, T.; Hiki, T.; Watanabe, R.; Hashimoto, T.; Nakajima, K. The Arabidopsis RWP-RK Protein RKD4 Triggers Gene Expression and Pattern Formation in Early Embryogenesis. Curr. Biol. 2011, 21, 1277–1281. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.; Palmer, T.M.; Lukowitz, W. The RWP-RK Factor GROUNDED Promotes Embryonic Polarity by Facilitating YODA MAP Kinase Signaling. Curr. Biol. 2011, 21, 1268–1276. [Google Scholar] [CrossRef] [PubMed]
- Mursyanti, E.; Purwantoro, A.; Moeljopawiro, S.; Semiarti, E. Induction of somatic embryogenesis through overexpression of ATRKD4 genes in Phalaenopsis “Sogo Vivien”. Indones. J. Biotechnol. 2015, 20, 42–53. [Google Scholar] [CrossRef]
- Lotan, T.; Ohto, M.-a.; Yee, K.M.; West, M.A.L.; Lo, R.; Kwong, R.W.; Yamagishi, K.; Fischer, R.L.; Goldberg, R.B.; Harada, J.J. Arabidopsis LEAFY COTYLEDON1 Is Sufficient to Induce Embryo Development in Vegetative Cells. Cell 1998, 93, 1195–1205. [Google Scholar] [CrossRef]
- Zhu, S.-P.; Wang, J.; Ye, J.-L.; Zhu, A.-D.; Guo, W.-W.; Deng, X.-X. Isolation and characterization of LEAFY COTYLEDON 1-LIKE gene related to embryogenic competence in Citrus sinensis. plant Cell Tissue Organ. Cult. 2014, 119, 1–13. [Google Scholar] [CrossRef]
- Uddenberg, D.; Abrahamsson, M.; von Arnold, S. Overexpression of PaHAP3A stimulates differentiation of ectopic embryos from maturing somatic embryos of Norway spruce. Tree Genet. Genomes 2016, 12, 18. [Google Scholar] [CrossRef]
- Gazzarrini, S.; Tsuchiya, Y.; Lumba, S.; Okamoto, M.; McCourt, P. The Transcription Factor FUSCA3 Controls Developmental Timing in Arabidopsis through the Hormones Gibberellin and Abscisic Acid. Dev. Cell 2004, 7, 373–385. [Google Scholar] [CrossRef] [PubMed]
- Stone, S.L.; Kwong, L.W.; Yee, K.M.; Pelletier, J.; Lepiniec, L.; Fischer, R.L.; Goldberg, R.B.; Harada, J.J. LEAFY COTYLEDON2 encodes a B3 domain transcription factor that induces embryo development. Proc. Natl. Acad. Sci. USA 2001, 98, 11806–11811. [Google Scholar] [CrossRef] [PubMed]
- Santos Mendoza, M.; Dubreucq, B.; Miquel, M.; Caboche, M.; Lepiniec, L. LEAFY COTYLEDON 2 activation is sufficient to trigger the accumulation of oil and seed specific mRNAs in Arabidopsis leaves. FEBS Lett. 2005, 579, 4666–4670. [Google Scholar] [CrossRef] [PubMed]
- Gallois, J.-L.; Woodward, C.; Reddy, G.V.; Sablowski, R. Combined SHOOT MERISTEMLESS and WUSCHEL trigger ectopic organogenesis in Arabidopsis. Development 2002, 129, 3207–3217. [Google Scholar] [CrossRef] [PubMed]
- Gallois, J.-L.; Nora, F.R.; Mizukami, Y.; Sablowski, R. WUSCHEL induces shoot stem cell activity and developmental plasticity in the root meristem. Genes Dev. 2004, 18, 375–380. [Google Scholar] [CrossRef]
- Rashid, S.Z.; Yamaji, N.; Kyo, M. Shoot formation from root tip region: A developmental alteration by WUS in transgenic tobacco. plant Cell Rep. 2007, 26, 1449–1455. [Google Scholar] [CrossRef]
- Luo, K.; Zheng, X.; Chen, Y.; Xiao, Y.; Zhao, D.; McAvoy, R.; Pei, Y.; Li, Y. The maize Knotted1 gene is an effective positive selectable marker gene for Agrobacterium-mediated tobacco transformation. plant Cell Rep. 2006, 25, 403–409. [Google Scholar] [CrossRef]
- Nishimura, A.; Tamaoki, M.; Sakamoto, T.; Matsuoka, M. Over-expression of tobacco knotted 1-type class1 homeobox genes alters various leaf morphology. plant Cell Physiol. 2000, 41, 583–590. [Google Scholar] [CrossRef]
- Daimon, Y.; Takabe, K.; Tasaka, M. The CUP-SHAPED COTYLEDON genes promote adventitious shoot formation on calli. plant Cell Physiol. 2003, 44, 113–121. [Google Scholar] [CrossRef]
- Banno, H.; Ikeda, Y.; Niu, Q.-W.; Chua, N.-H. Overexpression of Arabidopsis ESR1 induces initiation of shoot regeneration. plant Cell 2001, 13, 2609–2618. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, Y.; Banno, H.; Niu, Q.-W.; Howell, S.H.; Chua, N.-H. The ENHANCER OF SHOOT REGENERATION 2 gene in Arabidopsis regulates CUP-SHAPED COTYLEDON 1 at the transcriptional level and controls cotyledon development. plant Cell Physiol. 2006, 47, 1443–1456. [Google Scholar] [CrossRef] [PubMed]
- Ckurshumova, W.; Smirnova, T.; Marcos, D.; Zayed, Y.; Berleth, T. Irrepressible MONOPTEROS/ARF 5 promotes de novo shoot formation. New Phytol. 2014, 204, 556–566. [Google Scholar] [CrossRef] [PubMed]
- Wójcikowska, B.; Jaskóła, K.; Gąsiorek, P.; Meus, M.; Nowak, K.; Gaj, M.D. LEAFY COTYLEDON2 (LEC2) promotes embryogenic induction in somatic tissues of Arabidopsis, via YUCCA-mediated auxin biosynthesis. Planta 2013, 238, 425–440. [Google Scholar] [CrossRef]
- Deng, W.; Luo, K.; Li, Z.; Yang, Y. A novel method for induction of plant regeneration via somatic embryogenesis. plant Sci. 2009, 177, 43–48. [Google Scholar] [CrossRef]
- Lowe, K.; Wu, E.; Wang, N.; Hoerster, G.; Hastings, C.; Cho, M.-J.; Scelonge, C.; Lenderts, B.; Chamberlin, M.; Cushatt, J. Morphogenic regulators Baby boom and Wuschel improve monocot transformation. plant Cell 2016, 28, 1998–2015. [Google Scholar] [CrossRef]
- Mookkan, M.; Nelson-Vasilchik, K.; Hague, J.; Zhang, Z.J.; Kausch, A.P. Selectable marker independent transformation of recalcitrant maize inbred B73 and sorghum P898012 mediated by morphogenic regulators BABY BOOM and WUSCHEL2. plant Cell Rep. 2017, 36, 1477–1491. [Google Scholar] [CrossRef] [PubMed]
- Heidmann, I.; de Lange, B.; Lambalk, J.; Angenent, G.C.; Boutilier, K. Efficient sweet pepper transformation mediated by the BABY BOOM transcription factor. plant Cell Rep. 2011, 30, 1107–1115. [Google Scholar] [CrossRef] [PubMed]
- Lutz, K.A.; Martin, C.; Khairzada, S.; Maliga, P. Steroid-inducible BABY BOOM system for development of fertile Arabidopsis thaliana plants after prolonged tissue culture. plant Cell Rep. 2015, 34, 1849–1856. [Google Scholar] [CrossRef]
- Wang, X.; Niu, Q.-W.; Teng, C.; Li, C.; Mu, J.; Chua, N.-H.; Zuo, J. Overexpression of PGA37/MYB118 and MYB115 promotes vegetative-to-embryonic transition in Arabidopsis. Cell Res. 2009, 19, 224–235. [Google Scholar] [CrossRef]
- Shires, M.E.; Florez, S.L.; Lai, T.S.; Curtis, W.R. Inducible somatic embryogenesis in Theobroma cacao achieved using the DEX-activatable transcription factor-glucocorticoid receptor fusion. Biotechnol. Lett. 2017, 39, 1747–1755. [Google Scholar] [CrossRef]
- Kononov, M.E.; Bassuner, B.; Gelvin, S.B. Integration of T-DNA binary vector ‘backbone’sequences into the tobacco genome: Evidence for multiple complex patterns of integration. plant J. 1997, 11, 945–957. [Google Scholar] [CrossRef]
- Hanson, B.; Engler, D.; Moy, Y.; Newman, B.; Ralston, E.; Gutterson, N. A simple method to enrich an Agrobacterium-transformed population for plants containing only T-DNA sequences. plant J. 1999, 19, 727–734. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-R.; Lee, J.; Jun, S.-H.; Park, S.; Kang, H.-G.; Kwon, S.; An, G. Transgene structures in T-DNA-inserted rice plants. plant Mol. Biol. 2003, 52, 761–773. [Google Scholar] [CrossRef] [PubMed]
- Podevin, N.; De Buck, S.; De Wilde, C.; Depicker, A. Insights into recognition of the T-DNA border repeats as termination sites for T-strand synthesis by Agrobacterium tumefaciens. Transgenic Res. 2006, 15, 557. [Google Scholar] [CrossRef] [PubMed]
- Richael, C.M.; Kalyaeva, M.; Chretien, R.C.; Yan, H.; Adimulam, S.; Stivison, A.; Weeks, J.T.; Rommens, C.M. Cytokinin vectors mediate marker-free and backbone-free plant transformation. Transgenic Res. 2008, 17, 905–917. [Google Scholar] [CrossRef] [PubMed]
- Lowe, K.; La Rota, M.; Hoerster, G.; Hastings, C.; Wang, N.; Chamberlin, M.; Wu, E.; Jones, T.; Gordon-Kamm, W. Rapid genotype “independent” Zea mays L. (maize) transformation via direct somatic embryogenesis. Vitr. Cell. Dev. Biol. Plant 2018, 54, 240–252. [Google Scholar] [CrossRef] [PubMed]
- Kong, J.; Martin-Ortigosa, S.; Finer, J.; Orchard, N.; Gunadi, A.; Batts, L.A.; Thakare, D.; Rush, B.; Schmitz, O.; Stuiver, M.; et al. Overexpression of the Transcription Factor GROWTH-REGULATING FACTOR5 Improves Transformation of Dicot and Monocot Species. Front. plant Sci. 2020, 11, 572319. [Google Scholar] [CrossRef]
- Hills, M.J.; Hall, L.; Arnison, P.G.; Good, A.G. Genetic use restriction technologies (GURTs): Strategies to impede transgene movement. Trends plant Sci. 2007, 12, 177–183. [Google Scholar] [CrossRef]
- Daniell, H. Molecular strategies for gene containment in transgenic crops. Nat. Biotechnol. 2002, 20, 581–586. [Google Scholar] [CrossRef]
- Linder, C.R.; Taha, I.; Rieseberg, L.H.; Seiler, G.J.; Snow, A.A. Long-term introgression of crop genes into wild sunflower populations. Theor. Appl. Genet. 1998, 96, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Schernthaner, J.P.; Fabijanski, S.F.; Arnison, P.G.; Racicot, M.; Robert, L.S. Control of seed germination in transgenic plants based on the segregation of a two-component genetic system. Proc. Natl. Acad. Sci. USA 2003, 100, 6855. [Google Scholar] [CrossRef]
- Hüsken, A.; Prescher, S.; Schiemann, J. Evaluating biological containment strategies for pollen-mediated gene flow. Environ. Biosaf. Res. 2010, 9, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Bicknell, R.A.; Koltunow, A.M. Understanding Apomixis: Recent Advances and Remaining Conundrums. plant Cell 2004, 16, S228. [Google Scholar] [CrossRef] [PubMed]
- Haider, N.; Allainguillaume, J.; Wilkinson, M.J. Spontaneous capture of oilseed rape (Brassica napus) chloroplasts by wild B. rapa: Implications for the use of chloroplast transformation for biocontainment. Curr. Genet. 2009, 55, 139–150. [Google Scholar] [CrossRef]
- Lu, B.-R. Editorial. Transgene containment by molecular means—is it possible and cost effective. Environ. Biosaf. Res. 2003, 2, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Wang, X.; Tang, Q.; Wang, Z. Theoretical Basis of Gene Splitting Technique and Its Application in the Control of Transgene Flow. Agric. Biotechnol. 2015, 4, 1–4. [Google Scholar]
- Kelloniemi, J.; Mäkinen, K.; Valkonen, J.P.T. Three heterologous proteins simultaneously expressed from a chimeric potyvirus: Infectivity, stability and the correlation of genome and virion lengths. Virus Res. 2008, 135, 282–291. [Google Scholar] [CrossRef]
- Saurabh, S.; Vidyarthi, A.S.; Prasad, D. RNA interference: Concept to reality in crop improvement. Planta 2014, 239, 543–564. [Google Scholar] [CrossRef]
- Lv, J.; Yu, K.; Wei, J.; Gui, H.; Liu, C.; Liang, D.; Wang, Y.; Zhou, H.; Carlin, R.; Rich, R.; et al. Generation of paternal haploids in wheat by genome editing of the centromeric histone CENH3. Nat. Biotechnol. 2020. [Google Scholar] [CrossRef]
- Jacinto, F.V.; Link, W.; Ferreira, B.I. CRISPR/Cas9-mediated genome editing: From basic research to translational medicine. J. Cell. Mol. Med. 2020, 24, 3766–3778. [Google Scholar] [CrossRef]
- Hurgobin, B.; Tamiru-Oli, M.; Welling, M.T.; Doblin, M.S.; Bacic, A.; Whelan, J.; Lewsey, M.G. Recent advances in Cannabis sativa genomics research. New Phytol. 2020. [Google Scholar] [CrossRef]
- van Bakel, H.; Stout, J.M.; Cote, A.G.; Tallon, C.M.; Sharpe, A.G.; Hughes, T.R.; Page, J.E. The draft genome and transcriptome of Cannabis sativa. Genome Biol. 2011, 12, R102. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Cradick, T.J.; Brown, M.T.; Deshmukh, H.; Ranjan, P.; Sarode, N.; Wile, B.M.; Vertino, P.M.; Stewart, F.J.; Bao, G. CRISPR/Cas9 systems have off-target activity with insertions or deletions between target DNA and guide RNA sequences. Nucleic Acids Res. 2014, 42, 7473–7485. [Google Scholar] [CrossRef] [PubMed]
- Gerashchenkov, G.A.; Rozhnova, N.A.; Kuluev, B.R.; Kiryanova, O.Y.; Gumerova, G.R.; Knyazev, A.V.; Vershinina, Z.R.; Mikhailova, E.V.; Chemeris, D.A.; Matniyazov, R.T.; et al. Design of Guide RNA for CRISPR/Cas Plant Genome Editing. Mol. Biol. 2020, 54, 24–42. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, X.; Cheng, L.; Luo, Y. An overview and metanalysis of machine and deep learning-based CRISPR gRNA design tools. RNA Biol. 2020, 17, 13–22. [Google Scholar] [CrossRef]
- Hwang, G.-H.; Park, J.; Lim, K.; Kim, S.; Yu, J.; Yu, E.; Kim, S.-T.; Eils, R.; Kim, J.-S.; Bae, S. Web-based design and analysis tools for CRISPR base editing. BMC Bioinform. 2018, 19, 542. [Google Scholar] [CrossRef] [PubMed]
- Bae, S.; Park, J.; Kim, J.-S. Cas-OFFinder: A fast and versatile algorithm that searches for potential off-target sites of Cas9 RNA-guided endonucleases. Bioinformatics 2014, 30, 1473–1475. [Google Scholar] [CrossRef]
- Park, J.; Bae, S.; Kim, J.-S. Cas-Designer: A web-based tool for choice of CRISPR-Cas9 target sites. Bioinformatics 2015, 31, 4014–4016. [Google Scholar] [CrossRef]
- Bae, S.; Kweon, J.; Kim, H.S.; Kim, J.-S. Microhomology-based choice of Cas9 nuclease target sites. Nat. Methods 2014, 11, 705–706. [Google Scholar] [CrossRef]
- Haeussler, M.; Schönig, K.; Eckert, H.; Eschstruth, A.; Mianné, J.; Renaud, J.-B.; Schneider-Maunoury, S.; Shkumatava, A.; Teboul, L.; Kent, J.; et al. Evaluation of off-target and on-target scoring algorithms and integration into the guide RNA selection tool CRISPOR. Genome Biol. 2016, 17, 148. [Google Scholar] [CrossRef]
- Concordet, J.-P.; Haeussler, M. CRISPOR: Intuitive guide selection for CRISPR/Cas9 genome editing experiments and screens. Nucleic Acids Res. 2018, 46, W242–W245. [Google Scholar] [CrossRef] [PubMed]
- Stemmer, M.; Thumberger, T.; del Sol Keyer, M.; Wittbrodt, J.; Mateo, J.L. CCTop: An Intuitive, Flexible and Reliable CRISPR/Cas9 Target Prediction Tool. PLoS ONE 2015, 10, e0124633. [Google Scholar] [CrossRef] [PubMed]
- Labun, K.; Montague, T.G.; Gagnon, J.A.; Thyme, S.B.; Valen, E. CHOPCHOP v2: A web tool for the next generation of CRISPR genome engineering. Nucleic Acids Res. 2016, 44, W272–W276. [Google Scholar] [CrossRef] [PubMed]
- Oliveros, J.C.; Franch, M.; Tabas-Madrid, D.; San-León, D.; Montoliu, L.; Cubas, P.; Pazos, F. Breaking-Cas—interactive design of guide RNAs for CRISPR-Cas experiments for ENSEMBL genomes. Nucleic Acids Res. 2016, 44, W267–W271. [Google Scholar] [CrossRef] [PubMed]
- Svitashev, S.; Young, J.K.; Schwartz, C.; Gao, H.; Falco, S.C.; Cigan, A.M. Targeted Mutagenesis, Precise Gene Editing, and Site-Specific Gene Insertion in Maize Using Cas9 and Guide RNA. plant Physiol. 2015, 169, 931. [Google Scholar] [CrossRef]
- Li, C.; Zhang, R.; Meng, X.; Chen, S.; Zong, Y.; Lu, C.; Qiu, J.-L.; Chen, Y.-H.; Li, J.; Gao, C. Targeted, random mutagenesis of plant genes with dual cytosine and adenine base editors. Nat. Biotechnol. 2020, 38, 875–882. [Google Scholar] [CrossRef]
- Hua, K.; Tao, X.; Zhu, J.-K. Expanding the base editing scope in rice by using Cas9 variants. plant Biotechnol. J. 2019, 17, 499–504. [Google Scholar] [CrossRef] [PubMed]
- Anzalone, A.V.; Randolph, P.B.; Davis, J.R.; Sousa, A.A.; Koblan, L.W.; Levy, J.M.; Chen, P.J.; Wilson, C.; Newby, G.A.; Raguram, A.; et al. Search-and-replace genome editing without double-strand breaks or donor DNA. Nature 2019, 576, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Sukegawa, S.; Saika, H.; Toki, S. Plant genome editing: Ever more precise and wide reaching. plant J. 2021, 106, 1–11. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Secondary Metabolite | Structure | Medicinal Effects | References |
|---|---|---|---|
| Tetrahydrocannabinol |  | Anti-inflammatory, antispastic, analgesic, antineoplastic, antiemetic activity, antipruritic agent, bronchodilator | Maayah et al. [12] and Workman et al. [13] |
| Cannabidiol |  | Anticonvulsant, antipsychotic, analgésic and anti-inflammatory, neuroprotection, antibacterial, antiemetic, anxiolytic, immunomodulator, antidepressant, cytotoxic for some cancer cell lines | Maayah et al. [12], Cassano et al. [14], and Alves et al. [15] |
| Cannabinol |  | Anti-inflammatory, antibacterial, anticonvulsant | Maayah et al. [12] and Alves et al. [15] |
| Cannabigerol |  | Analgesic, antifungal, antibacterial, antitumor activity, decreasing intraocular pressure | Cassano et al. [14] and Alves et al. [15] |
| Tetrahydrocannabivarin |  | Anti-inflammatory, anti-nociceptive, anticonvulsant | Maayah et al. [12], Workman et al. [13], and Cassano et al. [14] |
| Myrcene |  | Anti-inflammatory, anti-nociceptive, antioxidative | Baron [16], Nuutinen [17], and Hwang et al. [18] |
| β-caryophyllene |  | Anti-inflammatory, anti-convulsive, the astrocyte activation, inhibition of microglia, modulate nociception, neuroprotection, dopaminergic cell protection, addiction and alcohol consumption, feeding behavior, preventing alcohol-induced damage | Baron [16], Nuutinen [17], and Aly et al. [19] |
| Caryophyllene oxide |  | Anticancer, antioxidant, bactericide, and analgesic | Baron [16], Nuutinen [17], and Ciftci et al. [20] |
| Humulene |  | Treatment of depression, insomnia, nervousness, anxiety, delirium, and digestive disorders | Baron [16], Nuutinen [17], and Shah et al. [21] |
| α-Pinene |  | Anti-tumor, anti-allergic bronchodilator, anti-metastatic, antioxidant, anti-inflammatory, anxiolytic, and hypnotic | Baron [16], Nuutinen [17], and Kołodziejczyk et al. [22] |
| β -Pinene |  | Treatment of cancer, diabetes, atherosclerosis, and obesity | Baron [16], Nuutinen [17], and Guzmán-Gutiérrez et al. [23] |
| Linalool |  | Antioxidative, anti-nociceptive, neuroprotective, anticonvulsant, anti-inflammatory, sedative, anti-microbial, anti-depressant, hepatoprotective, anti-tumor | Baron [16], Nuutinen [17], and Jana et al. [24] |
| Limonene |  | Anti-tumor, anticancer, ameliorate depression, stress, inflammation, viral infections, and spasms | Baron [16], Nuutinen [17], and Srividya et al. [25] |
| Perillyl alcohol |  | Anti-inflammatory, anticancer, antioxidant, anti-tumor, nociceptive, antifungal, hepatoprotective, anti-parasitic | Baron [16], Nuutinen [17], and Faria et al. [26] |
| Terpinolene |  | Anti-inflammatory, antioxidant, anti-nociceptive | Baron [16], Nuutinen [17], and Zhao et al. [27] |
| γ-Terpinene |  | Anti-nociceptive and anti-inflammatory | Baron [16], Nuutinen [17], and Castro et al. [28] |
| α -Terpinene |  | Antioxidant and antibiotic | Baron [16], Nuutinen [17], and de Oliveira et al. [29] |
| Terpineol |  | Anticancer, spasmolytic, anti-tumorigenic, antibiotic, anti-inflammatory, anticonvulsant, treatment of spasms, neurological damages, pain, and asthma | Baron [16], Nuutinen [17], and Vieira et al. [30] |
| Geraniol |  | Treatment of depression, cancer, cardiac dysfunction, pain, colitis, neuropathy, atherosclerosis, allergic asthma, inflammation, tissue injuries, PD, and diabetes. | Baron [16], Nuutinen [17], and Lira et al. [31] |
| Nerolidol |  | Antioxidant, anti-inflammatory, anticancer, sedative, fungicide, anxiolytic, bactericide, anti-parasitic, antidepressant, and antinociceptive. | Baron [16], Nuutinen [17], and Barros Silva Soares de Souza et al. [32] |
| Borneol |  | DNA preserving, antipyretic, anti-inflammatory, neuroprotective, antioxidant, and anti-nociceptive. | Baron [16], Nuutinen [17], and Yang et al. [33] |
| α -Bisabolol |  | Anti-parasitic, anti-inflammatory, anti-nociceptive, anticancer, antibiotic, anti-tumor, and anti-apoptotic. | Baron [16], Nuutinen [17], and Xu et al. [34] |
| Bisabolenes |  | Anti-bacterial, anticancer, anti-convulsive, and anti-tumor. | Baron [16], Nuutinen [17], and Gogineni et al. [35] |
| β -elemene |  | Anti-inflammatory, treatment of cancer, liver fibrosis, atherosclerosis, and MS. | Baron [16], Nuutinen [17], and Tong et al. [36] |
| Fenchone |  | Antimicrobial, anticancer, and anti-tumor. | Baron [16], Nuutinen [17], and Müller et al. [37] |
| Pulegone |  | Antimicrobial, anticancer, and anti-tumor. | Baron [16], Nuutinen [17], and Yang et al. [33] |
| α -Phellandrene |  | Pro-apoptotic, antimicrobial, anti-inflammatory, anti-depressive, immunomodulatory, and anti-nociceptive. | Baron [16], Nuutinen [17], and de Christo Scherer et al. [38] |
| β -eudesmol |  | Anti-convulsant, anti-inflammatory, and anticancer. | Baron [16], Nuutinen [17], and Kotawong et al. [39] |
| Genotype(s) | Explant(s) | Morphogenetic Response(s) | Media, PGRs (mg/L), and Additives | Culture Conditions (Temperature, Light Intensity, etc.) | Outcomes and Descriptions | Reference |
|---|---|---|---|---|---|---|
| OSU | Roots derived from in vitro grown seedling | Cell suspension cultures | Gamborg’s medium (67-V), 2,4-D (1.5) + NAA (0.1) + IAA (1) + Kin (0.25) + casein hydrolysate (1) | Light at 26 °C, light condition: NR | The maximum callogenesis was observed in the media containing 0.1 mg/L NAA + 0.25 mg/L Kin + 1 mg/L casein hydrolysate. | Veliky and Genest [62] |
| C-71, TU-A | Leaves, hypocotyl, root, and female and male floral parts | Callogenesis | MS +2,4-D (1) + Kin (0.01–0.1) | Light at 26 °C, light condition: NR | The maximum callogenesis was observed in MS medium supplemented with 1 mg/L 2,4-D + 0.1 mg/L Kin. | Itokawa et al. [63] |
| C-150, C-152 | Bracts, calyx | Callogenesis | Miller’s medium + Murashige’s iron source + IAA (0.25, 1) + NAA (0.1, 0.25) + 2,4-D (0.2) + Kin (1, 1.5, 2) + casein hydrolysate (1) | 12 h photoperiod (~700 lx); temperature: NR | The maximum callogenesis in different cultivars and explants was observed in the media containing 0.5 mg/L NAA + 2 mg/L Kin. Although root formation was observed on the surface of the callus, it was inhibited by using 0.2 mg/L 2,4-D. | Hemphill et al. [49] |
| OSU | Leaf, roots, and stem | Callogenesis and cell suspension cultures | MS salts +B5 vitamins medium + 2,4-D (0–5), 2,4,5-T (0–5), NAA (0–5), kin (0–5), 2iP (0–5), and BAP (0–5) | Light at 26 °C, light condition: NR | Callogenesis in stem segments was observed in 0.5 mg/L 2,4-D and 0.1 mg/L BAP. 2,4,5-T and NAA could not produce calli in stem segments. Callogenesis in root segments was observed in 0.1–1 mg/L NAA and 5 mg/L kin. as well as 5 mg/L BAP and 1 mg/L NAA. 2,4,5-T and 2,4-D could not produce calli in root segments. Generally, the response of Cannabis explants to PGRS was significantly affected by the type of explant. The maximum cell masses in cell suspension culture were produced in 3 mg/L 2,4,5-T without subculture. | Loh et al. [50] |
| OSU | Different parts of seedling | Callogenesis and cell suspension cultures | MS salts +B5 vitamins medium +2,4-D (0.1) + Kin (0.5) | Light at 27 °C, light condition: NR | Six to eight weeks after culturing, callogenesis was obtained. The maximum cell masses in cell suspension culture were produced in 3 mg/L 2,4,5-T. | Hartsel et al. [64] |
| F56 and F77 | Apical and axillary buds | Shoot organogenesis and in vitro rooting | MS + IBA (0–20) + BAP (0.45) + 3% glucose + 1% sucrose + charcoal (0–2 g/L) | 27 ± 2 °C under 16 h photoperiod (360 µmol/m2/s) | The highest shoot regeneration was observed in 2 mg/L IBA + 0.45 mg/L BAP + 3% glucose + 1% sucrose. The maximum root regeneration was observed in 20 mg/L IBA + 2g/l charcoal. | Richez-Dumanois et al. [65] |
| NR | Leaf | Cell suspension cultures | B5 medium + 1 mg/L 2,4 -D + 0.5 mg/L KIN + 3% glucose | Darkness at 25 °C | A cell suspension culture of Cannabis was able to convert CBD to bound CBE and THC to CBC. | Braemer and Paris [51] |
| Sud Italian | Leaf, hypocotyl, cotyledon, and root | Callogenesis and shoot regeneration | MS salts +B5 vitamins medium +2,4-D (3–10) + BAP (0.01–1) | 27 ± 2 °C under 16 h photoperiod (360 µmol/m2/s) | Although all explants produced callus, the maximum callogenesis was observed in leaf and hypocotyl segments. The maximum shoot regeneration was obtained from hypocotyl segments; however, leaf explants could not produce shoots. | Mandolino and Ranalli [66] |
| Silesia, Juso-15, Novosadska, Fibrimon-24, and Fedrina-74 | Leaves, petioles, internodes, and axillary buds | Callogenesis, shoot regeneration, and in vitro rooting | MS + 2,4-D (2 and 4), DIC (2 and 3), NAA (0.5, 1 and 2), and Kin (1, 2, and 4) | 22 °C under a 16 h photoperiod (~2000 lx) | Callogenesis and shoot regeneration responses were varied based on different explants and genotypes. The highest callogenesis was obtained by petiole segments of cv. Fibrimon-24. The maximum indirect shoot regeneration was observed on a medium containing DIC. In vitro rooting was obtained from 1.0 mg/L IAA and 1.0 mg/L NAA. | Slusarkiewicz-Jarzina et al. [67] |
| Beniko and Bialobrzeskie | Stems, roots, and adventitious shoots | Direct organogenesis and indirect embryogenesis | Knapp’s medium + BAP (NR) + NAA (NR) + IAA (NR) | NR | After two weeks direct organogenesis was observed. Somatic embryos were also obtained from the medium containing NAA and BAP along with 500 mg/L activated charcoal. | Plawuszewski et al. [68] |
| Finola | Lateral buds | Shoot regeneration and in vitro rooting | Shoot regeneration: MS + TDZ (0.1–0.5) + NAA (0.05–0.3) Rooting: 1/2MS or MS + IBA (0.01–0.5) + NAA (0.01–0.25) | 25 °C under a 16 h photoperiod (~3000 lx) | The maximum shoot regeneration was observed in 0.35 mg/L TDZ + 0.3 mg/L NAA. The highest root formation was observed in MS + 0.2 mg/L IBA + 0.15 mg/L NAA. | Bing et al. [69] |
| Bialobrzeskie, Silesia, and Beniko | Cotyledons, stems, and roots | Callogenesis, shoot regeneration, and in vitro rooting | Knopp’s medium + Kin (1), BAP (0.2), NAA (0.03–0.05), IAA (2) | 24–26 °C under a 16 h photoperiod (light intensity: NR) | Callogenesis and shoot regeneration responses were varied based on different explants and genotypes. The highest callogenesis was obtained from 1 mg/L Kin and 0.05 NAA mg/L. The maximum indirect shoot regeneration was observed in a medium containing 0.2 mg/L BAP and 0.03 mg/L NAA. In vitro rooting was obtained from 2.0 mg/L IAA. | Wielgus et al. [70] |
| Changtu | Shoot tips | Shoot proliferation and in vitro rooting | Shoot regeneration: MS + BAP (1.0, 2.0, 5.0), Kin (1.0, 2.0, 5.0), TDZ (0.1, 0.2, 0.5), NAA (0.05, 0.1, 0.5) Rooting: 1/2MS, MS, B5 or NN + NAA (0.05, 0.25), IAA (0.05, 0.25), IBA (0.1, 0.5) | 25 ± 1 °C under a 16 h photoperiod (2500 lx) | The highest shoot proliferation was obtained from 0.2 mg/L 1TDZ and 0.1 NAA mg/L. The highest in vitro rooting was obtained from MS + 0.1 mg/L IBA + 0.05 mg/L NAA. | Wang et al. [71] |
| MX-1 | Nodal segments containing axillary buds | Shoot proliferation and in vitro rooting | Shoot regeneration: MS + BAP (0.5–9 μM), Kin (0.5–9 μM), TDZ (0.5–9 μM), GA (0.7 μM) In vitro rooting: 1/2MS + 500 mg/L activated charcoal + IAA (2.5, 5 μM), IBA (2.5, 5 μM), NAA (2.5, 5 μM) | 25 ± 2 °C under a 16 h photoperiod (52 µmol/m2/s) | The highest shoot proliferation was obtained from 0.5 μM TDZ. The highest in vitro rooting was obtained from 2.5 μM IBA. | Lata et al. [72] and Lata et al. [73] |
| MXE-1 | Leaf | Callogenesis, shoot organogenesis, and in vitro rooting | Callogenesis: MS + 1.0 μM TDZ + (0.5, 1.0, 1.5, and 2.0 μM) of IAA, NAA, IBA Shoot organogenesis: MS + (0.5, 1.0, 2.5, 5.0, and 10.0 μM) of BAP, Kin, TDZIn vitro rooting: 1/2MS + (0.5, 1.0, 2.5, 5.0, and 10.0 μM) of IAA, IBA, and NAA | 25 ± 2 °C under a 16 h photoperiod (52 µmol/m2/s) | The maximum callogenesis was obtained from 0.5 μM NAA + 1.0 μM TDZ. The highest shoot organogenesis was observed in 0.5 μM TDZ. The highest in vitro rooting was obtained from 2.5 μM IBA. | Lata et al. [74] |
| NR | Cotyledon and epicotyl | Indirect shoot organogenesis, and in vitro rooting | Indirect shoot organogenesis: MS +BAP (0.1, 0.2, 0.5, 1, 2, 3), IBA (0.5), TDZ (0.1, 0.2, 0.5, 1, 2, 3), IAA (0.5) In vitro rooting: MS + IBA (0.1, 0.2, 0.5, 1) + NAA (0.1, 0.2, 0.5, 1) | NR | The maximum callogenesis was obtained from cotyledon explants in MS medium supplemented with 3 mg/L TDZ + 0.5 mg/L IBA. The maximum shoot organogenesis was achieved from epicotyl segments in MS medium supplemented with 2 mg/L BAP + 0.5 mg/L IBA. | Movahedi et al. [75] |
| NR | Leaf and hypocotyl | Indirect shoot organogenesis, and in vitro rooting | Callogenesis and shoot regeneration: MS+ 2,4-D (0.1, 0.2, 0.5, 1), NAA (0.5, 1, 2, 3), BAP (0.5) In vitro rooting: MS+ (0.1, 0.2, 0.5, 1) of IBA and NAA | 25 °C under a 16 h photoperiod (light intensity: NR) | The maximum callogenesis was observed from leaf segments in 1 mg/L 2,4-D + 0.5 mg/L BAP. However, indirect organogenesis was only obtained from hypocotyl explants in the medium containing 0.1 mg/L 2,4-D + 0.5 mg/L BAP. Successful in vitro rooting was observed in all of the treatments. | Movahedi et al. [76] |
| NR | Leaf and hypocotyl | Callogenesis | Callogenesis and shoot regeneration: MS+ BAP (0.1, 0.2, 0.5, 1, 2, 3), TDZ (0.1, 0.2, 0.5, 1, 2, 3), IBA (0.5) | 25 °C under a 16 h photoperiod (light intensity: NR) | The maximum callogenesis was obtained from MS medium containing 0.5 mg/L IBA + 2 mg/L TDZ using leaf segments. Indirect shoot formation was observed on various concentrations of BAP in hypocotyl segments. | Movahedi et al. [77] |
| Mexican variety | Nodal segments containing axillary buds | Shoot proliferation and in vitro rooting | Shoot regeneration: MS + 500 mg/L activated charcoal + TDZ (0.05, 0.50, 1, 2, 3, 4, and 5 μM), mT (0.05, 0.50, 1, 2, 3, 4, and 5 μM) In vitro rooting: 1/2MS + 500 mg/L activated charcoal + IBA (0.05, 0.50, 1, 2, 3, 4, and 5 μM) | 25 ± 2 °C under a 16 h photoperiod (52 µmol/m2/s) | 2 μM mT resulted in the highest shoot regeneration and in vitro rooting. | Lata et al. [78] |
| Kunming, Neimeng 700, YM535, Anhui727, DaliS1, Heilongjiang698, Heilongjiang449, BM2 | Cotyledons | Shoot regeneration and in vitro rooting | Callogenesis: MS +BAP (4,6,8), ZT (0.5, 1, 1.5), TDZ (0.1, 0.2, 0.4), NAA (0.2, 0.4, 0.6) Shoot organogenesis: MS + TDZ (0.1, 0.2, 0.3, 0.4, 0.5), NAA (0.2, 0.4, 0.6) In vitro rooting: 1/2MS + IBA (0.2, 0.5, 1, 2) | 22 ± 2 °C under a 16 h photoperiod (36 µmol/m2/s) | While BA and ZT produced Soft, flaky, green and yellow callus, TDZ produced Hard, green and nodular callus. The maximum shoot regeneration was obtained from 0.4 mg/L TDZ + 0.2 mg/L NAA. The regenerated micro-shoots had a high vitrification rate and a low chance of survival in the rooting step when higher than 0.5 mg/L TDZ was used. Shoot regeneration responses were varied based on cotyledon age and genotypes. The juvenile cotyledon (2-day-old) showed the best regeneration potential. | Chaohua et al. [79] |
| 1KG2TF, S1525, H5458 | Immature and mature inflorescences | Shoot regeneration | Shoot organogenesis: MS + TDZ (0.1, 2, 5, 10) | 23 °C under a 16 h photoperiod (10–30 µmol/m2/s) | Shoot regeneration was observed in 1 and 10 μmol TDZ. MS+ 0.03% also activated charcoal+ 1.86 μmol kin+ 0.54 μmol NAA resulted in shoot multiplication and in vitro rooting. | Piunno et al. [80] |
| Bialobriezskie, Tygra, Fibrol, Monoica, USO-31 | Cotyledonary node, epicotyl with first node, hypocotyl, epicotyl with first and second node, shoot apical meristem, and shoot apex | Shoot regeneration | MS + 9.31 μg/L NAA + 0.23 mg/L BAP + mT (1–5), BAP9THP (1–5), PEO-IAA (10 μmol/l) | 19 °C under a 16 h photoperiod (56 µmol/m2/s) | Epicotyl with the first node resulted in the highest shoot regeneration. The maximum shoot regeneration was also observed in the medium containing BAP9THP. | Smýkalová et al. [81] |
| U91, GRC, U37, RTG, U82, U42, U22, U38, U31, and U61 | Leaf | Callogenesis | MS and DKW + NAA (0.5 μM), TDZ (0.5 and 1 μM) | 25 °C under a 16 h photoperiod (10–41 ± 4 µmol/m2/s) | Although 1.0 μM TDZ + 0.5 μM NAA produced callus in all genotypes, callogenesis was determined to be species-specific. | Monthony et al. [82] |
| E1, E4, and E40 of Epsilon 68 | Nodal segments containing axillary buds, shoot tips | Shoot regeneration and in vitro rooting | Shoot regeneration: MS+ BAP (0.5–2), TDZ (0.1–0.5), mT (0.1–1) In vitro rooting: 1/2 MS + (0.25, 0.5, 0.75) of IBA and IAA | 25 ± 1 °C under a 18 h photoperiod (60 µmol/m2/s) | The highest shoot regeneration was observed in the media containing 1–2 mg/L ZEARIB + 0.02 mg/L NAA. | Wróbel et al. [83] |
| Felina32, Ferimon, Fedora17, Finola, and USO31 | Leaves, hypocotyl, and cotyledon | Direct shoot regeneration | MS+ BAP (0.5, 1, 2), TDZ (0.4, 1), NAA (0.02, 0.2), IBA (0.5), 2,4-D (0.1), 4-CPPU (1.0), ZTRIB (1, 2), BAPRIB (1) | 22 ± 1 °C under a 16 h photoperiod (90.15 µmol/m2/s) | Cotyledon and leaf explants had poor shoot regeneration responses, while hypocotyl segments were the best explant for shoot regeneration. | Galán-Ávila et al. [84] |
| U82 and U91 | Inflorescences (single florets vs. pairs of florets) | Direct shoot regeneration | DKW + BAP (0.0, 0.01, 0.1, 1.0, and 10 μM) for both cultivars DKW + mT (0.0, 0.01, 0.1, 1.0, and 10 μM) for U91 cultivar | 25 °C under a 16 h photoperiod (50 µmol/m2/s) | Floral reversion was observed in the meristematic florets. These explants can be applied to improve regeneration frequency. Although the pairs of florets had a significant effect on the reversion rate and production of healthier plantlets, PGRs and cultivars had no remarkable impact on the reversion rate. | Monthony et al. [85] |
| Aida, Juani, Magda, Moniek, Octavia, and Pilar | Axillary buds | Shoot regeneration | MS, B5 with vitamins (Formula βA), and B5 without MS vitamins (Formula βH) + 2μM mT, 2μM IBA NAA, 2μM IBA | 25 ± 0.5 °C under a 18 h photoperiod (50 µmol/m2/s) | Both Formula β media resulted in a better response. Also, results showed that success was cultivar-dependent. | Codesido et al. [86] |
| MX-CBD-11 and MX-CBD-707 | Axillary buds | Shoot regeneration | MS + TDZ (0.011, 0.1, 0.11, 0.22, 0.44, 0.88, 1.76 mg/L), mT (0.012, 0.12, 0.24, 0.48, 0.5, 0.96, 1.93 mg/L), BAP (1, 2.5, 5 mg/L), IAA (0.1 mg/L) | 25 °C under a 16-h photoperiod (light intensity: NR) | The results showed that the type and concentration of PGRs and genotype had a significant effect on cannabis shoot regeneration. MS medium supplemented with 0.1 mg/L TDZ also resulted in the highest regeneration frequency in both genotypes. | Mubi et al. [87] |
| a high CBD and a high CBG | Axillary buds | Shoot regeneration | Shoot regeneration: Full- or half-strength MS + BAP (1.0, 2.0, 4.0 and 8.0 μM), TDZ (1.0, 2.0, 4.0 and 8.0 μM) In vitro rooting: Full- or half-strength MS + IBA (1.0, 2.0, 4.0 and 8.0 μM), NAA (1.0, 2.0, 4.0 and 8.0 μM) | 23 ± 1 °C under a 16 h photoperiod (50 µmol/m2/s) | Both full and half-strength MS + 4.0 μM BA resulted in the maximum shoot number and shoot length in both genotypes. The highest root formation was also obtained from both full and half-strength MS + 4.0 μM IBA or NAA. | Ioannidis et al. [88] |
| Hemp cultivars (Wife and Dinamed CBD) | Stem tips | Shoot proliferation | MS, MS + Mesos components, 2.5× MS with vitamins, MS with vitamins + added Mesos, MS with vitamins + added vitamins, MS with vitamins + added Mesos and vitamins; MS with vitamins + added Mesos and vitamins + NH4NO3 (0, 500, 1000, or 1500 mg/L) | 25 °C under a 18 h photoperiod (40 µmol/m2/s) | The maximum shoot multiplication, leaf lamina development, and shoot extension were observed in MS with vitamins + added Mesos and vitamins + 500 mg/L NH4NO3. 75% to 100% ex vitro rooting was also obtained in Rockwool. | Jessica et al. [89] |
| US Nursery Cherry 1 | Apical shoot tip and single node | Shoot proliferation | DKW without PGRs | 23 ± 2 °C under a 14 h photoperiod (25, 46, 85, 167 µmol/m2/s) in vessels with vented or non-vented closures | The maximum number of harvested shoot tips was observed in 46 µmol/m2/s in non-vented vessels. | Murphy and Adelberg [90] |
| BCN Power Plant, Safari Cake 747, CD13, and Blue Widow | Stem segments | Shoot growth and development | Safari Flower (SF) vegetative fertilizer solution + Sigma-Aldrich Canada + ethanesulfonic acid + 5-mM MES (2-(Nmorpholino) | 22 ± 3 °C under a 18 h photoperiod (50, 100, 150 µmol/m2/s) | The roles of Rockwool medium pH, cutting length, the moisture content in the vessels, basal wounding methods, the capacity of culture vessel gas exchange, and light intensity were studied. The percent of rooted plants was increased by using both 5- and 7-cm explant lengths compared to 3-cm explant length. Rooting was improved by increasing gas exchange. | Zarei et al. [91] |
| BA-1, BA-21, BA-41, BA-49, BA-61, BA-71 | Stem segments with two nodes | Callogenesis, shoot proliferation | MS, DKW, WPM, B5, BABI media + TDZ (0.5 μM), 2,4-D (10, 20, 30 μM) | 25 °C under a 16 h photoperiod (10–41 ± 4 µmol/m2/s) | The maximum shoot regeneration was observed in DKW + 0.5 μM. DKW+ 10 μM 2,4-D was the best treatment for callogenesis. | Page et al. [92] |
| Hemp cultivar (YUNMA7) | Immature embryo hypocotyls, true leaves, cotyledons and hypocotyls | Indirect shoot organogenesis | Callus induction medium: MS+ 1 mg/L Nicotinic acid + 1 mg/L Pyridoxine-HCl + 10 mg/L Thiamine-HCl + 0.1 g/L Myo-inositol + 3% Sucrose + 2.5 g/L Phytagel + 1 mg/L 2,4-D + 0.25 mg/L Kin + 100 mg/L Casein hydrolysate Regeneration medium: 1/2 strength MS + 1.5% Sucrose + 3.5 g/L Phytagel + 0.5 mg/L TDZ + 0.3 mg/L 6-BA + 0.2 mg/L NAA + 0.2 mg/L IAA Rooting medium: 1/2 strength MS + 1.5% Sucrose + 3.5 g/L Phytagel + 0.2 mg/L NAA + 0.5 mg/L IBA + 0.01 mg/L ZeaRIB | 26 °C under continuous light (50 µmol/m2/s) | Over 20% of the immature embryo hypocotyls developed embryogenic calli within 5 days, and the hypocotyls collected 15 days after anthesis (D15) produced more calli (at an average of 31.08%) compared to those collected earlier or later. Throughout the 4-week incubation, the induction frequencies of only 5.97% in true leaves, 7.65% in cotyledons, and 5.31% in hypocotyls were observed. After an additional 2 weeks, proliferating tissues were transferred to the regeneration medium and 6.12% of the D15 calli produced shoots, and less than 3% of the calli developed proliferated shoots from the other three explants. | Zhang et al. [93] |
| Genotype(s) | Explant | Protoplast Isolation Procedure | References |
|---|---|---|---|
| Cherry x Otto II: Sweetened | Mesophyll of young, not fully expanded leaves of in vitro grown plantlets | Enzymolysis solution composed of 0.3% w/v Macerozyme R-10, 20 mM MES (2-(N-morpholino) ethanesulfonic acid), 1.25% w/v Cellulase R-10, 0.4 M mannitol, 0.1% w/v bovine serum albumin, 10 mM calcium chloride, 20 mM potassium chloride, and 0.075% w/v Pectolyase Y23, adjusted to pH 5.7 and heated to 55 °C for 10 min | Beard et al. [141] |
| Finola | Etiolated hypocotyls and mesophyll of leaf | Enzyme solution composed of 0.4% Macerozyme R–10 and 1.5% Cellulase Onozuka R-10 | Lazič [142] |
| Mexican strain | Leaf cells | Digestion solution supplemented with 88 mM sucrose, 0.4 M mannitol, 1% (w/v) Cellulase Onozuka R-10, 0.1% (w/v) pectolyase Y-23, and 0.2% (w/v) Macerozyme R-10 at 30 °C for 4 h with gentle agitation | Morimoto et al. [143] |
| Genotype(s) | Applied Antimitotic Agent(s) | Polyploidy Induction Efficiency (%) | Survival Rate of Induced Polyploids (%) | Method of Confirmation | Outcomes and Remarks | References |
|---|---|---|---|---|---|---|
| Unspecified Iranian Cultivar | Colchicine (0.1–0.2% w/v) | 59.1 for 24 h 42.1 for 48 h | 73.33 for 24 h 63.33 for 48 h | Stomate size/density, leaf morphology, and flow cytometry | 0.2% colchicine was required to induce polyploidy. Polyploids exhibit wider leaves, larger stomata, and larger male flowers. No effect on cannabinoid production in male and female flowers was reported however female polyploid leaves demonstrated a significant increase in CBD concentration. | Mansouri and Bagheri [162] |
| THC Dominant Indica and Balanced THC/CBD Indica Dominant Hybrid (Canopy Growth Corp.) | Oryzalin (20–150 µM) | 66.7 | 37.5 | Stomate size/density, flow cytometry, and chromosome count | The THC dominant cultivar was only induced under the 40 µM treatment and unsuccessful treatments produced many mixoploids. The balanced cultivar was successfully induced between a range of 20–60 µM. Tetraploid flowers had increased CBD and CBDA content however overall cannabinoid and terpene concentrations were not significantly different. Polyploids exhibit larger leaf area, larger stomata, reduced rooting success, increased sugar leaf trichome density, and decreased stomata density. | Parsons et al. [161] |
| Hemp cultivars (Youngsim10, Mountain Mango, Cherry Wine, Wife, and Abacus × Wife) | Colchicine (0.02% or 0.05%) | 26–64% for 12 h in different cultivars | Not reported | Stomate size/density, leaf morphology, and flow cytometry | 0.05% colchicine for 12 h was required to induce polyploidy. Polyploids exhibit thickened hypocotyls and cotyledons, as well as larger stomata. | Kurtz et al. [160] |
| Genotype(s) | Explant(s) | Agrobacterium Strain(s) | Additives (mg/L) | Selection Marker | Promoter (s) | CCP (day) | OD (nm) | Transgene(s) | Method of Confirmation | Efficiency (%) | References |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Fedora19, Felina34 | Shoot tips | NR | CF (NR) | Herbicide | NR | NR | NR | PGIP | Post-inoculation with Botrytis cinerea. | ≥50 | MacKinnon et al. [171] |
| UnikoB, Kompolti, Anka, Felina-34 | Stem, leaf | A. tumefaciens EHA101 | Spc (150), K (50), AS (100 µM), T (300), D-mannose (1, 2, 3%) | D-mannose, Spc | Ubq3, NOS | 3 | 1.6–1.8 (600) | PMI, Spc | PMI assay, PCR, Southern blot | 15.1–55.3 | Feeney and Punja [172] |
| Futura77, Delta405, Delta-llosa, CAN0221, CAN0111 | Cotyledonary node, hypocotyls, primary leaves, cotyledons | A. tumefaciens LBA4404, C58, IVIA 251, and A. rhizogenes 476, 477, 478, A424, AR10GUS, A4, AR10, R1601 | AS (20, 100, 200 µM), sucrose (0.5, 2%), sodium citrate (20 mM), MES (30 mM) | K, Carb, Rif | 35S::GUS-INT; p35S-CODA-CAMV3′ | 2 | NR | BamHI, XhoI, GUS; LBA-rolABC (EcoRI); LBA-rolA (EcoRI-BamHI); LBA-rolB (SmaI-HpaI); LBA-rolC (HindIIII-EcoRI) | GUS assay, PCR, | 43–98 for A. rhizogenes; 33.7–63 for A. tumefaciens | Wahby et al. [173] |
| UnikoB, Kompolti, Anka, Felina-34 | Stem, leaf | A. tumefaciens EHA101 | Spc (150), K (50), AS (100 µM),T (300), D-mannose (1, 2, 3%) | D-mannose, Spc | Ubq3, NOS | 3 | 1.6–1.8 (600) | PMI and Spc | PMI assay, PCR, Southern blot | 15.1–55.3 | Feeney and Punja [174] |
| Ferimon, Fedora 17, USO31, Felina 32, Santhica 27, Futura 75, CRS-1, CFX-2 | male and female flowers, stem, leaf, root | A. tumefaciens LBA4404, GV3101, EHA105 | 200 μM AS, 2% glucose, 10 mM MES, Silwett L-77; Pluronic F-68; L-Ascorbic acid; PVP | CmR | CAMV35s, OCS | 3 | 0.5 (600) | eGFP, uidA, attBI, attBII, CsPDS, CmR, His- | GFP assay, GUS assay, qPCR | 10–80 | Deguchi et al. [175] |
| Candida CD-1, Holy Grail x CD-1, Green Crack CBD, Nightingale | Cotyledons, leaves | A. tumefaciens EHA105 | 100 µM AS, 3 mM Silver thiosulfate | K, Rif | CAMV35s, Ubq3, NOS | 3 | 0.6 (600) | PMI, GUS | GUS assay, MUG assay, PCR | 45–70.6 | Sorokin et al. [176] |
| Hemp strain (DMG278) | immature embryo hypocotyls | A. tumefaciens AGL1 | 50 mg/L K, 20 mg/L Rif | K, Rif | CaMV 35S promoter and NOS terminator | 3 | 0.1 (600) | CsGRF3–CsGIF1 | GFP assay, PCR, | 63% | Zhang et al. [93] |
| Features | Tools | ||||
|---|---|---|---|---|---|
| CRISPOR | CCTop | CHOPCHOP | Breaking Cas | CRISPR RGEN Tools | |
| wtSpCas9 nuclease/orthologues and Cas9 mutants | +/+ | +/+ | +/+ | +/+ | +/+ |
| Cpf1 (Cas12a) | + | + | - | + | + |
| Custom PAM | + | - | + | + | - |
| nickases/FokI-Cas9 | - | - | +/+ | - | - |
| nuclease-deaminase | - | - | - | - | + |
| (proto)spacer length | - | + | - | + | + |
| 5′-end of gRNA/in vitro transcription promoter | +/- | +/+ | +/- | - | - |
| mismatch | - | + | + | + | + |
| indels in spacers and protospacers | - | - | - | - | + |
| GC-content in protospacers | - | + | + | - | - |
| input of DNA through the clipboard/as a file | +/- | +/+ | +/- | +/+ | +/+ |
| input of individual genomes vis gene name or Accession Number/input of DNA using the genome coordinates | -/+ | - | +/+ | - | +/- |
| multiple sequences | - | + | - | - | - |
| ranked gRNAs | + | + | + | + | - |
| off-target sites | + | + | - | + | + |
| microhomology | + | - | - | - | + |
| restriction sites | + | - | + | - | - |
| both DNA strands/edited region (exon, intron, intergenic spacer) | +/+ | +/+ | +/+ | +/+ | +/+ |
| presence of the TTT(T) sequence | - | - | - | + | - |
| GC-content in protospacers/secondary gRNA structure (constant and variable parts) | - | - | +/- | - | +/- |
| oligonucleotides and primers for cloning/PCR detection | +/+ | - | +/- | - | - |
| demo version | - | - | - | + | - |
| off-line | + | + | - | - | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hesami, M.; Baiton, A.; Alizadeh, M.; Pepe, M.; Torkamaneh, D.; Jones, A.M.P. Advances and Perspectives in Tissue Culture and Genetic Engineering of Cannabis. Int. J. Mol. Sci. 2021, 22, 5671. https://doi.org/10.3390/ijms22115671
Hesami M, Baiton A, Alizadeh M, Pepe M, Torkamaneh D, Jones AMP. Advances and Perspectives in Tissue Culture and Genetic Engineering of Cannabis. International Journal of Molecular Sciences. 2021; 22(11):5671. https://doi.org/10.3390/ijms22115671
Chicago/Turabian StyleHesami, Mohsen, Austin Baiton, Milad Alizadeh, Marco Pepe, Davoud Torkamaneh, and Andrew Maxwell Phineas Jones. 2021. "Advances and Perspectives in Tissue Culture and Genetic Engineering of Cannabis" International Journal of Molecular Sciences 22, no. 11: 5671. https://doi.org/10.3390/ijms22115671
APA StyleHesami, M., Baiton, A., Alizadeh, M., Pepe, M., Torkamaneh, D., & Jones, A. M. P. (2021). Advances and Perspectives in Tissue Culture and Genetic Engineering of Cannabis. International Journal of Molecular Sciences, 22(11), 5671. https://doi.org/10.3390/ijms22115671