
Intricate Connections between the Microbiota and Endometriosis

Abstract
| Term | Definition |
| Microbiota | The collection of all the microorganisms residing in and on the body, including bacteria, archaea, protists, fungi, and viruses |
| Microbiome | The aggregate of all the genetic material of the microbiota |
| Estrobolome | The total collection of genes, in the gut microbiota, responsible for estrogen metabolism |
| Metabolome | The total collection of metabolites in a given environment |
| Dysbiosis | Imbalance or impairment of the microbiota, characterized by gain of pathogenic microbes or loss of probiotics |
| Prebiotic | Compounds that promote growth and activity of beneficial microorganisms |
| Probiotic | Live microorganisms that are beneficial to host health |
1. Endometriosis
1.1. Introduction of Endometriosis
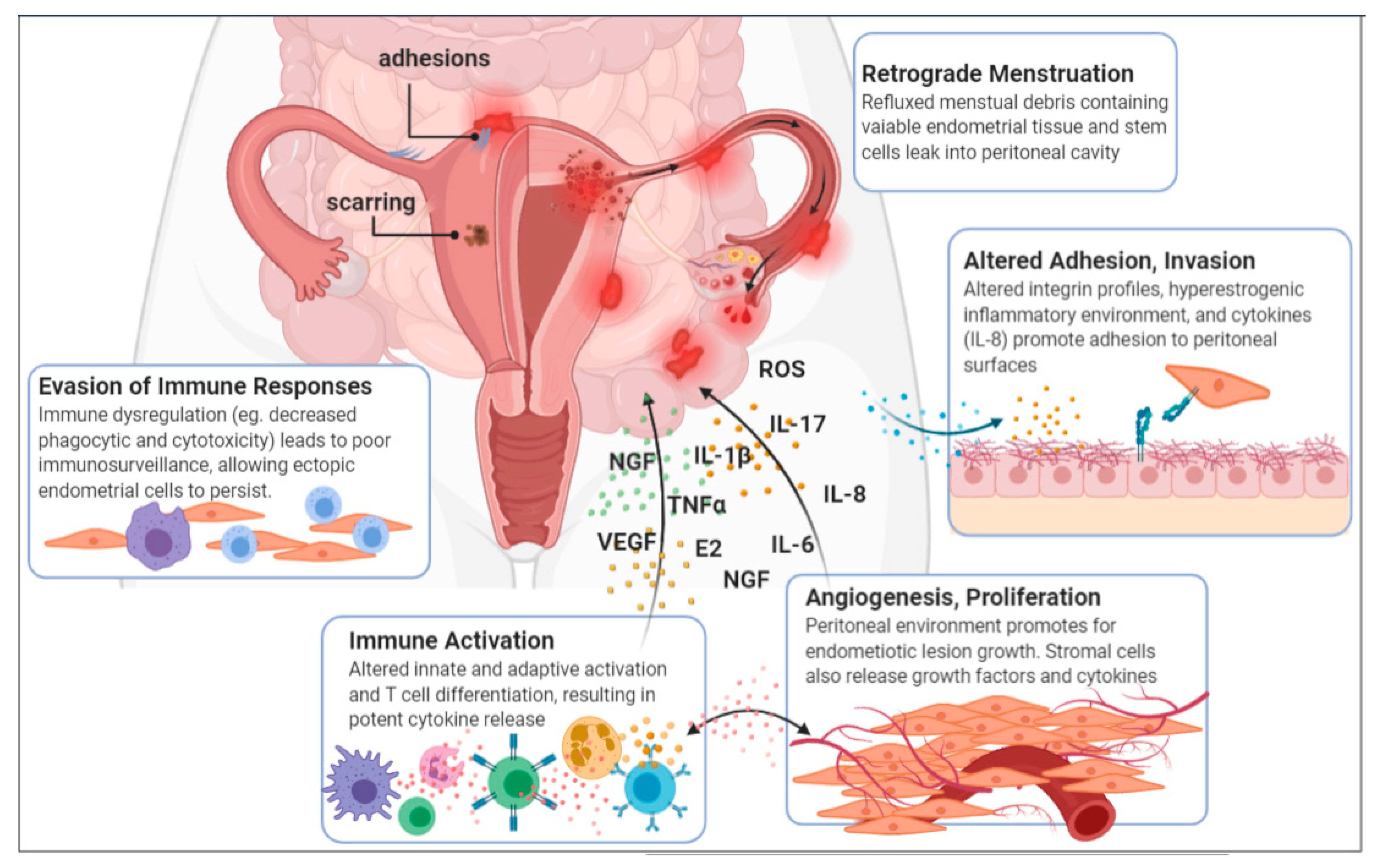
1.2. Aetiology and Pathogenesis
1.3. A Disease of the Immune System
1.3.1. Elevated Inflammatory Mediators
1.3.2. Macrophages: Principal Contributors to the Pathogenesis of Endometriosis
1.3.3. Preconditioned Neutrophils
1.3.4. Impaired Natural Killer Cells
1.3.5. Altered T cell Differentiation
1.3.6. Activated B Cells
1.4. Estrogen Levels and Signaling Is Altered in Endometriosis
2. The Microbiota
2.1. Introduction to the Microbiota
2.2. Dysbiosis
2.3. Gut Microbiota
Role of the Gut Microbiota in Host Immune Function
2.4. Female Reproductive Tract Microbiota
2.4.1. Proof of Existence
2.4.2. Female Reproductive Tract Microbiota Composition
3. Evidence of an Intricate Connection
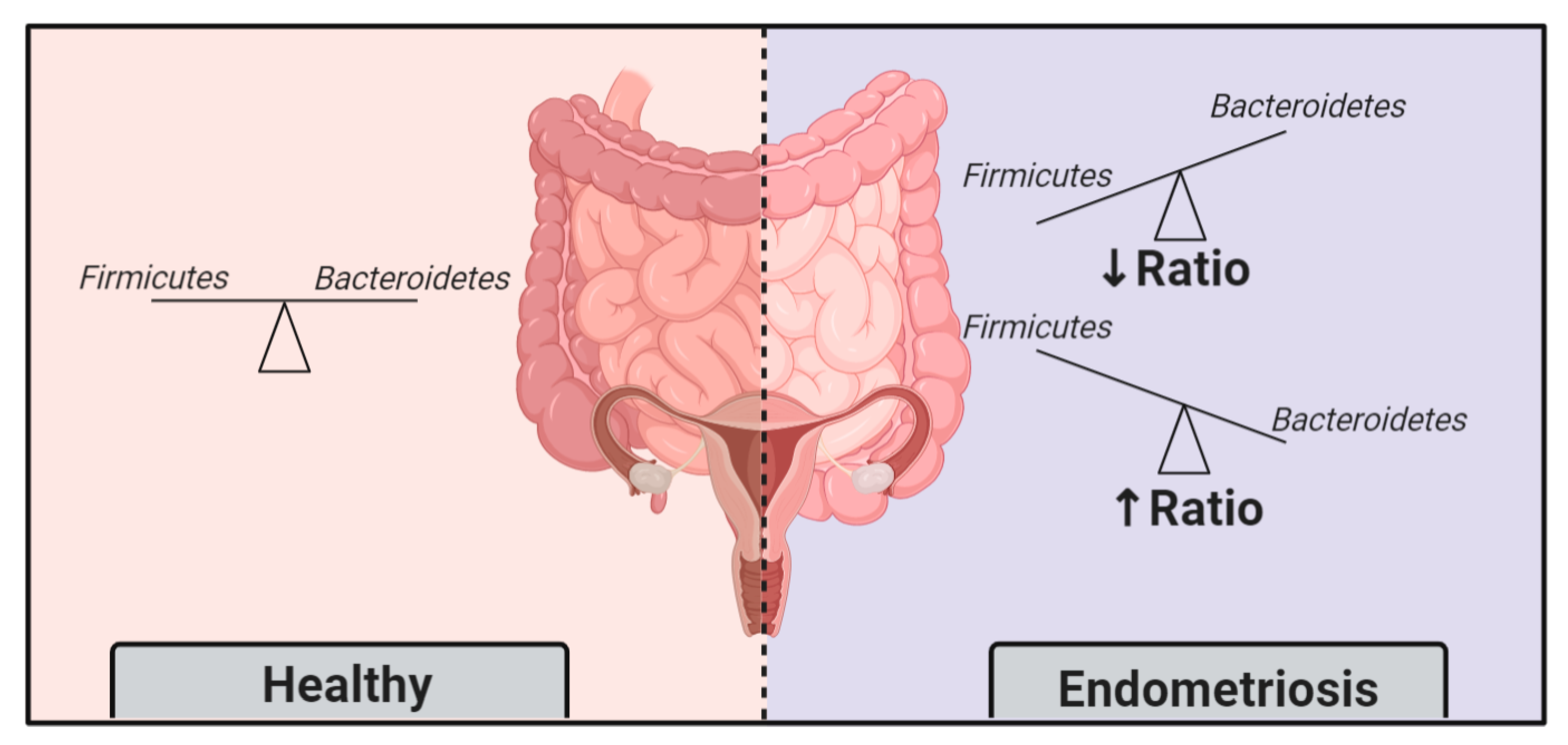
3.1. Endometriotic Women Exhibit Altered Microbiotas
3.2. Endometriosis Induces Gut Microbiota Alterations
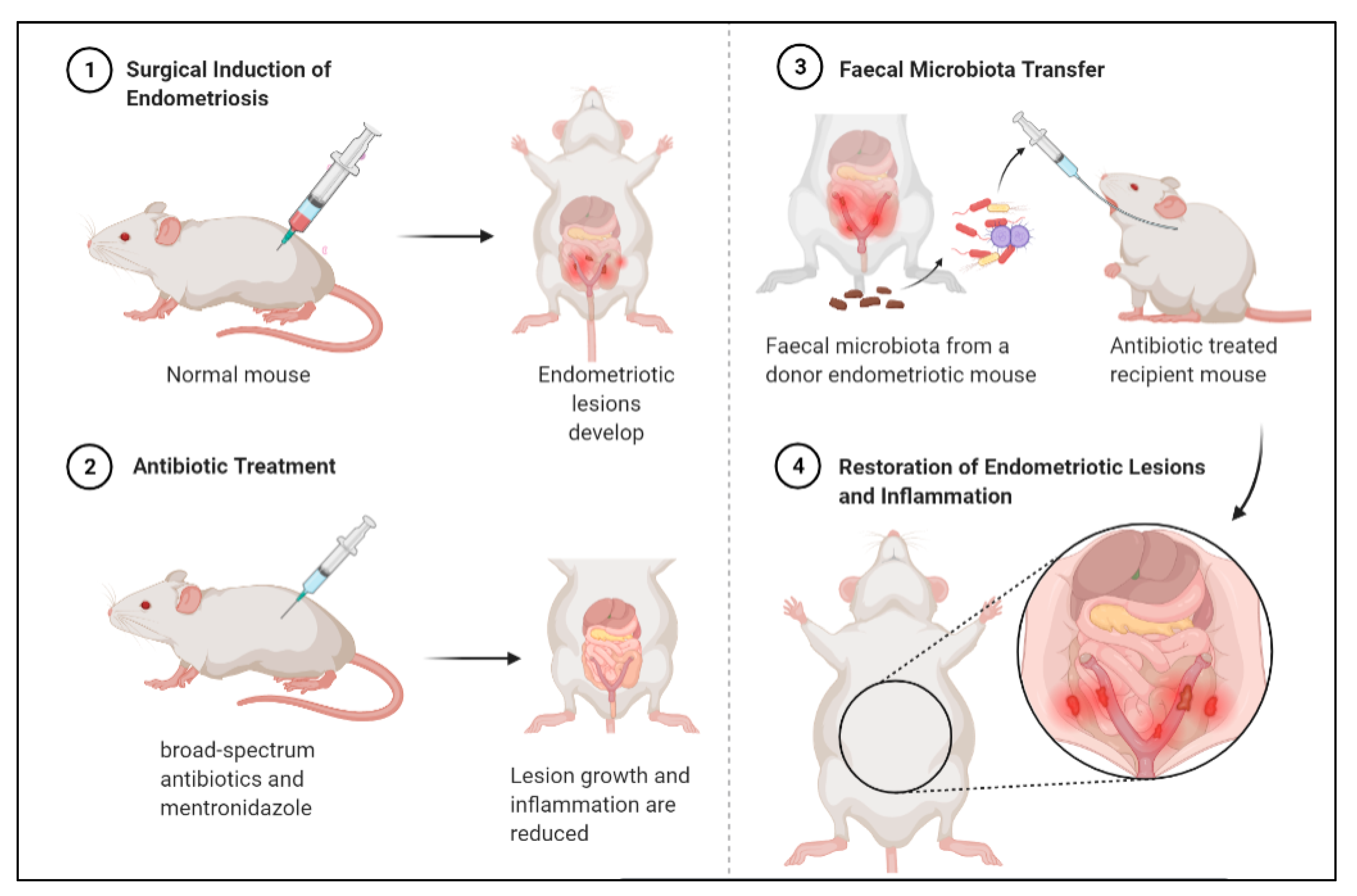
3.3. Faecal Microbiota Transfer Induces Endometriosis
3.4. Diet-Induced Gut Microbiota Changes Reduce Endometriosis Risk
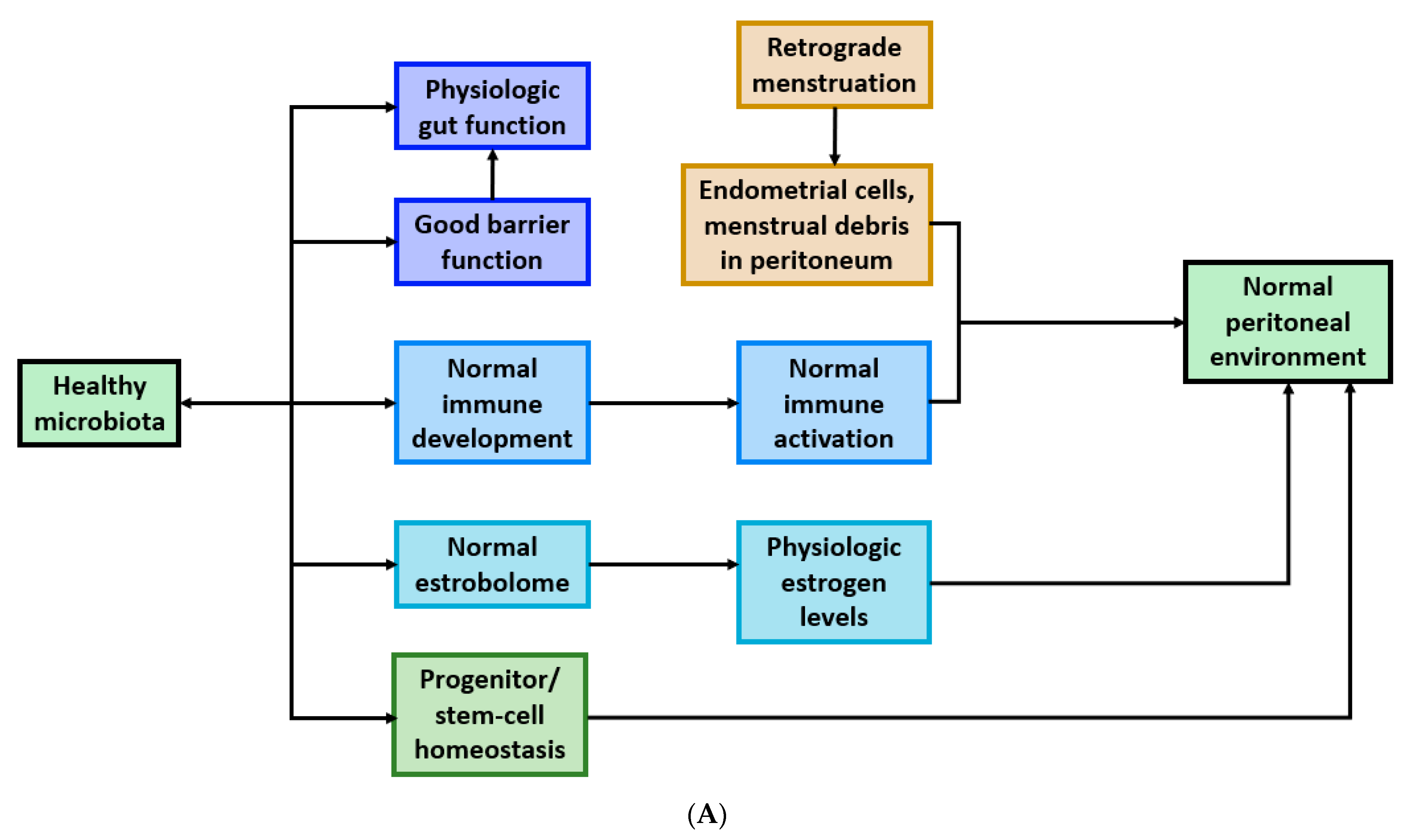
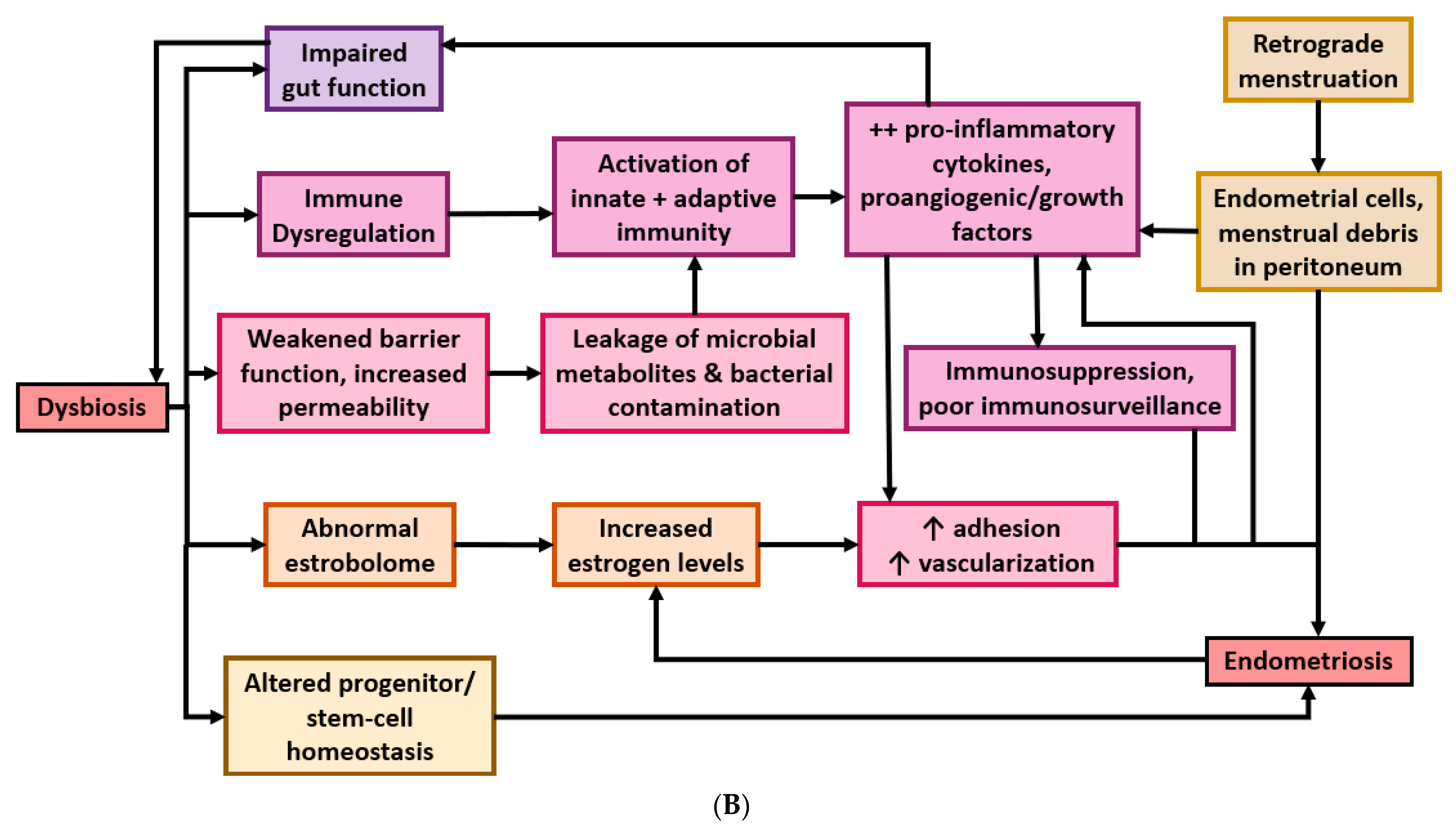
4. Postulated Mechanisms of Microbiota Involvement in Endometriosis
4.1. Bacterial Contamination Theory and Immune Activation
4.2. Cytokines Affect Gut Function
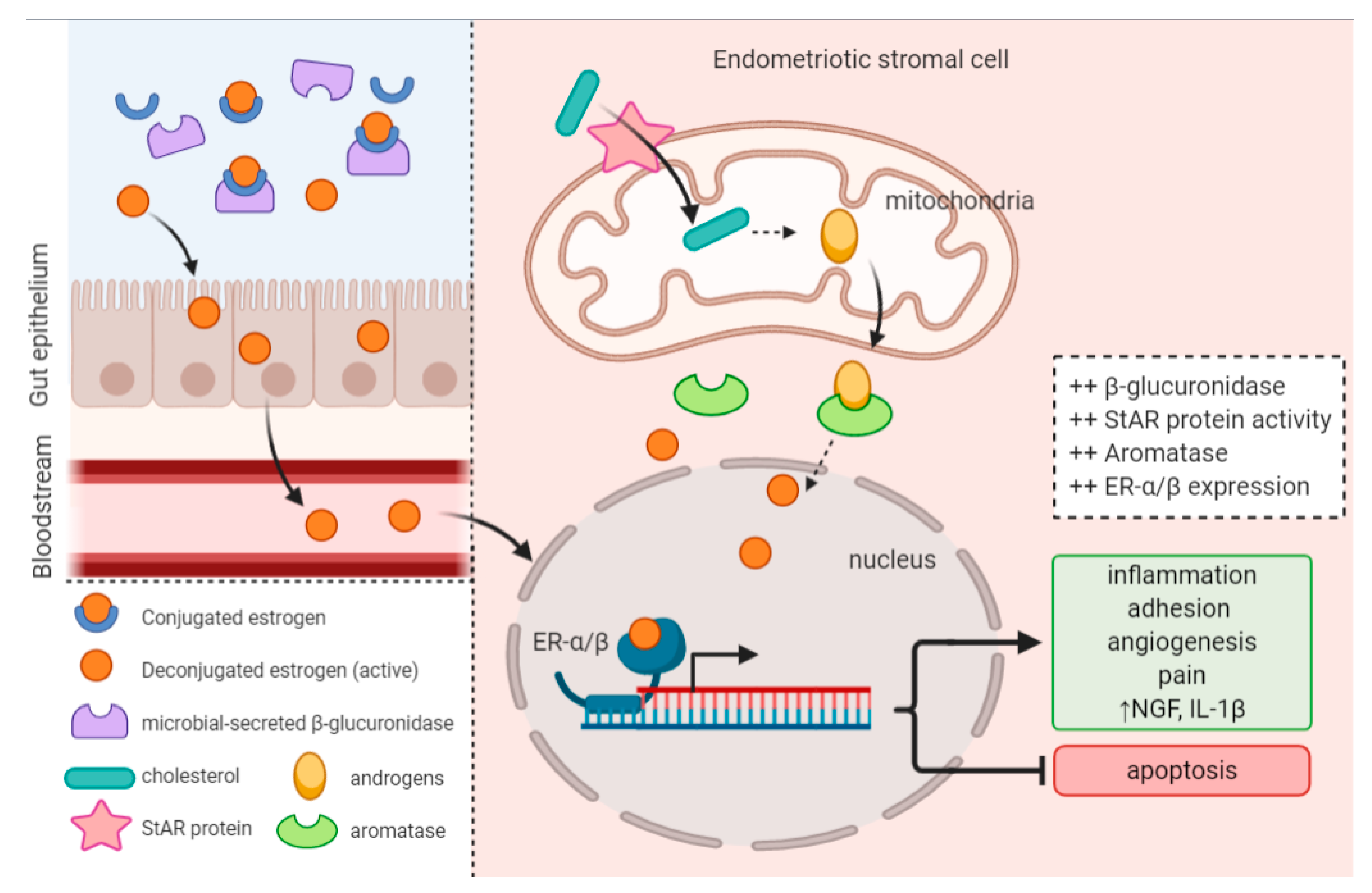
4.3. Microbiota Composition and Estrogen Availability
4.4. Microbiota Regulates Progenitor and Stem-Cell Homeostasis
5. What Can This Mean for Endometriosis Care
5.1. Gynaecologic and Obstetric Applications of Microbiota Modulation
5.2. Treating Endometriosis with Antibiotics
5.3. Treating Endometriosis with Probiotics
5.4. A Mechanism for Known Treatments
5.5. Side Effects and Challenges
5.6. Opportunities for Diagnostics
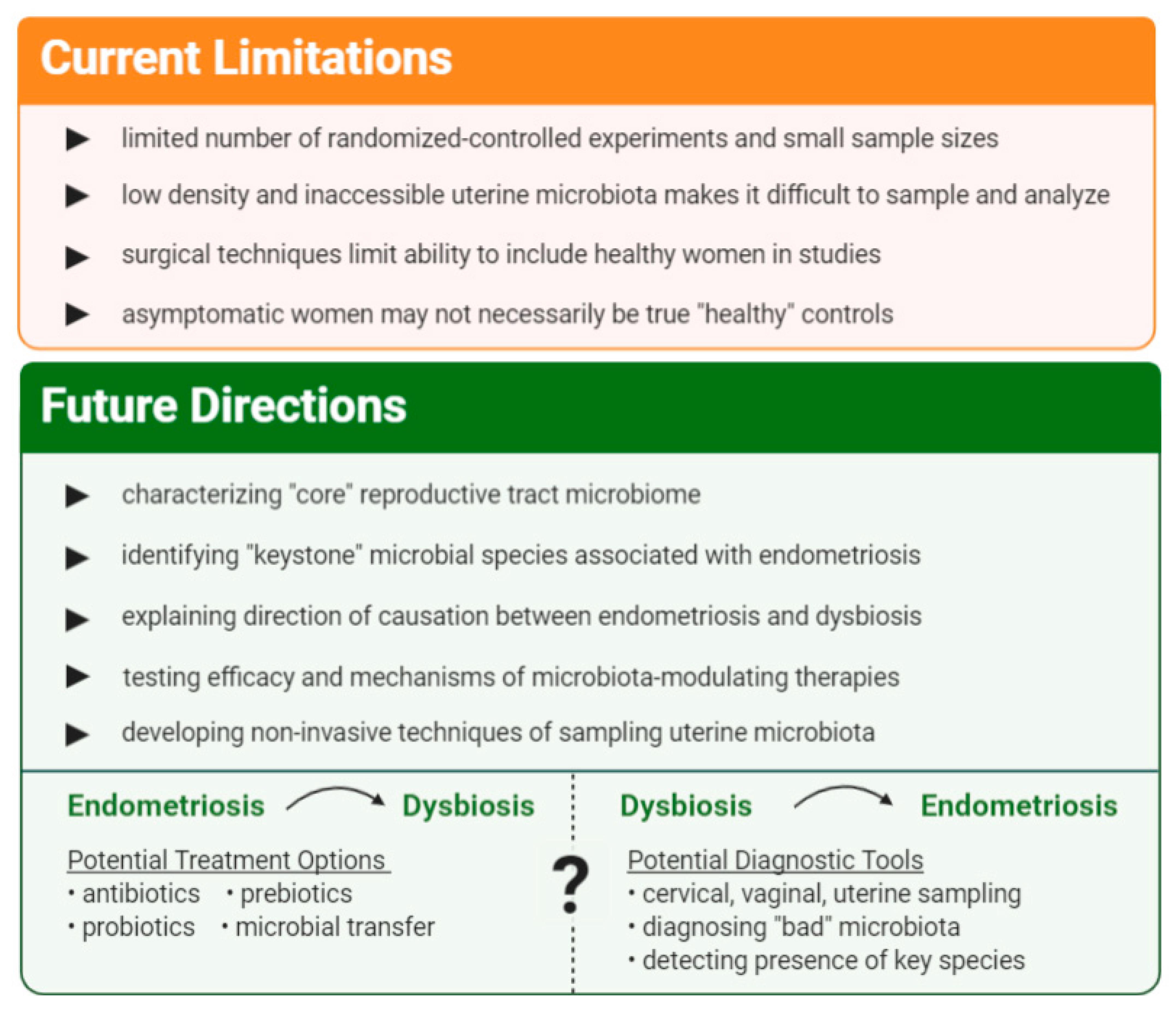
6. Limitations and Future Directions
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zondervan, K.T.; Becker, C.M.; Koga, K.; Missmer, S.A.; Taylor, R.N.; Viganò, P. Endometriosis. Nat. Rev. Dis. Primers 2018, 4, 9. [Google Scholar] [CrossRef]
- Zondervan, K.T.; Becker, C.M.; Missmer, S.A. Endometriosis. N. Engl. J. Med. 2020, 382, 1244–1256. [Google Scholar] [CrossRef] [PubMed]
- Busacca, M.; Vignali, M. Ovarian endometriosis: From pathogenesis to surgical treatment. Curr. Opin. Obstet. Gynecol. 2003, 15, 321–326. [Google Scholar] [CrossRef]
- Dunselman, G.A.J.; Vermeulen, N.; Becker, C.; Calhaz-Jorge, C.; D’Hooghe, T.; De Bie, B.; Heikinheimo, O.; Horne, A.W.; Kiesel, L.; Nap, A.; et al. ESHRE Guideline: Management of Women with Endometriosis. Hum. Reprod. 2014, 29, 400–412. [Google Scholar] [CrossRef]
- Eskenazi, B.; Warner, M.L. Epidemiology of endometriosis. Obstet. Gynecol. Clin. N. Am. 1997, 24, 235–258. [Google Scholar] [CrossRef]
- Vercellini, P.; Fedele, L.; Aimi, G.; Pietropaolo, G.; Consonni, D.; Crosignani, P.G. Association between Endometriosis Stage, Lesion Type, Patient Characteristics and Severity of Pelvic Pain Symptoms: A Multivariate Analysis of over 1000 Patients. Hum. Reprod. 2007, 22, 266–271. [Google Scholar] [CrossRef] [PubMed]
- Arion, K.; Orr, N.L.; Noga, H.; Allaire, C.; Williams, C.; Bedaiwy, M.A.; Yong, P.J. A Quantitative Analysis of Sleep Quality in Women with Endometriosis. J. Women’s Health 2020, 29, 1209–1215. [Google Scholar] [CrossRef]
- Shum, L.K.; Bedaiwy, M.A.; Allaire, C.; Williams, C.; Noga, H.; Albert, A.; Lisonkova, S.; Yong, P.J. Deep Dyspareunia and Sexual Quality of Life in Women with Endometriosis. Sex. Med. 2018, 6, 224–233. [Google Scholar] [CrossRef]
- Sepulcri, R.d.P.; do Amaral, V.F. Depressive Symptoms, Anxiety, and Quality of Life in Women with Pelvic Endometriosis. Eur. J. Obstet. Gynecol. Reprod. Biol. 2009, 142, 53–56. [Google Scholar] [CrossRef]
- Simoens, S.; Dunselman, G.; Dirksen, C.; Hummelshoj, L.; Bokor, A.; Brandes, I.; Brodszky, V.; Canis, M.; Colombo, G.L.; DeLeire, T.; et al. The Burden of Endometriosis: Costs and Quality of Life of Women with Endometriosis and Treated in Referral Centres. Hum. Reprod. 2014, 29, 2073. [Google Scholar] [CrossRef]
- Vercellini, P.; Vigano, P.; Somigliana, E.; Fedele, L. Endometriosis: Pathogenesis and Treatment. Nat. Rev. Endocrinol. 2014, 10, 261–276. [Google Scholar] [CrossRef]
- Bedaiwy, M.A.; Barker, N.M. Evidence Based Surgical Management of Endometriosis. Middle East. Fertil. Soc. J. 2012, 17, 57–60. [Google Scholar] [CrossRef]
- Bedaiwy, M.A.; Alfaraj, S.; Yong, P.; Casper, R. New Developments in the Medical Treatment of Endometriosis. Fertil. Steril. 2017, 107, 555–565. [Google Scholar] [CrossRef] [PubMed]
- Vercellini, P.; Somigliana, E.; Viganò, P.; Abbiati, A.; Daguati, R.; Crosignani, P.G. Endometriosis: Current and Future Medical Therapies. Best Pract. Res. Clin. Obstet. Gynaecol. 2008, 22, 275–306. [Google Scholar] [CrossRef] [PubMed]
- Yovich, J.L.; Rowlands, P.K.; Lingham, S.; Sillender, M.; Srinivasan, S. Pathogenesis of Endometriosis: Look No Further than John Sampson. Reprod. Biomed. Online 2020, 40, 7–11. [Google Scholar] [CrossRef]
- Halme, J.; Hammond, M.G.; Hulka, J.F.; Raj, S.G.; Talbert, L.M. Retrograde Menstruation in Healthy Women and in Patients with Endometriosis. Obstet. Gynecol. 1984, 64, 151–154. [Google Scholar] [PubMed]
- Laschke, M.W.; Menger, M.D. The Gut Microbiota: A Puppet Master in the Pathogenesis of Endometriosis? Am. J. Obstet. Gynecol. 2016, 215, 68.e1–68.e4. [Google Scholar] [CrossRef]
- Harada, T.; Iwabe, T.; Terakawa, N. Role of Cytokines in Endometriosis. Fertil. Steril. 2001, 76, 1–10. [Google Scholar] [CrossRef]
- Klemmt, P.A.B.; Carver, J.G.; Koninckx, P.; McVeigh, E.J.; Mardon, H.J. Endometrial Cells from Women with Endometriosis Have Increased Adhesion and Proliferative Capacity in Response to Extracellular Matrix Components: Towards a Mechanistic Model for Endometriosis Progression. Hum. Reprod. 2007, 22, 3139–3147. [Google Scholar] [CrossRef]
- Garcia-Velasco, J.A.; Arici, A. P-255. Interleukin-8 Stimulates Endometrial Stromal Cell Adhesion to Fibronectin. Hum. Reprod. 1999, 14, 268. [Google Scholar] [CrossRef]
- Symons, L.K.; Miller, J.E.; Kay, V.R.; Marks, R.M.; Liblik, K.; Koti, M.; Tayade, C. The Immunopathophysiology of Endometriosis. Trends Mol. Med. 2018, 24, 748–762. [Google Scholar] [CrossRef]
- Cosín, R.; Gilabert-Estellés, J.; Ramón, L.A.; Gómez-Lechón, M.J.; Gilabert, J.; Chirivella, M.; Braza-Boïls, A.; España, F.; Estellés, A. Influence of Peritoneal Fluid on the Expression of Angiogenic and Proteolytic Factors in Cultures of Endometrial Cells from Women with Endometriosis. Hum. Reprod. 2010, 25, 398–405. [Google Scholar] [CrossRef] [PubMed]
- McLaren, J.; Prentice, A.; Charnock-Jones, D.S.; Millican, S.A.; Müller, K.H.; Sharkey, A.M.; Smith, S.K. Vascular Endothelial Growth Factor Is Produced by Peritoneal Fluid Macrophages in Endometriosis and Is Regulated by Ovarian Steroids. J. Clin. Investig. 1996, 98, 482–489. [Google Scholar] [CrossRef] [PubMed]
- Iwabe, T.; Harada, T.; Tsudo, T.; Nagano, Y.; Yoshida, S.; Tanikawa, M.; Terakawa, N. Tumor Necrosis Factor-α Promotes Proliferation of Endometriotic Stromal Cells by Inducing Interleukin-8 Gene and Protein Expression. J. Clin. Endocrinol. Metab. 2000, 85, 824–829. [Google Scholar] [CrossRef] [PubMed]
- Leibovich, S.J.; Polverini, P.J.; Shepard, H.M.; Wiseman, D.M.; Shively, V.; Nuseir, N. Macrophage-Induced Angiogenesis Is Mediated by Tumour Necrosis Factor- α. Nature 1987, 329, 630–632. [Google Scholar] [CrossRef]
- Nothnick, W.B. Treating Endometriosis as an Autoimmune Disease. Fertil. Steril. 2001, 76, 223–231. [Google Scholar] [CrossRef]
- García-Peñarrubia, P.; Ruiz-Alcaraz, A.J.; Martínez-Esparza, M.; Marín, P.; Machado-Linde, F. Hypothetical Roadmap towards Endometriosis: Prenatal Endocrine-Disrupting Chemical Pollutant Exposure, Anogenital Distance, Gut-Genital Microbiota and Subclinical Infections. Hum. Reprod. Update 2020, 26, 214–246. [Google Scholar] [CrossRef]
- Shigesi, N.; Kvaskoff, M.; Kirtley, S.; Feng, Q.; Fang, H.; Knight, J.C.; Missmer, S.A.; Rahmioglu, N.; Zondervan, K.T.; Becker, C.M. The Association between Endometriosis and Autoimmune Diseases: A Systematic Review and Meta-Analysis. Hum. Reprod. Update 2019, 25, 486–503. [Google Scholar] [CrossRef]
- Tsudo, T.; Harada, T.; Iwabe, T.; Tanikawa, M.; Nagano, Y.; Ito, M.; Taniguchi, F.; Terakawa, N. Altered Gene Expression and Secretion of Interleukin-6 in Stromal Cells Derived from Endometriotic Tissues. Fertil. Steril. 2000, 73, 205–211. [Google Scholar] [CrossRef]
- Tseng, J.F.; Ryan, I.P.; Milam, T.D.; Murai, J.T.; Schriock, E.D.; Landers, D.V.; Taylor, R.N. Interleukin-6 Secretion in Vitro Is up-Regulated in Ectopic and Eutopic Endometrial Stromal Cells from Women with Endometriosis. J. Clin. Endocrinol. Metab. 1996, 81, 1118–1122. [Google Scholar] [CrossRef][Green Version]
- Beste, M.T.; Pfäffle-Doyle, N.; Prentice, E.A.; Morris, S.N.; Lauffenburger, D.A.; Isaacson, K.B.; Griffith, L.G. Molecular Network Analysis of Endometriosis Reveals a Role for C-Jun–Regulated Macrophage Activation. Sci. Transl. Med. 2014, 6, 222ra16. [Google Scholar] [CrossRef] [PubMed]
- Mu, F.; Harris, H.R.; Rich-Edwards, J.W.; Hankinson, S.E.; Rimm, E.B.; Spiegelman, D.; Missmer, S.A. A Prospective Study of Inflammatory Markers and Risk of Endometriosis. Am. J. Epidemiol. 2018, 187, 515–522. [Google Scholar] [CrossRef] [PubMed]
- Vallvé-Juanico, J.; Santamaria, X.; Vo, K.C.; Houshdaran, S.; Giudice, L.C. Macrophages Display Proinflammatory Phenotypes in the Eutopic Endometrium of Women with Endometriosis with Relevance to an Infectious Etiology of the Disease. Fertil. Steril. 2019, 112, 1118–1128. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.-J.; Lai, M.-D.; Lei, H.-Y.; Wing, L.-Y.C. Neutrophils and Macrophages Promote Angiogenesis in the Early Stage of Endometriosis in a Mouse Model. Endocrinology 2006, 147, 1278–1286. [Google Scholar] [CrossRef] [PubMed]
- Bacci, M.; Capobianco, A.; Monno, A.; Cottone, L.; Di Puppo, F.; Camisa, B.; Mariani, M.; Brignole, C.; Ponzoni, M.; Ferrari, S.; et al. Macrophages Are Alternatively Activated in Patients with Endometriosis and Required for Growth and Vascularization of Lesions in a Mouse Model of Disease. Am. J. Pathol. 2009, 175, 547–556. [Google Scholar] [CrossRef] [PubMed]
- Hirayama, D.; Iida, T.; Nakase, H. The Phagocytic Function of Macrophage-Enforcing Innate Immunity and Tissue Homeostasis. Int. J. Mol. Sci. 2017, 19, 92. [Google Scholar] [CrossRef] [PubMed]
- Salamonsen, L.A.; Zhang, J.; Brasted, M. Leukocyte Networks and Human Endometrial Remodelling. J. Reprod. Immunol. 2002, 57, 95–108. [Google Scholar] [CrossRef]
- Berbic, M.; Schulke, L.; Markham, R.; Tokushige, N.; Russell, P.; Fraser, I.S. Macrophage Expression in Endometrium of Women with and without Endometriosis. Hum. Reprod. 2009, 24, 325–332. [Google Scholar] [CrossRef]
- Bedaiwy, M.A. Endometrial Macrophages, Endometriosis, and Microbiota: Time to Unravel the Complexity of the Relationship. Fertil. Steril. 2019, 112, 1049–1050. [Google Scholar] [CrossRef]
- Lousse, J.-C.; Van Langendonckt, A.; González-Ramos, R.; Defrère, S.; Renkin, E.; Donnez, J. Increased Activation of Nuclear Factor-Kappa B (NF-ΚB) in Isolated Peritoneal Macrophages of Patients with Endometriosis. Fertil. Steril. 2008, 90, 217–220. [Google Scholar] [CrossRef] [PubMed]
- Milewski, Ł.; Dziunycz, P.; Barcz, E.; Radomski, D.; Roszkowski, P.I.; Korczak-Kowalska, G.; Kamiński, P.; Malejczyk, J. Increased Levels of Human Neutrophil Peptides 1, 2, and 3 in Peritoneal Fluid of Patients with Endometriosis: Association with Neutrophils, T Cells and IL-8. J. Reprod. Immunol. 2011, 91, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Karmarkar, D.; Rock, K.L. Microbiota Signalling through MyD88 Is Necessary for a Systemic Neutrophilic Inflammatory Response. Immunology 2013, 140, 483–492. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.-Y.; Yang, J.-H.; Chao, K.-H.; Hwang, J.-L.; Yang, Y.-S.; Ho, H.-N. Increase in the Expression of Killer Cell Inhibitory Receptors on Peritoneal Natural Killer Cells in Women with Endometriosis. Fertil. Steril. 2000, 74, 1187–1191. [Google Scholar] [CrossRef]
- Podgaec, S.; Abrao, M.S.; Dias, J.A., Jr.; Rizzo, L.V.; de Oliveira, R.M.; Baracat, E.C. Endometriosis: An Inflammatory Disease with a Th2 Immune Response Component. Hum. Reprod. 2007, 22, 1373–1379. [Google Scholar] [CrossRef] [PubMed]
- Antsiferova, Y.S.; Sotnikova, N.Y.; Posiseeva, L.V.; Shor, A.L. Changes in the T-Helper Cytokine Profile and in Lymphocyte Activation at the Systemic and Local Levels in Women with Endometriosis. Fertil. Steril. 2005, 84, 1705–1711. [Google Scholar] [CrossRef] [PubMed]
- Gogacz, M.; Winkler, I.; Bojarska-Junak, A.; Tabarkiewicz, J.; Semczuk, A.; Rechberger, T.; Adamiak, A. Increased Percentage of Th17 Cells in Peritoneal Fluid Is Associated with Severity of Endometriosis. J. Reprod. Immunol. 2016, 117, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Laux-Biehlmann, A.; d’Hooghe, T.; Zollner, T.M. Menstruation Pulls the Trigger for Inflammation and Pain in Endometriosis. Trends Pharmacol. Sci. 2015, 36, 270–276. [Google Scholar] [CrossRef]
- Ahn, S.H.; Edwards, A.K.; Singh, S.S.; Young, S.L.; Lessey, B.A.; Tayade, C. IL-17A Contributes to the Pathogenesis of Endometriosis by Triggering Proinflammatory Cytokines and Angiogenic Growth Factors. J. Immunol. 2015, 195, 2591–2600. [Google Scholar] [CrossRef]
- Shen, P.; Fillatreau, S. Antibody-Independent Functions of B Cells: A Focus on Cytokines. Nat. Rev. Immunol. 2015, 15, 441–452. [Google Scholar] [CrossRef]
- Randall, G.W.; Gantt, P.A.; Poe-Zeigler, R.L.; Bergmann, C.A.; Noel, M.E.; Strawbridge, W.R.; Richardson-Cox, B.; Hereford, J.R.; Reiff, R.H. Serum Antiendometrial Antibodies and Diagnosis of Endometriosis. Am. J. Reprod. Immunol. 2007, 58, 374–382. [Google Scholar] [CrossRef] [PubMed]
- Chantalat, E.; Valera, M.-C.; Vaysse, C.; Noirrit, E.; Rusidze, M.; Weyl, A.; Vergriete, K.; Buscail, E.; Lluel, P.; Fontaine, C.; et al. Estrogen Receptors and Endometriosis. Available online: https://doaj.org (accessed on 26 January 2021).
- Galvankar, M.; Singh, N.; Modi, D. Estrogen Is Essential but Not Sufficient to Induce Endometriosis. J. Biosci. 2017, 42, 251–263. [Google Scholar] [CrossRef]
- Reis, F.M.; Petraglia, F.; Taylor, R.N. Endometriosis: Hormone Regulation and Clinical Consequences of Chemotaxis and Apoptosis. Hum. Reprod. Update 2013, 19, 406–418. [Google Scholar] [CrossRef]
- Han, S.J.; Jung, S.Y.; Wu, S.-P.; Hawkins, S.M.; Park, M.J.; Kyo, S.; Qin, J.; Lydon, J.P.; Tsai, S.Y.; Tsai, M.-J.; et al. Estrogen Receptor β Modulates Apoptosis Complexes and the Inflammasome to Drive the Pathogenesis of Endometriosis. Cell 2015, 163, 960–974. [Google Scholar] [CrossRef] [PubMed]
- Burns, K.A.; Rodriguez, K.F.; Hewitt, S.C.; Janardhan, K.S.; Young, S.L.; Korach, K.S. Role of Estrogen Receptor Signaling Required for Endometriosis-Like Lesion Establishment in a Mouse Model. Endocrinology 2012, 153, 3960–3971. [Google Scholar] [CrossRef]
- Morotti, M.; Vincent, K.; Becker, C.M. Mechanisms of Pain in Endometriosis. Eur. J. Obstet. Gynecol. Reprod. Biol. 2017, 209, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Bulun, S.E. Endometriosis. N. Engl. J. Med. 2009, 360, 268–279. [Google Scholar] [CrossRef]
- Ervin, S.M.; Li, H.; Lim, L.; Roberts, L.R.; Liang, X.; Mani, S.; Redinbo, M.R. Gut Microbial β-Glucuronidases Reactivate Estrogens as Components of the Estrobolome That Reactivate Estrogens. J. Biol. Chem. 2019, 294, 18586–18599. [Google Scholar] [CrossRef] [PubMed]
- Baker, J.M.; Al-Nakkash, L.; Herbst-Kralovetz, M.M. Estrogen–Gut Microbiome Axis: Physiological and Clinical Implications. Maturitas 2017, 103, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Goedert, J.J.; Jones, G.; Hua, X.; Xu, X.; Yu, G.; Flores, R.; Falk, R.T.; Gail, M.H.; Shi, J.; Ravel, J.; et al. Investigation of the Association between the Fecal Microbiota and Breast Cancer in Postmenopausal Women: A Population-Based Case-Control Pilot Study. J. Natl. Cancer Inst. 2015, 107. [Google Scholar] [CrossRef]
- Mikhael, S.; Punjala-Patel, A.; Gavrilova-Jordan, L. Hypothalamic-Pituitary-Ovarian Axis Disorders Impacting Female Fertility. Available online: https://doaj.org (accessed on 18 February 2021).
- Chen, J.; Li, Y.; Tian, Y.; Huang, C. Interaction between Microbes and Host Intestinal Health: Modulation by Dietary Nutrients and Gut-Brain-Endocrine-Immune Axis. Curr. Protein Pept. Sci. 2015, 16, 592. [Google Scholar] [CrossRef]
- Baj, A.; Moro, E.; Bistoletti, M.; Orlandi, V.; Crema, F.; Giaroni, C. Glutamatergic Signaling Along the Microbiota-Gut-Brain Axis. Available online: https://doaj.org (accessed on 18 February 2021).
- Jones, L.A.; Sun, E.W.; Martin, A.M.; Keating, D.J. The Ever-Changing Roles of Serotonin. Int. J. Biochem. Cell Biol. 2020, 125, 105776. [Google Scholar] [CrossRef] [PubMed]
- Hiller-Sturmhöfel, S.; Bartke, A. The Endocrine System. Alcohol Health Res. World 1998, 22, 153–164. [Google Scholar]
- Ogunrinola, G.A.; Oyewale, J.O.; Oshamika, O.O.; Olasehinde, G.I. The Human Microbiome and Its Impacts on Health. Available online: https://doaj.org (accessed on 6 January 2021).
- Cho, I.; Blaser, M.J. The Human Microbiome: At the Interface of Health and Disease. Nat. Rev. Genet. 2012, 13, 260–270. [Google Scholar] [CrossRef] [PubMed]
- Hooper, L.V.; Littman, D.R.; Macpherson, A.J. Interactions Between the Microbiota and the Immune System. Science 2012, 336, 1268–1273. [Google Scholar] [CrossRef] [PubMed]
- Forbes, J.D.; Chen, C.; Knox, N.C.; Marrie, R.-A.; El-Gabalawy, H.; de Kievit, T.; Alfa, M.; Bernstein, C.N.; Domselaar, G.V. A Comparative Study of the Gut Microbiota in Immune-Mediated Inflammatory Diseases—Does a Common Dysbiosis Exist? Available online: https://doaj.org (accessed on 26 January 2021).
- West, C.E.; Renz, H.; Jenmalm, M.C.; Kozyrskyj, A.L.; Allen, K.J.; Vuillermin, P.; Prescott, S.L.; MacKay, C.; Salminen, S.; Wong, G.; et al. The Gut Microbiota and Inflammatory Noncommunicable Diseases: Associations and Potentials for Gut Microbiota Therapies. J. Allergy Clin. Immunol. 2015, 135, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Pickard, J.M.; Zeng, M.Y.; Caruso, R.; Núñez, G. Gut Microbiota: Role in Pathogen Colonization, Immune Responses, and Inflammatory Disease. Immunol. Rev. 2017, 279, 70–89. [Google Scholar] [CrossRef] [PubMed]
- Spurbeck, R.R.; Arvidson, C.G. Inhibition of Neisseria Gonorrhoeae Epithelial Cell Interactions by Vaginal Lactobacillus Species. Infect. Immun. 2012, 80, 3742. [Google Scholar] [CrossRef]
- Benner, M.; Ferwerda, G.; Joosten, I.; van der Molen, R.G. How Uterine Microbiota Might Be Responsible for a Receptive, Fertile Endometrium. Hum. Reprod. Update 2018, 24, 393–415. [Google Scholar] [CrossRef]
- Baker, J.M.; Chase, D.M.; Herbst-Kralovetz, M.M. Uterine Microbiota: Residents, Tourists, or Invaders? Front. Immunol. 2018. [Google Scholar] [CrossRef]
- Chen, C.; Song, X.; Wei, W.; Zhong, H.; Dai, J.; Lan, Z.; Li, F.; Yu, X.; Feng, Q.; Wang, Z.; et al. The Microbiota Continuum along the Female Reproductive Tract and Its Relation to Uterine-Related Diseases. Nat. Commun. 2017, 8, 875. [Google Scholar] [CrossRef]
- Moreno, I.; Codoñer, F.M.; Vilella, F.; Valbuena, D.; Martinez-Blanch, J.F.; Jimenez-Almazán, J.; Alonso, R.; Alamá, P.; Remohí, J.; Pellicer, A.; et al. Evidence That the Endometrial Microbiota Has an Effect on Implantation Success or Failure. Am. J. Obstet. Gynecol. 2016, 215, 684–703. [Google Scholar] [CrossRef]
- Mitchell, C.M.; Haick, A.; Nkwopara, E.; Garcia, R.; Rendi, M.; Agnew, K.; Fredricks, D.N.; Eschenbach, D. Colonization of the Upper Genital Tract by Vaginal Bacterial Species in Nonpregnant Women. Am. J. Obstet. Gynecol. 2015, 212, 611.e1–611.e9. [Google Scholar] [CrossRef] [PubMed]
- Onderdonk, A.B.; Delaney, M.L.; Fichorova, R.N. The Human Microbiome during Bacterial Vaginosis. Clin. Microbiol. Rev. 2016, 29, 223–238. [Google Scholar] [CrossRef] [PubMed]
- Ling, Z.; Liu, X.; Luo, Y.; Wu, X.; Yuan, L.; Tong, X.; Li, L.; Xiang, C. Associations between Vaginal Pathogenic Community and Bacterial Vaginosis in Chinese Reproductive-Age Women. PLoS ONE 2013, 8, e76589. [Google Scholar] [CrossRef]
- Skarin, A.; Sylwan, J. Vaginal Lactobacilli Inhibiting Growth of Gardnerella Vaginalis, Mobiluncus and Other Bacterial Species Cultured from Vaginal Content of Women with Bacterial Vaginosis. Acta Pathol. Microbiol. Immunol. Scand. Sect. B Microbiol. 1986, 94, 399. [Google Scholar] [CrossRef]
- Yamamoto, T.; Zhou, X.; Williams, C.J.; Hochwalt, A.; Forney, L.J. Bacterial Populations in the Vaginas of Healthy Adolescent Women. J. Pediatr. Adolesc. Gynecol. 2009, 22, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Dols, J.A.M.; Molenaar, D.; van der Helm, J.J.; Caspers, M.P.M.; de Kat Angelino-Bart, A.; Schuren, F.H.J.; Speksnijder, A.G.C.L.; Westerhoff, H.V.; Richardus, J.H.; Boon, M.E.; et al. Molecular Assessment of Bacterial Vaginosis by Lactobacillus Abundance and Species Diversity. BMC Infect. Dis. 2016, 16, 180. [Google Scholar] [CrossRef]
- Dols, J.A.M.; Smit, P.W.; Kort, R.; Reid, G.; Schuren, F.H.J.; Tempelman, H.; Bontekoe, T.R.; Korporaal, H.; Boon, M.E. Microarray-Based Identification of Clinically Relevant Vaginal Bacteria in Relation to Bacterial Vaginosis. Am. J. Obstet. Gynecol. 2011, 204, 305.e1–305.e7. [Google Scholar] [CrossRef] [PubMed]
- Muzny, C.A.; Łaniewski, P.; Schwebke, J.R.; Herbst-Kralovetz, M.M. Host–Vaginal Microbiota Interactions in the Pathogenesis of Bacterial Vaginosis. Curr. Opin. Infect. Dis. 2020, 33, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Martin, D.H.; Marrazzo, J.M. The Vaginal Microbiome: Current Understanding and Future Directions. J. Infect. Dis. 2016, 214, S36–S41. [Google Scholar] [CrossRef]
- Muzny, C.A.; Taylor, C.M.; Swords, W.E.; Tamhane, A.; Chattopadhyay, D.; Cerca, N.; Schwebke, J.R. An Updated Conceptual Model on the Pathogenesis of Bacterial Vaginosis. J. Infect. Dis. 2019, 220, 1399–1405. [Google Scholar] [CrossRef]
- Doerflinger, S.Y.; Throop, A.L.; Herbst-Kralovetz, M.M. Bacteria in the Vaginal Microbiome Alter the Innate Immune Response and Barrier Properties of the Human Vaginal Epithelia in a Species-Specific Manner. J. Infect. Dis. 2014, 209, 1989–1999. [Google Scholar] [CrossRef]
- Schwebke, J.R.; Weiss, H.L. Interrelationships of Bacterial Vaginosis and Cervical Inflammation. Sex. Transm. Dis. 2002, 29, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, C.; Marrazzo, J. Bacterial Vaginosis and the Cervicovaginal Immune Response. Am. J. Reprod. Immunol. 2014, 71, 555–563. [Google Scholar] [CrossRef]
- Belkaid, Y.; Hand, T.W. Role of the Microbiota in Immunity and Inflammation. Cell 2014, 157, 121–141. [Google Scholar] [CrossRef] [PubMed]
- Blander, J.M.; Longman, R.S.; Iliev, I.D.; Sonnenberg, G.F.; Artis, D. Regulation of Inflammation by Microbiota Interactions with the Host. Nat. Immunol. 2017, 18, 851–861. [Google Scholar] [CrossRef]
- Turovskiy, Y.; Noll, K.S.; Chikindas, M.L. THE ETIOLOGY OF BACTERIAL VAGINOSIS. J. Appl. Microbiol. 2011, 110, 1105–1128. [Google Scholar] [CrossRef] [PubMed]
- Møller, B.R.; Kristiansen, F.V.; Thorsen, P.; Frost, L. Sterility of the Uterine Cavity. Acta Obstet. Gynecol. Scand. 1995, 74, 216. [Google Scholar] [CrossRef]
- Tai, F.-W.; Chang, C.Y.-Y.; Chiang, J.-H.; Lin, W.-C.; Wan, L. Association of Pelvic Inflammatory Disease with Risk of Endometriosis: A Nationwide Cohort Study Involving 141,460 Individuals. J. Clin. Med. 2018, 7, 379. [Google Scholar] [CrossRef]
- Ata, B.; Yildiz, S.; Turkgeldi, E.; Brocal, V.P.; Dinleyici, E.C.; Moya, A.; Urman, B. The Endobiota Study: Comparison of Vaginal, Cervical and Gut Microbiota Between Women with Stage 3/4 Endometriosis and Healthy Controls. Sci. Rep. 2019, 9, 2204. [Google Scholar] [CrossRef]
- Walther-António, M.R.S.; Chen, J.; Multinu, F.; Hokenstad, A.; Distad, T.J.; Cheek, E.H.; Keeney, G.L.; Creedon, D.J.; Nelson, H.; Mariani, A.; et al. Potential Contribution of the Uterine Microbiome in the Development of Endometrial Cancer. Available online: https://doaj.org (accessed on 14 January 2021).
- Khan, K.N.; Fujishita, A.; Masumoto, H.; Muto, H.; Kitajima, M.; Masuzaki, H.; Kitawaki, J. Molecular Detection of Intrauterine Microbial Colonization in Women with Endometriosis. Eur. J. Obstet. Gynecol. Reprod. Biol. 2016, 199, 69–75. [Google Scholar] [CrossRef]
- Hernandes, C.; Silveira, P.; Sereia, A.F.R.; Christoff, A.P.; Mendes, H.; de Oliveira, L.F.V.; Podgaec, S. Microbiome Profile of Deep Endometriosis Patients: Comparison of Vaginal Fluid, Endometrium and Lesion. Available online: https://doaj.org (accessed on 19 January 2021).
- Wei, W.; Zhang, X.; Tang, H.; Zeng, L.; Wu, R. Microbiota Composition and Distribution along the Female Reproductive Tract of Women with Endometriosis. Ann. Clin. Microbiol. Antimicrob. 2020, 19, 15. [Google Scholar] [CrossRef]
- Yuan, M.; Li, D.; Zhang, Z.; Sun, H.; An, M.; Wang, G. Endometriosis Induces Gut Microbiota Alterations in Mice. Hum. Reprod. 2018, 33, 607–616. [Google Scholar] [CrossRef] [PubMed]
- Bailey, M.T.; Coe, C.L. Endometriosis Is Associated with an Altered Profile of Intestinal Microflora in Female Rhesus Monkeys. Hum. Reprod. 2002, 17, 1704–1708. [Google Scholar] [CrossRef]
- Chadchan, S.B.; Cheng, M.; Parnell, L.A.; Yin, Y.; Schriefer, A.; Mysorekar, I.U.; Kommagani, R. Antibiotic Therapy with Metronidazole Reduces Endometriosis Disease Progression in Mice: A Potential Role for Gut Microbiota. Hum. Reprod. 2019, 34, 1106–1116. [Google Scholar] [CrossRef] [PubMed]
- Missmer, S.A.; Chavarro, J.E.; Malspeis, S.; Bertone-Johnson, E.R.; Hornstein, M.D.; Spiegelman, D.; Barbieri, R.L.; Willett, W.C.; Hankinson, S.E. A Prospective Study of Dietary Fat Consumption and Endometriosis Risk. Hum. Reprod. 2010, 25, 1528–1535. [Google Scholar] [CrossRef]
- Hopeman, M.M.; Riley, J.K.; Frolova, A.I.; Jiang, H.; Jungheim, E.S. Serum Polyunsaturated Fatty Acids and Endometriosis. Reprod. Sci. 2015, 22, 1083–1087. [Google Scholar] [CrossRef]
- Tomio, K.; Kawana, K.; Taguchi, A.; Isobe, Y.; Iwamoto, R.; Yamashita, A.; Kojima, S.; Mori, M.; Nagamatsu, T.; Arimoto, T.; et al. Omega-3 Polyunsaturated Fatty Acids Suppress the Cystic Lesion Formation of Peritoneal Endometriosis in Transgenic Mouse Models. PLoS ONE 2013, 8, e73085. [Google Scholar] [CrossRef]
- Attaman, J.A.; Stanic, A.K.; Kim, M.; Lynch, M.P.; Rueda, B.R.; Styer, A.K. The Anti-Inflammatory Impact of Omega-3 Polyunsaturated Fatty Acids During the Establishment of Endometriosis-Like Lesions. Am. J. Reprod. Immunol. 2014, 72, 392–402. [Google Scholar] [CrossRef]
- Ilich, J.Z.; Kelly, O.J.; Kim, Y.; Spicer, M.T. Low-Grade Chronic Inflammation Perpetuated by Modern Diet as a Promoter of Obesity and Osteoporosis. Arch. Ind. Hyg. Toxicol. 2014, 65, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Kelly, O.J.; Gilman, J.C.; Kim, Y.; Ilich, J.Z. Long-Chain Polyunsaturated Fatty Acids May Mutually Benefit Both Obesity and Osteoporosis. Nutr. Res. 2013, 33, 521–533. [Google Scholar] [CrossRef] [PubMed]
- Khan, K.N.; Fujishita, A.; Hiraki, K.; Kitajima, M.; Nakashima, M.; Fushiki, S.; Kitawaki, J. Bacterial Contamination Hypothesis: A New Concept in Endometriosis. Reprod. Med. Biol. 2018, 17, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Khan, K.N.; Kitajima, M.; Hiraki, K.; Yamaguchi, N.; Katamine, S.; Matsuyama, T.; Nakashima, M.; Fujishita, A.; Ishimaru, T.; Masuzaki, H. Escherichia Coli Contamination of Menstrual Blood and Effect of Bacterial Endotoxin on Endometriosis. Fertil. Steril. 2010, 94, 2860–2863.e3. [Google Scholar] [CrossRef] [PubMed]
- Emani, R.; Alam, C.; Pekkala, S.; Zafar, S.; Emani, M.R.; Hänninen, A. Peritoneal Cavity Is a Route for Gut-Derived Microbial Signals to Promote Autoimmunity in Non-Obese Diabetic Mice. Scand. J. Immunol. 2015, 81, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, I.I.; Atarashi, K.; Manel, N.; Brodie, E.L.; Shima, T.; Karaoz, U.; Wei, D.; Goldfarb, K.C.; Santee, C.A.; Lynch, S.V.; et al. Induction of Intestinal Th17 Cells by Segmented Filamentous Bacteria. Cell 2009, 139, 485–498. [Google Scholar] [CrossRef]
- Bedaiwy, M.A.; Falcone, T.; Sharma, R.K.; Goldberg, J.M.; Attaran, M.; Nelson, D.R.; Agarwal, A. Prediction of Endometriosis with Serum and Peritoneal Fluid Markers: A Prospective Controlled Trial. Hum. Reprod. 2002, 17, 426–431. [Google Scholar] [CrossRef]
- Bedaiwy, M.A.; El-Nashar, S.A.; Sharma, R.K.; Falcone, T. Effect of Ovarian Involvement on Peritoneal Fluid Cytokine Concentrations in Endometriosis Patients. Reprod. Biomed. Online 2007, 14, 620–625. [Google Scholar] [CrossRef]
- Bedaiwy, M.A.; Falcone, T. Peritoneal Fluid Environment in Endometriosis. Minerva Ginecol. 2003, 55, 13. [Google Scholar]
- Plaza, M.A.; Fioramonti, J.; Bueno, L. Role of Central Interleukin-1β in Gastrointestinal Motor Disturbances Induced by Lipopolysaccharide in Sheep. Dig. Dis. Sci. 1997, 42, 242–250. [Google Scholar] [CrossRef]
- Taché, Y.; Saperas, E. Potent Inhibition of Gastric Acid Secretion and Ulcer Formation by Centrally and Peripherally Administered Interleukin-1a. Ann. N. Y. Acad. Sci. 1992, 664, 353–368. [Google Scholar] [CrossRef]
- Flores, R.; Shi, J.; Fuhrman, B.; Xu, X.; Veenstra, T.D.; Gail, M.H.; Gajer, P.; Ravel, J.; Goedert, J.J. Fecal Microbial Determinants of Fecal and Systemic Estrogens and Estrogen Metabolites: A Cross-Sectional Study. J. Transl. Med. 2012, 10, 253. [Google Scholar] [CrossRef] [PubMed]
- Plottel, C.S.; Blaser, M.J. Microbiome and Malignancy. Cell Host Microbe 2011, 10, 324–335. [Google Scholar] [CrossRef] [PubMed]
- Flores, R.; Shi, J.; Gail, M.H.; Gajer, P.; Ravel, J.; Goedert, J.J. Association of Fecal Microbial Diversity and Taxonomy with Selected Enzymatic Functions. PLoS ONE 2012, 7, e39745. [Google Scholar] [CrossRef] [PubMed]
- Gloux, K.; Berteau, O.; Oumami, H.E.; Béguet, F.; Leclerc, M.; Doré, J. A Metagenomic β-Glucuronidase Uncovers a Core Adaptive Function of the Human Intestinal Microbiome. Proc. Natl. Acad. Sci. USA 2011, 108, 4539–4546. [Google Scholar] [CrossRef] [PubMed]
- Dalile, B.; Van Oudenhove, L.; Vervliet, B.; Verbeke, K. The Role of Short-Chain Fatty Acids in Microbiota–Gut–Brain Communication. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 461–478. [Google Scholar] [CrossRef] [PubMed]
- Paredes, S.; Cantillo, S.; Candido, K.D.; Knezevic, N.N. An Association of Serotonin with Pain Disorders and Its Modulation by Estrogens. Int. J. Mol. Sci. 2019, 20, 5729. [Google Scholar] [CrossRef]
- Kanasaki, H.; Tumurbaatar, T.; Oride, A.; Hara, T.; Okada, H.; Kyo, S. Gamma-aminobutyric AcidA Receptor Agonist, Muscimol, Increases KiSS-1 Gene Expression in Hypothalamic Cell Models. Reprod. Med. Biol. 2017, 16, 386–391. [Google Scholar] [CrossRef]
- Amabebe, E.; Anumba, D.O.C. The Vaginal Microenvironment: The Physiologic Role of Lactobacilli. Available online: https://doaj.org (accessed on 19 February 2021).
- Hufnagel, D.; Li, F.; Cosar, E.; Krikun, G.; Taylor, H. The Role of Stem Cells in the Etiology and Pathophysiology of Endometriosis. Semin. Reprod. Med. 2015, 33, 333–340. [Google Scholar] [CrossRef]
- Kwon, O.; Lee, S.; Kim, J.-H.; Kim, H.; Lee, S.-W. Altered Gut Microbiota Composition in Rag1-Deficient Mice Contributes to Modulating Homeostasis of Hematopoietic Stem and Progenitor Cells. Immune Netw. 2015, 15, 252–259. [Google Scholar] [CrossRef]
- Molina, N.M.; Sola-Leyva, A.; Saez-Lara, M.J.; Plaza-Diaz, J.; Tubić-Pavlović, A.; Romero, B.; Clavero, A.; Mozas-Moreno, J.; Fontes, J.; Altmäe, S. New Opportunities for Endometrial Health by Modifying Uterine Microbial Composition: Present or Future? Available online: https://doaj.org (accessed on 18 January 2021).
- Kyono, K.; Hashimoto, T.; Kikuchi, S.; Nagai, Y.; Sakuraba, Y. A Pilot Study and Case Reports on Endometrial Microbiota and Pregnancy Outcome: An Analysis Using 16S RRNA Gene Sequencing among IVF Patients, and Trial Therapeutic Intervention for Dysbiotic Endometrium. Reprod. Med. Biol. 2019, 18, 72–82. [Google Scholar] [CrossRef]
- Cicinelli, E.; Matteo, M.; Tinelli, R.; Pinto, V.; Marinaccio, M.; Indraccolo, U.; De Ziegler, D.; Resta, L. Chronic Endometritis due to Common Bacteria Is Prevalent in Women with Recurrent Miscarriage as Confirmed by Improved Pregnancy Outcome after Antibiotic Treatment. Reprod. Sci. 2014, 21, 640–647. [Google Scholar] [CrossRef] [PubMed]
- Cicinelli, E.; Matteo, M.; Trojano, G.; Mitola, P.C.; Tinelli, R.; Vitagliano, A.; Crupano, F.M.; Lepera, A.; Miragliotta, G.; Resta, L. Chronic Endometritis in Patients with Unexplained Infertility: Prevalence and Effects of Antibiotic Treatment on Spontaneous Conception. Am. J. Reprod. Immunol. 2018, 79, e12782. [Google Scholar] [CrossRef] [PubMed]
- McQueen, D.B.; Bernardi, L.A.; Stephenson, M.D. Chronic Endometritis in Women with Recurrent Early Pregnancy Loss and/or Fetal Demise. Fertil. Steril. 2014, 101, 1026–1030. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xu, H.; Liu, Y.; Zheng, S.; Zhao, W.; Wu, D.; Lei, L.; Chen, G. Confirmation of Chronic Endometritis in Repeated Implantation Failure and Success Outcome in IVF-ET after Intrauterine Delivery of the Combined Administration of Antibiotic and Dexamethasone. Am. J. Reprod. Immunol. 2019, 82, e13177. [Google Scholar] [CrossRef]
- Khodaverdi, S.; Mohammadbeigi, R.; Khaledi, M.; Mesdaghinia, L.; Sharifzadeh, F.; Nasiripour, S.; Gorginzadeh, M. Beneficial Effects of Oral Lactobacillus on Pain Severity in Women Suffering from Endometriosis: A Pilot Placebo-Controlled Randomized Clinical Trial. Int. J. Fertil. Steril. 2019, 13, 178–183. [Google Scholar] [CrossRef]
- Itoh, H.; Uchida, M.; Sashihara, T.; Ji, Z.-S.; Li, J.; Tang, Q.; Ni, S.; Song, L.; Kaminogawa, S. Lactobacillus Gasseri OLL2809 Is Effective Especially on the Menstrual Pain and Dysmenorrhea in Endometriosis Patients: Randomized, Double-Blind, Placebo-Controlled Study. Cytotechnology 2011, 63, 153–161. [Google Scholar] [CrossRef]
- Uchida, M.; Kobayashi, O. Effects of Lactobacillus Gasseri OLL2809 on the Induced Endometriosis in Rats. Biosci. Biotechnol. Biochem. 2013, 77, 1879–1881. [Google Scholar] [CrossRef] [PubMed]
- Itoh, H.; Sashihara, T.; Hosono, A.; Kaminogawa, S.; Uchida, M. Lactobacillus Gasseri OLL2809 Inhibits Development of Ectopic Endometrial Cell in Peritoneal Cavity via Activation of NK Cells in a Murine Endometriosis Model. Cytotechnology 2011, 63, 205–210. [Google Scholar] [CrossRef]
- Cao, Y.; Jiang, C.; Jia, Y.; Xu, D.; Yu, Y. Letrozole and the Traditional Chinese Medicine, Shaofu Zhuyu Decoction, Reduce Endometriotic Disease Progression in Rats: A Potential Role for Gut Microbiota. Evid. Based Complement. Altern. Med. 2020, 2020, 3687498. [Google Scholar] [CrossRef]
- Meng, J.; Meng, J.; Banerjee, S.; Banerjee, S.; Zhang, L.; Sindberg, G.; Moidunny, S.; Li, B.; Robbins, D.J.; Girotra, M.; et al. Opioids Impair Intestinal Epithelial Repair in HIV-Infected Humanized Mice. Available online: https://doaj.org (accessed on 20 January 2021).
- Llor, C.; Bjerrum, L. Antimicrobial resistance: Risk associated with antibiotic overuse and initiatives to reduce the problem. Ther Adv. Drug Saf. 2014, 5, 229–241. [Google Scholar] [CrossRef] [PubMed]
- Gottschick, C.; Deng, Z.L.; Vital, M.; Masur, C.; Abels, C.; Pieper, D.H.; Wagner-Döbler, I. The urinary microbiota of men and women and its changes in women during bacterial vaginosis and antibiotic treatment. Microbiome. 2017, 5, 99. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Chen, D.C. Facing a new challenge: The adverse effects of antibiotics on gut microbiota and host immunity. Chin. Med. J. 2019, 132, 1135–1138. [Google Scholar] [CrossRef] [PubMed]
- Perrotta, A.R.; Borrelli, G.M.; Martins, C.O.; Kallas, E.G.; Sanabani, S.S.; Griffith, L.G.; Alm, E.J.; Abrao, M.S. The Vaginal Microbiome as a Tool to Predict RASRM Stage of Disease in Endometriosis: A Pilot Study. Reprod. Sci. 2020, 27, 1064–1073. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Macrophages | Neutrophils | NK Cells | T Cells | B Cells | |
|---|---|---|---|---|---|
 |  |  |  |  | |
| Immune Dysregulation |
|
|
|
|
|
| Implications on Endometriosis |
|
|
|
|
|
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, I.; Yong, P.J.; Allaire, C.; Bedaiwy, M.A. Intricate Connections between the Microbiota and Endometriosis. Int. J. Mol. Sci. 2021, 22, 5644. https://doi.org/10.3390/ijms22115644
Jiang I, Yong PJ, Allaire C, Bedaiwy MA. Intricate Connections between the Microbiota and Endometriosis. International Journal of Molecular Sciences. 2021; 22(11):5644. https://doi.org/10.3390/ijms22115644
Chicago/Turabian StyleJiang, Irene, Paul J. Yong, Catherine Allaire, and Mohamed A. Bedaiwy. 2021. "Intricate Connections between the Microbiota and Endometriosis" International Journal of Molecular Sciences 22, no. 11: 5644. https://doi.org/10.3390/ijms22115644
APA StyleJiang, I., Yong, P. J., Allaire, C., & Bedaiwy, M. A. (2021). Intricate Connections between the Microbiota and Endometriosis. International Journal of Molecular Sciences, 22(11), 5644. https://doi.org/10.3390/ijms22115644