
Exposure to RF-EMF Alters Postsynaptic Structure and Hinders Neurite Outgrowth in Developing Hippocampal Neurons of Early Postnatal Mice


Abstract
1. Introduction
2. Results
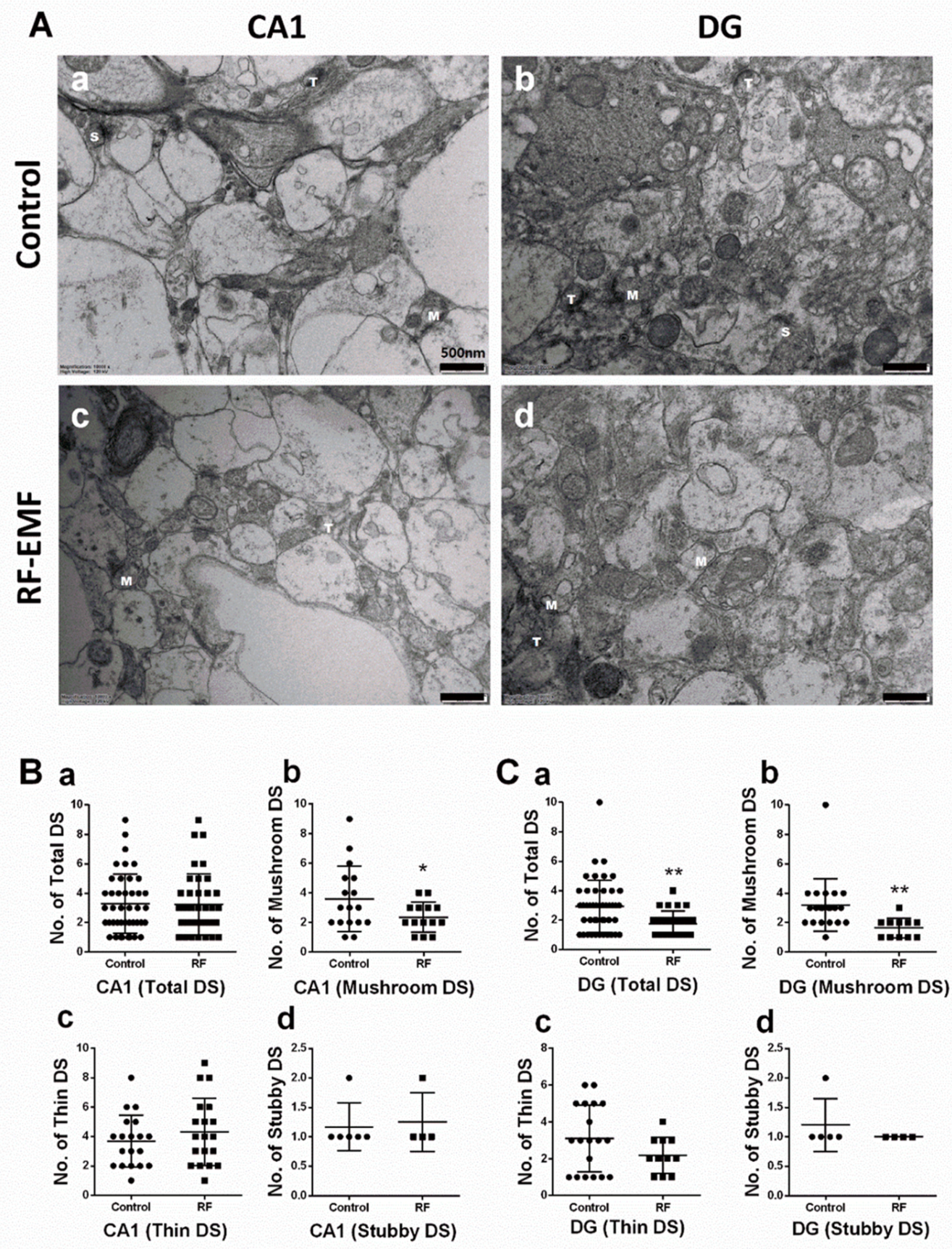
2.1. Exposure to RF-EMF Decreases the Number of DSs in Hippocampal Neurons
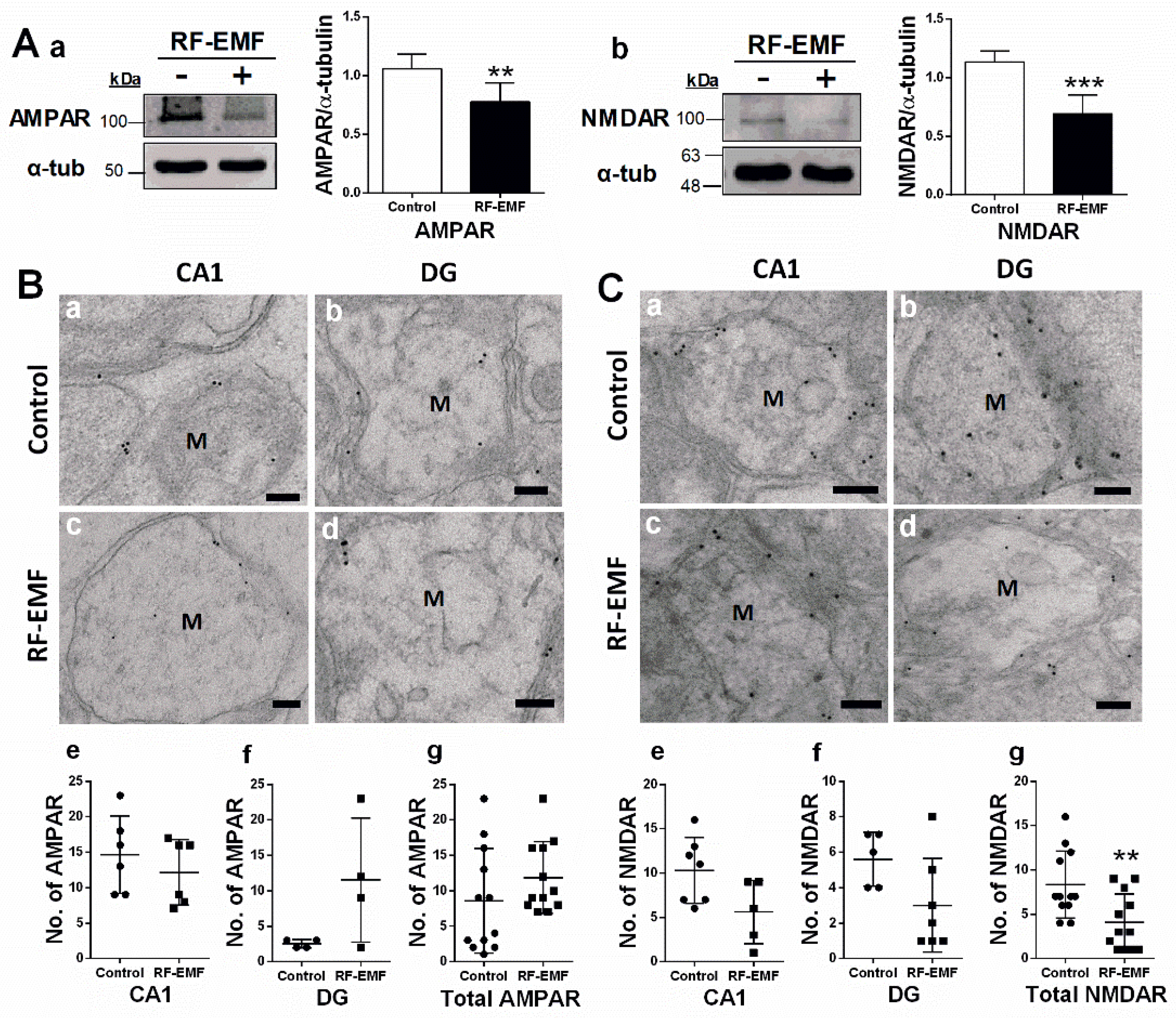
2.2. Glutamate Receptor Expression in Hippocampal Neurons Is Decreased Following RF-EMF Exposure
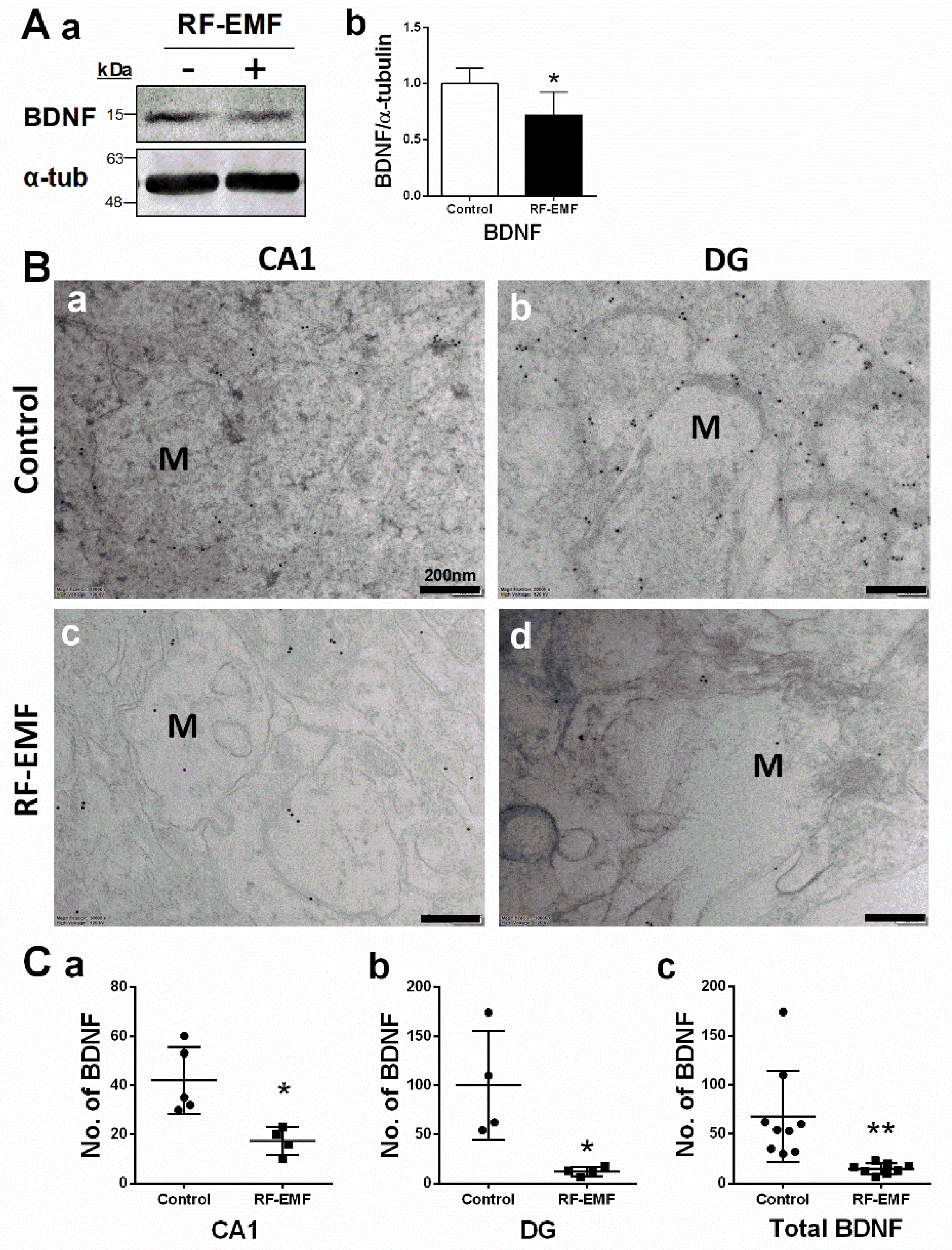
2.3. Brain-Derived Neurotrophic Factor (BDNF) Expression in Hippocampal Neurons Is Decreased Following RF-EMF Exposure
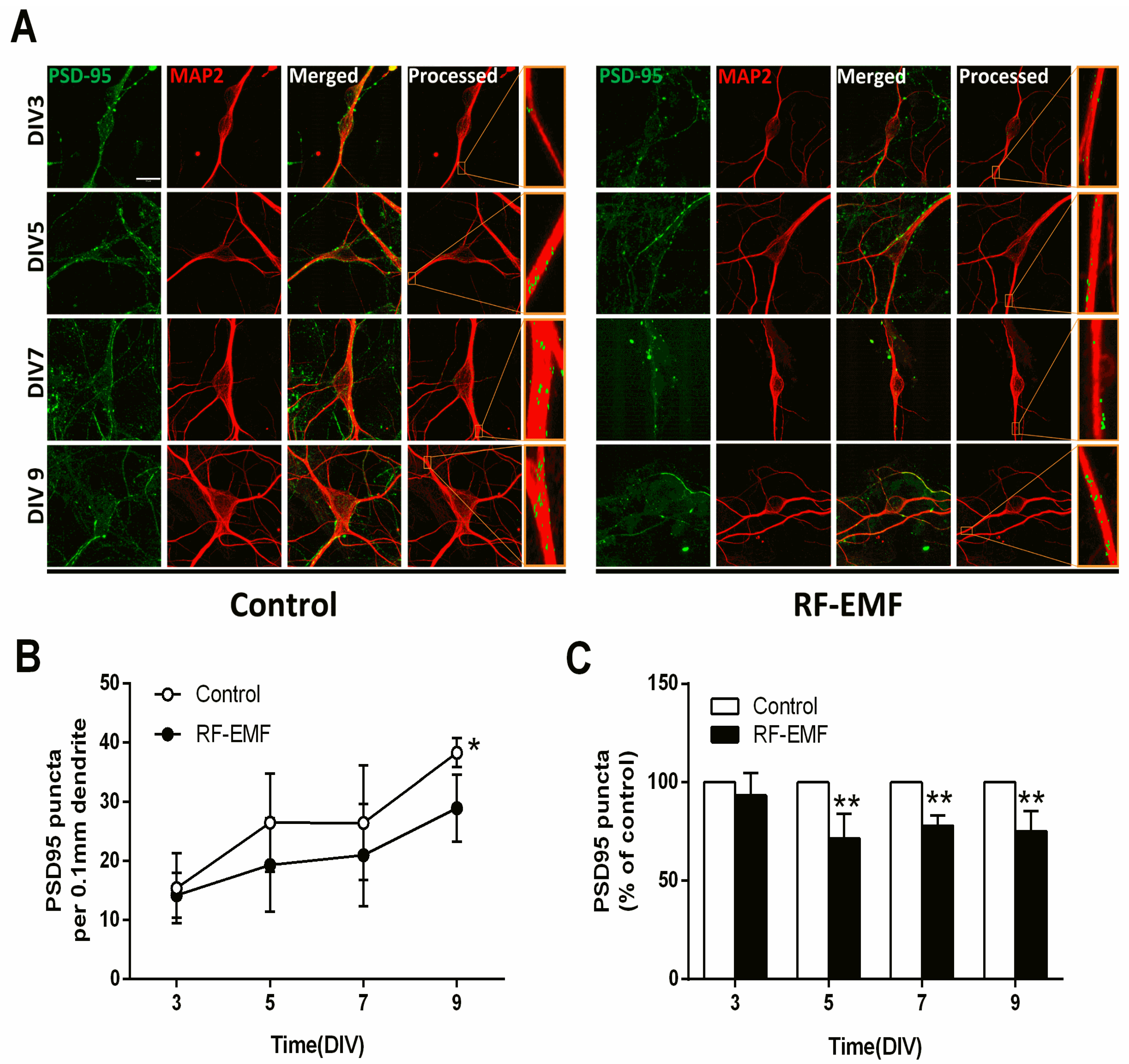
2.4. Synaptic Density in Hippocampal Neurons Is Decreased Following Exposure to RF-EMF
2.5. Glutamate Receptor Expression in Primary Cultured Hippocampal Neurons Is Decreased Following RF-EMF Exposure
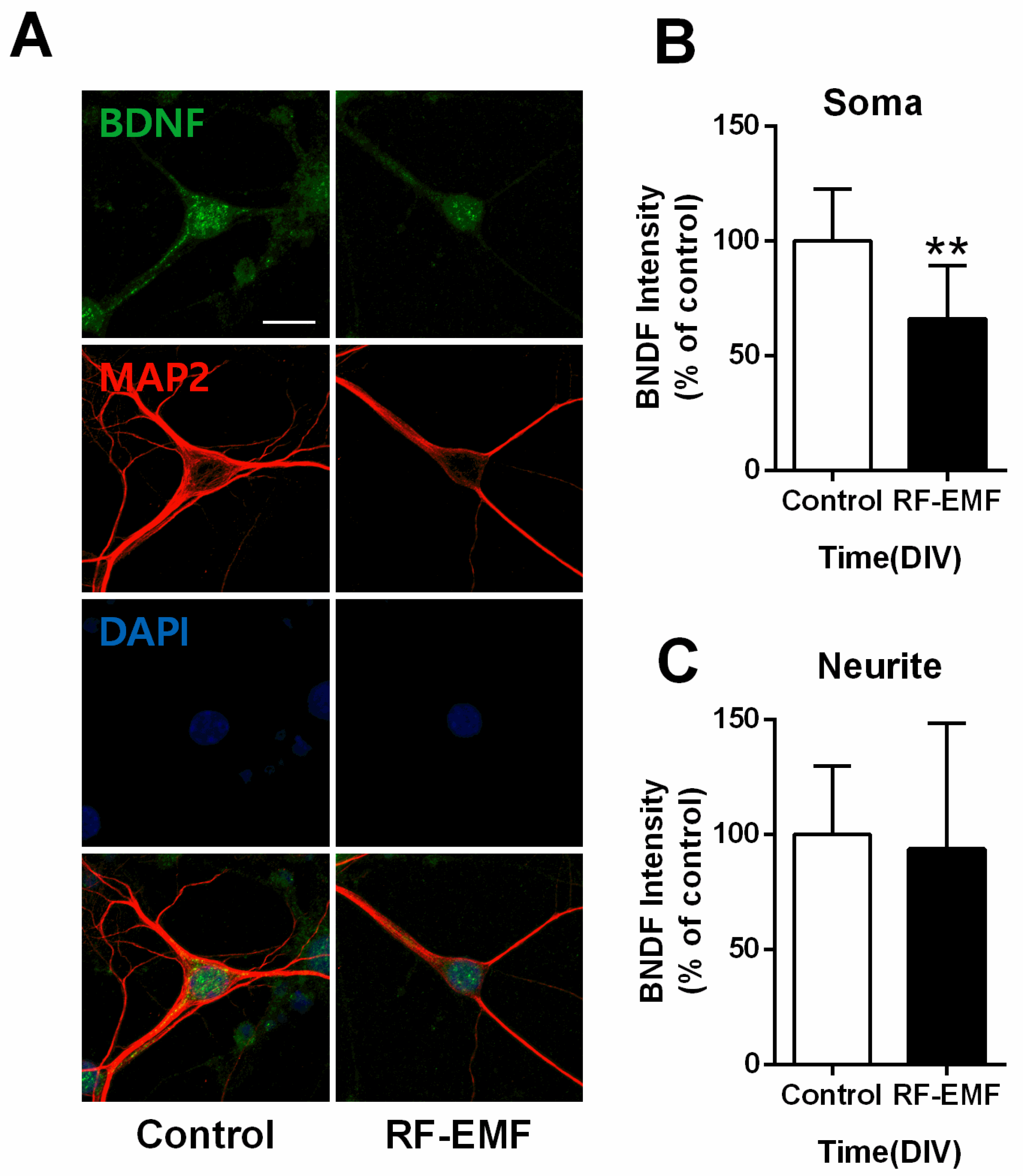
2.6. BDNF Expression Is Decreased at the Soma but Not Neurites of Hippocampal Neurons Following RF-EMF Exposure
2.7. Neurite Outgrowth of Hippocampal Neurons Is Affected by RF-EMF Exposure
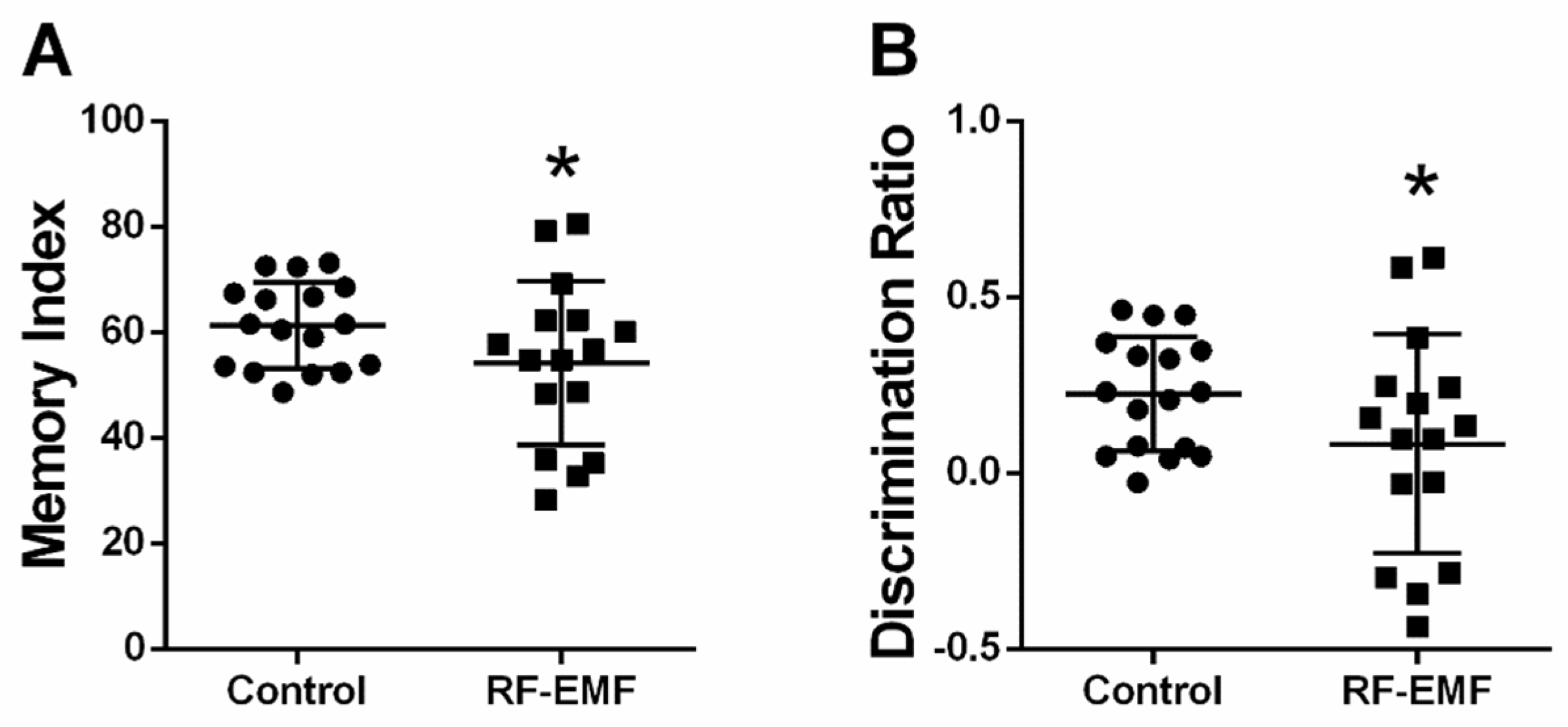
2.8. Memory Index and Discrimination Ratio Is Decreased Following RF-EMF Exposure
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. RF-EMF Exposure in Mice
4.3. Primary Cultures of Mouse Hippocampal Neurons
4.4. RF-EMF Exposure in Cultured Neurons
4.5. Transmission Electron Microscopy (TEM)
4.6. Immunogold Staining
4.7. Immunocytochemistry
4.8. Western Blot
4.9. Cellular Imaging and Morphological Analysis
4.10. Novel Object Recognition Test
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Measure | Control | RF-EMF | p | Cohen’s d | ||
|---|---|---|---|---|---|---|
| M | SD | M | SD | |||
| Figure 1Bb | 3.59 | 2.21 | 2.36 | 1.01 | 0.0391 | 0.71 |
| Figure 1Cb | 3.19 | 1.78 | 1.64 | 0.67 | 0.0093 | 1.03 |
| Figure 1Ca | 2.93 | 1.79 | 1.77 | 0.86 | 0.0027 | 0.77 |
| Figure 2Cg | 8.33 | 3.77 | 4.08 | 3.20 | 0.0070 | 1.21 |
| Figure 3Ca | 42.00 | 13.58 | 17.25 | 5.62 | 0.0117 | 2.27 |
| Figure 3Cb | 100.00 | 55.18 | 12.00 | 4.55 | 0.0191 | 2.25 |
| Figure 3Cc | 67.78 | 46.57 | 14.63 | 5.50 | 0.0060 | 1.55 |
| Figure 8A | 61.30 | 8.14 | 54.17 | 15.55 | 0.0389 | 0.58 |
| Figure 8B | 0.23 | 0.16 | 0.083 | 0.311 | 0.0389 | 0.58 |
References
- Christ, A.; Gosselin, M.-C.; Christopoulou, M.; Kühn, S.; Kuster, N. Age-dependent tissue-specific exposure of cell phone users. Phys. Med. Biol. 2010, 55, 1767–1783. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Sohn, U.D.; Kim, H.G.; Kim, H.R. Exposure to 835 MHz RF-EMF decreases the expression of calcium channels, inhibits apoptosis, but induces autophagy in the mouse hippocampus. Korean J. Physiol. Pharmacol. 2018, 22, 277–289. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Yu, D.H.; Huh, Y.H.; Lee, E.H.; Kim, H.G.; Kim, H.R. Long-term exposure to 835 MHz RF-EMF induces hyperactivity, autophagy and demyelination in the cortical neurons of mice. Sci. Rep. 2017, 7, 41129. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Lee, C.H.; Kim, H.G.; Kim, H.R. Decreased dopamine in striatum and difficult locomotor recovery from MPTP insult after exposure to radiofrequency electromagnetic fields. Sci. Rep. 2019, 9, 1201. [Google Scholar] [CrossRef]
- Birks, L.E.; Struchen, B.; Eeftens, M.; van Wel, L.; Huss, A.; Gajšek, P.; Kheifets, L.; Gallastegi, M.; Dalmau-Bueno, A.; Estarlich, M.; et al. Spatial and temporal variability of personal environmental exposure to radio frequency electromagnetic fields in children in Europe. Environ. Int. 2018, 117, 204–214. [Google Scholar] [CrossRef] [PubMed]
- Foster, K.R.; Chou, C.-K. Are Children More Exposed to Radio Frequency Energy From Mobile Phones Than Adults? IEEE Access 2014, 2, 1497–1509. [Google Scholar] [CrossRef]
- Gandhi, O.P.; Morgan, L.L.; De Salles, A.A.; Han, Y.-Y.; Herberman, R.B.; Davis, D.L. Exposure Limits: The underestimation of absorbed cell phone radiation, especially in children. Electromagn. Biol. Med. 2011, 31, 34–51. [Google Scholar] [CrossRef]
- Kim, J.H.; Huh, Y.H.; Lee, J.-H.; Jung, J.Y.; Ahn, S.C.; Kim, H.R. Early exposure to radiofrequency electromagnetic fields at 1850 MHz affects auditory circuits in early postnatal mice. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef]
- Konkel, A.; Cohen, N.J. Relational memory and the hippocampus: Representations and methods. Front. Neurosci. 2009, 3, 23. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.; Aimone, J.B.; Gage, F.H. New neurons and new memories: How does adult hippocampal neurogenesis affect learning and memory? Nat. Rev. Neurosci. 2010, 11, 339–350. [Google Scholar] [CrossRef]
- Eichenbaum, H.; Cohen, N.J. Can We Reconcile the Declarative Memory and Spatial Navigation Views on Hippocampal Function? Neuron 2014, 83, 764–770. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zang, Y.; He, Y.; Liang, M.; Zhang, X.; Tian, L.; Wu, T.; Jiang, T.; Li, K. Changes in hippocampal connectivity in the early stages of Alzheimer’s disease: Evidence from resting state fMRI. Neuroimage 2006, 31, 496–504. [Google Scholar] [CrossRef] [PubMed]
- Del Campo, M.; Hoozemans, J.J.; Dekkers, L.-L.; Rozemuller, A.J.; Korth, C.; Müller-Schiffmann, A.; Scheltens, P.; Blankenstein, M.A.; Jimenez, C.R.; Veerhuis, R. BRI2-BRICHOS is increased in human amyloid plaques in early stages of Alzheimer’s disease. Neurobiol. Aging 2014, 35, 1596–1604. [Google Scholar] [CrossRef] [PubMed]
- Campbell, S.; MacQueen, G. The role of the hippocampus in the pathophysiology of major depression. J. Psychiatr. Neurosci. 2004, 29, 417–426. [Google Scholar]
- Videbech, P.; Ravnkilde, B. Hippocampal volume and depression: A meta-analysis of MRI studies. Am. J. Psychiatr. 2004, 161, 1957–1966. [Google Scholar] [CrossRef] [PubMed]
- Morris, R.G.M.; Moser, E.I.; Riedel, G.; Martin, S.J.; Sandin, J.; Day, M.; O’Carroll, C. Elements of a neurobiological theory of the hippocampus: The role of activity-dependent synaptic plasticity in memory. Philos. Trans. R. Soc. B Biol. Sci. 2003, 358, 773–786. [Google Scholar] [CrossRef] [PubMed]
- Robinson, P.J. How to fill a synapse. Science 2007, 316, 551–553. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Kim, H.-J.; Yu, D.-H.; Kweon, H.-S.; Huh, Y.H.; Kim, H.R. Changes in numbers and size of synaptic vesicles of cortical neurons induced by exposure to 835 MHz radiofrequency-electromagnetic field. PLoS ONE 2017, 12, e0186416. [Google Scholar] [CrossRef]
- Kim, J.H.; Lee, J.-K.; Kim, H.-G.; Kim, K.-B.; Kim, H.R. Possible Effects of Radiofrequency Electromagnetic Field Exposure on Central Nerve System. Biomol. Ther. 2019, 27, 265–275. [Google Scholar] [CrossRef]
- Kim, J.H.; Huh, Y.H.; Kim, H.R. Trafficking of synaptic vesicles is changed at the hypothalamus by exposure to an 835 MHz radiofrequency electromagnetic field. Gen. Physiol. Biophys. 2019, 38, 379–388. [Google Scholar] [CrossRef] [PubMed]
- Südhof, T.C. The synaptic vesicle cycle. Annu. Rev. Neurosci. 2004, 27, 509–547. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Miller, E.C.; Pozzo-Miller, L. Dendritic spine dysgenesis in Rett syndrome. Front. Neuroanat. 2014, 8, 97. [Google Scholar] [CrossRef]
- Bourne, J.; Harris, K.M. Do thin spines learn to be mushroom spines that remember? Curr. Opin. Neurobiol. 2007, 17, 381–386. [Google Scholar] [CrossRef] [PubMed]
- Newpher, T.M.; Ehlers, M.D. Glutamate Receptor Dynamics in Dendritic Microdomains. Neuron 2008, 58, 472–497. [Google Scholar] [CrossRef]
- Debanne, D.; Daoudal, G.; Sourdet, V.; Russier, M. Brain plasticity and ion channels. J. Physiol. 2003, 97, 403–414. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, A.L.; Matthews, B.J.; Meynard, M.M.; Hu, B.; Javed, S.; Cohen-Cory, S. BDNF increases synapse density in dendrites of developing tectal neurons in vivo. Development 2006, 133, 2477–2486. [Google Scholar] [CrossRef]
- Koshimizu, H.; Kiyosue, K.; Hara, T.; Hazama, S.; Suzuki, S.; Uegaki, K.; Nagappan, G.; Zaitsev, E.; Hirokawa, T.; Tatsu, Y.; et al. Multiple functions of precursor BDNF to CNS neurons: Negative regulation of neurite growth, spine formation and cell survival. Mol. Brain 2009, 2, 27. [Google Scholar] [CrossRef] [PubMed]
- Harward, S.C.; Hedrick, N.G.; Hall, C.E.; Parra-Bueno, P.; Milner, T.A.; Pan, E.; Laviv, T.; Hempstead, B.L.; Yasuda, R.; McNamara, J.O. Autocrine BDNF-TrkB signalling within a single dendritic spine. Nature 2016, 538, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Von Bohlen und Halbach, O.; von Bohlen und Halbach, V. BDNF effects on dendritic spine morphology and hippocampal function. Cell Tissue Res. 2018, 373, 729–741. [Google Scholar] [CrossRef] [PubMed]
- Holmes, J.R.; Berkowitz, A. Dendritic orientation and branching distinguish a class of multifunctional turtle spinal interneurons. Front. Neural Circuits 2014, 8, 136. [Google Scholar] [CrossRef] [PubMed]
- Southam, K.A.; Stennard, F.; Pavez, C.; Small, D.H. Knockout of amyloid β protein precursor (APP) expression alters synaptogenesis, neurite branching and axonal morphology of hippocampal neurons. Neurochem. Res. 2019, 44, 1346–1355. [Google Scholar] [CrossRef]
- Huang, E.J.; Reichardt, L.F. Neurotrophins: Roles in Neuronal Development and Function. Annu. Rev. Neurosci. 2001, 24, 677–736. [Google Scholar] [CrossRef] [PubMed]
- Hering, H.; Sheng, M. Dentritic spines: Structure, dynamics and regulation. Nat. Rev. Neurosci. 2001, 2, 880–888. [Google Scholar] [CrossRef] [PubMed]
- Schikorski, T.; Stevens, C.F. Quantitative Ultrastructural Analysis of Hippocampal Excitatory Synapses. J. Neurosci. 1997, 17, 5858–5867. [Google Scholar] [CrossRef] [PubMed]
- Nusser, Z.; Lujan, R.; Laube, G.; Roberts, J.B.; Molnar, E.; Somogyi, P. Cell Type and Pathway Dependence of Synaptic AMPA Receptor Number and Variability in the Hippocampus. Neuron 1998, 21, 545–559. [Google Scholar] [CrossRef]
- Xiong, J.; He, C.; Li, C.; Tan, G.; Li, J.; Yu, Z.; Hu, Z.; Chen, F. Changes of Dendritic Spine Density and Morphology in the Superficial Layers of the Medial Entorhinal Cortex Induced by Extremely Low-Frequency Magnetic Field Exposure. PLoS ONE 2013, 8, e83561. [Google Scholar] [CrossRef] [PubMed]
- Lang, C.; Barco, A.; Zablow, L.; Kandel, E.R.; Siegelbaum, S.A.; Zakharenko, S.S. Transient expansion of synaptically connected dendritic spines upon induction of hippocampal long-term potentiation. Proc. Nat. Acad. Sci. USA 2004, 101, 16665. [Google Scholar] [CrossRef]
- Matsuzaki, M.; Honkura, N.; Ellis-Davies, G.C.R.; Kasai, H. Structural basis of long-term potentiation in single dendritic spines. Nat. Cell Biol. 2004, 429, 761–766. [Google Scholar] [CrossRef] [PubMed]
- McEntee, W.J.; Crook, T.H. Glutamate: Its role in learning, memory, and the aging brain. Psychopharmacology 1993, 111, 391–401. [Google Scholar] [CrossRef] [PubMed]
- Henley, J.M.; Wilkinson, K.A. AMPA receptor trafficking and the mechanisms underlying synaptic plasticity and cognitive aging. Dialog. Clin. Neurosci. 2013, 15, 11–27. [Google Scholar]
- Takumi, Y.; Ramírez-León, V.; Laake, P.; Rinvik, E.; Ottersen, O.P. Different modes of expression of AMPA and NMDA receptors in hippocampal synapses. Nat. Neurosci. 1999, 2, 618–624. [Google Scholar] [CrossRef]
- Cunha, C.; Brambilla, R.; Thomas, K.L. A simple role for BDNF in learning and memory? Front. Mol. Neurosci. 2010, 3, 1. [Google Scholar] [CrossRef] [PubMed]
- Yoshii, A.; Constantine-Paton, M. BDNF induces transport of PSD-95 to dendrites through PI3K-AKT signaling after NMDA receptor activation. Nat. Neurosci. 2007, 10, 702–711. [Google Scholar] [CrossRef] [PubMed]
- Xiong, H.; Futamura, T.; Jourdi, H.; Zhou, H.; Takei, N.; Diverse-Pierluissi, M.; Plevy, S.; Nawa, H. Neurotrophins induce BDNF expression through the glutamate receptor pathway in neocortical neurons. Neuropharmacology 2002, 42, 903–912. [Google Scholar] [CrossRef][Green Version]
- Zheng, F.; Wang, H. NMDA-mediated and self-induced bdnf exon IV transcriptions are differentially regulated in cultured cortical neurons. Neurochem. Int. 2009, 54, 385–392. [Google Scholar] [CrossRef] [PubMed]
- Dotti, C.G.; Sullivan, C.A.; Banker, G.A. The establishment of polarity by hippocampal neurons in culture. J. Neurosci. 1988, 8, 1454–1468. [Google Scholar] [CrossRef]
- Virdee, J.K.; Saro, G.; Fouillet, A.; Findlay, J.; Ferreira, F.; Eversden, S.; O’Neill, M.J.; Wolak, J.; Ursu, D. A high-throughput model for investigating neuronal function and synaptic transmission in cultured neuronal networks. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef]
- El-Husseini, A.E.-D.; Schnell, E.; Chetkovich, D.M.; Nicoll, R.A.; Bredt, D.S. PSD-95 involvement in maturation of excitatory synapses. Science 2000, 290, 1364–1368. [Google Scholar] [PubMed]
- De Roo, M.; Klauser, P.; Mendez, P.; Poglia, L.; Muller, D. Activity-Dependent PSD Formation and Stabilization of Newly Formed Spines in Hippocampal Slice Cultures. Cereb. Cortex 2007, 18, 151–161. [Google Scholar] [CrossRef]
- Xu, S.; Ning, W.; Xu, Z.; Zhou, S.; Chiang, H.; Luo, J. Chronic exposure to GSM 1800-MHz microwaves reduces excitatory synaptic activity in cultured hippocampal neurons. Neurosci. Lett. 2006, 398, 253–257. [Google Scholar] [CrossRef] [PubMed]
- Almeida, M.F.; Chaves, R.S.; Silva, C.M.; Chaves, J.C.; Melo, K.P.; Ferrari, M.F. BDNF trafficking and signaling impairment during early neurodegeneration is prevented by moderate physical activity. IBRO Rep. 2016, 1, 19–31. [Google Scholar] [CrossRef] [PubMed]
- Harada, A.; Teng, J.; Takei, Y.; Oguchi, K.; Hirokawa, N. MAP2 is required for dendrite elongation, PKA anchoring in dendrites, and proper PKA signal transduction. J. Cell Biol. 2002, 158, 541–549. [Google Scholar] [CrossRef] [PubMed]
- Mozzachiodi, R.; Byrne, J.H. More than synaptic plasticity: Role of nonsynaptic plasticity in learning and memory. Trends Neurosci. 2010, 33, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Raven, F.; Van der Zee, E.A.; Meerlo, P.; Havekes, R. The role of sleep in regulating structural plasticity and synaptic strength: Implications for memory and cognitive function. Sleep Med. Rev. 2018, 39, 3–11. [Google Scholar] [CrossRef]
- Tavosanis, G. Dendritic structural plasticity. Dev. Neurobiol. 2011, 72, 73–86. [Google Scholar] [CrossRef] [PubMed]
- Penzes, P.; Cahill, M.E.; Jones, K.A.; VanLeeuwen, J.-E.; Woolfrey, K.M. Dendritic spine pathology in neuropsychiatric disorders. Nat. Neurosci. 2011, 14, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.J.; Munchow, A.H.; Rios, L.M.; Zhang, G.; Ásgeirsdóttir, H.N.; Stackman, R.W. The Rodent Hippocampus Is Essential for Nonspatial Object Memory. Curr. Biol. 2013, 23, 1685–1690. [Google Scholar] [CrossRef]
- Götz, J.; Ittner, L.M. Animal models of Alzheimer’s disease and frontotemporal dementia. Nat. Rev. Neurosci. 2008, 9, 532–544. [Google Scholar] [CrossRef] [PubMed]
- Ntzouni, M.P.; Skouroliakou, A.; Kostomitsopoulos, N.; Margaritis, L.H. Transient and cumulative memory impairments induced by GSM 1.8 GHz cell phone signal in a mouse model. Electromagn. Biol. Med. 2012, 32, 95–120. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Lai, J. Modelling the vibration behaviour of infinite structures by FEM. J. Sound Vib. 2000, 229, 453–466. [Google Scholar] [CrossRef]
- International Commission on Non-Ionizing Radiation Protection (ICNIRP). Guidelines for Limiting Exposure to Electromagnetic Fields (100 kHz to 300 GHz). Health Phys. 2020, 118, 483–524. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Min, K.; Jeon, S.; Kim, N.; Pack, J.-K.; Song, K. Continuous Exposure to 1.7 GHz LTE Electromagnetic Fields Increases Intracellular Reactive Oxygen Species to Decrease Human Cell Proliferation and Induce Senescence. Sci. Rep. 2020, 10, 1–15. [Google Scholar] [CrossRef]
- Kim, H.J.; Martemyanov, K.A.; Thayer, S.A. Human immunodeficiency virus protein Tat induces synapse loss via a reversible process that is distinct from cell death. J. Neurosci. 2008, 28, 12604–12613. [Google Scholar] [CrossRef]
- Baj, G.; Patrizio, A.; Montalbano, A.; Sciancalepore, M.; Tongiorgi, E. Developmental and maintenance defects in Rett syndrome neurons identified by a new mouse staging system in vitro. Front. Cell. Neurosci. 2014, 8, 18. [Google Scholar] [CrossRef] [PubMed]
- Waataja, J.J.; Kim, H.J.; Roloff, A.M.; Thayer, S.A. Excitotoxic loss of post-synaptic sites is distinct temporally and mechanistically from neuronal death. J. Neurochem. 2008, 104, 364–375. [Google Scholar] [CrossRef] [PubMed]
- Sivakumaran, M.H.; MacKenzie, A.K.; Callan, I.R.; Ainge, J.A.; O’Connor, A.R. The Discrimination Ratio derived from Novel Object Recognition tasks as a Measure of Recognition Memory Sensitivity, not Bias. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.H.; Chung, K.H.; Hwang, Y.R.; Park, H.R.; Kim, H.J.; Kim, H.-G.; Kim, H.R. Exposure to RF-EMF Alters Postsynaptic Structure and Hinders Neurite Outgrowth in Developing Hippocampal Neurons of Early Postnatal Mice. Int. J. Mol. Sci. 2021, 22, 5340. https://doi.org/10.3390/ijms22105340
Kim JH, Chung KH, Hwang YR, Park HR, Kim HJ, Kim H-G, Kim HR. Exposure to RF-EMF Alters Postsynaptic Structure and Hinders Neurite Outgrowth in Developing Hippocampal Neurons of Early Postnatal Mice. International Journal of Molecular Sciences. 2021; 22(10):5340. https://doi.org/10.3390/ijms22105340
Chicago/Turabian StyleKim, Ju Hwan, Kyung Hwun Chung, Yeong Ran Hwang, Hye Ran Park, Hee Jung Kim, Hyung-Gun Kim, and Hak Rim Kim. 2021. "Exposure to RF-EMF Alters Postsynaptic Structure and Hinders Neurite Outgrowth in Developing Hippocampal Neurons of Early Postnatal Mice" International Journal of Molecular Sciences 22, no. 10: 5340. https://doi.org/10.3390/ijms22105340
APA StyleKim, J. H., Chung, K. H., Hwang, Y. R., Park, H. R., Kim, H. J., Kim, H.-G., & Kim, H. R. (2021). Exposure to RF-EMF Alters Postsynaptic Structure and Hinders Neurite Outgrowth in Developing Hippocampal Neurons of Early Postnatal Mice. International Journal of Molecular Sciences, 22(10), 5340. https://doi.org/10.3390/ijms22105340