Safeguarding Female Reproductive Health Against Endocrine Disrupting Chemicals—The FREIA Project

 , ,
, ,  , , , , , ,
, , , , , , {kind=link}
{kind=link}
Abstract
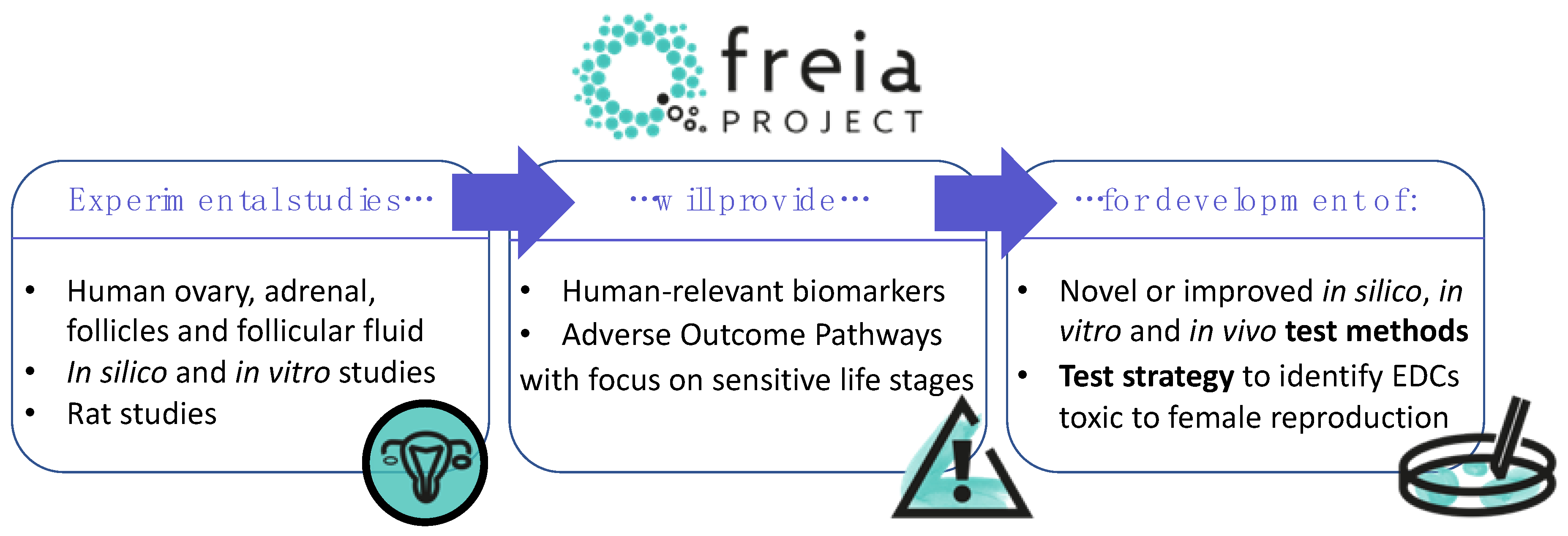
1. Introduction
2. EDC Exposure and Female Reproductive Disorders: The Current State of Knowledge
2.1. Female Reproductive Health
2.2. Characterisation of Susceptible Windows
2.3. In the Womb
2.4. Puberty
2.5. Adulthood
3. Advancing Test Methods for EDC Identification in Chemical Regulations
3.1. Mechanistic Descriptions of Female Reproductive Toxicity
3.2. Dedicated Endpoints and Bioassays to Address EDC-Related Female Reproductive Toxicity
- (1)
- Ovarian follicle counts performed in adult animals is included in selected OECD test guidelines, including the Extended One-Generation Reproductive Toxicity Study (TG 443). FREIA aims to improve follicle counting as currently recommended methods are highly affected by the size of the ovary and does not necessarily reflect the total follicle number.
- (2)
- Comparison of gene expression changes in key cell types in rat ovaries and in granulosa cells from human follicular fluids will help us to find biomarkers for oocyte quality, which are currently lacking in OECD test guidelines.
- (3)
- Morphological changes in early mammary development by image analysis of mammary gland whole mounts are considered sensitive markers of endocrine disruption in female rodents, yet is currently not part of any OECD test guideline. We will improve whole mount examination in the rat mammary gland. An unbiased and sensitive software tool will be applied for examination of mammary gland whole mounts collected at young age [40].
- (4)
- Puberty is a time with great hormonal changes, including a dramatic increase in DHEA over the adrenarche, yet this is largely overlooked in current test strategies dealing with EDCs. In FREIA we will evaluate the need for implementation of elements of the female pubertal assay, an established protocol (US EPA OPPTS 890.145), into current or future OECD test guidelines.
- (5)
- Changes in GnRH secretion preceding pubertal onset is considered a key event in ovarian dysfunction after EDC exposure. The effects of DES and ketoconazole on pulsatile GnRH release and hypothalamic transcriptional markers will be studied in primary rodent hypothalamic explants and compared with effects on GnRH release of in vivo exposed hypothalamus. This will allow the study of effects of prenatal exposure on the programming of maturation of GnRH secretion.
- (1)
- Quantitative structure-activity relationships (QSARs) allow us to predict molecular targets based on physico-chemical properties of a chemical. We will improve existing QSARs for aromatase and PPAR-gamma, two of the key events in a proposed adverse outcome pathway for female reproductive toxicity. These QSAR models and predictions will be made publicly available via the freely available DTU Danish (Q)SAR database (http://qsar.food.dtu.dk/) to open the possibility of integrating dedicated QSARs into toxicological risk assessment strategies.
- (2)
- Oestrogen receptor ER-beta (ESR2) plays a central role in ovarian development and function [41], yet concentration-effect data of EDCs on this receptor is limited. A high content analysis (HCA) assay utilizing an enhanced green-fluorescence tagged ER-beta nuclear receptor transfected into the U2OS cell line will be optimised and employed for screening of selected and human relevant EDCs over a concentration range for ER-beta concentration-response activity.
- (3)
- G-coupled protein oestrogen receptor (GPER) is suggested to act as selector during folliculogenesis [42] and may be involved in various ovarian pathologies [43]. GPER is also often more highly expressed in human fetal tissues than ER-alpha (ERS1) or ESR2. A novel high content analysis (HCA) assay for the GPER will be developed alongside HCA assays to assess subtle pre-lethal cytotoxicity in female reproductive cells.
- (4)
- It is not known whether the standardised steroidogenic assay OECD TG 456, which uses the adrenal corticocarcinoma cell line H295R and focuses only on progesterone, testosterone, and oestradiol synthesis, is sufficiently applicable to describe (interactions with) ovarian-specific steroidogenesis. This issue will be clarified by comparing steroidomic profiles obtained from human fetal ovarian and adrenal gland cultures, adult primary human ovarian tissue cultures, and human-derived granulosa cell lines.
- (5)
- For obvious ethical reasons, in vitro studies to assess human oocyte maturation and competency after fertilisation cannot be used for routine screening of EDCs in a regulatory context. In contrast, bovine ovarian tissues and follicles can easily be obtained from slaughterhouses. From these collected follicles, oocytes can be isolated and matured and used for in vitro fertilisation experiments, where embryonic development can be followed routinely until blastocyst hatching and elongation. Several studies have demonstrated the value of such bovine oocytes as experimental model for human reproductive parameters [44]. This test method adheres fully to the 3R-principle of toxicity testing in the 21st Century. Bovine oocyte studies will be performed to assess the effects of EDCs on oocyte maturation and the post-fertilisation competence to develop into a healthy embryo will be explored.
3.3. Testing for EDCs in the Regulatory Context
4. Sharing and Safeguarding
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AOP | Adverse outcome pathway |
| BPA | Bisphenol A |
| DDT | Dichlorodiphenyltrichloroethane |
| DEHP | Bis(2-ethylhexyl) phthalate |
| DES | Diethylstilbestrol |
| DHEA(S) | Dehydroepiandrosterone(-sulfate) |
| EATS | Estrogen-androgen-thyroid-steroidogenesis |
| EDC | Endocrine disrupting chemical |
| FREIA | “Female Reproductive toxicity of EDCs: a human evidence-based screening and Identification Approach” |
| FSH | Follicle stimulating hormone |
| GnRH | Gonadotropin-releasing hormone |
| GPER | G-coupled protein oestrogen receptor |
| HP(A)G axis | Hypothalamic-pituitary-(adrenal)-gonadal axis |
| IATA | Integrated approach to testing and assessment |
| LH | Luteinizing hormone |
| OECD | Organisation for Economic Cooperation and Development |
| PCOS | Polycystic ovary syndrome |
| POF | Premature ovarian failure |
| POI | Primary ovarian insufficiency |
| REACH | Registration, Evaluation, Authorisation and restriction of CHemicals |
| TG | Test guideline |
References
- WHO/UNEP. State of the Science of Endocrine Disrupting Chemicals—2012; World Health Organisation: Geneva, Switzerland, 2013. [Google Scholar]
- WHO/IPCS. Global Assessment of the State-of-the-Science of Endocrine Disruptors; International Programme on Chemical Safety; World Health Organisation and United Nations Environment Programme; World Health Organisation: Geneva, Switzerland, 2002. [Google Scholar]
- Toppari, J.; Virtanen, H.E.; Main, K.M.; Skakkebaek, N.E. Cryptorchidism and hypospadias as a sign of testicular dysgenesis syndrome (TDS): Environmental connection. Birth Defects Res. A Clin. Mol. Teratol. 2010, 88, 910–919. [Google Scholar] [CrossRef]
- Buck Louis, G.; Cooney, M.; Peterson, C. The ovarian dysgenesis syndrome. J. Dev. Orig. Health Dis. 2011, 2, 25–35. [Google Scholar] [CrossRef]
- OECD. Revised Guidance Document 150 on Standardised Test Guidelines for Evaluating Chemicals for Endocrine Disruption; OECD Series on Testing and Assessment, No. 150; OECD Publishing: Paris, France; Available online: https://doi.org/10.1787/9789264304741-en (accessed on 30 April 2020).
- Farquhar, C.M.; Bhattacharya, S.; Repping, S.; Mastenbroek, S.; Kamath, M.S.; Marjoribanks, J.; Boivin, J. Female subfertility. Nat. Rev. Dis Primers 2019, 5, 7. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Wen, S.; Yuan, D.; Peng, L.; Zeng, R.; Yang, Z.; Liu, Q.; Xu, L.; Kang, D. The association between the environmental endocrine disruptor bisphenol A and polycystic ovary syndrome: A systematic review and meta-analysis. Gynecol. Endocrinol. 2018, 34, 370–377. [Google Scholar] [CrossRef] [PubMed]
- Vagi, S.J.; Azziz-Baumgartner, E.; Sjodin, A.; Calafat, A.M.; Dumesic, D.; Gonzalez, L.; Kato, K.; Silva, M.J.; Ye, X.; Azziz, R. Exploring the potential association between brominated diphenyl ethers, polychlorinated biphenyls, organochlorine pesticides, perfluorinated compounds, phthalates, and bisphenol A in polycystic ovary syndrome: A case-control study. BMC Endocr. Disord. 2014, 14, 86. [Google Scholar] [CrossRef] [PubMed]
- Vabre, P.; Gatimel, N.; Moreau, J.; Gayrard, V.; Picard-Hagen, N.; Parinaud, J.; Leandri, R.D. Environmental pollutants, a possible etiology for premature ovarian insufficiency: A narrative review of animal and human data. Environ. Health 2017, 16, 37. [Google Scholar] [CrossRef]
- Viguie, C.; Mhaouty-Kodja, S.; Habert, R.; Chevrier, C.; Michel, C.; Pasquier, E. Evidence-based adverse outcome pathway approach for the identification of BPA as en endocrine disruptor in relation to its effect on the estrous cycle. Mol. Cell Endocrinol. 2018, 475, 10–28. [Google Scholar] [CrossRef]
- Conlon, J.L. Diethylstilbestrol: Potential health risks for women exposed in utero and their offspring. JAAPA 2017, 30, 49–52. [Google Scholar] [CrossRef]
- Titus, L.; Hatch, E.E.; Drake, K.M.; Parker, S.E.; Hyer, M.; Palmer, J.R.; Strohsnitter, W.C.; Adam, E.; Herbst, A.L.; Huo, D.; et al. Reproductive and hormone-related outcomes in women whose mothers were exposed in utero to diethylstilbestrol (DES): A report from the US National Cancer Institute DES Third Generation Study. Reprod. Toxicol. 2019, 84, 32–38. [Google Scholar] [CrossRef]
- Wallace, W.H.; Kelsey, T.W. Human ovarian reserve from conception to the menopause. PLoS ONE 2010, 5, e8772. [Google Scholar] [CrossRef]
- Wagner, M.; Yoshihara, M.; Douagi, I.; Damdimopoulos, A.; Panula, S.; Petropoulos, S.; Lu, H.; Pettersson, K.; Palm, K.; Katayama, S.; et al. Single-cell analysis of human ovarian cortex identifies distinct cell populations but no oogonial stem cells. Nat. Commun. 2020, 11, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Johansson, H.K.L.; Svingen, T.; Fowler, P.A.; Vinggaard, A.M.; Boberg, J. Environmental influences on ovarian dysgenesis—Developmental windows sensitive to chemical exposures. Nat. Rev. Endocrinol. 2017, 13, 400–414. [Google Scholar] [CrossRef]
- Carpenter, T.; Grecian, S.M.; Reynolds, R.M. Sex differences in early-life programming of the hypothalamic-pituitary-adrenal axis in humans suggest increased vulnerability in females: A systematic review. J. Dev. Orig. Health Dis. 2017, 8, 244–255. [Google Scholar] [CrossRef] [PubMed]
- Pinto, C.L.; Markey, K.; Dix, D.; Browne, P. Identification of candidate reference chemicals for in vitro steroidogenesis assays. Toxicol. Vitr. 2018, 47, 103–119. [Google Scholar] [CrossRef] [PubMed]
- Roelofs, M.J.; Piersma, A.H.; van den Berg, M.; van Duursen, M.B. The relevance of chemical interactions with CYP17 enzyme activity: Assessment using a novel in vitro assay. Toxicol. Appl. Pharm. 2013, 268, 309–317. [Google Scholar] [CrossRef] [PubMed]
- Franssen, D.; Ioannou, Y.S.; Alvarez-real, A.; Gerard, A.; Mueller, J.K.; Heger, S.; Bourguignon, J.P.; Parent, A.S. Pubertal timing after neonatal diethylstilbestrol exposure in female rats: Neuroendocrine vs peripheral effects and additive role of prenatal food restriction. Reprod. Toxicol. 2014, 44, 63–72. [Google Scholar] [CrossRef]
- Franssen, D.; Gerard, A.; Hennuy, B.; Donneau, A.F.; Bourguignon, J.P.; Parent, A.S. Delayed Neuroendocrine Sexual Maturation in Female Rats After a Very Low Dose of Bisphenol A Through Altered GABAergic Neurotransmission and Opposing Effects of a High Dose. Endocrinology 2016, 157, 1740–1750. [Google Scholar] [CrossRef]
- Kasper-Sonnenberg, M.; Wittsiepe, J.; Wald, K.; Koch, H.M.; Wilhelm, M. Pre-pubertal exposure with phthalates and bisphenol A and pubertal development. PLoS ONE 2017, 12, e0187922. [Google Scholar] [CrossRef]
- Rasier, G.; Parent, A.S.; Gerard, A.; Lebrethon, M.C.; Bourguignon, J.P. Early maturation of gonadotropin-releasing hormone secretion and sexual precocity after exposure of infant female rats to estradiol or dichlorodiphenyltrichloroethane. Biol. Reprod. 2007, 77, 734–742. [Google Scholar] [CrossRef]
- Parent, A.S.; Franssen, D.; Fudvoye, J.; Gerard, A.; Bourguignon, J.P. Developmental variations in environmental influences including endocrine disruptors on pubertal timing and neuroendocrine control: Revision of human observations and mechanistic insight from rodents. Front. Neuroendocr. 2015, 38, 12–36. [Google Scholar] [CrossRef]
- Greenspan, L.C.; Lee, M.M. Endocrine disrupters and pubertal timing. Curr Opin Endocrinol Diabetes Obes 2018, 25, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Binder, A.M.; Corvalan, C.; Pereira, A.; Calafat, A.M.; Ye, X.; Shepherd, J.; Michels, K.B. Prepubertal and Pubertal Endocrine-Disrupting Chemical Exposure and Breast Density among Chilean Adolescents. Cancer Epidemiol. Biomark. Prev. 2018, 27, 1491–1499. [Google Scholar] [CrossRef] [PubMed]
- Wolff, M.S.; Teitelbaum, S.L.; McGovern, K.; Pinney, S.M.; Windham, G.C.; Galvez, M.; Pajak, A.; Rybak, M.; Calafat, A.M.; Kushi, L.H.; et al. Environmental phenols and pubertal development in girls. Environ. Int. 2015, 84, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Acevedo, N.; Davis, B.; Schaeberle, C.M.; Sonnenschein, C.; Soto, A.M. Perinatally administered bisphenol a as a potential mammary gland carcinogen in rats. Environ. Health Perspect. 2013, 121, 1040–1046. [Google Scholar] [CrossRef] [PubMed]
- Mandrup, K.; Boberg, J.; Isling, L.K.; Christiansen, S.; Hass, U. Low-dose effects of bisphenol A on mammary gland development in rats. Andrology 2016, 4, 673–683. [Google Scholar] [CrossRef] [PubMed]
- Marti, N.; Galvan, J.A.; Pandey, A.V.; Trippel, M.; Tapia, C.; Muller, M.; Perren, A.; Fluck, C.E. Genes and proteins of the alternative steroid backdoor pathway for dihydrotestosterone synthesis are expressed in the human ovary and seem enhanced in the polycystic ovary syndrome. Mol. Cell Endocrinol. 2017, 441, 116–123. [Google Scholar] [CrossRef]
- Moenter, S.M.; Silveira, M.A.; Wang, L.; Adams, C. Central aspects of systemic oestradiol negative- and positive-feedback on the reproductive neuroendocrine system. J. Neuroendocr. 2020, 32, e12724. [Google Scholar] [CrossRef]
- Gal, M.; Orly, J. Ketoconazole inhibits ovulation as a result of arrest of follicular steroidogenesis in the rat ovary. Clin. Med. Insights Reprod. Health 2014, 8, 37–44. [Google Scholar] [CrossRef]
- Parsanezhad, M.E.; Alborzi, S.; Pakniat, M.; Schmidt, E.H. A double-blind, randomized, placebo-controlled study to assess the efficacy of ketoconazole for reducing the risk of ovarian hyperstimulation syndrome. Fertil. Steril. 2003, 80, 1151–1155. [Google Scholar] [CrossRef][Green Version]
- Bjorvang, R.D.; Damdimopoulou, P. Persistent environmental endocrine-disrupting chemicals in ovarian follicular fluid and in vitro fertilization treatment outcome in women. Upsala J. Med. Sci. 2020, 1–10. [Google Scholar] [CrossRef]
- O’Shaughnessy, P.J.; Antignac, J.P.; Le Bizec, B.; Morvan, M.L.; Svechnikov, K.; Soder, O.; Savchuk, I.; Monteiro, A.; Soffientini, U.; Johnston, Z.C.; et al. Alternative (backdoor) androgen production and masculinization in the human fetus. PLoS Biol. 2019, 17, e3000002. [Google Scholar] [CrossRef] [PubMed]
- Craig, Z.R.; Wang, W.; Flaws, J.A. Endocrine-disrupting chemicals in ovarian function: Effects on steroidogenesis, metabolism and nuclear receptor signaling. Reproduction 2011, 142, 633–646. [Google Scholar] [CrossRef] [PubMed]
- Petro, E.M.; Leroy, J.L.; Covaci, A.; Fransen, E.; De Neubourg, D.; Dirtu, A.C.; De Pauw, I.; Bols, P.E. Endocrine-disrupting chemicals in human follicular fluid impair in vitro oocyte developmental competence. Hum. Reprod 2012, 27, 1025–1033. [Google Scholar] [CrossRef]
- Petro, E.M.L.; D’Hollander, W.; Covaci, A.; Bervoets, L.; Fransen, E.; De Neubourg, D.; De Pauw, I.; Leroy, J.; Jorssen, E.P.A.; Bols, P.E.J. Perfluoroalkyl acid contamination of follicular fluid and its consequence for in vitro oocyte developmental competence. Sci. Total Environ. 2014, 496, 282–288. [Google Scholar] [CrossRef] [PubMed]
- Darde, T.A.; Gaudriault, P.; Beranger, R.; Lancien, C.; Caillarec-Joly, A.; Sallou, O.; Bonvallot, N.; Chevrier, C.; Mazaud-Guittot, S.; Jegou, B.; et al. TOXsIgN: A cross-species repository for toxicogenomic signatures. Bioinformatics 2018, 34, 2116–2122. [Google Scholar] [CrossRef]
- EFSA/ECHA. Guidance for the identification of endocrine disruptors in the context of Regulations (EU) No 528/2012 and (EC) No 1107/2009. EFSA J. 2018, 16, e05311. [Google Scholar]
- Montévil, A.R.; Acevedo, N.; Schaeberle, C.; Bharadwai, M.; Fenton, S.E.; Soto, A.M. A combined morphometric and statistical approach to assess non-monotonicity in the developing mammary gland of rats in the CLARITY-BPA study. Environ. Health Perspect. 2020. accepted for publication. [Google Scholar]
- Drummond, A.E.; Fuller, P.J. The importance of ERbeta signalling in the ovary. J. Endocrinol. 2010, 205, 15–23. [Google Scholar] [CrossRef]
- Heublein, S.; Lenhard, M.; Vrekoussis, T.; Schoepfer, J.; Kuhn, C.; Friese, K.; Makrigiannakis, A.; Mayr, D.; Jeschke, U. The G-protein-coupled estrogen receptor (GPER) is expressed in normal human ovaries and is upregulated in ovarian endometriosis and pelvic inflammatory disease involving the ovary. Reprod. Sci. 2012, 19, 1197–1204. [Google Scholar] [CrossRef]
- Tang, Z.R.; Zhang, R.; Lian, Z.X.; Deng, S.L.; Yu, K. Estrogen-Receptor Expression and Function in Female Reproductive Disease. Cells 2019, 8, 1123. [Google Scholar] [CrossRef]
- Santos, R.R.; Schoevers, E.J.; Roelen, B.A. Usefulness of bovine and porcine IVM/IVF models for reproductive toxicology. Reprod. Biol. Endocrinol. 2014, 12, 117. [Google Scholar] [CrossRef] [PubMed]
- Demeneix, B.; Slama, R. Endocrine Disruptors: From Scientific Evidence to Human Health Protection. 2019. Available online: http://www.europarl.europa.eu/supporting-analyses (accessed on 30 April 2020).


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
van Duursen, M.B.M.; Boberg, J.; Christiansen, S.; Connolly, L.; Damdimopoulou, P.; Filis, P.; Fowler, P.A.; Gadella, B.M.; Holte, J.; Jääger, K.; et al. Safeguarding Female Reproductive Health Against Endocrine Disrupting Chemicals—The FREIA Project. Int. J. Mol. Sci. 2020, 21, 3215. https://doi.org/10.3390/ijms21093215
van Duursen MBM, Boberg J, Christiansen S, Connolly L, Damdimopoulou P, Filis P, Fowler PA, Gadella BM, Holte J, Jääger K, et al. Safeguarding Female Reproductive Health Against Endocrine Disrupting Chemicals—The FREIA Project. International Journal of Molecular Sciences. 2020; 21(9):3215. https://doi.org/10.3390/ijms21093215
Chicago/Turabian Stylevan Duursen, Majorie B. M., Julie Boberg, Sofie Christiansen, Lisa Connolly, Pauliina Damdimopoulou, Panagiotis Filis, Paul A. Fowler, Bart M. Gadella, Jan Holte, Kersti Jääger, and et al. 2020. "Safeguarding Female Reproductive Health Against Endocrine Disrupting Chemicals—The FREIA Project" International Journal of Molecular Sciences 21, no. 9: 3215. https://doi.org/10.3390/ijms21093215
APA Stylevan Duursen, M. B. M., Boberg, J., Christiansen, S., Connolly, L., Damdimopoulou, P., Filis, P., Fowler, P. A., Gadella, B. M., Holte, J., Jääger, K., Johansson, H. K. L., Li, T., Mazaud-Guittot, S., Parent, A.-S., Salumets, A., Soto, A. M., Svingen, T., Velthut-Meikas, A., Bay Wedebye, E., ... van den Berg, M. (2020). Safeguarding Female Reproductive Health Against Endocrine Disrupting Chemicals—The FREIA Project. International Journal of Molecular Sciences, 21(9), 3215. https://doi.org/10.3390/ijms21093215