The Significance of MicroRNAs Expression in Regulation of Extracellular Matrix and Other Drug Resistant Genes in Drug Resistant Ovarian Cancer Cell Lines

 , ,
, ,  ,
,  and
and
Abstract
1. Introduction
2. Results
2.1. Gene Chip Quality Assessment
2.2. Gene Expression Evaluation and Gene Expression Lists
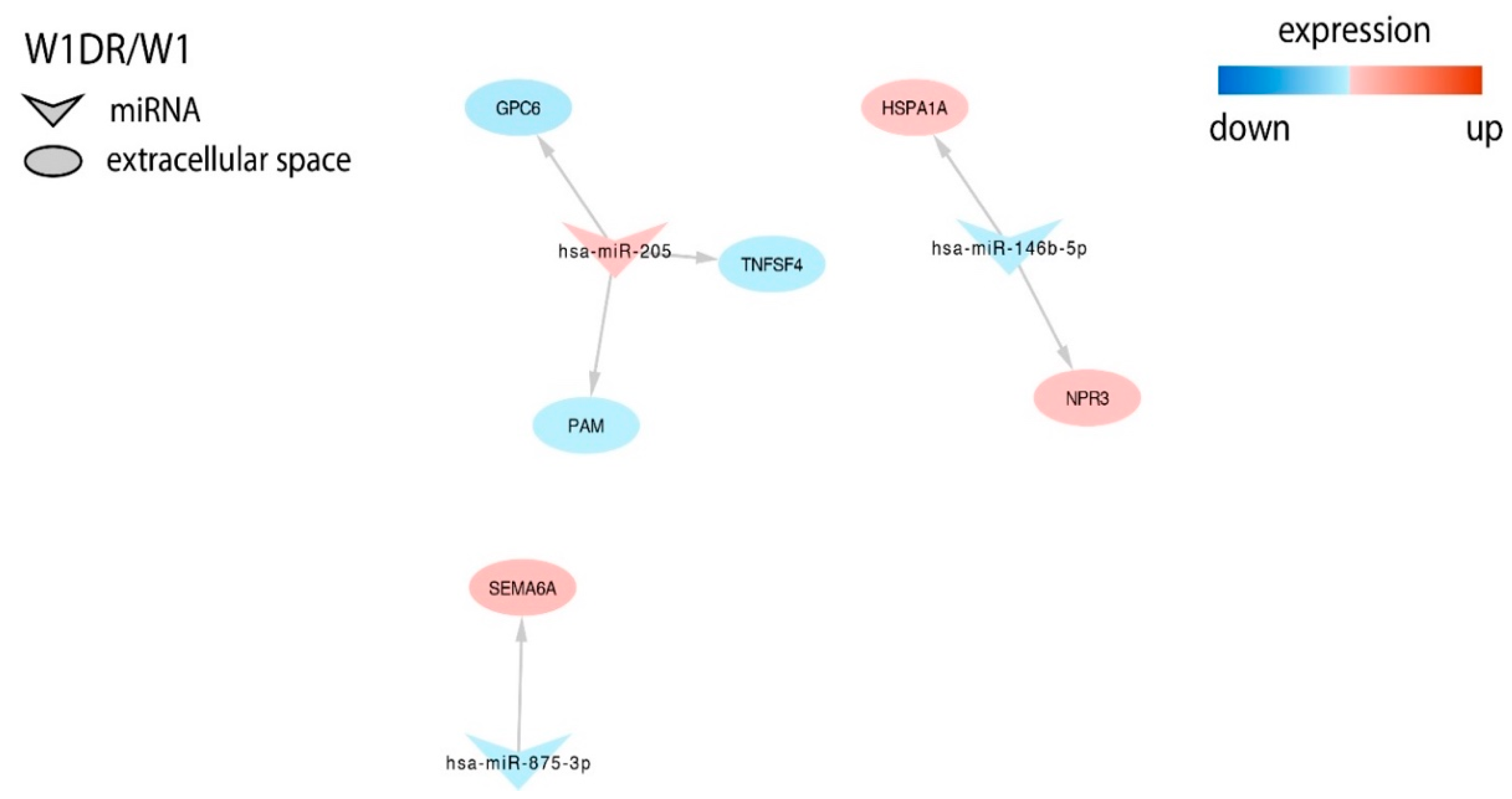
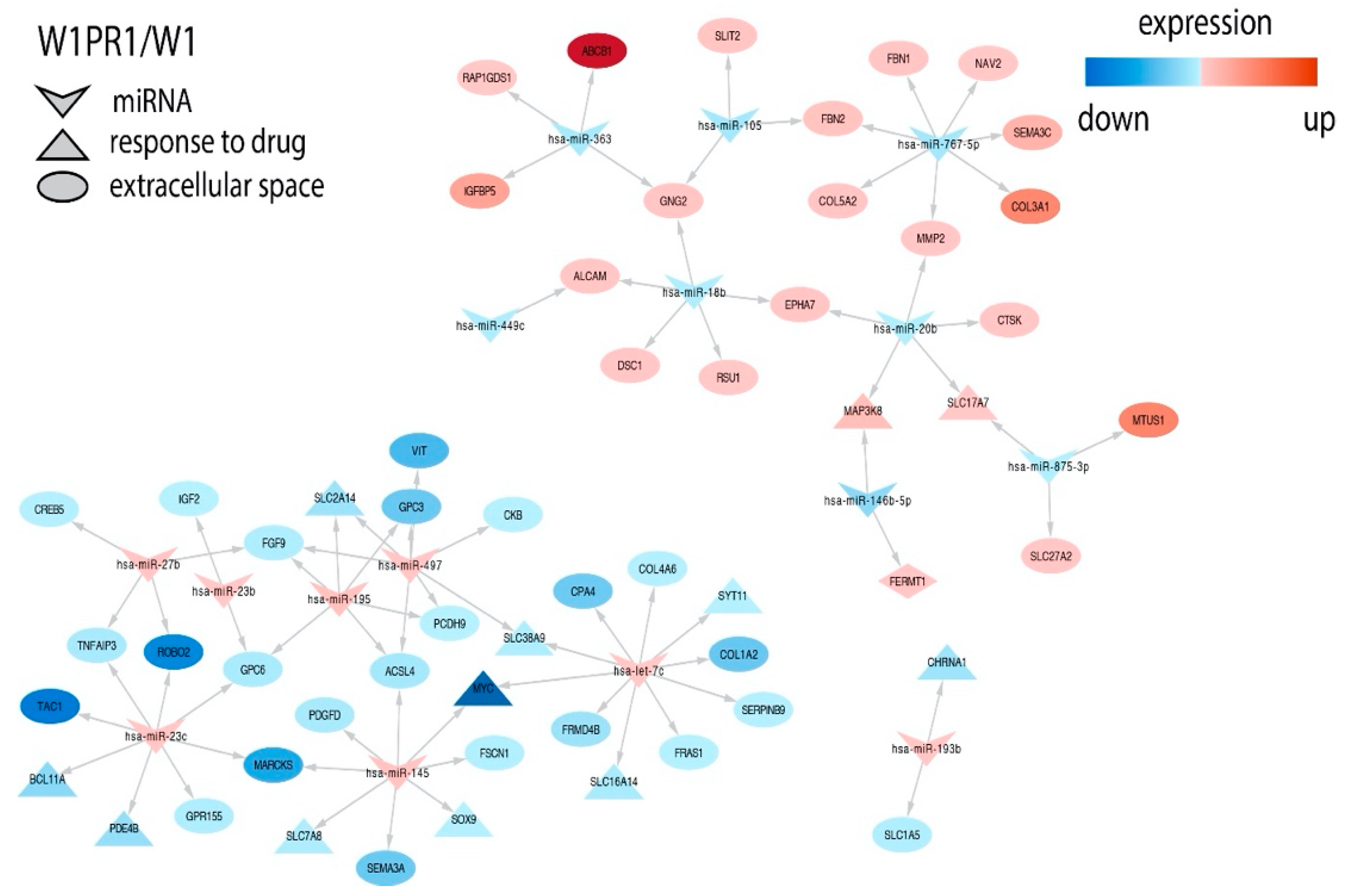
2.3. miRNAs Expression in Drug Resistant Cell Lines
2.4. Analysis of Target Genes Expression
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Cell Lines and Cell Culture
4.3. miRNA Isolation
4.4. Microarray Preparation, Hybridization, and Scanning
4.5. Microarray Analysis and miRNA Gene Screening
4.6. miRNA-Target Gene Prediction
4.7. Analysis of The Cancer Genome Atlas (TCGA) Dataset
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| CIS | Cisplatin |
| PAC | Paclitaxel |
| DOX | Doxorubicin |
| TOP | Topotecan |
| FIGO | fr. Fédération internationale de gynécologie et d’obstétrique |
| ABCB1 (MDR1) | ATP-binding cassette subfamily B member 1 (multidrug resistance protein 1) |
| W1PR1, W1PR2, W1CR, W1TR, W1DR | Resistant Variants of W1 Sensitive Ovarian Cancer Cell Line |
| ECM | Extracellular Matrix |
| FDR | False Discovery Rate |
| GEO | Gene Expression Omnibus |
| GO | Gene Ontology |
| COL3A1 | Collagen Type III Alpha 1 Chain |
| COL5A2 | Collagen Type V Alpha 2 Chain |
| MYC | Myelocytomatosis Viral Oncogene Homolog |
| EPHA7 | Ephrin Type-A Receptor 7 |
| MAP3K8 | Mitogen-Activated Protein Kinase Kinase Kinase 8 |
| LOX | Lysyl Oxidase |
| EPHA7 | Ephrin type-A receptor 7 |
| DCN | Decorin |
| MAP3K8 | Mitogen-Activated Protein Kinase 8 |
| SEM3A | Semaphorin 3A |
| PCDH9 | Protocadherin 9 |
| SPP1/OPN | Secreted Phosphoprotein 1/Osteopontin |
| TGFBI | Transforming Growth Factor Beta Induced |
References
- Roett, M.A.; Evans, P. Ovarian cancer: An overview. Am. Fam. Physician 2009, 80, 609–616. [Google Scholar] [PubMed]
- Hennessy, B.T.; Coleman, R.L.; Markman, M. Ovarian cancer. Lancet (Lond. Engl.) 2009, 374, 1371–1382. [Google Scholar] [CrossRef]
- Webb, P.M.; Jordan, S.J. Epidemiology of epithelial ovarian cancer. Best Pract. Res. Clin. Obstet. Gynaecol. 2017, 41, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Giulia, F.; Luffer, S.; Kumar, R.; Wu, B.; Tavallai, M.; Bekele, R.T.; Birrer, M.J. How can molecular abnormalities influence our clinical approach. Ann. Oncol. 2017, 28, viii16–viii24. [Google Scholar] [CrossRef]
- Gottesman, M.M. Mechanisms of cancer drug resistance. Ann. Rev. Med. 2002, 53, 615–627. [Google Scholar] [CrossRef] [PubMed]
- Greenlee, R.T.; Murray, T.; Bolden, S.; Wingo, P.A. Cancer statistics, 2000. CA Cancer J. Clin. 2000, 50, 7–33. [Google Scholar] [CrossRef]
- Gottesman, M.M.; Fojo, T.; Bates, S.E. Multidrug resistance in cancer: Role of ATP-dependent transporters. Nat. Rev. Cancer 2002, 2, 48–58. [Google Scholar] [CrossRef]
- Leonard, G.D.; Fojo, T.; Bates, S.E. The role of ABC transporters in clinical practice. Oncologist 2003, 8, 411–424. [Google Scholar] [CrossRef]
- Mansoori, B.; Mohammadi, A.; Davudian, S.; Shirjang, S.; Baradaran, B. The Different Mechanisms of Cancer Drug Resistance: A Brief Review. Adv. Pharm. Bull. 2017, 7, 339–348. [Google Scholar] [CrossRef]
- Correia, A.L.; Bissell, M.J. The tumor microenvironment is a dominant force in multidrug resistance. Drug Resist. Updates Rev. Comment. Antimicrob. Anticancer Chemother. 2012, 15, 39–49. [Google Scholar] [CrossRef]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Mihanfar, A.; Fattahi, A.; Nejabati, H.R. MicroRNA-mediated drug resistance in ovarian cancer. J. Cell. Physiol. 2019, 234, 3180–3191. [Google Scholar] [CrossRef]
- Baranwal, S.; Alahari, S.K. miRNA control of tumor cell invasion and metastasis. Int. J. Cancer 2010, 126, 1283–1290. [Google Scholar] [CrossRef]
- Lee, Y.; Kim, M.; Han, J.; Yeom, K.H.; Lee, S.; Baek, S.H.; Kim, V.N. MicroRNA genes are transcribed by RNA polymerase II. EMBO J. 2004, 23, 4051–4060. [Google Scholar] [CrossRef] [PubMed]
- Kamanu, T.K.; Radovanovic, A.; Archer, J.A.; Bajic, V.B. Exploration of miRNA families for hypotheses generation. Sci. Rep. 2013, 3, 2940. [Google Scholar] [CrossRef] [PubMed]
- Zou, Q.; Mao, Y.; Hu, L.; Wu, Y.; Ji, Z. miRClassify: An advanced web server for miRNA family classification and annotation. Comput. Biol. Med. 2014, 45, 157–160. [Google Scholar] [CrossRef]
- Zhang, W.; Dahlberg, J.E.; Tam, W. MicroRNAs in tumorigenesis: A primer. Am. J. Pathol. 2007, 171, 728–738. [Google Scholar] [CrossRef]
- Lu, J.; Getz, G.; Miska, E.A.; Alvarez-Saavedra, E.; Lamb, J.; Peck, D.; Sweet-Cordero, A.; Ebert, B.L.; Mak, R.H.; Ferrando, A.A.; et al. MicroRNA expression profiles classify human cancers. Nature 2005, 435, 834–838. [Google Scholar] [CrossRef] [PubMed]
- Shenouda, S.K.; Alahari, S.K. MicroRNA function in cancer: Oncogene or a tumor suppressor? Cancer Metastasis Rev. 2009, 28, 369–378. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Zhang, L.; Huang, H.; Huang, Y.; Huang, L.; Wang, J.; Huang, S.; He, L.; Zhou, Y.; Jia, W.; et al. MiR-26b/KPNA2 axis inhibits epithelial ovarian carcinoma proliferation and metastasis through downregulating OCT4. Oncotarget 2015, 6, 23793–23806. [Google Scholar] [CrossRef] [PubMed]
- Teng, Y.; Zhang, Y.; Qu, K.; Yang, X.; Fu, J.; Chen, W.; Li, X. MicroRNA-29B (mir-29b) regulates the Warburg effect in ovarian cancer by targeting AKT2 and AKT3. Oncotarget 2015, 6, 40799–40814. [Google Scholar] [CrossRef] [PubMed]
- Sestito, R.; Cianfrocca, R.; Rosano, L.; Tocci, P.; Semprucci, E.; Di Castro, V.; Caprara, V.; Ferrandina, G.; Sacconi, A.; Blandino, G.; et al. miR-30a inhibits endothelin A receptor and chemoresistance in ovarian carcinoma. Oncotarget 2016, 7, 4009–4023. [Google Scholar] [CrossRef]
- Chen, X.; Chen, S.; Xiu, Y.L.; Sun, K.X.; Zong, Z.H.; Zhao, Y. RhoC is a major target of microRNA-93-5P in epithelial ovarian carcinoma tumorigenesis and progression. Mol. Cancer 2015, 14, 31. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Chen, S.; Chen, X.; Xiu, Y.L.; Sun, K.X.; Zhao, Y. Inhibition of Ovarian Epithelial Carcinoma Tumorigenesis and Progression by microRNA 106b Mediated through the RhoC Pathway. PLoS ONE 2015, 10, e0125714. [Google Scholar] [CrossRef]
- Ying, X.; Wei, K.; Lin, Z.; Cui, Y.; Ding, J.; Chen, Y.; Xu, B. MicroRNA-125b Suppresses Ovarian Cancer Progression via Suppression of the Epithelial-Mesenchymal Transition Pathway by Targeting the SET Protein. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2016, 39, 501–510. [Google Scholar] [CrossRef]
- Chen, H.; Zhang, L.; Zhang, L.; Du, J.; Wang, H.; Wang, B. MicroRNA-183 correlates cancer prognosis, regulates cancer proliferation and bufalin sensitivity in epithelial ovarian caner. Am. J. Transl. Res. 2016, 8, 1748–1755. [Google Scholar]
- Yang, L.; Wei, Q.M.; Zhang, X.W.; Sheng, Q.; Yan, X.T. MiR-376a promotion of proliferation and metastases in ovarian cancer: Potential role as a biomarker. Life Sci. 2017, 173, 62–67. [Google Scholar] [CrossRef]
- Liu, J.; Dou, Y.; Sheng, M. Inhibition of microRNA-383 has tumor suppressive effect in human epithelial ovarian cancer through the action on caspase-2 gene. Biomed. Pharmacother. 2016, 83, 1286–1294. [Google Scholar] [CrossRef]
- Chaluvally-Raghavan, P.; Jeong, K.J.; Pradeep, S.; Silva, A.M.; Yu, S.; Liu, W.; Moss, T.; Rodriguez-Aguayo, C.; Zhang, D.; Ram, P.; et al. Direct Upregulation of STAT3 by MicroRNA-551b-3p Deregulates Growth and Metastasis of Ovarian Cancer. Cell Rep. 2016, 15, 1493–1504. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, J.; Zang, D.; Wu, S.; Liu, A.; Zhu, J.; Wu, G.; Li, J.; Jiang, L. Upregulation of miR-572 transcriptionally suppresses SOCS1 and p21 and contributes to human ovarian cancer progression. Oncotarget 2015, 6, 15180–15193. [Google Scholar] [CrossRef] [PubMed]
- Su, L.; Liu, M. Correlation analysis on the expression levels of microRNA-23a and microRNA-23b and the incidence and prognosis of ovarian cancer. Oncol. Lett. 2018, 16, 262–266. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.W.; Kim, E.Y.; Jeon, D.; Liu, J.L.; Kim, H.S.; Choi, J.W.; Ahn, W.S. Differential microRNA expression signatures and cell type-specific association with Taxol resistance in ovarian cancer cells. Drug Des. Dev. Ther. 2014, 8, 293–314. [Google Scholar] [CrossRef]
- Mitamura, T.; Watari, H.; Wang, L.; Kanno, H.; Hassan, M.K.; Miyazaki, M.; Katoh, Y.; Kimura, T.; Tanino, M.; Nishihara, H.; et al. Downregulation of miRNA-31 induces taxane resistance in ovarian cancer cells through increase of receptor tyrosine kinase MET. Oncogenesis 2013, 2, e40. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Bao, W.; Liu, Y.; Wang, S.; Xu, S.; Li, X.; Li, Y.; Wu, S. miR-98-5p contributes to cisplatin resistance in epithelial ovarian cancer by suppressing miR-152 biogenesis via targeting Dicer1. Cell Death Dis. 2018, 9, 447. [Google Scholar] [CrossRef]
- Wang, H.; Ren, S.; Xu, Y.; Miao, W.; Huang, X.; Qu, Z.; Li, J.; Liu, X.; Kong, P. MicroRNA-195 reverses the resistance to temozolomide through targeting cyclin E1 in glioma cells. Anti-Cancer Drugs 2019, 30, 81–88. [Google Scholar] [CrossRef]
- Chen, L.Z.; Ding, Z.; Zhang, Y.; He, S.T.; Wang, X.H. MiR-203 over-expression promotes prostate cancer cell apoptosis and reduces ADM resistance. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 3734–3741. [Google Scholar] [CrossRef]
- Januchowski, R.; Sterzynska, K.; Zawierucha, P.; Rucinski, M.; Swierczewska, M.; Partyka, M.; Bednarek-Rajewska, K.; Brazert, M.; Nowicki, M.; Zabel, M.; et al. Microarray-based detection and expression analysis of new genes associated with drug resistance in ovarian cancer cell lines. Oncotarget 2017, 8, 49944–49958. [Google Scholar] [CrossRef]
- Januchowski, R.; Zawierucha, P.; Andrzejewska, M.; Rucinski, M.; Zabel, M. Microarray-based detection and expression analysis of ABC and SLC transporters in drug-resistant ovarian cancer cell lines. Biomed. Pharmacother. 2013, 67, 240–245. [Google Scholar] [CrossRef]
- Januchowski, R.; Zawierucha, P.; Rucinski, M.; Zabel, M. Microarray-based detection and expression analysis of extracellular matrix proteins in drug resistant ovarian cancer cell lines. Oncol. Rep. 2014, 32, 1981–1990. [Google Scholar] [CrossRef]
- Ma, J.; Dong, C.; Ji, C. MicroRNA and drug resistance. Cancer Gene Ther. 2010, 17, 523–531. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.K.; Watari, H.; Mitamura, T.; Mohamed, Z.; El-Khamisy, S.F.; Ohba, Y.; Sakuragi, N. P18/Stathmin1 is regulated by miR-31 in ovarian cancer in response to taxane. Oncoscience 2015, 2, 294–308. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Chen, W.; Zhang, H.; Zhang, Y.; Ke, F.; Wu, X.; Zhang, Y.; Weng, M.; Liu, Y.; Gong, W. MiR-31 regulates the cisplatin resistance by targeting Src in gallbladder cancer. Oncotarget 2016, 7, 83060–83070. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Januchowski, R.; Wojtowicz, K.; Sterzyska, K.; Sosiska, P.; Andrzejewska, M.; Zawierucha, P.; Nowicki, M.; Zabel, M. Inhibition of ALDH1A1 activity decreases expression of drug transporters and reduces chemotherapy resistance in ovarian cancer cell lines. Int. J. Biochem. Cell Biol. 2016, 78, 248–259. [Google Scholar] [CrossRef]
- Sterzynska, K.; Klejewski, A.; Wojtowicz, K.; Swierczewska, M.; Nowacka, M.; Kazmierczak, D.; Andrzejewska, M.; Rusek, D.; Brazert, M.; Brazert, J.; et al. Mutual Expression of ALDH1A1, LOX, and Collagens in Ovarian Cancer Cell Lines as Combined CSCs- and ECM-Related Models of Drug Resistance Development. Int. J. Mol. Sci. 2018, 20, 54. [Google Scholar] [CrossRef]
- Lv, C.; Li, F.; Li, X.; Tian, Y.; Zhang, Y.; Sheng, X.; Song, Y.; Meng, Q.; Yuan, S.; Luan, L.; et al. MiR-31 promotes mammary stem cell expansion and breast tumorigenesis by suppressing Wnt signaling antagonists. Nat. Commun. 2017, 8, 1036. [Google Scholar] [CrossRef]
- Yang, M.; Shan, X.; Zhou, X.; Qiu, T.; Zhu, W.; Ding, Y.; Shu, Y.; Liu, P. miR-1271 regulates cisplatin resistance of human gastric cancer cell lines by targeting IGF1R, IRS1, mTOR, and BCL2. Anti-Cancer Agents Med. Chem. 2014, 14, 884–891. [Google Scholar] [CrossRef]
- Yang, L.; Wang, Y.; Li, Y.J.; Zeng, C.C. Chemo-resistance of A172 glioblastoma cells is controlled by miR-1271-regulated Bcl-2. Biomed. Pharmacother. 2018, 108, 734–740. [Google Scholar] [CrossRef]
- Lu, G.; Fu, D.; Jia, C.; Chai, L.; Han, Y.; Liu, J.; Wu, T.; Xie, R.; Chang, Z.; Yang, H.; et al. Reduced miR-105-1 levels are associated with poor survival of patients with non-small cell lung cancer. Oncol. Lett. 2017, 14, 7842–7848. [Google Scholar] [CrossRef]
- Li, H.Y.; Liang, J.L.; Kuo, Y.L.; Lee, H.H.; Calkins, M.J.; Chang, H.T.; Lin, F.C.; Chen, Y.C.; Hsu, T.I.; Hsiao, M.; et al. miR-105/93-3p promotes chemoresistance and circulating miR-105/93-3p acts as a diagnostic biomarker for triple negative breast cancer. Breast Cancer Res. BCR 2017, 19, 133. [Google Scholar] [CrossRef]
- Xiang, Y.; Ma, N.; Wang, D.; Zhang, Y.; Zhou, J.; Wu, G.; Zhao, R.; Huang, H.; Wang, X.; Qiao, Y.; et al. MiR-152 and miR-185 co-contribute to ovarian cancer cells cisplatin sensitivity by targeting DNMT1 directly: A novel epigenetic therapy independent of decitabine. Oncogene 2014, 33, 378–386. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.; Wei, R.; Zhang, P.; Kong, B. Overexpression of microRNA-195-5p reduces cisplatin resistance and angiogenesis in ovarian cancer by inhibiting the PSAT1-dependent GSK3beta/beta-catenin signaling pathway. J. Transl. Med. 2019, 17, 190. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.Z.; Xie, J.; Jin, B.; Chen, X.W.; Sun, Z.F.; Wang, B.X.; Dong, P. Gene and microRNA expression reveals sensitivity to paclitaxel in laryngeal cancer cell line. Int. J. Clin. Exp. Pathol. 2013, 6, 1351–1361. [Google Scholar] [PubMed]
- Dai, Y.; Xie, C.H.; Neis, J.P.; Fan, C.Y.; Vural, E.; Spring, P.M. MicroRNA expression profiles of head and neck squamous cell carcinoma with docetaxel-induced multidrug resistance. Head Neck 2011, 33, 786–791. [Google Scholar] [CrossRef]
- Sun, W.; Ma, Y.; Chen, P.; Wang, D. MicroRNA-10a silencing reverses cisplatin resistance in the A549/cisplatin human lung cancer cell line via the transforming growth factor-beta/Smad2/STAT3/STAT5 pathway. Mol. Med. Rep. 2015, 11, 3854–3859. [Google Scholar] [CrossRef]
- Yan, M.; Yang, X.; Shen, R.; Wu, C.; Wang, H.; Ye, Q.; Yang, P.; Zhang, L.; Chen, M.; Wan, B.; et al. miR-146b promotes cell proliferation and increases chemosensitivity, but attenuates cell migration and invasion via FBXL10 in ovarian cancer. Cell Death Dis. 2018, 9, 1123. [Google Scholar] [CrossRef]
- Cheng, R.; Lu, C.; Zhang, G.; Zhang, G.; Zhao, G. Overexpression of miR-203 increases the sensitivity of NSCLC A549/H460 cell lines to cisplatin by targeting Dickkopf-1. Oncol. Rep. 2017, 37, 2129–2136. [Google Scholar] [CrossRef]
- Liu, Y.; Gao, S.; Chen, X.; Liu, M.; Mao, C.; Fang, X. Overexpression of miR-203 sensitizes paclitaxel (Taxol)-resistant colorectal cancer cells through targeting the salt-inducible kinase 2 (SIK2). Tumour Biol. J. Int. Soc. Oncodev. Biol. Med. 2016, 37, 12231–12239. [Google Scholar] [CrossRef]
- Mohamed, Z.; Hassan, M.K.; Okasha, S.; Mitamura, T.; Keshk, S.; Konno, Y.; Kato, T.; El-Khamisy, S.F.; Ohba, Y.; Watari, H. miR-363 confers taxane resistance in ovarian cancer by targeting the Hippo pathway member, LATS2. Oncotarget 2018, 9, 30053–30065. [Google Scholar] [CrossRef]
- Ou, Y.; Zhai, D.; Wu, N.; Li, X. Downregulation of miR-363 increases drug resistance in cisplatin-treated HepG2 by dysregulating Mcl-1. Gene 2015, 572, 116–122. [Google Scholar] [CrossRef]
- Cao, L.; Wan, Q.; Li, F.; Tang, C.E. MiR-363 inhibits cisplatin chemoresistance of epithelial ovarian cancer by regulating snail-induced epithelial-mesenchymal transition. BMB Rep. 2018, 51, 456–461. [Google Scholar] [CrossRef] [PubMed]
- Yu, P.N.; Yan, M.D.; Lai, H.C.; Huang, R.L.; Chou, Y.C.; Lin, W.C.; Yeh, L.T.; Lin, Y.W. Downregulation of miR-29 contributes to cisplatin resistance of ovarian cancer cells. Int. J. Cancer 2014, 134, 542–551. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Li, N.; Yan, Z.; Li, C.; Zhao, Z. MiR-29a-Mediated CD133 Expression Contributes to Cisplatin Resistance in CD133(+) Glioblastoma Stem Cells. J. Mol. Neurosci. MN 2018, 66, 369–377. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Li, Z.; Zhu, X.; Xu, R.; Xu, Y. miR-29 Family Inhibits Resistance to Methotrexate and Promotes Cell Apoptosis by Targeting COL3A1 and MCL1 in Osteosarcoma. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2018, 24, 8812–8821. [Google Scholar] [CrossRef] [PubMed]
- Sorrentino, A.; Liu, C.G.; Addario, A.; Peschle, C.; Scambia, G.; Ferlini, C. Role of microRNAs in drug-resistant ovarian cancer cells. Gynecol. Oncol. 2008, 111, 478–486. [Google Scholar] [CrossRef]
- Guo, Y.; Zhang, H.; Xie, D.; Hu, X.; Song, R.; Zhu, L. Non-coding RNA NEAT1/miR-214-3p contribute to doxorubicin resistance of urothelial bladder cancer preliminary through the Wnt/beta-catenin pathway. Cancer Manag. Res. 2018, 10, 4371–4380. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, W.; Ni, P.; Li, A.; Zhou, J.; Xu, S. MiR-99a and MiR-491 Regulate Cisplatin Resistance in Human Gastric Cancer Cells by Targeting CAPNS1. Int. J. Biol. Sci. 2016, 12, 1437–1447. [Google Scholar] [CrossRef]
- Akbari Moqadam, F.; Lange-Turenhout, E.A.; Aries, I.M.; Pieters, R.; den Boer, M.L. MiR-125b, miR-100 and miR-99a co-regulate vincristine resistance in childhood acute lymphoblastic leukemia. Leuk. Res. 2013, 37, 1315–1321. [Google Scholar] [CrossRef]
- Zhu, X.; Li, Y.; Xie, C.; Yin, X.; Liu, Y.; Cao, Y.; Fang, Y.; Lin, X.; Xu, Y.; Xu, W.; et al. miR-145 sensitizes ovarian cancer cells to paclitaxel by targeting Sp1 and Cdk6. Int. J. Cancer 2014, 135, 1286–1296. [Google Scholar] [CrossRef]
- Khordadmehr, M.; Shahbazi, R.; Sadreddini, S.; Baradaran, B. miR-193: A new weapon against cancer. J. Cell. Physiol. 2019, 234, 16861–16872. [Google Scholar] [CrossRef]
- Janikova, M.; Zizkova, V.; Skarda, J.; Kharaishvili, G.; Radova, L.; Kolar, Z. Prognostic significance of miR-23b in combination with P-gp, MRP and LRP/MVP expression in non-small cell lung cancer. Neoplasma 2016, 63, 576–587. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Han, Y.F.; Ye, B.; Zhang, Y.L.; Dong, J.D.; Zhu, S.J.; Chen, J. miR-27b-3p is Involved in Doxorubicin Resistance of Human Anaplastic Thyroid Cancer Cells via Targeting Peroxisome Proliferator-Activated Receptor Gamma. Basic Clin. Pharmacol. Toxicol. 2018, 123, 670–677. [Google Scholar] [CrossRef] [PubMed]
- Husted, S.; Sokilde, R.; Rask, L.; Cirera, S.; Busk, P.K.; Eriksen, J.; Litman, T. MicroRNA expression profiles associated with development of drug resistance in Ehrlich ascites tumor cells. Mol. Pharm. 2011, 8, 2055–2062. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Jiang, X.; Chen, L.; Zhao, Y.; Huang, Z.; Zhou, H.; Shi, M. MiR-181a Promotes Apoptosis and Reduces Cisplatin Resistance by Inhibiting Osteopontin in Cervical Cancer Cells. Cancer Biother. Radiopharm. 2019, 34, 559–565. [Google Scholar] [CrossRef]
- Wang, X.; Chen, X.; Meng, Q.; Jing, H.; Lu, H.; Yang, Y.; Cai, L.; Zhao, Y. MiR-181b regulates cisplatin chemosensitivity and metastasis by targeting TGFbetaR1/Smad signaling pathway in NSCLC. Sci. Rep. 2015, 5, 17618. [Google Scholar] [CrossRef]
- Cheng, W.; Liu, T.; Wan, X.; Gao, Y.; Wang, H. MicroRNA-199a targets CD44 to suppress the tumorigenicity and multidrug resistance of ovarian cancer-initiating cells. FEBS J. 2012, 279, 2047–2059. [Google Scholar] [CrossRef]
- Liu, M.X.; Siu, M.K.; Liu, S.S.; Yam, J.W.; Ngan, H.Y.; Chan, D.W. Epigenetic silencing of microRNA-199b-5p is associated with acquired chemoresistance via activation of JAG1-Notch1 signaling in ovarian cancer. Oncotarget 2014, 5, 944–958. [Google Scholar] [CrossRef]
- Januchowski, R.; Zawierucha, P.; Rucinski, M.; Andrzejewska, M.; Wojtowicz, K.; Nowicki, M.; Zabel, M. Drug transporter expression profiling in chemoresistant variants of the A2780 ovarian cancer cell line. Biomed. Pharmacother. 2014, 68, 447–453. [Google Scholar] [CrossRef]
- Januchowski, R.; Zawierucha, P.; Rucinski, M.; Nowicki, M.; Zabel, M. Extracellular matrix proteins expression profiling in chemoresistant variants of the A2780 ovarian cancer cell line. BioMed Res. Int. 2014, 2014, 365867. [Google Scholar] [CrossRef]
- Prislei, S.; Mozzetti, S.; Filippetti, F.; De Donato, M.; Raspaglio, G.; Cicchillitti, L.; Scambia, G.; Ferlini, C. From plasma membrane to cytoskeleton: A novel function for semaphorin 6A. Mol. Cancer Ther. 2008, 7, 233–241. [Google Scholar] [CrossRef]
- Januchowski, R.; Wojtowicz, K.; Sujka-Kordowska, P.; Andrzejewska, M.; Zabel, M. MDR gene expression analysis of six drug-resistant ovarian cancer cell lines. BioMed Res. Int. 2013, 2013, 241763. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Chen, J.; Ding, C.M.; Jin, X.; Jia, Z.M.; Peng, J. LncRNA NR2F1-AS1 regulates hepatocellular carcinoma oxaliplatin resistance by targeting ABCC1 via miR-363. J. Cell. Mol. Med. 2018, 22, 3238–3245. [Google Scholar] [CrossRef] [PubMed]
- Januchowski, R.; Swierczewska, M.; Sterzynska, K.; Wojtowicz, K.; Nowicki, M.; Zabel, M. Increased Expression of Several Collagen Genes is Associated with Drug Resistance in Ovarian Cancer Cell Lines. J. Cancer 2016, 7, 1295–1310. [Google Scholar] [CrossRef] [PubMed]
- Swierczewska, M.; Sterzynska, K.; Wojtowicz, K.; Kazmierczak, D.; Izycki, D.; Nowicki, M.; Zabel, M.; Januchowski, R. PTPRK Expression Is Downregulated in Drug Resistant Ovarian Cancer Cell Lines, and Especially in ALDH1A1 Positive CSCs-Like Populations. Int. J. Mol. Sci. 2019, 20, E2053. [Google Scholar] [CrossRef]
- Zou, J.; Yin, F.; Wang, Q.; Zhang, W.; Li, L. Analysis of microarray-identified genes and microRNAs associated with drug resistance in ovarian cancer. Int. J. Clin. Exp. Pathol. 2015, 8, 6847–6858. [Google Scholar]
- Zhou, R. The Eph family receptors and ligands. Pharmacol. Ther. 1998, 77, 151–181. [Google Scholar] [CrossRef]
- Zhang, S.J.; Zhang, G.; Zhao, Y.F.; Wu, Y.; Li, J.; Chai, Y.X. [Expression of EphA7 protein in primary hepatocellular carcinoma and its clinical significance]. Zhonghua wai ke za zhi (Chine. J. Surg.) 2010, 48, 53–56. [Google Scholar]
- Giani, F.; Russo, G.; Pennisi, M.; Sciacca, L.; Frasca, F.; Pappalardo, F. Computational modeling reveals MAP3K8 as mediator of resistance to vemurafenib in thyroid cancer stem cells. Bioinformatics (Oxf. Engl.) 2019, 35, 2267–2275. [Google Scholar] [CrossRef]
- Izycka, N.; Sterzynska, K.; Januchowski, R.; Nowak-Markwitz, E. Semaphorin 3A (SEMA3A), protocadherin 9 (PCdh9), and S100 calcium binding protein A3 (S100A3) as potential biomarkers of carcinogenesis and chemoresistance of different neoplasms, including ovarian cancer-review of literature. Ginekol. Polska 2019, 90, 223–227. [Google Scholar] [CrossRef]
- Tang, C.; Gao, X.; Liu, H.; Jiang, T.; Zhai, X. Decreased expression of SEMA3A is associated with poor prognosis in gastric carcinoma. Int. J. Clin. Exp. Pathol. 2014, 7, 4782–4794. [Google Scholar]
- Jiang, H.; Qi, L.; Wang, F.; Sun, Z.; Huang, Z.; Xi, Q. Decreased semaphorin 3A expression is associated with a poor prognosis in patients with epithelial ovarian carcinoma. Int. J. Mol. Med. 2015, 35, 1374–1380. [Google Scholar] [CrossRef] [PubMed]
- Swierczewska, M.; Klejewski, A.; Brazert, M.; Kazmierczak, D.; Izycki, D.; Nowicki, M.; Zabel, M.; Januchowski, R. New and Old Genes Associated with Primary and Established Responses to Paclitaxel Treatment in Ovarian Cancer Cell Lines. Molecules (Basel Switz.) 2018, 23, 891. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Hong, F.; Bao, Q.; Xu, Q.; Duan, R.; Zhu, Z.; Zhang, W.; Ma, C. MicroRNA-145 suppresses osteogenic differentiation of human jaw bone marrow mesenchymal stem cells partially via targeting semaphorin 3A. Connect. Tissue Res. 2019, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Xia, J.; Wang, N.; Zong, H. miR-145 inhibits proliferation and invasion of esophageal squamous cell carcinoma in part by targeting c-Myc. Onkologie 2013, 36, 754–758. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, X.; Wu, L.; Pei, M.; Li, H.; Jiang, Y. miR-145 inhibits glutamine metabolism through c-myc/GLS1 pathways in ovarian cancer cells. Cell Biol. Int. 2019, 43, 921–930. [Google Scholar] [CrossRef]
- Nadiminty, N.; Tummala, R.; Lou, W.; Zhu, Y.; Zhang, J.; Chen, X.; eVere White, R.W.; Kung, H.J.; Evans, C.P.; Gao, A.C. MicroRNA let-7c suppresses androgen receptor expression and activity via regulation of Myc expression in prostate cancer cells. J. Biol. Chem. 2012, 287, 1527–1537. [Google Scholar] [CrossRef]
- Frank, M.; Kemler, R. Protocadherins. Curr. Opin. Cell Biol. 2002, 14, 557–562. [Google Scholar] [CrossRef]
- Kim, S.Y.; Yasuda, S.; Tanaka, H.; Yamagata, K.; Kim, H. Non-clustered protocadherin. Cell Adhes. Migr. 2011, 5, 97–105. [Google Scholar] [CrossRef]
- Shi, C.; Zhang, Z. Screening of potentially crucial genes and regulatory factors involved in epithelial ovarian cancer using microarray analysis. Oncol. Lett. 2017, 14, 725–732. [Google Scholar] [CrossRef]
- Chen, Y.; Xiang, H.; Zhang, Y.; Wang, J.; Yu, G. Loss of PCDH9 is associated with the differentiation of tumor cells and metastasis and predicts poor survival in gastric cancer. Clin. Exp. Metastasis 2015, 32, 417–428. [Google Scholar] [CrossRef]
- Sofeu Feugaing, D.D.; Gotte, M.; Viola, M. More than matrix: The multifaceted role of decorin in cancer. Eur. J. Cell Biol. 2013, 92, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kwan, P.; Ding, J.; Tredget, E.E. MicroRNA 181b regulates decorin production by dermal fibroblasts and may be a potential therapy for hypertrophic scar. PLoS ONE 2015, 10, e0123054. [Google Scholar] [CrossRef] [PubMed]
- Lucero, H.A.; Kagan, H.M. Lysyl oxidase: An oxidative enzyme and effector of cell function. Cell. Mol. Life Sci. CMLS 2006, 63, 2304–2316. [Google Scholar] [CrossRef]
- Erler, J.T.; Bennewith, K.L.; Nicolau, M.; Dornhofer, N.; Kong, C.; Le, Q.T.; Chi, J.T.; Jeffrey, S.S.; Giaccia, A.J. Lysyl oxidase is essential for hypoxia-induced metastasis. Nature 2006, 440, 1222–1226. [Google Scholar] [CrossRef] [PubMed]
- Ji, F.; Wang, Y.; Qiu, L.; Li, S.; Zhu, J.; Liang, Z.; Wan, Y.; Di, W. Hypoxia inducible factor 1alpha-mediated LOX expression correlates with migration and invasion in epithelial ovarian cancer. Int. J. Oncol. 2013, 42, 1578–1588. [Google Scholar] [CrossRef] [PubMed]
- Ween, M.P.; Oehler, M.K.; Ricciardelli, C. Transforming growth Factor-Beta-Induced Protein (TGFBI)/(betaig-H3): A matrix protein with dual functions in ovarian cancer. Int. J. Mol. Sci. 2012, 13, 10461–10477. [Google Scholar] [CrossRef]
- Karlan, B.Y.; Dering, J.; Walsh, C.; Orsulic, S.; Lester, J.; Anderson, L.A.; Ginther, C.L.; Fejzo, M.; Slamon, D. POSTN/TGFBI-associated stromal signature predicts poor prognosis in serous epithelial ovarian cancer. Gynecol. Oncol. 2014, 132, 334–342. [Google Scholar] [CrossRef]
- Bhushan, R.; Grunhagen, J.; Becker, J.; Robinson, P.N.; Ott, C.E.; Knaus, P. miR-181a promotes osteoblastic differentiation through repression of TGF-beta signaling molecules. Int. J. Biochem. Cell Biol. 2013, 45, 696–705. [Google Scholar] [CrossRef]
- Young, M.F.; Kerr, J.M.; Termine, J.D.; Wewer, U.M.; Wang, M.G.; McBride, O.W.; Fisher, L.W. cDNA cloning, mRNA distribution and heterogeneity, chromosomal location, and RFLP analysis of human osteopontin (OPN). Genomics 1990, 7, 491–502. [Google Scholar] [CrossRef]
- Kim, J.H.; Skates, S.J.; Uede, T.; Wong, K.K.; Schorge, J.O.; Feltmate, C.M.; Berkowitz, R.S.; Cramer, D.W.; Mok, S.C. Osteopontin as a potential diagnostic biomarker for ovarian cancer. Jama 2002, 287, 1671–1679. [Google Scholar] [CrossRef]
- Bao, L.H.; Sakaguchi, H.; Fujimoto, J.; Tamaya, T. Osteopontin in metastatic lesions as a prognostic marker in ovarian cancers. J. Biomed. Sci. 2007, 14, 373–381. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Samant, R.S.; Shevde, L.A. Nonclassical activation of Hedgehog signaling enhances multidrug resistance and makes cancer cells refractory to Smoothened-targeting Hedgehog inhibition. J. Biol. Chem. 2013, 288, 11824–11833. [Google Scholar] [CrossRef] [PubMed]
- Cui, R.; Takahashi, F.; Ohashi, R.; Yoshioka, M.; Gu, T.; Tajima, K.; Unnoura, T.; Iwakami, S.; Hirama, M.; Ishiwata, T.; et al. Osteopontin is involved in the formation of malignant pleural effusion in lung cancer. Lung Cancer (Amst. Neth.) 2009, 63, 368–374. [Google Scholar] [CrossRef] [PubMed]
- Huo, X.; Zhang, K.; Yi, L.; Mo, Y.; Liang, Y.; Zhao, J.; Zhang, Z.; Xu, Y.; Zhen, G. Decreased epithelial and plasma miR-181b-5p expression associates with airway eosinophilic inflammation in asthma. Clin. Exp. Allergy J. Br. Soc. Allergy Clin. Immunol. 2016, 46, 1281–1290. [Google Scholar] [CrossRef]
- Helleman, J.; Jansen, M.P.; Span, P.N.; van Staveren, I.L.; Massuger, L.F.; Meijer-van Gelder, M.E.; Sweep, F.C.; Ewing, P.C.; van der Burg, M.E.; Stoter, G.; et al. Molecular profiling of platinum resistant ovarian cancer. Int. J. Cancer 2006, 118, 1963–1971. [Google Scholar] [CrossRef]
- Stelcer, E.; Kulcenty, K.; Rucinski, M.; Jopek, K.; Richter, M.; Trzeciak, T.; Suchorska, W.M. The Role of MicroRNAs in Early Chondrogenesis of Human Induced Pluripotent Stem Cells (hiPSCs). Int. J. Mol. Sci. 2019, 20, 4371. [Google Scholar] [CrossRef]
- Kulcenty, K.; Wroblewska, J.P.; Rucinski, M.; Kozlowska, E.; Jopek, K.; Suchorska, W.M. MicroRNA Profiling During Neural Differentiation of Induced Pluripotent Stem Cells. Int. J. Mol. Sci. 2019, 20, 3651. [Google Scholar] [CrossRef]
- Gautier, L.; Cope, L.; Bolstad, B.M.; Irizarry, R.A. Affy—Analysis of Affymetrix GeneChip data at the probe level. Bioinformatics (Oxf. Engl.) 2004, 20, 307–315. [Google Scholar] [CrossRef]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]
- Cava, C.; Colaprico, A.; Bertoli, G.; Graudenzi, A.; Silva, T.C.; Olsen, C.; Noushmehr, H.; Bontempi, G.; Mauri, G.; Castiglioni, I. SpidermiR: An R/Bioconductor Package for Integrative Analysis with miRNA Data. Int. J. Mol. Sci. 2017, 18, 274. [Google Scholar] [CrossRef]
- Deng, M.; Brägelmann, J.; Kryukov, I.; Saraiva-Agostinho, N.; Perner, S. FirebrowseR: An R client to the Broad Institute’s Firehose Pipeline. Database 2017, 6. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Szyszka, M.; Paschke, L.; Tyczewska, M.; Jopek, K.; Celichowski, P.; Milecka, P.; Sultanova, G.; Stelcer, E.; Malinska, A.; Malendowicz, L.K.; et al. Analysis of transcriptome, selected intracellular signaling pathways, proliferation and apoptosis of LNCaP cells exposed to high leptin concentrations. Int. J. Mol. Sci. 2019, 20, e5412. [Google Scholar] [CrossRef] [PubMed]
- Weinstein, J.N.; Collisson, E.A.; Mills, G.B.; Shaw, K.R.M.; Ozenberger, B.A.; Ellrott, K.; Sander, C.; Stuart, J.M.; Chang, K.; Creighton, C.J.; et al. The cancer genome atlas pan-cancer analysis project. Nat. Genet. 2013, 45, 1113–1120. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | miRBase Accession Number | Fold Change (adj.p.val) | ||||
|---|---|---|---|---|---|---|
| W1PR1 | W1PR2 | W1CR | W1TR | W1DR | ||
| hsa-let-7c | MIMAT0000064 | 5.40 (0.001) | N.S | N.S | N.S | N.S |
| hsa-miR-100 | MIMAT0000098 | −5.01 (0.0003) | N.S | N.S | N.S | N.S |
| hsa-miR-105 | MIMAT0000102 | −19.22 (4.6 × 10−7) | −21.80 (1.5 × 10−6) | −15.92 (1.5 × 10−5) | N.S | N.S |
| hsa-miR-10a | MIMAT0000253 | −34.42 (2.3 × 10−7) | −9.07 (0.0002) | N.S | N.S | N.S |
| hsa-miR-1271 | MIMAT0005796 | 9.08 (0.004) | N.S | 6.24 (0.048) | 5.54 (0.049) | N.S |
| hsa-miR-1285 | MIMAT0005876 | N.S | 5.54 (0.01) | N.S | N.S | N.S |
| hsa-miR-143 | MIMAT0000437 | 8.81 (0.003) | N.S | N.S | N.S | N.S |
| hsa-miR-145 | MIMAT0000437 | 12.07 (4.8 × 10−5) | N.S | N.S | 7.05 (0.002) | N.S |
| hsa-miR-146b-5p | MIMAT0002809 | −39.13 (3.5 × 10−8) | N.S | N.S | N.S | −6.93 (0.0003) |
| hsa-miR-152 | MIMAT0000438 | 18.19 (6.5 × 10−5) | 16.52 (0.0003) | 8.53 (0.0006) | N.S | N.S |
| hsa-miR-181a | MIMAT0000256 | N.S | N.S | N.S | −12.28 (0.0003) | N.S |
| hsa-miR-181b | MIMAT0000257 | N.S | N.S | N.S | −9.39 (1.4 × 10−7) | N.S |
| hsa-miR-18b | MIMAT0001412 | −10.22 (0.0006) | N.S | N.S | N.S | N.S |
| hsa-miR-193a-5p | MIMAT0004614 | 5.32 (0.01) | N.S | N.S | N.S | N.S |
| hsa-miR-193b | MIMAT0002819 | 7.18 (0.01) | N.S | N.S | N.S | N.S |
| hsa-miR-195 | MIMAT0000461 | 13.94 (0.003) | N.S | 6.67 (0.046) | N.S | N.S |
| hsa-miR-199a-3p | MIMAT0000462 | N.S | N.S | N.S | N.S | −23.09 (0.01) |
| hsa-miR-199b-3p | MIMAT0000464 | N.S | N.S | N.S | N.S | −28.18 (0.02) |
| hsa-miR-203 | MIMAT0000264 | 14.45 (6.9 × 10−6) | N.S | 22.33 (1.5 × 10−5) | N.S | N.S |
| hsa-miR-205 | MIMAT0000465 | N.S | N.S | N.S | N.S | 6.23 (0.004) |
| hsa-miR-20b | MIMAT0000466 | −8.87 (6 × 10−8) | N.S | N.S | N.S | N.S |
| hsa-miR-214 | MIMAT0000271 | N.S | N.S | N.S | N.S | −26.05 (0.001) |
| hsa-miR-23b | MIMAT0000468 | 6.06 (0.0009) | N.S | N.S | N.S | N.S |
| hsa-miR-23c | MIMAT0000469 | 7.47 (0.02) | N.S | N.S | N.S | N.S |
| hsa-miR-27b | MIMAT0000470 | 9.21 (0.0003) | N.S | N.S | N.S | N.S |
| hsa-miR-29a | MIMAT0000086 | N.S | N.S | N.S | −22.04 (0.0009) | N.S |
| hsa-miR-30a | MIMAT0000087 | N.S | 14.82 (0.001) | N.S | N.S | N.S |
| hsa-miR-31 | MIMAT0000089 | −217.59 (4.6 × 10−5) | −51.72 (0.003) | −13.51 (0.048) | −36.90 (0.008) | N.S |
| hsa-miR-335 | MIMAT0000765 | N.S | N.S | N.S | 21.31 (0.0009) | N.S |
| hsa-miR-363 | MIMAT0000707 | −16.37 (1.55 × 10−7) | N.S | N.S | N.S | N.S |
| hsa-miR-4269 | MIMAT0016897 | N.S | N.S | N.S | N.S | −5.13 (0.04) |
| hsa-miR-449c | MIMAT0016897 | −11.97 (0.006) | −8.09 (0.04) | −5.61 (0.049) | −6.53 (0.047) | N.S |
| hsa-miR-4721 | MIMAT0019835 | N.S | 7.18 (0.03) | N.S | N.S | N.S |
| hsa-miR-497 | MIMAT0002820 | 8.22 (0.02) | N.S | N.S | N.S | N.S |
| hsa-miR-501-5p | MIMAT0002872 | N.S | −5.60 (0.002) | N.S | N.S | N.S |
| hsa-miR-572 | MIMAT0003237 | N.S | 6.23 (0.04) | N.S | N.S | N.S |
| hsa-miR-708 | MIMAT0004926 | N.S | N.S | 5.52 (0.007) | N.S | N.S |
| hsa-miR-767-5p | MIMAT0003882 | −20.61 (9 × 10−6) | −19.99 (3 × 10−5) | −13.95 (0.0004) | N.S | N.S |
| hsa-miR-875-3p | MIMAT0004923 | −7.93 (0.02) | −7.32 (0.048) | N.S | N.S | −6.28 (0.049) |
| hsa-miR-99a | MIMAT0000097 | 11.02 (0.004) | N.S | N.S | N.S | N.S |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kazmierczak, D.; Jopek, K.; Sterzynska, K.; Ginter-Matuszewska, B.; Nowicki, M.; Rucinski, M.; Januchowski, R. The Significance of MicroRNAs Expression in Regulation of Extracellular Matrix and Other Drug Resistant Genes in Drug Resistant Ovarian Cancer Cell Lines. Int. J. Mol. Sci. 2020, 21, 2619. https://doi.org/10.3390/ijms21072619
Kazmierczak D, Jopek K, Sterzynska K, Ginter-Matuszewska B, Nowicki M, Rucinski M, Januchowski R. The Significance of MicroRNAs Expression in Regulation of Extracellular Matrix and Other Drug Resistant Genes in Drug Resistant Ovarian Cancer Cell Lines. International Journal of Molecular Sciences. 2020; 21(7):2619. https://doi.org/10.3390/ijms21072619
Chicago/Turabian StyleKazmierczak, Dominika, Karol Jopek, Karolina Sterzynska, Barbara Ginter-Matuszewska, Michal Nowicki, Marcin Rucinski, and Radoslaw Januchowski. 2020. "The Significance of MicroRNAs Expression in Regulation of Extracellular Matrix and Other Drug Resistant Genes in Drug Resistant Ovarian Cancer Cell Lines" International Journal of Molecular Sciences 21, no. 7: 2619. https://doi.org/10.3390/ijms21072619
APA StyleKazmierczak, D., Jopek, K., Sterzynska, K., Ginter-Matuszewska, B., Nowicki, M., Rucinski, M., & Januchowski, R. (2020). The Significance of MicroRNAs Expression in Regulation of Extracellular Matrix and Other Drug Resistant Genes in Drug Resistant Ovarian Cancer Cell Lines. International Journal of Molecular Sciences, 21(7), 2619. https://doi.org/10.3390/ijms21072619