Sublethal Radiation Affects Antigen Processing and Presentation Genes to Enhance Immunogenicity of Cancer Cells


Abstract
1. Introduction
2. Results
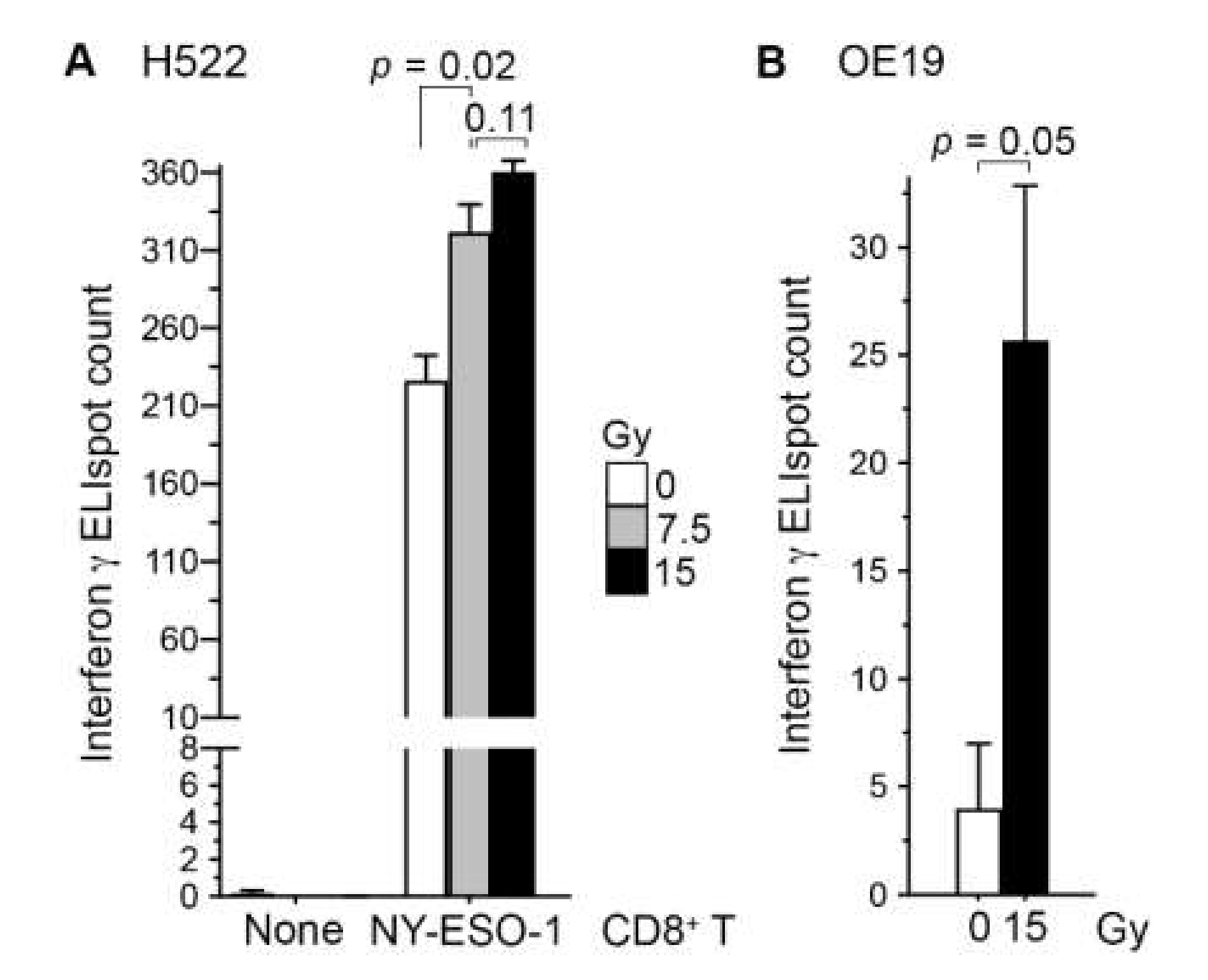
2.1. Irradiation of Cancer Cells Can Enhance Their Recognition by CD8+ T Cells
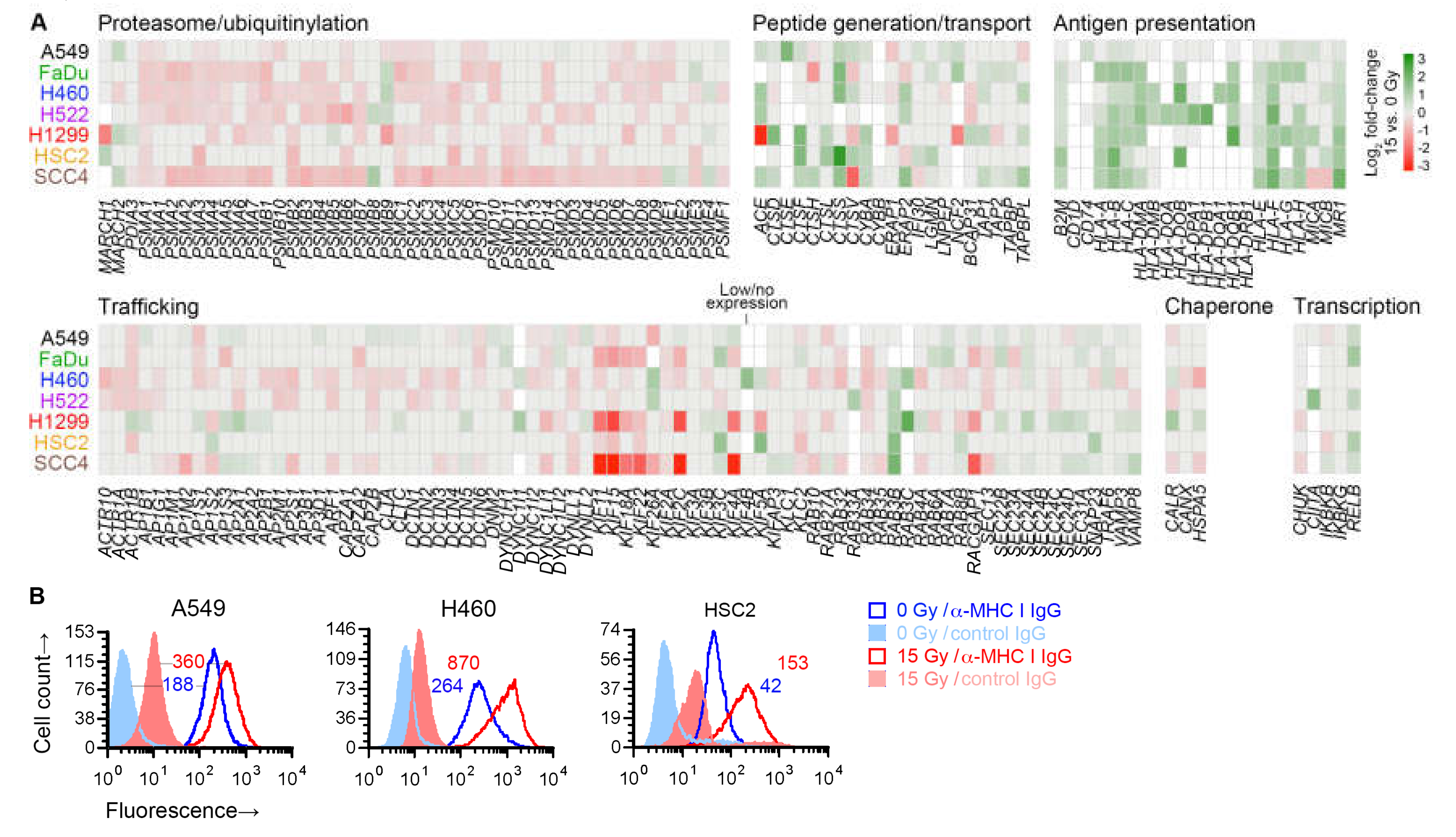
2.2. Cell Surface Proteins of Tumor Antigens May Not Be Increased by Radiation Treatment of Cancer Cells
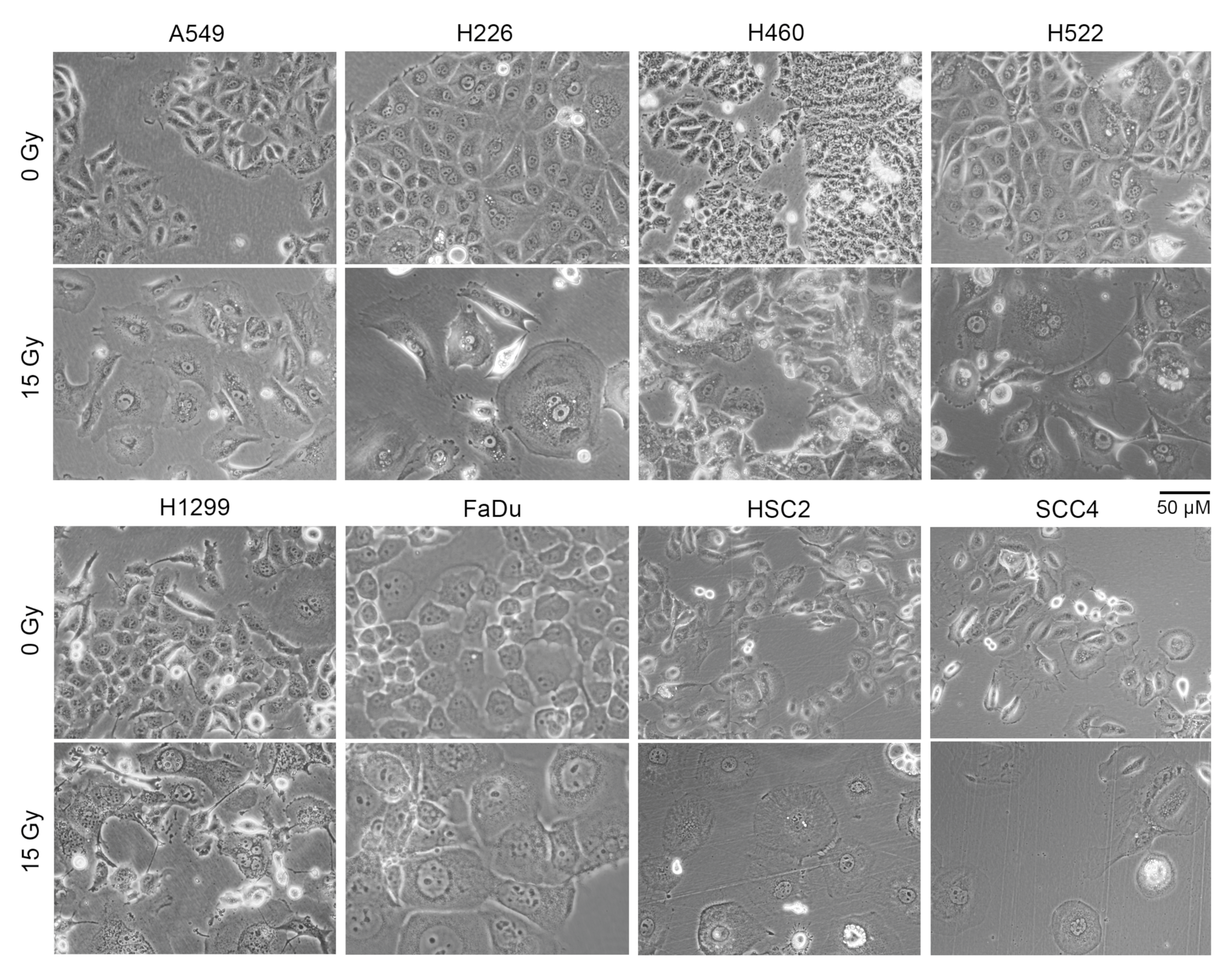
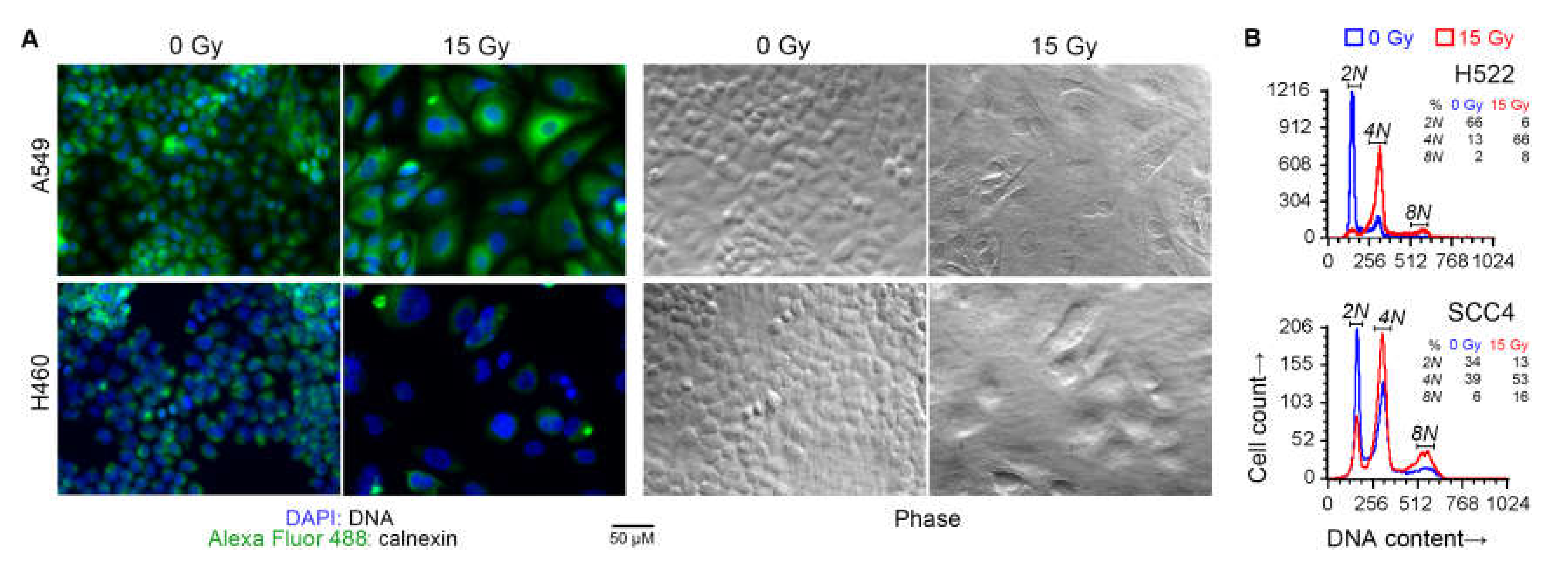
2.3. Irradiated Cancer Cells Were Larger Because of Mitosis Arrest and Polyploidization
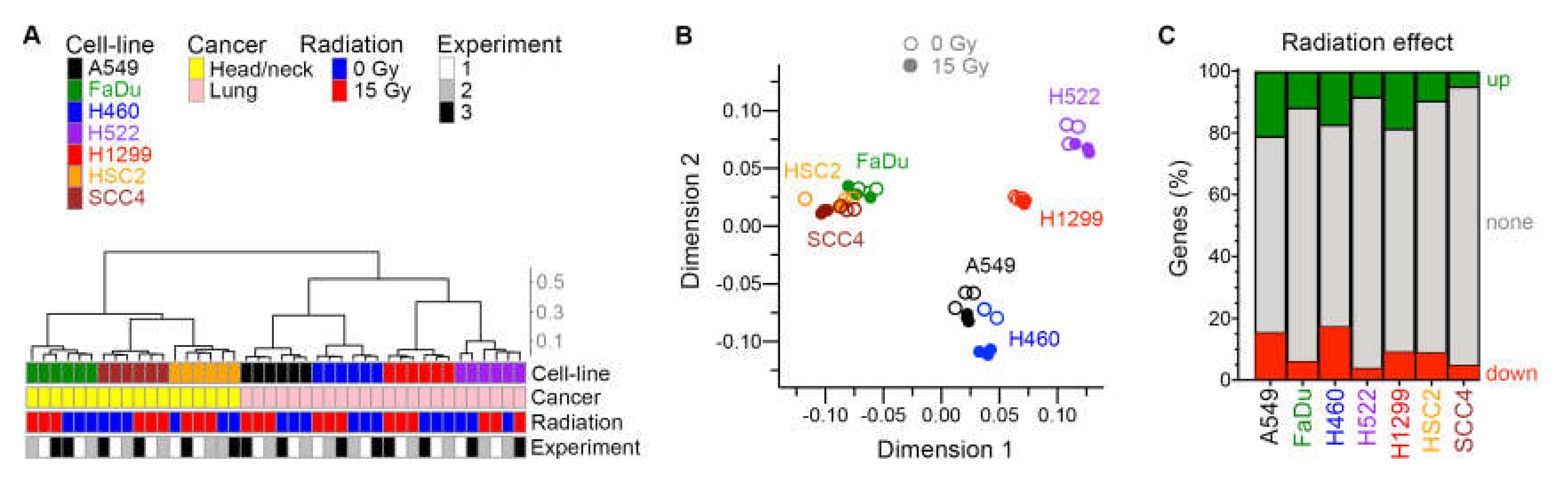
2.4. Radiation Had a Greater Effect on Gene Expression of Cells That Were More Proliferative
2.5. Gene Sets Downregulated by Radiation Were Predominantly Proliferation-Related
2.6. Radiation Enhanced Expression of Subsets of Antigen Processing and Presentation Genes
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Their Culture
4.2. Radiation Treatment of Cells
4.3. NY-ESO-1-Specific T-Cell Activation Assay
4.4. Antibodies
4.5. Flow Cytometry
4.6. DNA Content Analysis
4.7. Immunofluorescence Microscopy of Cells
4.8. Total RNA Isolation
4.9. Reverse Transcription and PCR
4.10. Total RNA Sequencing
4.11. Processing of RNA Sequencing Data
4.12. Analyses of Gene Expression
4.13. Other
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AC | Adenocarcinoma |
| CTA | Cancer/testis antigen |
| FDR | False discovery rate |
| GSVA | Gene set variation analysis |
| HLA | Human leukocyte antigen |
| HNSCC | Head and neck squamous cell carcinoma |
| IgG | Immunoglobulin G |
| mAb | Monoclonal antibody |
| MHC | Major histocompatibility complex |
| NSCLC | Non-small cell lung cancer |
| NY-ESO-1 | New York esophageal squamous cell carcinoma 1 |
| SCC | Squamous cell lung carcinoma |
References
- Stone, H.B.; Peters, L.J.; Milas, L. Effect of host immune capability on radiocurability and subsequent transplantability of a murine fibrosarcoma. J. Natl. Cancer Inst. 1979, 63, 1229–1235. [Google Scholar] [PubMed]
- Meng, X.; Feng, R.; Yang, L.; Xing, L.; Yu, J. The role of radiation oncology in immuno-oncology. Oncologist 2019, 24, S42–S52. [Google Scholar] [CrossRef] [PubMed]
- Solanki, A.A.; Bossi, A.; Efstathiou, J.A.; Lock, D.; Mondini, M.; Ramapriyan, R.; Welsh, J.; Kang, J. Combining immunotherapy with radiotherapy for the treatment of genitourinary malignancies. Eur. Urol. Oncol. 2019, 2, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Zhang, J. Immunotherapy-based combination strategies for treatment of gastrointestinal cancers: Current status and future prospects. Front. Med. 2019, 13, 12–23. [Google Scholar] [CrossRef]
- Lee, L.; Matulonis, U. Immunotherapy and radiation combinatorial trials in gynecologic cancer: A potential synergy? Gynecol. Oncol. 2019, 154, 236–245. [Google Scholar] [CrossRef]
- Ko, E.C.; Raben, D.; Formenti, S.C. The integration of radiotherapy with immunotherapy for the treatment of non-small cell lung cancer. Clin. Cancer Res. 2018, 24, 5792–5806. [Google Scholar] [CrossRef]
- Karam, S.D.; Raben, D. Radioimmunotherapy for the treatment of head and neck cancer. Lancet Oncol. 2019, 20, e404–e416. [Google Scholar] [CrossRef]
- Gameiro, S.R.; Jammeh, M.L.; Wattenberg, M.M.; Tsang, K.Y.; Ferrone, S.; Hodge, J.W. Radiation-induced immunogenic modulation of tumor enhances antigen processing and calreticulin exposure, resulting in enhanced T-cell killing. Oncotarget 2014, 5, 403–416. [Google Scholar] [CrossRef]
- Kaur, P.; Asea, A. Radiation-induced effects and the immune system in cancer. Front. Oncol. 2012, 2, 191. [Google Scholar] [CrossRef]
- Lhuillier, C.; Rudqvist, N.P.; Elemento, O.; Formenti, S.C.; Demaria, S. Radiation therapy and anti-tumor immunity: Exposing immunogenic mutations to the immune system. Genome Med. 2019, 11, 40. [Google Scholar] [CrossRef]
- Spiotto, M.; Fu, Y.X.; Weichselbaum, R.R. The intersection of radiotherapy and immunotherapy: Mechanisms and clinical implications. Sci. Immunol. 2016, 1. [Google Scholar] [CrossRef] [PubMed]
- Natale, N.; Reiner, J.; Southam, C.M. Immunogenic efficacy of various syngeneic tumor cell preparations. Cancer 1971, 28, 1118–1125. [Google Scholar] [CrossRef]
- Wennerberg, E.; Vanpouille-Box, C.; Bornstein, S.; Yamazaki, T.; Demaria, S.; Galluzzi, L. Immune recognition of irradiated cancer cells. Immunol. Rev. 2017, 280, 220–230. [Google Scholar] [CrossRef] [PubMed]
- Thomas, R.; Al-Khadairi, G.; Roelands, J.; Hendrickx, W.; Dermime, S.; Bedognetti, D.; Decock, J. NY-ESO-1 based immunotherapy of cancer: Current perspectives. Front. Immunol. 2018, 9, 947. [Google Scholar] [CrossRef]
- Matsuzaki, J.; Tsuji, T.; Luescher, I.F.; Shiku, H.; Mineno, J.; Okamoto, S.; Old, L.J.; Shrikant, P.; Gnjatic, S.; Odunsi, K. Direct tumor recognition by a human CD4(+) T-cell subset potently mediates tumor growth inhibition and orchestrates anti-tumor immune responses. Sci. Rep. 2015, 5, 14896. [Google Scholar] [CrossRef]
- Chueh, A.C.; Liew, M.S.; Russell, P.A.; Walkiewicz, M.; Jayachandran, A.; Starmans, M.H.W.; Boutros, P.C.; Wright, G.; Barnett, S.A.; Mariadason, J.M.; et al. Promoter hypomethylation of NY-ESO-1, association with clinicopathological features and PD-L1 expression in non-small cell lung cancer. Oncotarget 2017, 8, 74036–74048. [Google Scholar] [CrossRef]
- Singh, A.K.; Winslow, T.B.; Kermany, M.H.; Goritz, V.; Heit, L.; Miller, A.; Hoffend, N.C.; Stein, L.C.; Kumaraswamy, L.K.; Warren, G.W.; et al. A pilot study of stereotactic body radiation therapy combined with cytoreductive nephrectomy for metastatic renal cell carcinoma. Clin. Cancer Res. 2017, 23, 5055–5065. [Google Scholar] [CrossRef]
- Eleftheriadou, I.; Brett, S.; Domogala, A.; Patasic, L.; Kijewska, M.A.; Soor, K.; Georgouli, M.; Dopierala, J.; Fisher, P.; Jing, J.; et al. 1229P—NY-ESO-1 and LAGE1A: An emerging target for cell therapies in solid tumours. Ann. Oncol. 2019, 30, v503. [Google Scholar] [CrossRef]
- Amundson, S.A.; Do, K.T.; Vinikoor, L.C.; Lee, R.A.; Koch-Paiz, C.A.; Ahn, J.; Reimers, M.; Chen, Y.; Scudiero, D.A.; Weinstein, J.N.; et al. Integrating global gene expression and radiation survival parameters across the 60 cell lines of the National Cancer Institute Anticancer Drug Screen. Cancer Res. 2008, 68, 415–424. [Google Scholar] [CrossRef]
- Rangan, S.R. A new human cell line (FaDu) from a hypopharyngeal carcinoma. Cancer 1972, 29, 117–121. [Google Scholar] [CrossRef]
- Cowley, G.S.; Weir, B.A.; Vazquez, F.; Tamayo, P.; Scott, J.A.; Rusin, S.; East-Seletsky, A.; Ali, L.D.; Gerath, W.F.; Pantel, S.E.; et al. Parallel genome-scale loss of function screens in 216 cancer cell lines for the identification of context-specific genetic dependencies. Sci. Data 2014, 1, 140035. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.; Quan, F.; Li, H.H.; Yao, X.B.; Zhao, Q.; Zhao, R.M. The radiosensitizing effect of resveratrol on hopypharyngeal carcinoma cell line FADU and its effect on the cell cycle. Zhongguo Zhong Xi Yi Jie He Za Zhi 2015, 35, 699–703. [Google Scholar] [PubMed]
- Song, S.Y.; Das, A.K.; Minna, J.D. Comparison between concurrent and sequential chemoradiation for non-small cell lung cancer in vitro. Oncol. Lett. 2014, 7, 307–310. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Shintani, S.; Mihara, M.; Li, C.; Nakahara, Y.; Hino, S.; Nakashiro, K.; Hamakawa, H. Up-regulation of DNA-dependent protein kinase correlates with radiation resistance in oral squamous cell carcinoma. Cancer Sci. 2003, 94, 894–900. [Google Scholar] [CrossRef]
- Zhang, M.; Rose, B.; Lee, C.S.; Hong, A.M. In vitro 3-dimensional tumor model for radiosensitivity of HPV positive OSCC cell lines. Cancer Biol. Ther 2015, 16, 1231–1240. [Google Scholar] [CrossRef]
- Lu, Y.C.; Weng, W.C.; Lee, H. Functional roles of calreticulin in cancer biology. Biomed. Res. Int. 2015, 2015, 526524. [Google Scholar] [CrossRef]
- Syrkina, M.S.; Rubtsov, M.A. MUC1 in cancer immunotherapy—New hope or phantom menace? Biochemistry (Moscow) 2019, 84, 773–781. [Google Scholar] [CrossRef]
- Hanzelmann, S.; Castelo, R.; Guinney, J. GSVA: Gene set variation analysis for microarray and RNA-seq data. BMC Bioinform. 2013, 14, 7. [Google Scholar] [CrossRef]
- Kelly, A.; Trowsdale, J. Genetics of antigen processing and presentation. Immunogenetics 2019, 71, 161–170. [Google Scholar] [CrossRef]
- Wang, J.-S.; Wang, H.-J.; Qian, H.-L. Biological effects of radiation on cancer cells. Mil. Med. Res. 2018, 5, 20. [Google Scholar] [CrossRef]
- Barcellos-Hoff, M.H.; Park, C.; Wright, E.G. Radiation and the microenvironment—Tumorigenesis and therapy. Nat. Rev. Cancer 2005, 5, 867–875. [Google Scholar] [CrossRef] [PubMed]
- Frey, B.; Rubner, Y.; Kulzer, L.; Werthmoller, N.; Weiss, E.M.; Fietkau, R.; Gaipl, U.S. Antitumor immune responses induced by ionizing irradiation and further immune stimulation. Cancer Immunol. Immunother. 2014, 63, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Chajon, E.; Castelli, J.; Marsiglia, H.; De Crevoisier, R. The synergistic effect of radiotherapy and immunotherapy: A promising but not simple partnership. Crit. Rev. Oncol. Hematol. 2017, 111, 124–132. [Google Scholar] [CrossRef]
- De Ruysscher, D. Combination of radiotherapy and immune treatment: First clinical data. Cancer Radiother. 2018, 22, 564–566. [Google Scholar] [CrossRef] [PubMed]
- Formenti, S.C.; Demaria, S. Combining radiotherapy and cancer immunotherapy: A paradigm shift. J. Natl. Cancer Inst. 2013, 105, 256–265. [Google Scholar] [CrossRef]
- Sharma, A.; Bode, B.; Wenger, R.H.; Lehmann, K.; Sartori, A.A.; Moch, H.; Knuth, A.; Boehmer, L.; Broek, M. Gamma-Radiation promotes immunological recognition of cancer cells through increased expression of cancer-testis antigens in vitro and in vivo. PLoS ONE 2011, 6, e28217. [Google Scholar] [CrossRef]
- Malamas, A.S.; Gameiro, S.R.; Knudson, K.M.; Hodge, J.W. Sublethal exposure to alpha radiation (223Ra dichloride) enhances various carcinomas’ sensitivity to lysis by antigen-specific cytotoxic T lymphocytes through calreticulin-mediated immunogenic modulation. Oncotarget 2016, 7, 86937–86947. [Google Scholar] [CrossRef]
- Garnett, C.T.; Palena, C.; Chakraborty, M.; Tsang, K.Y.; Schlom, J.; Hodge, J.W. Sublethal irradiation of human tumor cells modulates phenotype resulting in enhanced killing by cytotoxic T lymphocytes. Cancer Res. 2004, 64, 7985–7994. [Google Scholar] [CrossRef]
- Rock, K.L.; Reits, E.; Neefjes, J. Present yourself! By MHC class I and MHC class II molecules. Trends Immunol. 2016, 37, 724–737. [Google Scholar] [CrossRef]
- Axelrod, M.L.; Cook, R.S.; Johnson, D.B.; Balko, J.M. Biological consequences of MHC-II expression by tumor cells in cancer. Clin. Cancer Res. 2019, 25, 2392–2402. [Google Scholar] [CrossRef]
- Accolla, R.S.; Lombardo, L.; Abdallah, R.; Raval, G.; Forlani, G.; Tosi, G. Boosting the MHC class II-restricted tumor antigen presentation to CD4+ T helper cells: A critical issue for triggering protective immunity and re-orienting the tumor microenvironment toward an anti-tumor state. Front. Oncol. 2014, 4, 32. [Google Scholar] [CrossRef] [PubMed]
- Karamooz, E.; Harriff, M.J.; Lewinsohn, D.M. MR1-dependent antigen presentation. Semin. Cell Dev. Biol. 2018, 84, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Crowther, M.D.; Dolton, G.; Legut, M.; Caillaud, M.E.; Lloyd, A.; Attaf, M.; Galloway, S.A.E.; Rius, C.; Farrell, C.P.; Szomolay, B.; et al. Genome-wide CRISPR-Cas9 screening reveals ubiquitous T cell cancer targeting via the monomorphic MHC class I-related protein MR1. Nat. Immunol. 2020, 21, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Gettinger, S.; Choi, J.; Hastings, K.; Truini, A.; Datar, I.; Sowell, R.; Wurtz, A.; Dong, W.; Cai, G.; Melnick, M.A.; et al. Impaired HLA class I antigen processing and presentation as a mechanism of acquired resistance to immune checkpoint inhibitors in lung cancer. Cancer Discov. 2017, 7, 1420–1435. [Google Scholar] [CrossRef] [PubMed]
- Seliger, B.; Maeurer, M.J.; Ferrone, S. Antigen-processing machinery breakdown and tumor growth. Immunol. Today 2000, 21, 455–464. [Google Scholar] [CrossRef]
- Wang, S.; He, Z.; Wang, X.; Li, H.; Liu, X.S. Antigen presentation and tumor immunogenicity in cancer immunotherapy response prediction. Elife 2019, 8. [Google Scholar] [CrossRef]
- Chiriva-Internati, M.; Grizzi, F.; Pinkston, J.; Morrow, K.J.; D’Cunha, N.; Frezza, E.E.; Muzzio, P.C.; Kast, W.M.; Cobos, E. Gamma-radiation upregulates MHC class I/II and ICAM-I molecules in multiple myeloma cell lines and primary tumors. Cell. Dev. Biol. Anim. 2006, 42, 89–95. [Google Scholar] [CrossRef]
- Reits, E.A.; Hodge, J.W.; Herberts, C.A.; Groothuis, T.A.; Chakraborty, M.; Wansley, E.K.; Camphausen, K.; Luiten, R.M.; De Ru, A.H.; Neijssen, J.; et al. Radiation modulates the peptide repertoire, enhances MHC class I expression, and induces successful antitumor immunotherapy. J. Exp. Med. 2006, 203, 1259–1271. [Google Scholar] [CrossRef]
- Santin, A.D.; Hermonat, P.L.; Hiserodt, J.C.; Chiriva-Internati, M.; Woodliff, J.; Theus, J.W.; Barclay, D.; Pecorelli, S.; Parham, G.P. Effects of irradiation on the expression of major histocompatibility complex class I antigen and adhesion costimulation molecules ICAM-1 in human cervical cancer. Int. J. Radiat. Oncol. Biol. Phys. 1997, 39, 737–742. [Google Scholar] [CrossRef]
- Larkins, B.A.; Dilkes, B.P.; Dante, R.A.; Coelho, C.M.; Woo, Y.M.; Liu, Y. Investigating the hows and whys of DNA endoreduplication. J. Exp. Bot. 2001, 52, 183–192. [Google Scholar] [CrossRef]
- Kaur, E.; Rajendra, J.; Jadhav, S.; Shridhar, E.; Goda, J.S.; Moiyadi, A.; Dutt, S. Radiation-induced homotypic cell fusions of innately resistant glioblastoma cells mediate their sustained survival and recurrence. Carcinogenesis 2015, 36, 685–695. [Google Scholar] [CrossRef] [PubMed]
- Mirzayans, R.; Andrais, B.; Scott, A.; Wang, Y.W.; Kumar, P.; Murray, D. Multinucleated giant cancer cells produced in response to ionizing radiation retain viability and replicate their genome. Int. J. Mol. Sci. 2017, 18, 360. [Google Scholar] [CrossRef] [PubMed]
- Erenpreisa, J.; Ivanov, A.; Wheatley, S.P.; Kosmacek, E.A.; Ianzini, F.; Anisimov, A.P.; Mackey, M.; Davis, P.J.; Plakhins, G.; Illidge, T.M. Endopolyploidy in irradiated p53-deficient tumour cell lines: Persistence of cell division activity in giant cells expressing Aurora-B kinase. Cell Biol. Int. 2008, 32, 1044–1056. [Google Scholar] [CrossRef] [PubMed]
- Schwarz-Finsterle, J.; Scherthan, H.; Huna, A.; Gonzalez, P.; Mueller, P.; Schmitt, E.; Erenpreisa, J.; Hausmann, M. Volume increase and spatial shifts of chromosome territories in nuclei of radiation-induced polyploidizing tumour cells. Mutat. Res. 2013, 756, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Rene, A.A.; Nardone, R.M. The effect of gamma radiation on cell enlargement and size distribution of strain L. Curr. Mod. Biol. 1968, 2, 207–214. [Google Scholar] [CrossRef]
- Bairoch, A. The cellosaurus, a cell-line knowledge resource. J. Biomol. Tech. 2018, 29, 25–38. [Google Scholar] [CrossRef]
- Scholtalbers, J.; Boegel, S.; Bukur, T.; Byl, M.; Goerges, S.; Sorn, P.; Loewer, M.; Sahin, U.; Castle, J.C. TCLP: An online cancer cell line catalogue integrating HLA type, predicted neo-epitopes, virus and gene expression. Genome Med. 2015, 7, 118. [Google Scholar] [CrossRef]
- Jung, M.; Ramankulov, A.; Roigas, J.; Johannsen, M.; Ringsdorf, M.; Kristiansen, G.; Jung, K. In search of suitable reference genes for gene expression studies of human renal cell carcinoma by real-time PCR. BMC Mol. Biol. 2007, 8, 47. [Google Scholar] [CrossRef]
- Hamann, M.V.; Mullers, E.; Reh, J.; Stanke, N.; Effantin, G.; Weissenhorn, W.; Lindemann, D. The cooperative function of arginine residues in the Prototype Foamy Virus Gag C-terminus mediates viral and cellular RNA encapsidation. Retrovirology 2014, 11, 87. [Google Scholar] [CrossRef]
- Meissner, T.B.; Li, A.; Biswas, A.; Lee, K.H.; Liu, Y.J.; Bayir, E.; Iliopoulos, D.; Van den Elsen, P.J.; Kobayashi, K.S. NLR family member NLRC5 is a transcriptional regulator of MHC class I genes. Proc. Natl. Acad. Sci. USA 2010, 107, 13794–13799. [Google Scholar] [CrossRef]
- Wang, X.; Xiong, L.; Yu, G.; Li, D.; Peng, T.; Luo, D.; Xu, J. Cathepsin S silencing induces apoptosis of human hepatocellular carcinoma cells. Am. J. Transl. Res. 2015, 7, 100–110. [Google Scholar] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Smyth, G.K.; Shi, W. The Subread aligner: Fast, accurate and scalable read mapping by seed-and-vote. Nucleic Acids Res. 2013, 41, e108. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]
- Smyth, G. Limma: Linear models for microarray data. In Bioinformatics and Computational Biology Solutions using R and Bioconductor; Gentleman, R., Carey, V.J., Huber, W., Dudoit, S., Irizarry, R.A., Eds.; Springer: New York, NY, USA, 2005; pp. 397–420. [Google Scholar]
- Almeida, L.G.; Sakabe, N.J.; de Oliveira, A.R.; Silva, M.C.; Mundstein, A.S.; Cohen, T.; Chen, Y.T.; Chua, R.; Gurung, S.; Gnjatic, S.; et al. CTdatabase: A knowledge-base of high-throughput and curated data on cancer-testis antigens. Nucleic Acids Res. 2009, 37, D816–D819. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parent Tissue | Parent Cancer | Patient Demographic | Karyotype Modal Number | Doubling Time c (Hours) | SF2 d | SF8 | |
|---|---|---|---|---|---|---|---|
| A549 | Lung | Lung AC | White M, 58 y | 66 | 22 | 0.68 | 0.05 |
| FaDu | Hypopharynx | Hypopharynx SCC | White M, 56 y | 64 | 50 | 0.72 | |
| H226 | Metastatic pleural effusion | Lung SCC | M | 47 | 64 | 0.63 | 0.12 |
| H460 | Metastatic pleural effusion | Lung LCC | M | 57 | 23 | 0.75 | 0.25 |
| H522 | Lung | Lung AC | White M, 58 y | 53 | 50 | 0.43 | 0.003 |
| H1299 | Metastatic lymph node | Lung NSCLC | White M, 43 y | 30 | 0.52 | ||
| HSC2 | Mouth | Mouth SCC | M, 69 y | 27–38 | 0.66 | ||
| SCC4 | Tongue | Tongue SCC | M, 55 y | 50–80 | 61–96 | 0.59 |
| Gene | Description | Log2 Fold-Change | P |
|---|---|---|---|
| ABTB2 | Ankyrin repeat and BTB (POZ) domain containing 2 | 0.3–1.9 | 2.3 × 10−43–1.4 × 10−2 |
| AMPD3 | Adenosine monophosphate deaminase 3 | 0.6–1.6 | 2.0 × 10−30–2.6 × 10−3 |
| BCL6 | B-cell CLL/lymphoma 6 | 0.3–1.9 | 1.2 × 10−5–1.2 × 10−2 |
| BTBD19 | BTB (POZ) domain containing 19 | 0.6–2.4 | 1.2 × 10−29–1.1 × 10−2 |
| CDKN1A | Cyclin-dependent kinase inhibitor 1A (p21, Cip1) | 0.9–3.0 | 2.1 × 10−165–1.7 × 10−5 |
| COL1A1 | Collagen, type I, alpha 1 | 0.6–1.9 | 1.3 × 10−13–1.2 × 10−3 |
| COL7A1 | Collagen, type VII, alpha 1 | 0.4–2.5 | 1.5 × 10−35–3.0 × 10−2 |
| DYSF | Dysferlin | 0.2–3.1 | 1.7 × 10−25–4.1 × 10−2 |
| FAM214A | Family with sequence similarity 214, member A | 0.3–1.8 | 3.9 × 10−50–2.1 × 10−2 |
| FAM71F2 | Family with sequence similarity 71, member F2 | 0.4–2.5 | 4.8 × 10−21–3.2 × 10−2 |
| FBXO32 | F-box protein 32 | 0.6–2.2 | 4.6 × 10−43–4.6 × 10−2 |
| HAP1 | Huntingtin-associated protein 1 | 0.7–1.8 | 1.4 × 10−8–1.1 × 10−2 |
| HIVEP1 | Human immunodeficiency virus type I enhancer binding protein 1 | 0.4–0.9 | 6.3 × 10−9–1.6 × 10−2 |
| HIVEP2 | Human immunodeficiency virus type I enhancer binding protein 2 | 0.7–1.4 | 4.5 × 10−25–8.0 × 10−9 |
| HLA-B | Major histocompatibility complex, class I, B | 0.4–1.9 | 1.3 × 10−33–4.5 × 10−6 |
| HLA-C | Major histocompatibility complex, class I, C | 0.3–1.4 | 2.7 × 10−22–2.2 × 10−2 |
| LAMA3 | Laminin, alpha 3 | 0.4–1.3 | 8.7 × 10−13–8.4 × 10−3 |
| LIPH | Lipase, member H | 0.4–2.2 | 7.9 × 10−12–7.5 × 10−3 |
| LSMEM1 | Leucine-rich single-pass membrane protein 1 | 0.8–1.7 | 7.6 × 10−10–4.6 × 10−2 |
| LTBP3 | Latent transforming growth factor beta binding protein 3 | 0.4–1.4 | 5.2 × 10−60–8.3 × 10−3 |
| MCAM | Melanoma cell adhesion molecule | 0.7–3.6 | 2.1 × 10−25–9.9 × 10−3 |
| MR1 | Major histocompatibility complex, class I-related | 0.2–2.0 | 3.7 × 10−93–3.3 × 10−2 |
| NEB | Nebulin | 0.3–2.8 | 7.6 × 10−24–3.4 × 10−2 |
| NEK10 | NIMA-related kinase 10 | 0.9–1.9 | 3.6 × 10−35–3.7 × 10−3 |
| NOTCH2NL | Notch 2 N-terminal like | 0.4–1.1 | 1.1 × 10−9–4.0 × 10−2 |
| PLA2G4C | Phospholipase A2, group IVC (cytosolic, calcium-independent) | 0.8–2.7 | 5.0 × 10−30–2.1 × 10−3 |
| PLEKHA6 | Pleckstrin homology domain containing, family A member 6 | 0.6–3.8 | 2.1 × 10−18–1.2 × 10−2 |
| PRDM1 | PR domain containing 1, with ZNF domain | 0.5–2.4 | 3.2 × 10−25–2.6 × 10−3 |
| RGL1 | Ral guanine nucleotide dissociation stimulator-like 1 | 0.4–2.4 | 1.9 × 10−98–4.3 × 10−2 |
| RP11-288H12.3 | Calcineurin like EF-hand protein 1 pseudogene (LOC729603) | 0.8–1.1 | 2.2 × 10−5–1.7 × 10−2 |
| RP11-734K2.4 | POC1B antisense RNA 1 (POC1B-AS1) | 0.8–1.8 | 1.4 × 10−9–8.4 × 10−4 |
| SERPINE1 | Serpin peptidase inhibitor, clade E, member 1 | 0.7–2.0 | 8.6 × 10−19–2.2 × 10−4 |
| SERPINE2 | Serpin peptidase inhibitor, clade E, member 2 | 0.7–2.8 | 8.1 × 10−58–4.2 × 10−4 |
| SGK1 | Serum/glucocorticoid regulated kinase 1 | 0.5–3.2 | 2.8 × 10−25–8.8 × 10−3 |
| STX3 | Syntaxin 3 | 0.4–1.6 | 4.7 × 10−15–1.2 × 10−2 |
| TAGLN | Transgelin | 0.2–3.2 | 5.3 × 10−37–1.8 × 10−2 |
| TNFAIP3 | Tumor necrosis factor, alpha-induced protein 3 | 1.0–2.7 | 1.1 × 10−28–1.5 × 10−3 |
| TRIB1 | Tribbles pseudokinase 1 | 0.4–0.9 | 4.6 × 10−10–2.9 × 10−2 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Punnanitinont, A.; Kannisto, E.D.; Matsuzaki, J.; Odunsi, K.; Yendamuri, S.; Singh, A.K.; Patnaik, S.K. Sublethal Radiation Affects Antigen Processing and Presentation Genes to Enhance Immunogenicity of Cancer Cells. Int. J. Mol. Sci. 2020, 21, 2573. https://doi.org/10.3390/ijms21072573
Punnanitinont A, Kannisto ED, Matsuzaki J, Odunsi K, Yendamuri S, Singh AK, Patnaik SK. Sublethal Radiation Affects Antigen Processing and Presentation Genes to Enhance Immunogenicity of Cancer Cells. International Journal of Molecular Sciences. 2020; 21(7):2573. https://doi.org/10.3390/ijms21072573
Chicago/Turabian StylePunnanitinont, Achamaporn, Eric D. Kannisto, Junko Matsuzaki, Kunle Odunsi, Sai Yendamuri, Anurag K. Singh, and Santosh K. Patnaik. 2020. "Sublethal Radiation Affects Antigen Processing and Presentation Genes to Enhance Immunogenicity of Cancer Cells" International Journal of Molecular Sciences 21, no. 7: 2573. https://doi.org/10.3390/ijms21072573
APA StylePunnanitinont, A., Kannisto, E. D., Matsuzaki, J., Odunsi, K., Yendamuri, S., Singh, A. K., & Patnaik, S. K. (2020). Sublethal Radiation Affects Antigen Processing and Presentation Genes to Enhance Immunogenicity of Cancer Cells. International Journal of Molecular Sciences, 21(7), 2573. https://doi.org/10.3390/ijms21072573