LncRNAs in HCV Infection and HCV-Related Liver Disease



Abstract
1. Hepatitis C Virus (HCV)
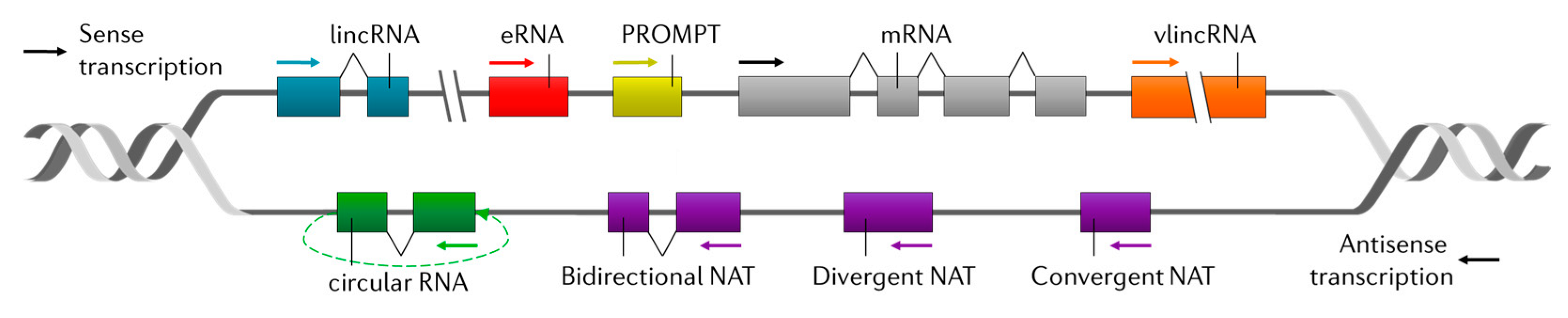
2. Long Non-Coding RNAs (lncRNAs)
3. General Considerations of HCV and lncRNAs
4. HCV lncRNAs
5. Cellular lncRNAs Related to HCV Cell Cycle and the Antiviral Response
5.1. LncRNAs that Affect the Viral Cell Cycle
5.2. LncRNAs that Affect the Antiviral Response
5.2.1. IFN-Related lncRNAs
5.2.2. IFN-Related lncRNAs that Affect HCV Replication
5.2.3. LncRNAs Affecting the IFN Synthesis Pathway
5.2.4. LncRNAs Affecting the Expression of Specific ISGs Located Nearby
5.2.5. LncRNAs Affecting General ISG Transcription
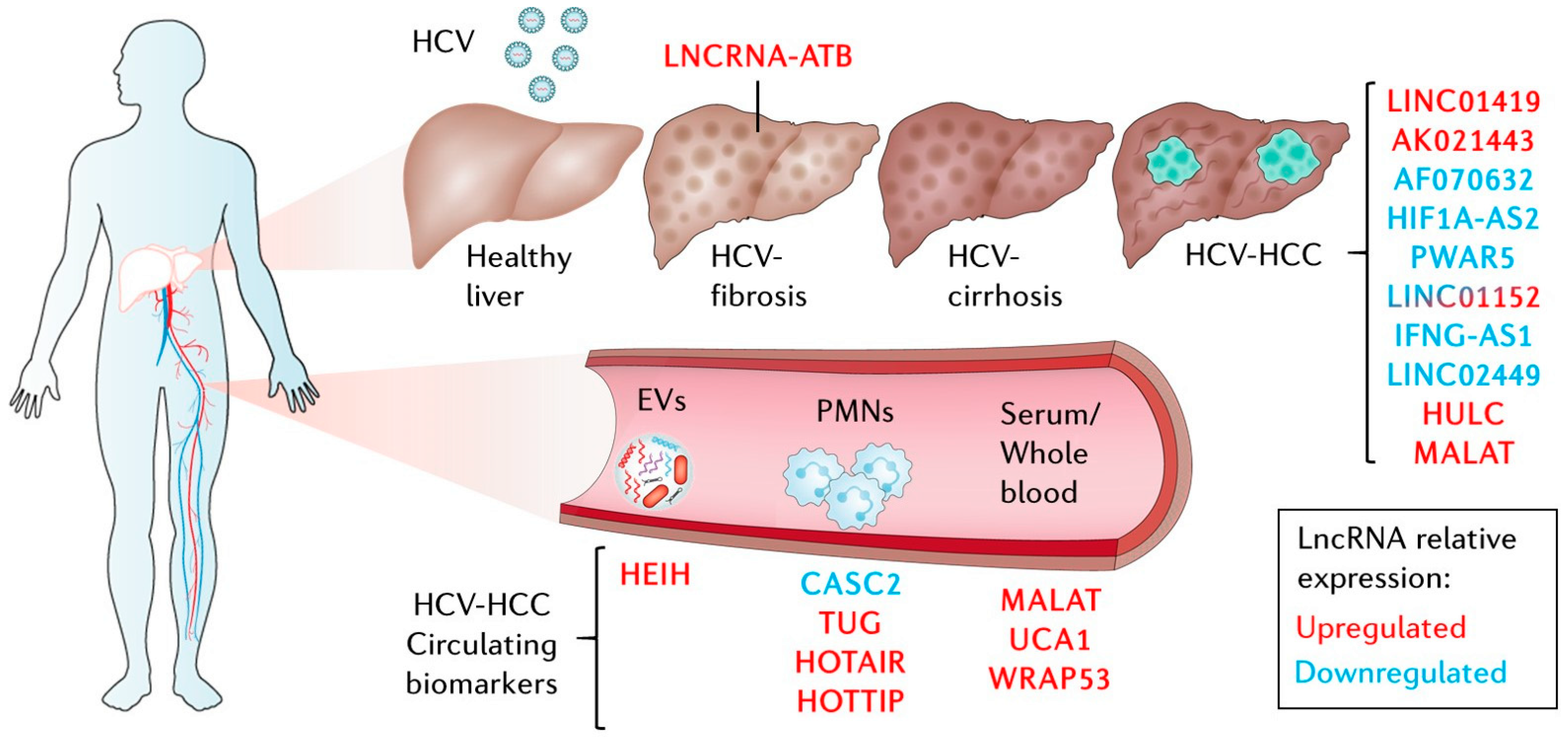
6. LncRNAs in HCV-Related Liver Diseases
6.1. LncRNAs Related to Extrahepatic Manifestations and HCV-Related Liver Fibrosis
6.2. LncRNAs Related to HCV-Related Hepatocellular Carcinoma (HCV-HCC)
6.3. LncRNAs in Liquid Biopsies as Biomarkers in HCV-HCC
6.4. LncRNAs with Potential HCV-HCC Roles From Cell Studies
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| LncRNAs | Long non-coding RNAs |
| HCV | Hepatitis C virus |
| HCC | Hepatocellular carcinoma |
| LDLR | Low-density lipoprotein receptor |
| CLDN1 | Claudin-1 |
| IRES | Internal ribosome entry site |
| IFN | Interferon |
| PAMPs | Pathogen-associated molecular patterns |
| LPS | Lipopolysaccharide |
| RIG-I | Retinoic acid-inducible gene I |
| UTR | Untranslated region |
| PKR | Protein kinase R |
| DDX3X | DEAD (Asp-Glu-Ala-Asp) box helicase 3 X-linked |
| TLR3 | Toll-like receptor 3 |
| dsRNA | Double-stranded RNA |
| MDA5 | Melanoma differentiation-associated protein 5 |
| MAVS | Mitochondrial antiviral signaling protein |
| TRIF | Toll-interleukin-1 receptor domain-containing adapter protein inducing interferon beta |
| NF-κB | Nuclear factor κb |
| IRF | Interferon regulatory factors |
| IFNAR | IFN-α/β receptor |
| JAK | Janus kinase |
| STAT | Signal transducer and activator of transcription |
| ISGF3 | IFN-stimulated gene factor 3 |
| ISG | IFN-stimulated genes |
| Mx | Myxoma resistance protein |
| TRIM | Tripartite motif containing |
| IFITM | Interferon induced transmembrane protein |
| CH25H | Cholesterol 25-hydroxylase |
| OAS | Oligoadenylate synthetase |
| IFIT | Interferon induced protein |
| GBP1 | Guanylate binding protein 1 |
| BST2 | Bone marrow stromal cell antigen 2 |
| SOCS3 | Suppressor of cytokine signaling 3 |
| PP2A | Protein phosphatase 2 |
| eIF2α | Eukaryotic translation initiation factor 2 alpha |
| ncRNAs | Non-coding RNAs |
| rRNAs | Ribosomal RNAs |
| tRNAs | Transfer RNAs |
| snRNAs | Small nuclear RNAs |
| snoRNAs | Small nucleolar RNAs |
| miRNAs | MicroRNAs |
| siRNAs | Small interfering RNAs |
| piRNAs | P-element-induced wimpy testis (PIWI)-interacting RNAs |
| mRNAs | Messenger RNAs |
| ORF | Open reading frame |
| LincRNAs | Long intergenic long non-coding RNAs |
| eRNAs | Enhancer RNAs |
| PROMPTs | Promoter upstream transcripts |
| NAT | Natural antisense transcript |
| vlincRNA | Very long lincRNA |
| HOTAIR | Homeobox C transcript antisense RNA |
| PVT1 | LncRNA plasmacytoma variant translocation 1 |
| MEG3 | Maternally expressed 3 |
| PANDAR | Promoter of CDKN1A (cyclin dependent kinase inhibitor 1A) antisense DNA damage-activated RNA |
| XRN1 | 5′-3′ exoribonuclease 1 |
| GAS5 | Growth arrest-specific transcript 5 |
| PRC2 | Polycomb repressive complex 2 |
| SIRT1 | Sirtuin 1 |
| HULC | Highly upregulated in liver cancer |
| RXRA | Retinoid X receptor alpha |
| IGF2-AS | Insulin-like growth factor 2 antisense RNA |
| 7SK | RNA component of 7SK nuclear ribonucleoprotein |
| PI4P | Phosphatidylinositol 4-phosphate |
| TSPOAP1-AS1 | Translocator protein associated protein 1 (TSPOAP1) antisense RNA 1 |
| MIR155HG | miRNA-155 host gene |
| pI:C | Polyinosinic–polycytidylic acid |
| PME-1 | Phosphatase methylesterase 1 |
| CSTB | Cystatin B |
| lncLRRC55-AS | LncRNA antisense to leucine rich repeat containing 55 |
| NEAT1 | Nuclear paraspeckle assembly transcript 1 |
| DDX60 | DExD/H-Box helicase 60 |
| lnc-Lsm3b | LncRNA U6 snRNA-associated sm-like protein |
| lnc-MxA | LncRNA interferon-induced GTP-binding protein MxA |
| CFAP58-DT | Cilia and flagella associated protein 58 (CFAP58) divergent transcript |
| EGFR | Epidermal growth factor receptor |
| CD81 | Cluster of differentiation 81 |
| IFI6 | Interferon alpha inducible protein 6 |
| BISPR | BST2 IFN-stimulated positive regulator |
| LUARIS | LncRNA upregulator of antiviral response interferon signaling |
| hnRNPU | Heterogeneous nuclear ribonucleoprotein U |
| ITPRIP-1 | Inositol 1,4,5-Trisphosphate Receptor Interacting Protein |
| ATF2 | Activating transcription factor 2 |
| HBV | Hepatitis B virus |
| cAMP | Cyclic adenosine monophosphate |
| NRIR | Negative regulator of the IFN response |
| CMPK2 | Cytidine/uridine monophosphate kinase 2 |
| CXCL10 | C-X-C motif chemokine ligand 10 |
| GCSIR | G protein-coupled receptor 55 (GPR55) cis regulatory suppressor of immune response RNA |
| EGOT | Eosinophil granule ontogeny transcript |
| EC | Endocannabinoid |
| SFV | Semliki Forest virus |
| CHC | Chronic HCV infection |
| LncRNA-ATB | Long noncoding RNA activated by TGFβ (transforming growth factor-beta) |
| COL1A1 | Alpha-1 type I collagen |
| α-SMA | Alpha-smooth muscle actin |
| HCV-HCC | HCV-related HCC |
| PRLH1 | p53-regulated lncRNA for homologous recombination (HR) repair 1 |
| TCGA | The cancer genome atlas network |
| HDV | Hepatitis Delta virus |
| aHIF | LncRNA antisense to HIF1A (hypoxia-inducible factor 1 alpha) |
| PAR5/PWAR5 | Prader Willi/Angelman region RNA 5 |
| EZH2 | Enhancer of zeste 2 polycomb repressive complex 2 subunit |
| TMEVPG1 | Theiler’s murine encephalomyelitis virus persistence candidate gene 1 |
| IFNG-AS1 | IFNG antisense RNA 1 |
| IL-23 | Interleukin 23 |
| HBx | HBV-encoded protein x |
| MALAT1 | Metastasis-associated lung adenocarcinoma transcript 1 |
| AFP | Alpha-fetoprotein |
| EVs | Extracellular vesicles |
| UCA1 | Urothelial carcinoma associated-1 |
| WRAP53 | Trp-Asp (WD) repeat containing antisense to TP53 (tumor protein p53) |
| HEIH | HCC up-regulated EZH2-associated long non-coding RNA |
| TUG1 | Taurine up-regulated 1 |
| CASC2 | Cancer susceptibility 2 |
| HOTTIP | Homeobox A (HOXA) distal transcript antisense RNA |
| NOP2 | Nucleolar protein 2 |
| MYC | Myc proto-oncogene protein |
| NORAD | Non-coding RNA activated by DNA damage |
| Wee1 | G2 checkpoint kinase |
References
- Webster, D.P.; Klenerman, P.; Dusheiko, G.M. Hepatitis C. Lancet 2015, 385, 1124–1135. [Google Scholar] [CrossRef]
- Bukh, J. The history of hepatitis C virus (HCV): Basic research reveals unique features in phylogeny, evolution and the viral life cycle with new perspectives for epidemic control. J. Hepatol. 2016, 65, S2–S21. [Google Scholar] [CrossRef]
- D’Ambrosio, R.; Degasperi, E.; Colombo, M.; Aghemo, A. Direct-acting antivirals: The endgame for hepatitis C? Curr. Opin. Virol. 2017, 24, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Ghany, M.G.; Strader, D.B.; Thomas, D.L.; Seeff, L.B. Diagnosis, management, and treatment of hepatitis C: An update. Hepatology 2009, 49, 1335–1374. [Google Scholar] [CrossRef] [PubMed]
- Llovet, J.M.; Villanueva, A. Effect of HCV clearance with direct-acting antiviral agents on HCC. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 561–562. [Google Scholar] [CrossRef] [PubMed]
- Neufeldt, C.J.; Cortese, M.; Acosta, E.G.; Bartenschlager, R. Rewiring cellular networks by members of the Flaviviridae family. Nat. Rev. Microbiol. 2018, 16, 125–142. [Google Scholar] [CrossRef]
- Catanese, M.T.; Uryu, K.; Kopp, M.; Edwards, T.J.; Andrus, L.; Rice, W.J.; Silvestry, M.; Kuhn, R.J.; Rice, C.M. Ultrastructural analysis of hepatitis C virus particles. Proc. Natl. Acad. Sci. USA 2013, 110, 9505–9510. [Google Scholar] [CrossRef]
- Ploss, A.; Evans, M.J.; Gaysinskaya, V.A.; Panis, M.; You, H.; de Jong, Y.P.; Rice, C.M. Human occludin is a hepatitis C virus entry factor required for infection of mouse cells. Nature 2009, 457, 882–886. [Google Scholar] [CrossRef]
- Meyers, N.L.; Fontaine, K.A.; Kumar, G.R.; Ott, M. Entangled in a membranous web: ER and lipid droplet reorganization during hepatitis C virus infection. Curr. Opin. Cell Biol. 2016, 41, 117–124. [Google Scholar] [CrossRef]
- Piñeiro, D.; Martinez-Salas, E. RNA structural elements of hepatitis C virus controlling viral RNA translation and the implications for viral pathogenesis. Viruses 2012, 4, 2233–2250. [Google Scholar] [CrossRef]
- Rusinova, I.; Forster, S.; Yu, S.; Kannan, A.; Masse, M.; Cumming, H.; Chapman, R.; Hertzog, P.J. INTERFEROME v2.0: An updated database of annotated interferon-regulated genes. Nucleic Acids Res. 2012, 41, D1040–D1046. [Google Scholar] [CrossRef] [PubMed]
- Porritt, R.A.; Hertzog, P.J. Dynamic control of type I IFN signalling by an integrated network of negative regulators. Trends Immunol. 2015, 36, 150–160. [Google Scholar] [CrossRef] [PubMed]
- Valadkhan, S.; Fortes, P. Regulation of the interferon response by lncRNAs in HCV infection. Front. Microbiol. 2018, 9, 181. [Google Scholar] [CrossRef] [PubMed]
- Saito, T.; Owen, D.M.; Jiang, F.; Marcotrigiano, J.; Gale, M. Innate immunity induced by composition-dependent RIG-I recognition of hepatitis C virus RNA. Nature 2008, 454, 523–527. [Google Scholar] [CrossRef]
- Arnaud, N.; Dabo, S.; Maillard, P.; Budkowska, A.; Kalliampakou, K.I.; Mavromara, P.; Garcin, D.; Hugon, J.; Gatignol, A.; Akazawa, D.; et al. Hepatitis C virus controls interferon production through PKR activation. PLoS ONE 2010, 5, e10575. [Google Scholar] [CrossRef]
- Arnaud, N.; Dabo, S.; Akazawa, D.; Fukasawa, M.; Shinkai-Ouchi, F.; Hugon, J.; Wakita, T.; Meurs, E.F. Hepatitis C virus reveals a novel early control in acute immune response. PLoS Pathog. 2011, 7, e1002289. [Google Scholar] [CrossRef]
- Pène, V.; Li, Q.; Sodroski, C.; Hsu, C.-S.; Liang, T.J. Dynamic Interaction of Stress Granules, DDX3X, and IKK-α Mediates Multiple Functions in Hepatitis C Virus Infection. J. Virol. 2015, 89, 5462–5477. [Google Scholar] [CrossRef]
- Binder, M.; Eberle, F.; Seitz, S.; Mücke, N.; Hüber, C.M.; Kiani, N.; Kaderali, L.; Lohmann, V.; Dalpke, A.; Bartenschlager, R. Molecular Mechanism of Signal Perception and Integration by the Innate Immune Sensor Retinoic Acid-inducible Gene-I (RIG-I). J. Biol. Chem. 2011, 286, 27278–27287. [Google Scholar] [CrossRef]
- Li, K.; Li, N.L.; Wei, D.; Pfeffer, S.R.; Fan, M.; Pfeffer, L.M. Activation of chemokine and inflammatory cytokine response in hepatitis C virus-infected hepatocytes depends on Toll-like receptor 3 sensing of hepatitis C virus double-stranded RNA intermediates. Hepatology 2012, 55, 666–675. [Google Scholar] [CrossRef]
- Xie, Q.; Chen, S.; Tian, R.; Huang, X.; Deng, R.; Xue, B.; Qin, Y.; Xu, Y.; Wang, J.; Guo, M.; et al. Long Noncoding RNA ITPRIP-1 Positively Regulates the Innate Immune Response through Promotion of Oligomerization and Activation of MDA5. J. Virol. 2018, 92. [Google Scholar] [CrossRef]
- Ivashkiv, L.B.; Donlin, L.T. Regulation of type I interferon responses. Nat. Rev. Immunol. 2014, 14, 36–49. [Google Scholar] [CrossRef]
- Schneider, W.M.; Chevillotte, M.D.; Rice, C.M. Interferon-stimulated genes: A complex web of host defenses. Annu. Rev. Immunol. 2014, 32, 513–545. [Google Scholar] [CrossRef]
- Barriocanal, M.; Fortes, P. Long Non-coding RNAs in hepatitis C virus-infected cells. Front. Microbiol. 2017, 8, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Carnero, E.; Fortes, P. HCV infection, IFN response and the coding and non-coding host cell genome. Virus Res. 2016, 212, 85–102. [Google Scholar] [CrossRef]
- Malakhova, O.A.; Kim, K., II; Luo, J.-K.; Zou, W.; Kumar, K.G.S.; Fuchs, S.Y.; Shuai, K.; Zhang, D.-E. UBP43 is a novel regulator of interferon signaling independent of its ISG15 isopeptidase activity. EMBO J. 2006, 25, 2358–2367. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, A.; Naka, T.; Kubo, M. SOCS proteins, cytokine signalling and immune regulation. Nat. Rev. Immunol. 2007, 7, 454–465. [Google Scholar] [CrossRef] [PubMed]
- Duong, F.H.T.; Filipowicz, M.; Tripodi, M.; La Monica, N.; Heim, M.H. Hepatitis C virus inhibits interferon signaling through up-regulation of protein phosphatase 2A. Gastroenterology 2004, 126, 263–277. [Google Scholar] [CrossRef] [PubMed]
- Kawaguchi, T.; Yoshida, T.; Harada, M.; Hisamoto, T.; Nagao, Y.; Ide, T.; Taniguchi, E.; Kumemura, H.; Hanada, S.; Maeyama, M.; et al. Hepatitis C virus down-regulates insulin receptor substrates 1 and 2 through up-regulation of suppressor of cytokine signaling 3. Am. J. Pathol. 2004, 165, 1499–1508. [Google Scholar] [CrossRef]
- Walsh, M.J.; Jonsson, J.R.; Richardson, M.M.; Lipka, G.M.; Purdie, D.M.; Clouston, A.D.; Powell, E.E. Non-response to antiviral therapy is associated with obesity and increased hepatic expression of suppressor of cytokine signalling 3 (SOCS-3) in patients with chronic hepatitis C, viral genotype 1. Gut 2006, 55, 529–535. [Google Scholar] [CrossRef]
- Li, K.; Foy, E.; Ferreon, J.C.; Nakamura, M.; Ferreon, A.C.M.; Ikeda, M.; Ray, S.C.; Gale, M.; Lemon, S.M. Immune evasion by hepatitis C virus NS3/4A protease-mediated cleavage of the Toll-like receptor 3 adaptor protein TRIF. Proc. Natl. Acad. Sci. USA 2005, 102, 2992–2997. [Google Scholar] [CrossRef]
- Meylan, E.; Curran, J.; Hofmann, K.; Moradpour, D.; Binder, M.; Bartenschlager, R.; Tschopp, J. Cardif is an adaptor protein in the RIG-I antiviral pathway and is targeted by hepatitis C virus. Nature 2005, 437, 1167–1172. [Google Scholar] [CrossRef] [PubMed]
- Ke, P.-Y.; Chen, S.S.-L. Activation of the unfolded protein response and autophagy after hepatitis C virus infection suppresses innate antiviral immunity in vitro. J. Clin. Invest. 2011, 121, 37–56. [Google Scholar] [CrossRef] [PubMed]
- Hedegaard, D.L.; Tully, D.C.; Rowe, I.A.; Reynolds, G.M.; Bean, D.J.; Hu, K.; Davis, C.; Wilhelm, A.; Ogilvie, C.B.; Power, K.A.; et al. High resolution sequencing of hepatitis C virus reveals limited intra-hepatic compartmentalization in end-stage liver disease. J. Hepatol. 2017, 66, 28–38. [Google Scholar] [CrossRef] [PubMed]
- Garaigorta, U.; Chisari, F. V Hepatitis C virus blocks interferon effector function by inducing protein kinase R phosphorylation. Cell Host Microbe 2009, 6, 513–522. [Google Scholar] [CrossRef] [PubMed]
- Broering, R.; Zhang, X.; Kottilil, S.; Trippler, M.; Jiang, M.; Lu, M.; Gerken, G.; Schlaak, J.F. The interferon stimulated gene 15 functions as a proviral factor for the hepatitis C virus and as a regulator of the IFN response. Gut 2010, 59, 1111–1119. [Google Scholar] [CrossRef]
- Li, Q.; Pène, V.; Krishnamurthy, S.; Cha, H.; Liang, T.J. Hepatitis C virus infection activates an innate pathway involving IKK-α in lipogenesis and viral assembly. Nat. Med. 2013, 19, 722–729. [Google Scholar] [CrossRef]
- Durfee, L.A.; Lyon, N.; Seo, K.; Huibregtse, J.M. The ISG15 conjugation system broadly targets newly synthesized proteins: Implications for the antiviral function of ISG15. Mol. Cell 2010, 38, 722–732. [Google Scholar] [CrossRef]
- Kim, M.-J.; Hwang, S.-Y.; Imaizumi, T.; Yoo, J.-Y. Negative feedback regulation of RIG-I-mediated antiviral signaling by interferon-induced ISG15 conjugation. J. Virol. 2008, 82, 1474–1483. [Google Scholar] [CrossRef]
- Fortes, P.; Morris, K.V. Long noncoding RNAs in viral infections. Virus Res. 2016, 212, 1–11. [Google Scholar] [CrossRef]
- Ransohoff, J.D.; Wei, Y.; Khavari, P.A. The functions and unique features of long intergenic non-coding RNA. Nat. Rev. Mol. Cell Biol. 2018, 19, 143–157. [Google Scholar] [CrossRef]
- Cech, T.R.; Steitz, J.A. The noncoding RNA revolution - Trashing old rules to forge new ones. Cell 2014, 157, 77–94. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, H.; Tomari, Y. RISC assembly: Coordination between small RNAs and Argonaute proteins. Biochim. Biophys. Acta - Gene Regul. Mech. 2016, 1859, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Sarnow, P.; Sagan, S.M. Unraveling the Mysterious Interactions Between Hepatitis C Virus RNA and Liver-Specific MicroRNA-122. Annu. Rev. Virol. 2016, 3, 309–332. [Google Scholar] [CrossRef] [PubMed]
- Sedano, C.D.; Sarnow, P. Interaction of host cell microRNAs with the HCV RNA genome during infection of liver cells. Semin. Liver Dis. 2015, 35, 75–80. [Google Scholar] [CrossRef]
- Iyer, M.K.; Niknafs, Y.S.; Malik, R.; Singhal, U.; Sahu, A.; Hosono, Y.; Barrette, T.R.; Prensner, J.R.; Evans, J.R.; Zhao, S.; et al. The landscape of long noncoding RNAs in the human transcriptome. Nat. Genet. 2015, 47, 199–208. [Google Scholar] [CrossRef]
- Necsulea, A.; Soumillon, M.; Warnefors, M.; Liechti, A.; Daish, T.; Zeller, U.; Baker, J.C.; Grutzner, F.; Kaessmann, H. The evolution of lncRNA repertoires and expression patterns in tetrapods. Nature 2014, 505, 635–640. [Google Scholar] [CrossRef]
- Quinn, J.J.; Chang, H.Y. Unique features of long non-coding RNA biogenesis and function. Nat. Rev. Genet. 2015, 17, 47–62. [Google Scholar] [CrossRef]
- Beckedorff, F.C.; Ayupe, A.C.; Verjovski-Almeida, S.; Tahira, A.C.; Reis, E.M.; Camargo, L. Global analysis of biogenesis, stability and sub-cellular localization of lncRNAs mapping to intragenic regions of the human genome. RNA Biol. 2015, 12, 877–892. [Google Scholar]
- Ulitsky, I. Evolution to the rescue: Using comparative genomics to understand long non-coding RNAs. Nat. Rev. Genet. 2016, 17, 601–614. [Google Scholar] [CrossRef]
- Laurent, G.S.; Wahlestedt, C.; Kapranov, P. The Landscape of long noncoding RNA classification. Trends Genet. 2015, 31, 239–251. [Google Scholar] [CrossRef]
- Kopp, F.; Mendell, J.T. Functional Classification and Experimental Dissection of Long Noncoding RNAs. Cell 2018, 172, 393–407. [Google Scholar] [CrossRef] [PubMed]
- Batista, P.J.; Chang, H.Y. Long noncoding RNAs: Cellular address codes in development and disease. Cell 2013, 152, 1298–1307. [Google Scholar] [CrossRef] [PubMed]
- Flynn, R.A.; Chang, H.Y. Long noncoding RNAs in cell-fate programming and reprogramming. Cell Stem Cell 2014, 14, 752–761. [Google Scholar] [CrossRef] [PubMed]
- Unfried, J.P.; Serrano, G.; Suárez, B.; Sangro, P.; Ferretti, V.; Prior, C.; Boix, L.; Bruix, J.; Sangro, B.; Segura, V.; et al. Identification of Coding and Long Noncoding RNAs Differentially Expressed in Tumors and Preferentially Expressed in Healthy Tissues. Cancer Res. 2019, 79, 5167–5180. [Google Scholar] [CrossRef] [PubMed]
- Gutschner, T.; Diederichs, S. The Hallmarks of Cancer: A long non-coding RNA point of view. RNA Biol. 2012, 9, 703–719. [Google Scholar] [CrossRef]
- Tseng, Y.-Y.; Moriarity, B.S.; Gong, W.; Akiyama, R.; Tiwari, A.; Kawakami, H.; Ronning, P.; Reuland, B.; Guenther, K.; Beadnell, T.C.; et al. PVT1 dependence in cancer with MYC copy-number increase. Nature 2014, 512, 82–86. [Google Scholar] [CrossRef]
- Gupta, R.A.; Shah, N.; Wang, K.C.; Kim, J.; Horlings, H.M.; Wong, D.J.; Tsai, M.-C.; Hung, T.; Argani, P.; Rinn, J.L.; et al. Long non-coding RNA HOTAIR reprograms chromatin state to promote cancer metastasis. Nature 2010, 464, 1071–1076. [Google Scholar] [CrossRef]
- Braconi, C.; Kogure, T.; Valeri, N.; Huang, N.; Nuovo, G.; Costinean, S.; Negrini, M.; Miotto, E.; Croce, C.M.; Patel, T. microRNA-29 can regulate expression of the long non-coding RNA gene MEG3 in hepatocellular cancer. Oncogene 2011, 30, 4750–4756. [Google Scholar] [CrossRef]
- Han, L.; Zhang, E.; Yin, D.; Kong, R.; Xu, T.; Chen, W.; Xia, R.; Shu, Y.; De, W. Low expression of long noncoding RNA PANDAR predicts a poor prognosis of non-small cell lung cancer and affects cell apoptosis by regulating Bcl-2. Cell Death Dis. 2015, 6, e1665. [Google Scholar] [CrossRef]
- Cabanski, C.R.; White, N.M.; Dang, H.X.; Silva-Fisher, J.M.; Rauck, C.E.; Cicka, D.; Maher, C.A. Pan-cancer transcriptome analysis reveals long noncoding RNAs with conserved function. RNA Biol. 2015, 12, 628–642. [Google Scholar] [CrossRef]
- Chiu, H.-S.; Somvanshi, S.; Patel, E.; Chen, T.-W.; Singh, V.P.; Zorman, B.; Patil, S.L.; Pan, Y.; Chatterjee, S.S.; Cancer Genome Atlas Research Network, S.J.; et al. Pan-Cancer Analysis of lncRNA Regulation Supports Their Targeting of Cancer Genes in Each Tumor Context. Cell Rep. 2018, 23, 297–312.e12. [Google Scholar] [CrossRef] [PubMed]
- Kamel, M.M.; Matboli, M.; Sallam, M.; Montasser, I.F.; Saad, A.S.; El-Tawdi, A.H.F. Investigation of long noncoding RNAs expression profile as potential serum biomarkers in patients with hepatocellular carcinoma. Transl. Res. 2016, 168, 134–145. [Google Scholar] [CrossRef] [PubMed]
- Yang, N.; Li, S.; Li, G.; Zhang, S.; Tang, X.; Ni, S.; Jian, X.; Xu, C.; Zhu, J.; Lu, M. The role of extracellular vesicles in mediating progression, metastasis and potential treatment of hepatocellular carcinoma. Oncotarget 2017, 8, 3683–3695. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.-X.; Zhang, X.; Miao, R.-C.; Xiang, X.-H.; Fu, Y.-N.; Zhang, J.-Y.; Liu, C.; Qu, K. Six-long non-coding RNA signature predicts recurrence-free survival in hepatocellular carcinoma. World J. Gastroenterol. 2019, 25, 220–232. [Google Scholar] [CrossRef] [PubMed]
- Sui, J.; Miao, Y.; Han, J.; Nan, H.; Shen, B.; Zhang, X.; Zhang, Y.; Wu, Y.; Wu, W.; Liu, T.; et al. Systematic analyses of a novel lncRNA-associated signature as the prognostic biomarker for Hepatocellular Carcinoma. Cancer Med. 2018, 7, 3240–3256. [Google Scholar] [CrossRef]
- Tang, J.; Jiang, R.; Deng, L.; Zhang, X.; Wang, K.; Sun, B. Circulation long non-coding RNAs act as biomarkers for predicting tumorigenesis and metastasis in hepatocellular carcinoma. Oncotarget 2015, 6, 4505. [Google Scholar] [CrossRef]
- Carnero, E.; Barriocanal, M.; Prior, C.; Pablo Unfried, J.; Segura, V.; Guruceaga, E.; Enguita, M.; Smerdou, C.; Gastaminza, P.; Fortes, P. Long noncoding RNA EGOT negatively affects the antiviral response and favors HCV replication. EMBO Rep. 2016, 17, 1013–1028. [Google Scholar] [CrossRef]
- Moon, S.L.; Blackinton, J.G.; Anderson, J.R.; Dozier, M.K.; Dodd, B.J.T.; Keene, J.D.; Wilusz, C.J.; Bradrick, S.S.; Wilusz, J. XRN1 stalling in the 5’ UTR of Hepatitis C virus and Bovine Viral Diarrhea virus is associated with dysregulated host mRNA stability. PLoS Pathog. 2015, 11, e1004708. [Google Scholar] [CrossRef]
- Xu, C.; Zhang, Y.; Wang, Q.; Xu, Z.; Jiang, J.; Gao, Y.; Gao, M.; Kang, J.; Wu, M.; Xiong, J.; et al. Long non-coding RNA GAS5 controls human embryonic stem cell self-renewal by maintaining NODAL signalling. Nat. Commun. 2016, 7, 13287. [Google Scholar] [CrossRef]
- Hudson, W.H.; Pickard, M.R.; de Vera, I.M.S.; Kuiper, E.G.; Mourtada-Maarabouni, M.; Conn, G.L.; Kojetin, D.J.; Williams, G.T.; Ortlund, E.A. Conserved sequence-specific lincRNA-steroid receptor interactions drive transcriptional repression and direct cell fate. Nat. Commun. 2014, 5, 5395. [Google Scholar] [CrossRef]
- Qian, X.; Xu, C.; Zhao, P.; Qi, Z. Long non-coding RNA GAS5 inhibited hepatitis C virus replication by binding viral NS3 protein. Virology 2016, 492, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Murray, C.L.; Jones, C.T.; Rice, C.M. Architects of assembly: Roles of Flaviviridae non-structural proteins in virion morphogenesis. Nat. Rev. Microbiol. 2008, 6, 699–708. [Google Scholar] [CrossRef] [PubMed]
- Moradpour, D.; Penin, F. Hepatitis C virus proteins: From structure to function. Curr. Top. Microbiol. Immunol. 2013, 369, 113–142. [Google Scholar] [PubMed]
- Li, Z.-Q.; Gu, X.-Y.; Hu, J.-X.; Ping, Y.; Li, H.; Yan, J.-Y.; Li, J.; Sun, R.; Yu, Z.-J.; Zhang, Y. Hepatitis C virus core protein impairs metabolic disorder of liver cell via HOTAIR-Sirt1 signalling. Biosci. Rep. 2016, 36, e00336. [Google Scholar] [CrossRef]
- Sharma, G.; Tripathi, S.K.; Das, S. lncRNA HULC facilitates efficient loading of HCV-core protein onto lipid droplets and subsequent virus-particle release. Cell. Microbiol. 2019, 21, e13086. [Google Scholar] [CrossRef]
- Xiong, Y.; Jia, M.; Yuan, J.; Zhang, C.; Zhu, Y.; Kuang, X.; Lan, L.; Wang, X. STAT3-regulated long non-coding RNAs lnc-7SK and lnc-IGF2-AS promote hepatitis C virus replication. Mol. Med. Rep. 2015, 12, 6738–6744. [Google Scholar] [CrossRef]
- Fan, J.; Cheng, M.; Chi, X.; Liu, X.; Yang, W. A Human Long Non-coding RNA LncATV Promotes Virus Replication Through Restricting RIG-I-Mediated Innate Immunity. Front. Immunol. 2019, 10, 1711. [Google Scholar] [CrossRef]
- Liu, X.; Duan, X.; Holmes, J.A.; Li, W.; Lee, S.H.; Tu, Z.; Zhu, C.; Salloum, S.; Lidofsky, A.; Schaefer, E.A.; et al. A Long Noncoding RNA Regulates Hepatitis C Virus Infection Through Interferon Alpha-Inducible Protein 6. Hepatology 2019, 69, 1004–1019. [Google Scholar] [CrossRef]
- Barriocanal, M.; Carnero, E.; Segura, V.; Fortes, P. Long Non-Coding RNA BST2/BISPR is Induced by IFN and Regulates the Expression of the Antiviral Factor Tetherin. Front. Immunol. 2014, 5, 655. [Google Scholar] [CrossRef]
- Kambara, H.; Gunawardane, L.; Zebrowski, E.; Kostadinova, L.; Jobava, R.; Krokowski, D.; Hatzoglou, M.; Anthony, D.D.; Valadkhan, S. Regulation of Interferon-Stimulated Gene BST2 by a lncRNA Transcribed from a Shared Bidirectional Promoter. Front. Immunol. 2014, 5, 676. [Google Scholar] [CrossRef]
- Nishitsuji, H.; Ujino, S.; Yoshio, S.; Sugiyama, M.; Mizokami, M.; Kanto, T.; Shimotohno, K. Long noncoding RNA #32 contributes to antiviral responses by controlling interferon-stimulated gene expression. Proc. Natl. Acad. Sci. USA 2016, 113, 10388–10393. [Google Scholar] [PubMed]
- Kambara, H.; Niazi, F.; Kostadinova, L.; Moonka, D.K.; Siegel, C.T.; Post, A.B.; Carnero, E.; Barriocanal, M.; Fortes, P.; Anthony, D.D.; et al. Negative regulation of the interferon response by an interferon-induced long non-coding RNA. Nucleic Acids Res. 2014, 42, 10668–10680. [Google Scholar] [CrossRef] [PubMed]
- Hu, P.; Wilhelm, J.; Gerresheim, G.K.; Shalamova, L.A.; Niepmann, M. Lnc-ITM2C-1 and GPR55 Are Proviral Host Factors for Hepatitis C Virus. Viruses 2019, 11, 549. [Google Scholar] [CrossRef] [PubMed]
- Bishé, B.; Syed, G.H.; Field, S.J.; Siddiqui, A. Role of phosphatidylinositol 4-phosphate (PI4P) and its binding protein GOLPH3 in hepatitis C virus secretion. J. Biol. Chem. 2012, 287, 27637–27647. [Google Scholar] [CrossRef]
- Wang, Q.; Zhang, D.; Feng, W.; Guo, Y.; Sun, X.; Zhang, M.; Guan, Z.; Duan, M. Long noncoding RNA TSPOAP1 antisense RNA 1 negatively modulates type I IFN signaling to facilitate influenza A virus replication. J. Med. Virol. 2019. [Google Scholar] [CrossRef]
- Maarouf, M.; Chen, B.; Chen, Y.; Wang, X.; Rai, K.R.; Zhao, Z.; Liu, S.; Li, Y.; Xiao, M.; Chen, J.-L. Identification of lncRNA-155 encoded by MIR155HG as a novel regulator of innate immunity against influenza A virus infection. Cell. Microbiol. 2019, 21, e13036. [Google Scholar] [CrossRef]
- Zhou, Y.; Li, M.; Xue, Y.; Li, Z.; Wen, W.; Liu, X.; Ma, Y.; Zhang, L.; Shen, Z.; Cao, X. Interferon-inducible cytoplasmic lncLrrc55-AS promotes antiviral innate responses by strengthening IRF3 phosphorylation. Cell Res. 2019, 29, 641–654. [Google Scholar] [CrossRef]
- Ma, H.; Han, P.; Ye, W.; Chen, H.; Zheng, X.; Cheng, L.; Zhang, L.; Yu, L.; Wu, X.; Xu, Z.; et al. The Long Noncoding RNA NEAT1 Exerts Antihantaviral Effects by Acting as Positive Feedback for RIG-I Signaling. J. Virol. 2017, 91. [Google Scholar] [CrossRef]
- Jiang, M.; Zhang, S.; Yang, Z.; Lin, H.; Zhu, J.; Liu, L.; Wang, W.; Liu, S.; Liu, W.; Ma, Y.; et al. Self-Recognition of an Inducible Host lncRNA by RIG-I Feedback Restricts Innate Immune Response. Cell 2018, 173, 906–919.e13. [Google Scholar] [CrossRef]
- Li, X.; Guo, G.; Lu, M.; Chai, W.; Li, Y.; Tong, X.; Li, J.; Jia, X.; Liu, W.; Qi, D.; et al. Long Noncoding RNA Lnc-MxA Inhibits Beta Interferon Transcription by Forming RNA-DNA Triplexes at Its Promoter. J. Virol. 2019, 93. [Google Scholar] [CrossRef]
- Carnero, E.; Barriocanal, M.; Segura, V.; Guruceaga, E.; Prior, C.; Börner, K.; Grimm, D.; Fortes, P. Type I Interferon Regulates the Expression of Long Non-Coding RNAs. Front. Immunol. 2014, 5, 548. [Google Scholar] [CrossRef] [PubMed]
- Meyer, K.; Kwon, Y.-C.; Liu, S.; Hagedorn, C.H.; Ray, R.B.; Ray, R. Interferon-α inducible protein 6 impairs EGFR activation by CD81 and inhibits hepatitis C virus infection. Sci. Rep. 2015, 5, 9012. [Google Scholar] [CrossRef] [PubMed]
- Neil, S.J.D.; Zang, T.; Bieniasz, P.D. Tetherin inhibits retrovirus release and is antagonized by HIV-1 Vpu. Nature 2008, 451, 425–430. [Google Scholar] [CrossRef] [PubMed]
- Dafa-Berger, A.; Kuzmina, A.; Fassler, M.; Yitzhak-Asraf, H.; Shemer-Avni, Y.; Taube, R. Modulation of hepatitis C virus release by the interferon-induced protein BST-2/tetherin. Virology 2012, 428, 98–111. [Google Scholar] [CrossRef] [PubMed]
- Watson, G.; Ronai, Z.A.; Lau, E. ATF2, a paradigm of the multifaceted regulation of transcription factors in biology and disease. Pharmacol. Res. 2017, 119, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Reimold, A.M.; Kim, J.; Finberg, R.; Glimcher, L.H. Decreased immediate inflammatory gene induction in activating transcription factor-2 mutant mice. Int. Immunol. 2001, 13, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Helbig, K.J.; Lau, D.T.-Y.; Semendric, L.; Harley, H.A.J.; Beard, M.R. Analysis of ISG expression in chronic hepatitis C identifies viperin as a potential antiviral effector. Hepatology 2005, 42, 702–710. [Google Scholar] [CrossRef]
- Helbig, K.J.; Eyre, N.S.; Yip, E.; Narayana, S.; Li, K.; Fiches, G.; McCartney, E.M.; Jangra, R.K.; Lemon, S.M.; Beard, M.R. The antiviral protein viperin inhibits hepatitis C virus replication via interaction with nonstructural protein 5A. Hepatology 2011, 54, 1506–1517. [Google Scholar] [CrossRef]
- Patsenker, E.; Sachse, P.; Chicca, A.; Gachet, M.S.; Schneider, V.; Mattsson, J.; Lanz, C.; Worni, M.; de Gottardi, A.; Semmo, M.; et al. Elevated levels of endocannabinoids in chronic hepatitis C may modulate cellular immune response and hepatic stellate cell activation. Int. J. Mol. Sci. 2015, 16, 7057–7076. [Google Scholar] [CrossRef]
- Mallat, A.; Teixeira-Clerc, F.; Lotersztajn, S. Cannabinoid signaling and liver therapeutics. J. Hepatol. 2013, 59, 891–896. [Google Scholar] [CrossRef]
- Rose, D.; Stadler, P.F. Molecular evolution of the non-coding eosinophil granule ontogeny transcript. Front. Genet. 2011, 2, 69. [Google Scholar] [CrossRef] [PubMed]
- Wagner, L.A.; Christensen, C.J.; Dunn, D.M.; Spangrude, G.J.; Georgelas, A.; Kelley, L.; Esplin, M.S.; Weiss, R.B.; Gleich, G.J. EGO, a novel, noncoding RNA gene, regulates eosinophil granule protein transcript expression. Blood 2007, 109, 5191–5198. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.; Li, C.; Kang, B.; Gao, G.; Li, C.; Zhang, Z. GEPIA: A web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res. 2017, 45, W98–W102. [Google Scholar] [CrossRef] [PubMed]
- Younossi, Z.; Park, H.; Henry, L.; Adeyemi, A.; Stepanova, M. Extrahepatic Manifestations of Hepatitis C: A Meta-analysis of Prevalence, Quality of Life, and Economic Burden. Gastroenterology 2016, 150, 1599–1608. [Google Scholar] [CrossRef] [PubMed]
- Sabry, D.; Elamir, A.; Mahmoud, R.H.; Abdelaziz, A.A.; Fathy, W. Role of LncRNA-AF085935, IL-10 and IL-17 in Rheumatoid Arthritis Patients With Chronic Hepatitis C. J. Clin. Med. Res. 2017, 9, 416–425. [Google Scholar] [CrossRef] [PubMed]
- Cui, M.; Chang, Y.; Du, W.; Liu, S.; Qi, J.; Luo, R.; Luo, S. Upregulation of lncRNA-ATB by Transforming Growth Factor β1 (TGF-β1) Promotes Migration and Invasion of Papillary Thyroid Carcinoma Cells. Med. Sci. Monit. 2018, 24, 5152–5158. [Google Scholar] [CrossRef] [PubMed]
- Fu, N.; Niu, X.; Wang, Y.; Du, H.; Wang, B.; Du, J.; Li, Y.; Wang, R.; Zhang, Y.; Zhao, S.; et al. Role of LncRNA-activated by transforming growth factor beta in the progression of hepatitis C virus-related liver fibrosis. Discov. Med. 2016, 22, 29–42. [Google Scholar]
- Manns, M.P.; Buti, M.; Gane, E.; Pawlotsky, J.M.; Razavi, H.; Terrault, N.; Younossi, Z. Hepatitis C virus infection. Nat. Rev. Dis. Prim. 2017, 3, 1–19. [Google Scholar] [CrossRef]
- Arzumanyan, A.; Reis, H.M.G.P.V.; Feitelson, M.A. Pathogenic mechanisms in HBV- and HCV-associated hepatocellular carcinoma. Nat. Rev. Cancer 2013, 13, 123–135. [Google Scholar] [CrossRef]
- Vescovo, T.; Refolo, G.; Vitagliano, G.; Fimia, G.M.; Piacentini, M. Molecular mechanisms of hepatitis C virus–induced hepatocellular carcinoma. Clin. Microbiol. Infect. 2016, 22, 853–861. [Google Scholar] [CrossRef]
- Ukwaja, K.N.; Malekzadeh, R.; Dey, S.; Chitheer, A.; Wakayo, T.; Kumar, G.A.; Abera, S.; Pourmalek, F.; Fürst, T.; Ekwueme, D.U.; et al. The Burden of Primary Liver Cancer and Underlying Etiologies From 1990 to 2015 at the Global, Regional, and National Level. JAMA Oncol. 2017, 3, 1683. [Google Scholar]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2019. CA. Cancer J. Clin. 2019, 69, 7–34. [Google Scholar] [CrossRef] [PubMed]
- Llovet, J.M.; Zucman-Rossi, J.; Pikarsky, E.; Sangro, B.; Schwartz, M.; Sherman, M.; Gores, G. Hepatocellular carcinoma. Nat. Rev. Dis. Prim. 2016, 2, 16018. [Google Scholar] [CrossRef] [PubMed]
- Zucman-Rossi, J.; Villanueva, A.; Nault, J.-C.; Llovet, J.M. Genetic Landscape and Biomarkers of Hepatocellular Carcinoma. Gastroenterology 2015, 149, 1226–1239.e4. [Google Scholar] [CrossRef] [PubMed]
- Tsang, F.H.C.; Au, S.L.K.; Wei, L.; Fan, D.N.Y.; Lee, J.M.F.; Wong, C.C.L.; Ng, I.O.L.; Wong, C.M. Long non-coding RNA HOTTIP is frequently up-regulated in hepatocellular carcinoma and is targeted by tumour suppressive miR-125b. Liver Int. 2015, 35, 1597–1606. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.-M.; Tsang, F.H.-C.; Ng, I.O.-L. Non-coding RNAs in hepatocellular carcinoma: Molecular functions and pathological implications. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 137–151. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Matsuura, K.; Kleiner, D.E.; Zamboni, F.; Alter, H.J.; Farci, P. Analysis of long noncoding RNA expression in hepatocellular carcinoma of different viral etiology. J. Transl. Med. 2016, 14, 328. [Google Scholar] [CrossRef] [PubMed]
- Fang, T.-T.; Sun, X.-J.; Chen, J.; Zhao, Y.; Sun, R.-X.; Ren, N.; Liu, B.-B. Long non-coding RNAs are differentially expressed in hepatocellular carcinoma cell lines with differing metastatic potential. Asian Pac. J. Cancer Prev. 2014, 15, 10513–10524. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zhang, H.; Zhu, C.; Zhao, Y.; Li, M.; Wu, L.; Yang, X.; Wan, X.; Wang, A.; Zhang, M.Q.; Sang, X.; et al. Long non-coding RNA expression profiles of hepatitis C virus-related dysplasia and hepatocellular carcinoma. Oncotarget 2015, 6, 43770–43778. [Google Scholar] [CrossRef]
- Deng, B.; Xu, W.; Wang, Z.; Liu, C.; Lin, P.; Li, B.; Huang, Q.; Yang, J.; Zhou, H.; Qu, L. An LTR retrotransposon-derived lnc RNA interacts with RNF 169 to promote homologous recombination. EMBO Rep. 2019, 20, 1–18. [Google Scholar] [CrossRef]
- Li, Y.C.; Wang, D.; Zhu, G.Y. Increased expression of long noncoding RNA AK021443 predicts worse clinical outcome in hepatocellular carcinoma. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 4855–4860. [Google Scholar] [PubMed]
- Petermann, F.; Pękowska, A.; Johnson, C.A.; Jankovic, D.; Shih, H.Y.; Jiang, K.; Hudson, W.H.; Brooks, S.R.; Sun, H.W.; Villarino, A.V.; et al. The Magnitude of IFN-γ Responses Is Fine-Tuned by DNA Architecture and the Non-coding Transcript of Ifng-as1. Mol. Cell 2019, 75, 1229–1242.e5. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Li, P.; Kwok, J.G.; Korrapati, A.; Li, W.T.; Qu, Y.; Wang, X.Q.; Kisseleva, T.; Wang-Rodriguez, J.; Ongkeko, W.M. Alcohol and hepatitis virus-dysregulated lncRNAs as potential biomarkers for hepatocellular carcinoma. Oncotarget 2018, 9, 224–235. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Mo, M.; Tan, H.J.J.; Tan, C.; Zeng, X.; Zhang, G.; Huang, D.; Liang, J.; Liu, S.; Qiu, X. LINC02499, a novel liver-specific long non-coding RNA with potential diagnostic and prognostic value, inhibits hepatocellular carcinoma cell proliferation, migration, and invasion. Hepatol. Res. 2020. [Google Scholar] [CrossRef]
- Sonohara, F.; Inokawa, Y.; Hayashi, M.; Yamada, S.; Sugimoto, H.; Fujii, T.; Kodera, Y.; Nomoto, S. Prognostic Value of Long Non-Coding RNA HULC and MALAT1 Following the Curative Resection of Hepatocellular Carcinoma. Sci. Rep. 2017, 7, 1–9. [Google Scholar] [CrossRef]
- Li, S.P.; Xu, H.X.; Yu, Y.; He, J.D.; Wang, Z.; Xu, Y.J.; Wang, C.Y.; Zhang, H.M.; Zhang, R.X.; Zhang, J.J.; et al. LncRNA HULC enhances epithelial-mesenchymal transition to promote tumorigenesis and metastasis of hepatocellular carcinoma via the miR-200a-3p/ZEB1 signaling pathway. Oncotarget 2016, 7, 42431–42446. [Google Scholar] [CrossRef]
- Toraih, E.A.; Ellawindy, A.; Fala, S.Y.; Al Ageeli, E.; Gouda, N.S.; Fawzy, M.S.; Hosny, S. Oncogenic long noncoding RNA MALAT1 and HCV-related hepatocellular carcinoma. Biomed. Pharmacother. 2018, 102, 653–669. [Google Scholar] [CrossRef]
- Malakar, P.; Shilo, A.; Mogilevsky, A.; Stein, I.; Pikarsky, E.; Nevo, Y.; Benyamini, H.; Elgavish, S.; Zong, X.; Prasanth, K.V.; et al. Long noncoding RNA MALAT1 promotes hepatocellular carcinoma development by SRSF1 upregulation and mTOR activation. Cancer Res. 2017, 77, 1155–1167. [Google Scholar] [CrossRef]
- Wang, F.; Ying, H.-Q.; He, B.-S.; Pan, Y.-Q.; Deng, Q.-W.; Sun, H.-L.; Chen, J.; Liu, X.; Wang, S.-K. Upregulated lncRNA-UCA1 contributes to progression of hepatocellular carcinoma through inhibition of miR-216b and activation of FGFR1/ERK signaling pathway. Oncotarget 2015, 6, 7899–7917. [Google Scholar] [CrossRef]
- Mahmoudi, S.; Henriksson, S.; Farnebo, L.; Roberg, K.; Farnebo, M. WRAP53 promotes cancer cell survival and is a potential target for cancer therapy. Cell Death Dis. 2011, 2, 1–10. [Google Scholar] [CrossRef]
- Zhang, C.; Yang, X.; Qi, Q.; Gao, Y.; Wei, Q.; Han, S. lncRNA-HEIH in serum and exosomes as a potential biomarker in the HCV-related hepatocellular carcinoma. Cancer Biomark. 2018, 21, 651–659. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Zhang, L.; Huo, X.S.; Yuan, J.H.; Xu, D.; Yuan, S.X.; Zhu, N.; Zhou, W.P.; Yang, G.S.; Wang, Y.Z.; et al. Long noncoding RNA high expression in hepatocellular carcinoma facilitates tumor growth through enhancer of zeste homolog 2 in humans. Hepatology 2011, 54, 1679–1689. [Google Scholar] [CrossRef] [PubMed]
- Refai, N.S.; Louka, M.L.; Halim, H.Y.; Montasser, I. Long non-coding RNAs (CASC2 and TUG1) in hepatocellular carcinoma: Clinical significance. J. Gene Med. 2019, 21, e3112. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, Z.; Yao, B.; Li, Q.; Wang, L.; Wang, C.; Dou, C.; Xu, M.; Liu, Q.; Tu, K. Long non-coding RNA CASC2 suppresses epithelial-mesenchymal transition of hepatocellular carcinoma cells through CASC2/miR-367/FBXW7 axis. Mol. Cancer 2017, 16, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.D.; Chen, W.M.; Qi, F.Z.; Sun, M.; Xu, T.P.; Ma, P.; Shu, Y.Q. Long non-coding RNA TUG1 is up-regulated in hepatocellular carcinoma and promotes cell growth and apoptosis by epigenetically silencing of KLF2. Mol. Cancer 2015, 14, 1–12. [Google Scholar] [CrossRef]
- Roshdy, F.; Farag, M.M.S.; El-Ahwany, E.; Mahmode, O.; Mousa, A.A.; Talkawy, M.E.; Essawy, F. Long non-coding RNA HOTAIR and HOTTIP as potential biomarkers for hepatitis C virus genotype 4-induced hepatocellular carcinoma. Egypt. J. Med Hum. Genet. 2020, 21, 1–13. [Google Scholar] [CrossRef]
- Cheng, D.; Deng, J.; Zhang, B.; He, X.; Meng, Z.; Li, G.; Ye, H.; Zheng, S.; Wei, L.; Deng, X.; et al. LncRNA HOTAIR epigenetically suppresses miR-122 expression in hepatocellular carcinoma via DNA methylation. EBioMedicine 2018, 36, 159–170. [Google Scholar] [CrossRef]
- Li, Z.; Zhao, X.; Zhou, Y.; Liu, Y.; Zhou, Q.; Ye, H.; Wang, Y.X.; Zeng, J.; Song, Y.; Gao, W.; et al. The long non-coding RNA HOTTIP promotes progression and gemcitabine resistance by regulating HOXA13 in pancreatic cancer. J. Transl. Med. 2015, 13, 1–16. [Google Scholar] [CrossRef]
- Gomez, J.A.; Wapinski, O.L.; Yang, Y.W.; Bureau, J.F.; Gopinath, S.; Monack, D.M.; Chang, H.Y.; Brahic, M.; Kirkegaard, K. The NeST long ncRNA controls microbial susceptibility and epigenetic activation of the interferon-γ locus. Cell 2013, 152, 743–754. [Google Scholar] [CrossRef]
- Chen, T.; Pei, J.; Wang, J.; Luo, R.; Liu, L.; Wang, L.; Jia, H. HBx-related long non-coding RNA 01152 promotes cell proliferation and survival by IL-23 in hepatocellular carcinoma. Biomed. Pharmacother. 2019, 115, 0–8. [Google Scholar] [CrossRef]
- Wu, Y.; Li, H.; Chen, R.; Fang, S.; Li, X.; Teng, X.; Zhao, Y.; Zhao, L.; Sun, L.; Sun, X.; et al. NONCODEV5: A comprehensive annotation database for long non-coding RNAs. Nucleic Acids Res. 2017, 46, D308–D314. [Google Scholar]
- Guerriero, P.; Moshiri, F.; Lupini, L.; Sabbioni, S.; Negrini, M.; Callegari, E. Circulating tumor DNAs and non-coding RNAs as potential biomarkers for hepatocellular carcinoma diagnosis, prognosis and response to therapy. Hepatoma Res. 2019, 2019. [Google Scholar] [CrossRef]
- Heitzer, E.; Haque, I.S.; Roberts, C.E.S.; Speicher, M.R. Current and future perspectives of liquid biopsies in genomics-driven oncology. Nat. Rev. Genet. 2019, 20, 71–88. [Google Scholar] [CrossRef] [PubMed]
- Siravegna, G.; Marsoni, S.; Siena, S.; Bardelli, A. Integrating liquid biopsies into the management of cancer. Nat. Rev. Clin. Oncol. 2017, 14, 531–548. [Google Scholar] [CrossRef] [PubMed]
- Zaporozhchenko, I.A.; Ponomaryova, A.A.; Rykova, E.Y.; Laktionov, P.P. The potential of circulating cell-free RNA as a cancer biomarker: Challenges and opportunities. Expert Rev. Mol. Diagn. 2018, 18, 133–145. [Google Scholar] [CrossRef] [PubMed]
- Anfossi, S.; Babayan, A.; Pantel, K.; Calin, G.A. Clinical utility of circulating non-coding RNAs — an update. Nat. Rev. Clin. Oncol. 2018, 15, 541–563. [Google Scholar] [CrossRef]
- Wang, X.; Peng, J.; Wang, J.; Li, M.; Wu, D.; Wu, S.; Liao, J.; Dou, J. Hepatitis C virus core impacts expression of miR122 and miR204 involved in carcinogenic progression via regulation of TGFBRAP1 and HOTTIP expression. Onco. Targets. Ther. 2018, 11, 1173–1182. [Google Scholar] [CrossRef]
- Zhang, S.; Ouyang, X.; Jiang, X.; Gu, D.; Lin, Y.; Kong, S.K.; Xie, W. Dysregulated serum microRNA expression profile and potential biomarkers in hepatitis C virus-infected patients. Int. J. Med. Sci. 2015, 12, 590–598. [Google Scholar] [CrossRef]
- Takaki, Y.; Saito, Y.; Takasugi, A.; Toshimitsu, K.; Yamada, S.; Muramatsu, T.; Kimura, M.; Sugiyama, K.; Suzuki, H.; Arai, E.; et al. Silencing of microRNA-122 is an early event during hepatocarcinogenesis from non-alcoholic steatohepatitis. Cancer Sci. 2014, 105, 1254–1260. [Google Scholar] [CrossRef]
- Wang, W.; Liu, W.B.; Huang, D.B.; Jia, W.; Ji, C.S.; Hu, B. Targeting PCDH20 gene by microRNA-122 confers 5-FU resistance in hepatic carcinoma. Am. J. Cancer Res. 2016, 6, 1681–1694. [Google Scholar]
- Wang, F.; Yuan, J.H.; Wang, S.B.; Yang, F.; Yuan, S.X.; Ye, C.; Yang, N.; Zhou, W.P.; Li, W.L.; Li, W.; et al. Oncofetal long noncoding RNA PVT1 promotes proliferation and stem cell-like property of hepatocellular carcinoma cells by stabilizing NOP2. Hepatology 2014, 60, 1278–1290. [Google Scholar] [CrossRef] [PubMed]
- Ding, C.; Yang, Z.; Lv, Z.; Du, C.; Xiao, H.; Peng, C.; Cheng, S.; Xie, H.; Zhou, L.; Wu, J.; et al. Long non-coding RNA PVT1 is associated with tumor progression and predicts recurrence in hepatocellular carcinoma patients. Oncol. Lett. 2015, 9, 955–963. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.W.; Xu, J.; Sun, R.; Mumbach, M.R.; Carter, A.C.; Chen, Y.G.; Yost, K.E.; Kim, J.; He, J.; Nevins, S.A.; et al. Promoter of lncRNA Gene PVT1 Is a Tumor-Suppressor DNA Boundary Element. Cell 2018, 173, 1398–1412.e22. [Google Scholar] [CrossRef] [PubMed]
- Sur, S.; Sasaki, R.; Devhare, P.; Steele, R.; Ray, R.; Ray, R.B. Association between MicroRNA-373 and Long Noncoding RNA NORAD in Hepatitis C Virus-Infected Hepatocytes Impairs Wee1 Expression for Growth Promotion. J. Virol. 2018, 92, 1–11. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| GENCODE Name | Alternative Name | HCV | Regulation | Impact | Role | Function | References |
|---|---|---|---|---|---|---|---|
| GAS5 | - | Up | 5′end blocks NS3 | Blocks NS3 | Antiviral | [71] | |
| HOTAIR | - | Up | Core | Silences SIRT1 promoter and affects glucose and lipid metabolism | Viral release | Proviral | [74] |
| HULC | - | Up | RXRA | Required for lipid biogenesis. Facilitates binding of core with lipid droplets | Viral release | Proviral | [75] |
| IGF2-AS | - | Up | STAT3 | Increases phosphatidylinositol 4-phosphate (PI4P) | Viral release | Proviral | [76] |
| 7SK | - | Up | STAT3 | Increases phosphatidylinositol 4-phosphate (PI4P) | Viral release | Proviral | [76] |
| AL391832.2 | lncATV | Up | IFN | Binds to RIG-I and inhibits signaling and IFN production | Antiviral response | Proviral | [77] |
| CFAP58-DT | lncITPRIP-1 | Up | IFN | Binds MDA5 and induces oligomerization, activation, and binding to viral RNA | Antiviral response | Antiviral | [20] |
| AL445490.1 | lncRNA-IFI6 | Up | Inhibits expression of neighbor IFI6, by affecting histone modification | Antiviral response | Proviral | [78] | |
| BISPR | - | Up | IFN | Induces BST2 expression | Antiviral response | Antiviral | [79,80] |
| LUARIS | lncRNA-32 | pI:C/down IFN | Binds ATF2 and induces ISG expression | Antiviral response | Antiviral | [81] | |
| NRIR | lnc-CMPK2 | Up | IFN | Negative regulator of several ISGs | Antiviral response | Proviral | [82] |
| GCSIR | lnc-ITM2C-1 | Up | HCV/down IFN | Promotes expression of neighbor GPR55, negative regulator of ISGs | Antiviral response | Proviral | [83] |
| EGOT | - | Up | IFN/NF-κB | Negative regulator of several ISGs | Antiviral response | Proviral | [67] |
| GENCODE Name | Alternative Name | Relative Expression | Function in HCC | References |
|---|---|---|---|---|
| LINC01419 | PRLH1 | up | Promotes cell proliferation by helping HR repair of DNA damage | [119,120] |
| AK021443 | up | Promotes cell cycle progression 1 | [119] | |
| AF070632 | down | Involved in cofactor binding and cell metabolism 1 | [119] | |
| HIF1A-AS2 | aHIF | down | Unknown | [117] |
| PWAR5 | PAR5 | down | Unknown | [117] |
| IFNG-AS1 | TMEVPG1 | down | Activates IFN-γ production and the antiviral response | [117,122] |
| LINC01152 | up/down | Promotes tumorigenesis by binding IL-23 and activating STAT3/pSTAT3 in HBV-related HCC | [117] | |
| LINC02499 | lnc-AFM-2 | down | Overexpression inhibits HCC cells proliferation, migration, and invasion in vitro | [123,124] |
| HULC | up | Enhances tumorigenesis and metastasis via activation of miR-200a-3p/ZEB1 pathway | [125,126] | |
| MALAT1 | up | Acts as a proto-oncogene through Wnt activation and induction of SRSF1 splicing | [127,128] | |
| Biomarkers in Liquid Biopsies | ||||
| MALAT1 | up 2 | Same as above | - | |
| UCA1 | up 2 | Contributes to HCC through inhibition of miR-216b and activation of FGFR1/ERK pathway | [62,129] | |
| WRAP53 | up 2 | Promotes cancer cell survival by regulating p53 | [62,130] | |
| HEIH | up 3 | Promotes tumor progression by binding to a causing the repression of EZH2 target genes | [131,132] | |
| CASC2 | down 4 | Suppresses EMT of HCC cells through CASC2/miR-367/FBXW7 axis | [133,134] | |
| TUG1 | up 4 | Promotes cell growth and apoptosis by epigenetically silencing of KLF2 | [133,135] | |
| HOTAIR | up 5 | Promotes tumorigenesis by suppression of miR-122 and activation of Cyclin G1 | [136,137] | |
| HOTTIP | up 5 | Unknown in HCC; promotes cell proliferation and invasion by modulating HOXA13 in pancreatic cancer | [136,138] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Unfried, J.P.; Fortes, P. LncRNAs in HCV Infection and HCV-Related Liver Disease. Int. J. Mol. Sci. 2020, 21, 2255. https://doi.org/10.3390/ijms21062255
Unfried JP, Fortes P. LncRNAs in HCV Infection and HCV-Related Liver Disease. International Journal of Molecular Sciences. 2020; 21(6):2255. https://doi.org/10.3390/ijms21062255
Chicago/Turabian StyleUnfried, Juan P., and P. Fortes. 2020. "LncRNAs in HCV Infection and HCV-Related Liver Disease" International Journal of Molecular Sciences 21, no. 6: 2255. https://doi.org/10.3390/ijms21062255
APA StyleUnfried, J. P., & Fortes, P. (2020). LncRNAs in HCV Infection and HCV-Related Liver Disease. International Journal of Molecular Sciences, 21(6), 2255. https://doi.org/10.3390/ijms21062255