Combined Chibby and β-Catenin Predicts Clinical Outcomes in Patients with Hepatocellular Carcinoma

 , , ,
, , ,
Abstract
1. Introduction
2. Results
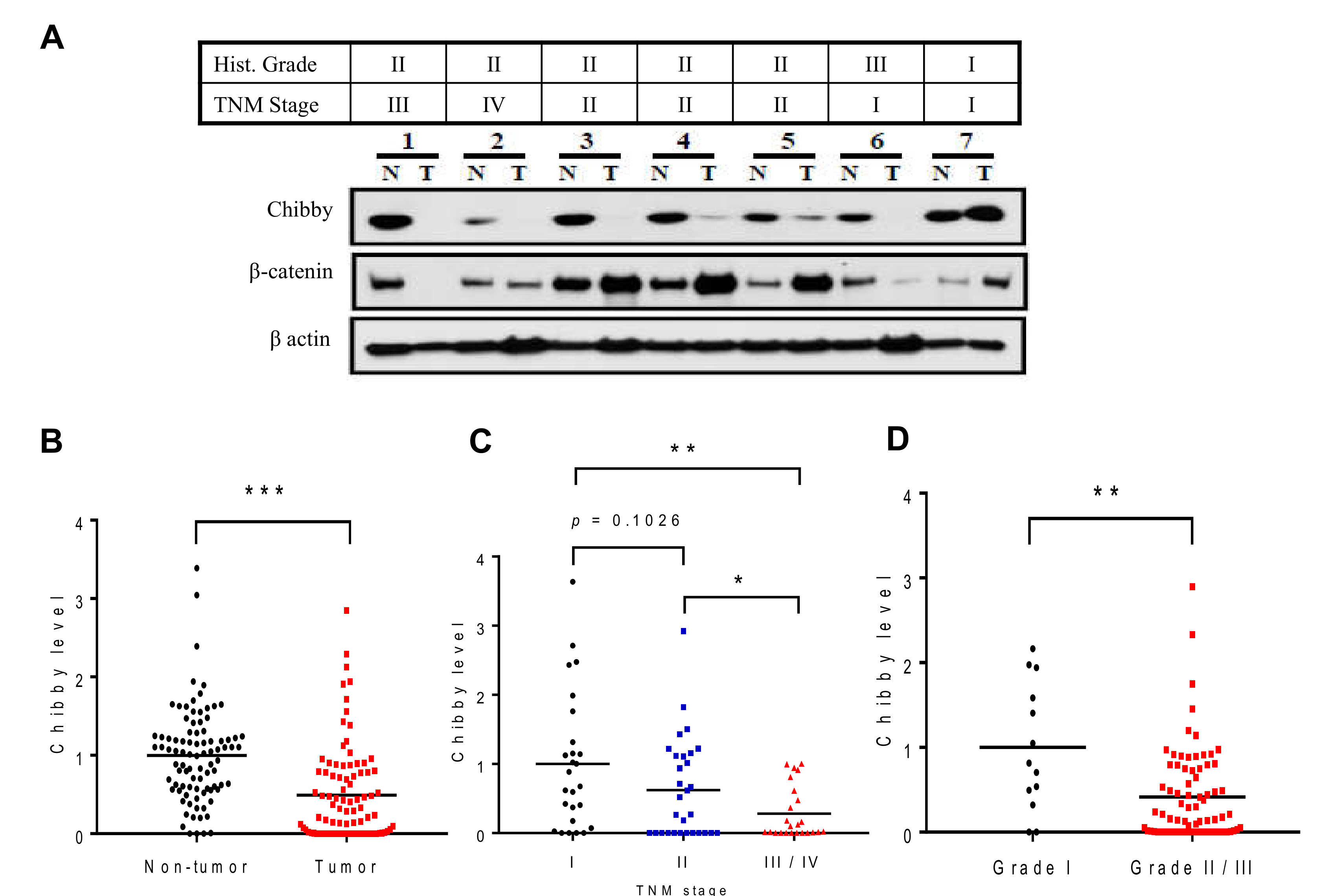
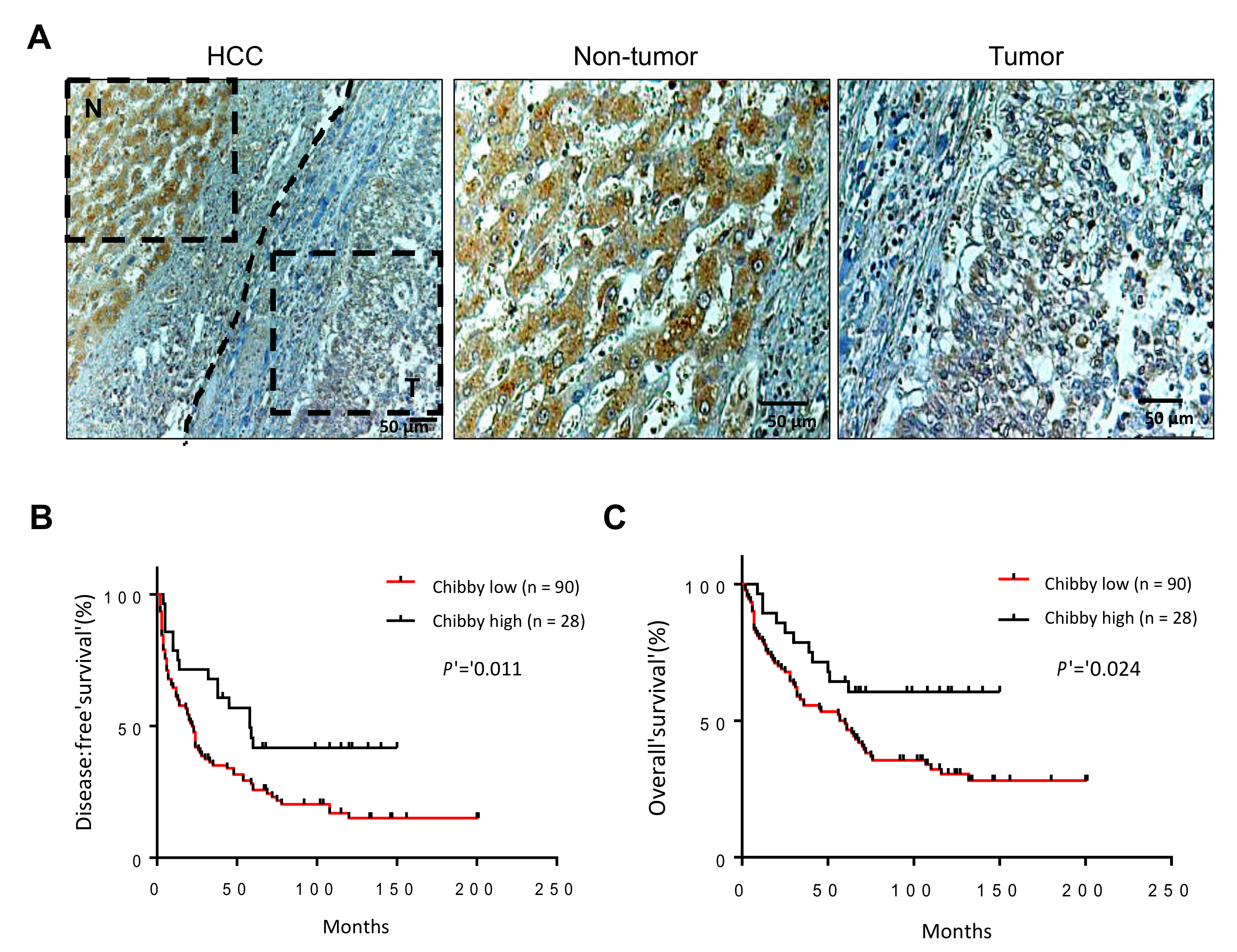
2.1. Low Expression of Chibby Correlates with High Stage of HCC
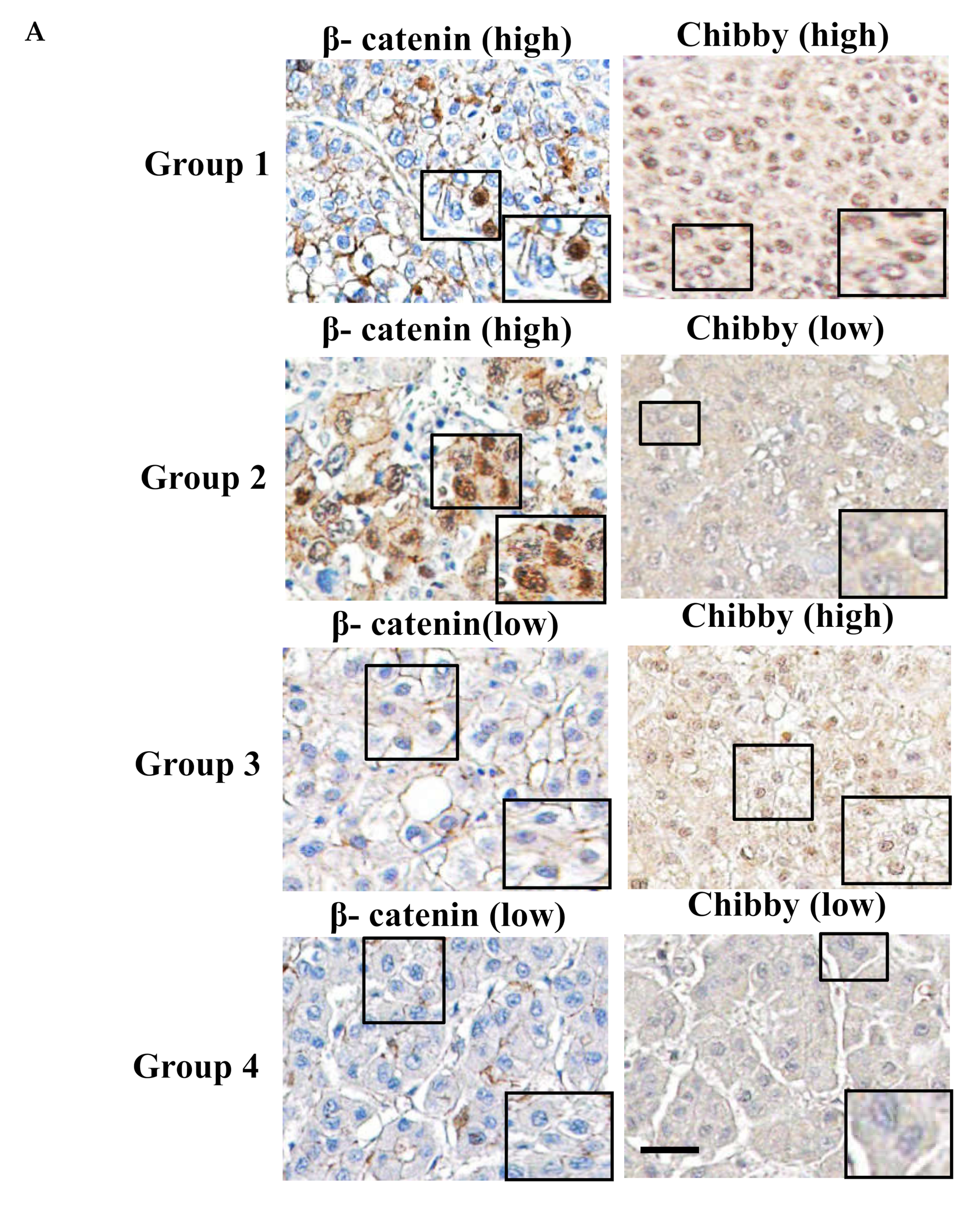
2.2. Correlation between Chibby Expression and β-Catenin Expression in HCC Specimens
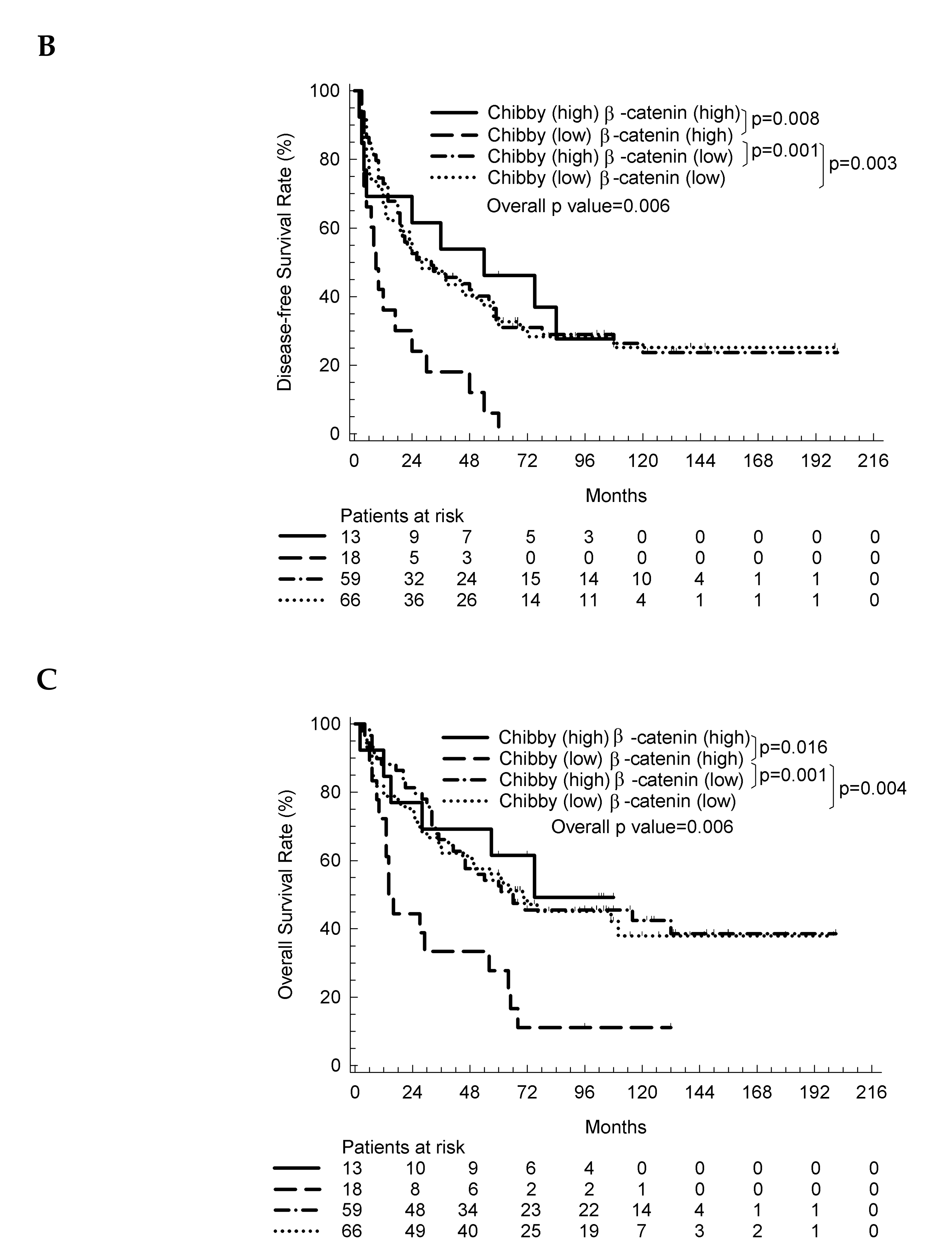
2.3. The Nuclear Expression of Chibby Predict Disease-Free Survival and Overall Survival in HCC Patients with Nuclear Expression of β-Catenin
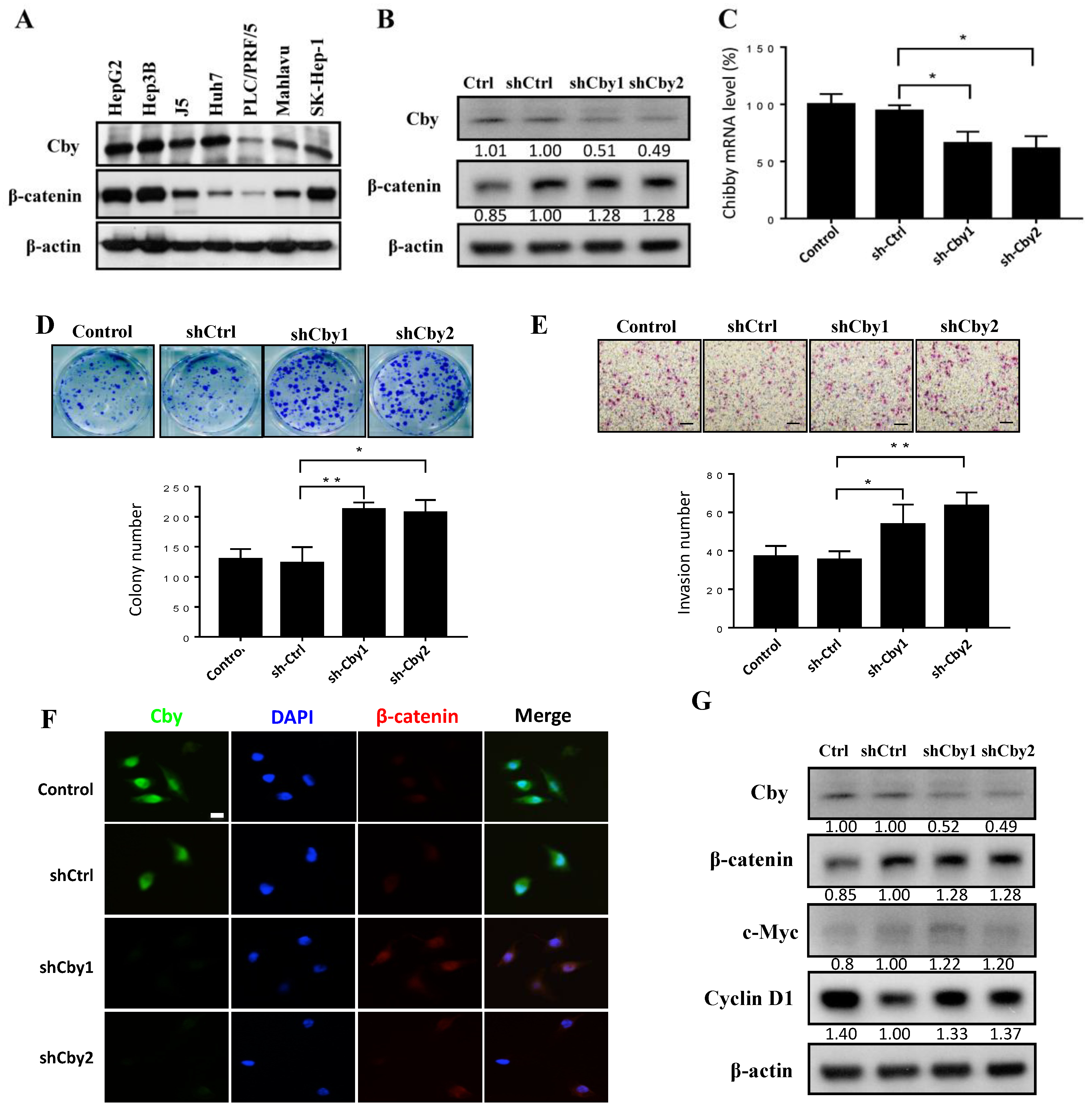
2.4. Knockdown of Chibby Promotes β-Catenin Signaling and HCC Cell Proliferation and Invasion
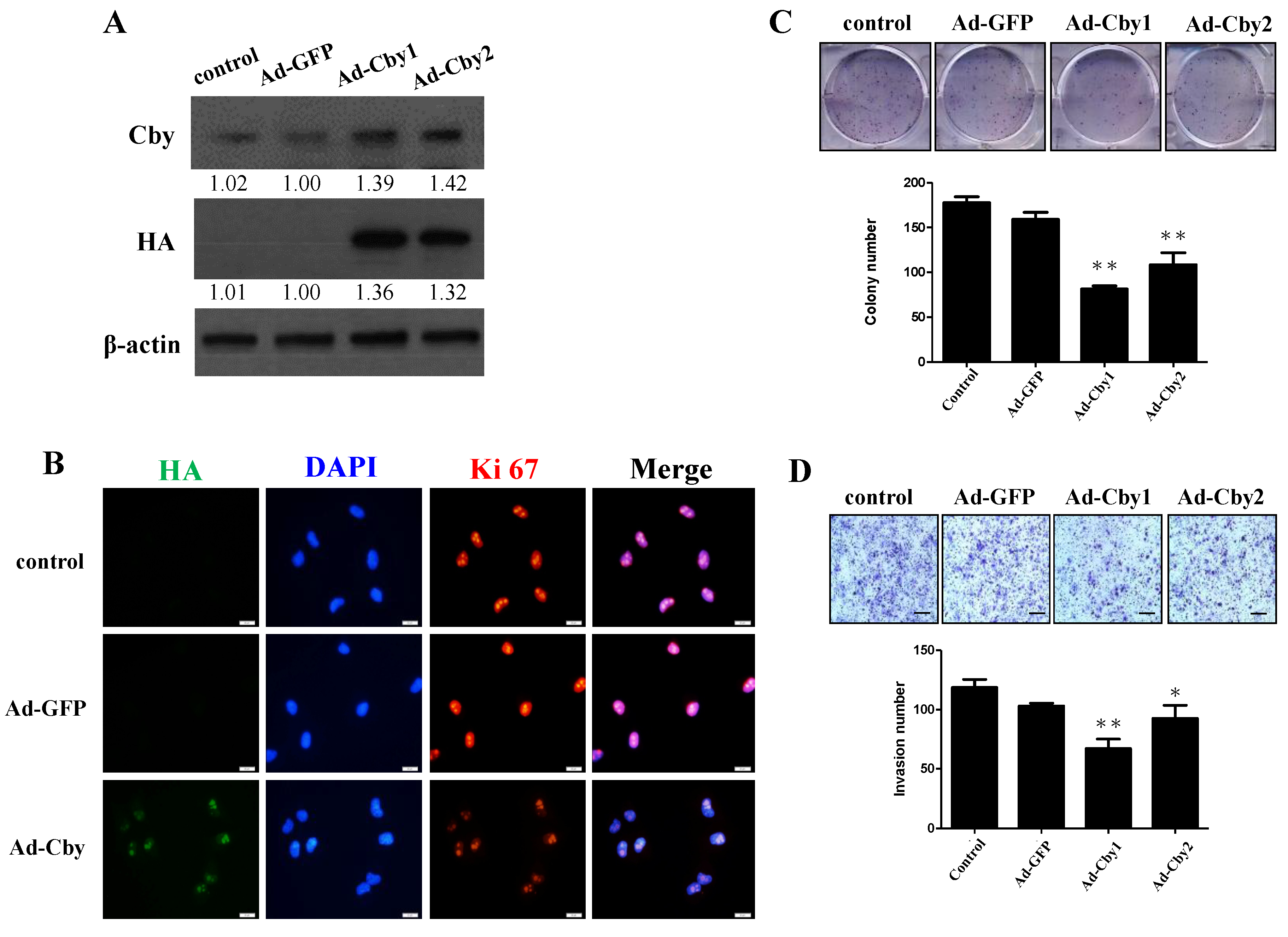
2.5. Expression of Chibby Inhibits β-Catenin Signaling and HCC Cell Proliferation and Invasion
3. Discussion
4. Materials and Methods
4.1. Patients and Tissue Samples
4.2. Immunohistochemical Staining and Scoring
4.3. RNA Isolation and Quantitative Reverse-Transcriptase Polymerase Chain Reaction
4.4. Western Blot Analysis
4.5. Immunofluorescence Staining
4.6. Cell Culture and Transfection Protocols
4.7. Adenoviral Vectors-Mediated Gene Delivery
4.8. Knockdown of Endogenous Chibby
4.9. Colonies Formation Assay
4.10. Invasion Assay
4.11. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Torre, L.A.; Bray, F.; Siegel, R.L.; Ferlay, J.; Lortet-Tieulent, J.; Jemal, A. Global cancer statistics, 2012. CA A Cancer J. Clin. 2015, 65, 87–108. [Google Scholar] [CrossRef] [PubMed]
- Llovet, J.M.; Burroughs, A.; Bruix, J. Hepatocellular carcinoma. Lancet 2003, 362, 1907–1917. [Google Scholar] [CrossRef]
- Thorgeirsson, S.S.; Grisham, J.W. Molecular pathogenesis of human hepatocellular carcinoma. Nat. Genet. 2002, 31, 339–346. [Google Scholar] [CrossRef] [PubMed]
- De La Coste, A.; Romagnolo, B.; Billuart, P.; Renard, C.A.; Buendia, M.A.; Soubrane, O.; Fabre, M.; Chelly, J.; Beldjord, C.; Kahn, A.; et al. Somatic mutations of the beta-catenin gene are frequent in mouse and human hepatocellular carcinomas. Proc. Natl. Acad. Sci. USA 1998, 95, 8847–8851. [Google Scholar] [CrossRef]
- Llovet, J.M.; Zucman-Rossi, J.; Pikarsky, E.; Sangro, B.; Schwartz, M.; Sherman, M.; Gores, G. Hepatocellular carcinoma. Nat. Rev. Dis. Primers 2016, 2, 16018. [Google Scholar] [CrossRef]
- Schulze, K.; Imbeaud, S.; Letouze, E.; Alexandrov, L.B.; Calderaro, J.; Rebouissou, S.; Couchy, G.; Meiller, C.; Shinde, J.; Soysouvanh, F.; et al. Exome sequencing of hepatocellular carcinomas identifies new mutational signatures and potential therapeutic targets. Nat. Genet. 2015, 47, 505–511. [Google Scholar] [CrossRef]
- Anastas, J.N.; Moon, R.T. WNT signalling pathways as therapeutic targets in cancer. Nat. Rev. Cancer 2013, 13, 11–26. [Google Scholar] [CrossRef]
- Inagawa, S.; Itabashi, M.; Adachi, S.; Kawamoto, T.; Hori, M.; Shimazaki, J.; Yoshimi, F.; Fukao, K. Expression and prognostic roles of beta-catenin in hepatocellular carcinoma: Correlation with tumor progression and postoperative survival. Clin. Cancer Res. 2002, 8, 450–456. [Google Scholar]
- Nhieu, J.T.; Renard, C.A.; Wei, Y.; Cherqui, D.; Zafrani, E.S.; Buendia, M.A. Nuclear accumulation of mutated beta-catenin in hepatocellular carcinoma is associated with increased cell proliferation. Am. J. Pathol. 1999, 155, 703–710. [Google Scholar] [CrossRef]
- Takemaru, K.; Yamaguchi, S.; Lee, Y.S.; Zhang, Y.; Carthew, R.W.; Moon, R.T. Chibby, a nuclear beta-catenin-associated antagonist of the Wnt/Wingless pathway. Nature 2003, 422, 905–909. [Google Scholar] [CrossRef]
- Tolwinski, N.S.; Wieschaus, E. A nuclear function for armadillo/beta-catenin. PLoS Biol. 2004, 2, E95. [Google Scholar] [CrossRef] [PubMed]
- Shtutman, M.; Zhurinsky, J.; Simcha, I.; Albanese, C.; D’Amico, M.; Pestell, R.; Ben-Ze’ev, A. The cyclin D1 gene is a target of the beta-catenin/LEF-1 pathway. Proc. Natl. Acad. Sci. USA 1999, 96, 5522–5527. [Google Scholar] [CrossRef] [PubMed]
- He, T.C.; Sparks, A.B.; Rago, C.; Hermeking, H.; Zawel, L.; da Costa, L.T.; Morin, P.J.; Vogelstein, B.; Kinzler, K.W. Identification of c-MYC as a target of the APC pathway. Science 1998, 281, 1509–1512. [Google Scholar] [CrossRef] [PubMed]
- Takemaru, K.; Fischer, V.; Li, F.Q. Fine-tuning of nuclear-catenin by Chibby and 14-3-3. Cell Cycle 2009, 8, 210–213. [Google Scholar] [CrossRef] [PubMed]
- Fischer, V.; Brown-Grant, D.A.; Li, F.Q. Chibby suppresses growth of human SW480 colon adenocarcinoma cells through inhibition of beta-catenin signaling. J. Mol. Signal 2012, 7, 6. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Ren, G.; Zhao, D.A.; Li, B.A.; Cai, C.F.; Zhou, Y.; Luo, X.Y. Downregulated chibby in laryngeal squamous cell carcinoma with increased expression in laryngeal carcinoma Hep-2 cells. Oncol. Rep. 2014, 32, 1947–1956. [Google Scholar] [CrossRef][Green Version]
- Xu, H.T.; Li, Q.C.; Dai, S.D.; Xie, X.M.; Liu, D.I.; Wang, E.H. The expression patterns and correlations of chibby, beta-catenin, and DNA methyltransferase-1 and their clinicopathological significance in lung cancers. APMIS 2011, 119, 750–758. [Google Scholar] [CrossRef]
- Collins, F.S.; Varmus, H. A new initiative on precision medicine. N. Engl. J. Med. 2015, 372, 793–795. [Google Scholar] [CrossRef]
- Bruix, J.; Sherman, M. Management of hepatocellular carcinoma. Hepatology 2005, 42, 1208–1236. [Google Scholar] [CrossRef]
- Bruix, J.; Sherman, M.; Llovet, J.M.; Beaugrand, M.; Lencioni, R.; Burroughs, A.K.; Christensen, E.; Pagliaro, L.; Colombo, M.; Rodes, J.; et al. Clinical management of hepatocellular carcinoma. Conclusions of the barcelona-2000 easl conference. european association for the study of the liver. J. Hepatol. 2001, 35, 421–430. [Google Scholar] [CrossRef]
- Edge, S.B.; Compton, C.C. The American Joint Committee on Cancer: The 7th edition of the AJCC cancer staging manual and the future of TNM. Ann. Surg. Oncol. 2010, 17, 1471–1474. [Google Scholar] [CrossRef] [PubMed]
- Edmondson, H.A.; Steiner, P.E. Primary carcinoma of the liver: A study of 100 cases among 48,900 necropsies. Cancer 1954, 7, 462–503. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinicopathologic | β-Catenin (high)/Chibby (high) (n = 13) | β-Catenin (high)/Chibby (low) (n = 18) | β-Catenin (low)/Chibby (high) (n = 59) | β-Catenin (low)/Chibby (low) (n = 66) | p value |
|---|---|---|---|---|---|
| Age (>60 vs. ≤60 years) | 4/9 | 9/9 | 15/44 | 28/38 | 0.125 |
| Sex (Male vs. Female) | 11/2 | 13/5 | 46/13 | 55/11 | 0.689 |
| AFP (>200 vs. ≤200 ng/L) | 6/7 | 8/10 | 26/33 | 25/41 | 0.875 |
| HBsAg (positive vs. negative) | 9/4 | 12/6 | 40/16 | 47/16 | 0.914 |
| HCV Ab (positive vs. negative) | 3/10 | 3/15 | 14/42 | 15/48 | 0.909 |
| Liver cirrhosis (yes vs. no) | 7/6 | 11/7 | 38/21 | 78/28 | 0.837 |
| Tumor size (>5 vs. ≤5 cm) | 10/3 | 11/7 | 30/29 | 32/34 | 0.250 |
| Tumor number (multiple vs. single) * | 2/11 | 9/9 | 16/43 | 11/55 | 0.025 |
| TNM Stage (III + IV vs. I + II) | 9/4 | 13/5 | 28/31 | 30/36 | 0.108 |
| Histology grade (Poor vs. Well + Moderate) | 6/7 | 6/12 | 17/42 | 15/51 | 0.349 |
| Univariate | Multivariate | ||||
|---|---|---|---|---|---|
| Variable | Comparison | HR (95%CI) | p value | HR (95%CI) | p value |
| Age (years) | >60 vs. ≤60 | 0.755 (0.513–1.110) | 0.153 | NS | |
| Sex | Male vs. Female | 1.362 (0.856–2.167) | 0.193 | NS | |
| AFP (ng/mL) | >200 vs. ≤200 | 1.676 (1.160–2.421) | 0.006 | 1.907 (1.297–2.803) | 0.001 |
| HBsAg | Positive vs. Negative | 1.306 (0.855–1.996) | 0.217 | NS | |
| HCV Ab | Positive vs. Negative | 0.811 (0.519–1.267) | 0.358 | NA | |
| Liver cirrhosis | Yes vs. No | 1.494 (1.001–2.169) | 0.049 | 1.795 (1.201–2.683) | 0.004 |
| Tumor size (cm) | >5 vs. ≤5 | 1.904 (1.313–2.761) | 0.001 | NS | |
| Tumor number | Multiple vs. Single | 1.534 (1.019–2.310) | 0.041 | NS | |
| TNM stages | III + IV vs. I + II | 2.730 (1.870–3.983) | <0.001 | 2.808 (1.898–4.156) | <0.001 |
| Histology grade | Poor vs. Well + Moderate | 1.557 (1.052–2.304) | 0.027 | NS | |
| β-catenin | High vs. Low | 1.505 (0.971–2.332) | 0.068 | NS | |
| Chibby | High vs. Low | 0.802 (0.556–1.157) | 0.239 | NS | |
| β-catenin/Chibby | High/Low vs. others | 2.450 (1.448–4.144) | 0.001 | 1.986 (1.160–3.400) | 0.012 |
| Univariate | Multivariate | ||||
|---|---|---|---|---|---|
| Variable | Comparison | HR (95%CI) | p value | HR (95%CI) | p value |
| Age (years) | >60 vs. ≤60 | 0.966 (0.633–1.474) | 0.892 | NA | |
| Sex | Male vs. Female | 1.204 (0.719–2.014) | 0.480 | NA | |
| AFP (ng/mL) | >200 vs. ≤200 | 1.580 (1.050–2.377) | 0.028 | 1.638 (1.076–2.493) | 0.021 |
| HBsAg | Positive vs. Negative | 1.264 (0.787–2.031) | 0.333 | NA | |
| HCV Ab | Positive vs. Negative | 0.804 (0.489–1.322) | 0.389 | NA | |
| Liver cirrhosis | Yes vs. No | 1.307 (0.854–2.001) | 0.217 | NS | |
| Tumor size (cm) | >5 vs. ≤5 | 2.407 (1.343–3.119) | 0.001 | NS | |
| Tumor number | Multiple vs. Single | 1.221 (0.762–1.956) | 0.407 | NA | |
| TNM stages | III + IV vs. I + II | 3.351 (2.165–5.188) | <0.001 | 3.135 (1.979–4.968) | <0.001 |
| Histology grade | Poor vs. well + moderate | 1.858 (1.212–2.850) | 0.004 | NS | |
| β-catenin | High vs. Low | 1.612 (0.996–2.609) | 0.052 | NS | |
| Chibby | High vs. Low | 0.756 (0.501–1.140) | 0.182 | NS | |
| β-catenin/Chibby | High/Low vs. others | 2.534 (1.471–4.363) | 0.001 | 2.207 (1.270–3.835) | 0.005 |
| Patient Demographics | |
|---|---|
| Age [years; median (range)] | 56 (5–82) |
| Sex (M:F) | 125:31 |
| AFP [ng/mL; median (range)] | 87 (2–80000) |
| Tumor size [cm; median (range)] a | 5 (1–20) |
| Tumor number (single:multiple) | 37:119 |
| Liver cirrhosis, n (%) | 94 (59.9) |
| Hepatitis (B:C:B+C:NBNC) | 101:28:7:20 |
| TNM stage (I:II:III:IV) | 22:54:46:34 |
| Pathological Features | |
| Capsule (Yes:No) | 69:86 |
| Microvascular invasion (Yes:No) | 76:77 |
| Histological grade (well:moderate:poor) | 38:74:44 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsai, M.-C.; Huang, C.-C.; Wei, Y.-C.; Liu, T.-T.; Lin, M.-T.; Yi, L.-N.; Lin, P.-R.; Wang, C.-C.; Chu, T.-H.; Hsiao, C.-C.; et al. Combined Chibby and β-Catenin Predicts Clinical Outcomes in Patients with Hepatocellular Carcinoma. Int. J. Mol. Sci. 2020, 21, 2060. https://doi.org/10.3390/ijms21062060
Tsai M-C, Huang C-C, Wei Y-C, Liu T-T, Lin M-T, Yi L-N, Lin P-R, Wang C-C, Chu T-H, Hsiao C-C, et al. Combined Chibby and β-Catenin Predicts Clinical Outcomes in Patients with Hepatocellular Carcinoma. International Journal of Molecular Sciences. 2020; 21(6):2060. https://doi.org/10.3390/ijms21062060
Chicago/Turabian StyleTsai, Ming-Chao, Chao-Cheng Huang, Yu-Ching Wei, Ting-Ting Liu, Ming-Tsung Lin, Li-Na Yi, Pey-Ru Lin, Chih-Chi Wang, Tian-Huei Chu, Chang-Chun Hsiao, and et al. 2020. "Combined Chibby and β-Catenin Predicts Clinical Outcomes in Patients with Hepatocellular Carcinoma" International Journal of Molecular Sciences 21, no. 6: 2060. https://doi.org/10.3390/ijms21062060
APA StyleTsai, M.-C., Huang, C.-C., Wei, Y.-C., Liu, T.-T., Lin, M.-T., Yi, L.-N., Lin, P.-R., Wang, C.-C., Chu, T.-H., Hsiao, C.-C., Hu, T.-H., & Tai, M.-H. (2020). Combined Chibby and β-Catenin Predicts Clinical Outcomes in Patients with Hepatocellular Carcinoma. International Journal of Molecular Sciences, 21(6), 2060. https://doi.org/10.3390/ijms21062060