Silver Nanoparticles in Zebrafish (Danio rerio) Embryos: Uptake, Growth and Molecular Responses

 , ,
, , 
Abstract
1. Introduction
2. Results
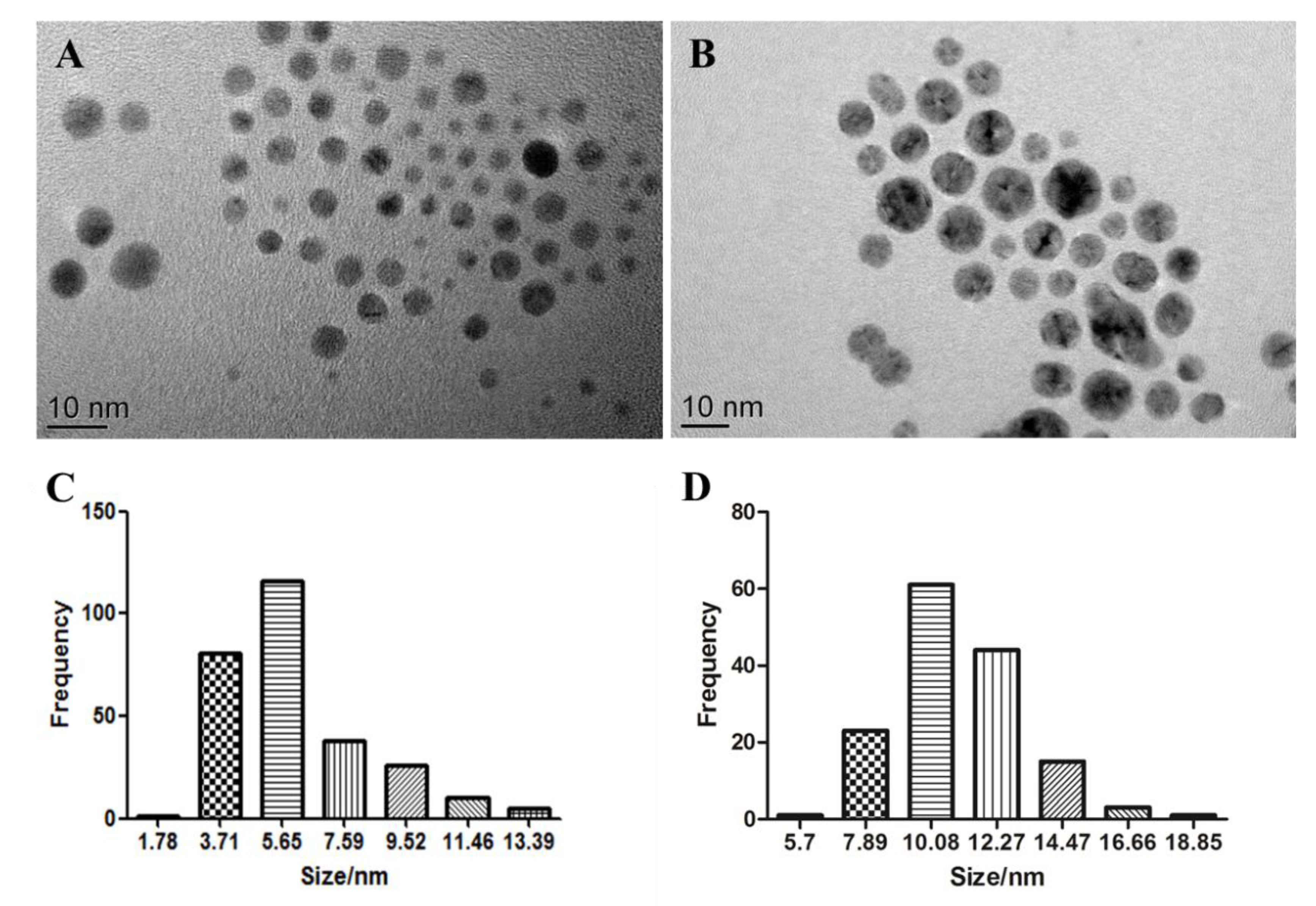
2.1. Characterisation of AgNPs
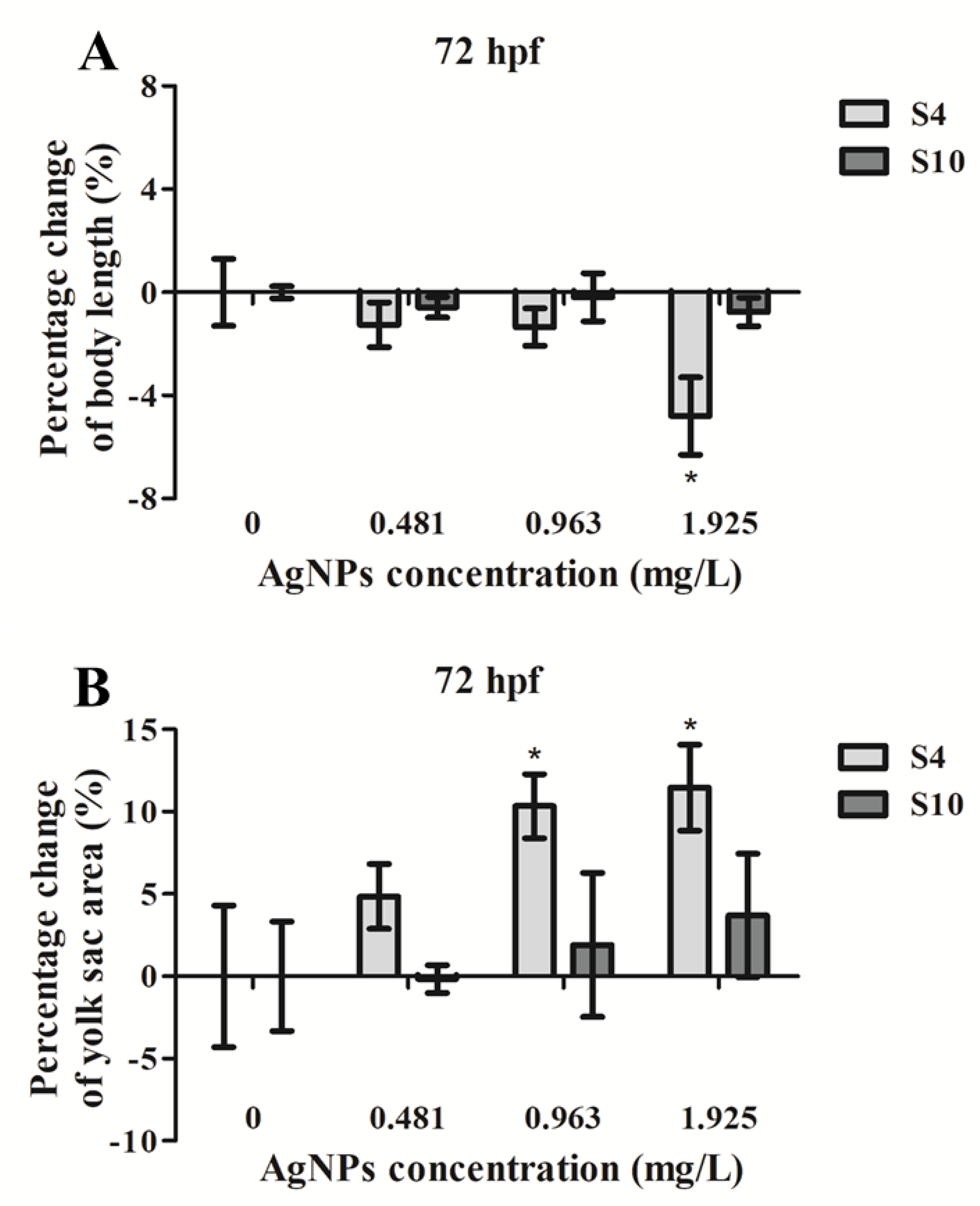
2.2. Embryo Body Length and Yolk Sac Area Impacts
2.3. AgNP Uptake
2.4. qPCR Analysis of Candidate Gene Expression in Fish Embryos
3. Discussion
4. Materials and Methods
4.1. Materials and Characterisation of AgNPs
4.2. Zebrafish Maintenance and Embryo Collection
4.3. Exposure Design
4.4. Uptake of AgNPs in Fish Embryos
4.5. Target Gene Isolation and Characterisation
4.6. Quantitative (Q) PCR Analysis of mRNA Expression
4.7. Statistical Analyses
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Jeevanandam, J.; Barhoum, A.; Chan, Y.S.; Dufresne, A.; Danquah, M.K. Review on nanoparticles and nanostructured materials: history, sources, toxicity and regulations. Beilstein J. Nanotechnol. 2018, 9, 1050–1074. [Google Scholar] [CrossRef] [PubMed]
- Pulit-Prociak, J.; Chwastowski, J.; Bittencourt Rodrigues, L.; Banach, M. Analysis of the physicochemical properties of antimicrobial compositions with zinc oxide nanoparticles. Sci. Technol. Adv. Mater. 2019, 20, 1150–1163. [Google Scholar] [CrossRef] [PubMed]
- McGillicuddy, E.; Murray, I.; Shevlin, D.; Morrisson, L.; Cormican, M.; Fogarty, A.; Cummins, E.; Dockery, E.; Dunlop, P.; Rowan, N.J. Detection, Toxicology, Environmental Fate and Risk Assessment of Nanoparticles in the Aquatic Environment (DeTER); Report No. 259; Environmental Protection Agency: Washington, DC, USA, 2018.
- Hedberg, J.; Oromieh, A.G.; Kleja, D.B.; Wallinder, I.O. Sorption and dissolution of bare and coated silver nanoparticles in soil suspensions—Influence of soil and particle characteristics. J. Environ. Sci. Health 2015, 50, 891–900. [Google Scholar]
- Saleeb, N.; Gooneratne, R.; Cavanagh, J.; Bunt, C.; Hossain, A.; Gaw, S.; Robinson, B. The Mobility of Silver Nanoparticles and Silver Ions in the Soil-Plant System. J. Environ. Qual. 2019, 48, 1835–1841. [Google Scholar] [CrossRef]
- Pulit-Prociak, J.; Banach, M. Silver nanoparticles–a material of the futu? Open Chem. 2016, 14, 76–91. [Google Scholar] [CrossRef]
- Al-Sid-Cheikh, M.; Rouleau, C.; Bussolaro, D.; Oliveira Ribeiro, C.A.; Pelletier, E. Tissue Distribution of Radiolabeled 110mAg Nanoparticles in Fish: Arctic Charr (Salvelinus alpinus). Environ. Sci. Technol. 2019, 53, 12043–12053. [Google Scholar] [CrossRef]
- Keller, A.A.; McFerran, S.; Lazareva, A.; Suh, S. Global life cycle releases of engineered nanomaterials. J. Nanoparticle Res. 2013, 15, 1692. [Google Scholar] [CrossRef]
- Massarsky, A.; Trudeau, V.L.; Moon, T.W. Predicting the environmental impact of nanosilver. Environ. Toxicol. Pharmacol. 2014, 38, 861–873. [Google Scholar] [CrossRef]
- Gao, J.; Mahapatra, C.T.; Mapes, C.D.; Khlebnikova, M.; Wei, A.; Sepúlveda, M.S. Vascular toxicity of silver nanoparticles to developing zebrafish (Danio rerio). Nanotoxicology 2016, 10, 1363–1372. [Google Scholar] [CrossRef]
- Mosselhy, D.A.; He, W.; Li, D.; Meng, Y.; Feng, Q. Silver nanoparticles: in vivo toxicity in zebrafish embryos and a comparison to silver nitrate. J. Nanopart. Res. 2016, 18, 222. [Google Scholar] [CrossRef]
- Boyle, D.; Goss, G.G. Effects of silver nanoparticles in early life-stage zebrafish are associated with particle dissolution and the toxicity of soluble silver. Nano Impact 2018, 12, 1–8. [Google Scholar] [CrossRef]
- Wu, Y.; Zhou, Q. Dose-and time-related changes in aerobic metabolism, chorionic disruption, and oxidative stress in embryonic medaka (Oryzias latipes): underlying mechanisms for silver nanoparticle developmental toxicity. Aquat. Toxicol. 2012, 124, 238–246. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.E.; Kim, S.; Ahn, J.H.; Youn, P.; Kang, J.S.; Park, K.; Yi, J.; Ryu, D.-Y. Induction of oxidative stress and apoptosis by silver nanoparticles in the liver of adult zebrafish. Aquat. Toxicol. 2010, 100, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Bilberg, K.; Hovgaard, M.B.; Besenbacher, F.; Baatrup, E. In vivo toxicity of silver nanoparticles and silver ions in zebrafish (Danio rerio). J. Toxicol. 2012, 2012. [Google Scholar] [CrossRef]
- Kashiwada, S.; Ariza, M.E.; Kawaguchi, T.; Nakagame, Y.; Jayasinghe, B.S.; Gärtner, K.; Nakamura, H.; Kagami, Y.; Sabo-Attwood, T.; Ferguson, P.L. Silver nanocolloids disrupt medaka embryogenesis through vital gene expressions. Environ. Sci. Technol. 2012, 46, 6278–6287. [Google Scholar] [CrossRef]
- Ašmonaitė, G.; Boyer, S.; de Souza, K.B.; Wassmur, B.; Sturve, J. Behavioural toxicity assessment of silver ions and nanoparticles on zebrafish using a locomotion profiling approach. Aquat. Toxicol. 2016, 173, 143–153. [Google Scholar] [CrossRef]
- van Aerle, R.; Lange, A.; Moorhouse, A.; Paszkiewicz, K.; Ball, K.; Johnston, B.D.; de-Bastos, E.; Booth, T.; Tyler, C.R.; Santos, E.M. Molecular mechanisms of toxicity of silver nanoparticles in zebrafish embryos. Environ. Sci. Technol. 2013, 47, 8005–8014. [Google Scholar] [CrossRef]
- Hawkins, A.D.; Thornton, C.; Kennedy, A.J.; Bu, K.; Cizdziel, J.; Jones, B.W.; Steevens, J.A.; Willett, K.L. Gill histopathologies following exposure to nanosilver or silver nitrate. J. Toxicol. Environ. Health Part A 2015, 78, 301–315. [Google Scholar] [CrossRef]
- Powers, C.M.; Slotkin, T.A.; Seidler, F.J.; Badireddy, A.R.; Padilla, S. Silver nanoparticles alter zebrafish development and larval behavior: distinct roles for particle size, coating and composition. Neurotoxicol. Teratol. 2011, 33, 708–714. [Google Scholar] [CrossRef]
- Griffitt, R.J.; Hyndman, K.; Denslow, N.D.; Barber, D.S. Comparison of molecular and histological changes in zebrafish gills exposed to metallic nanoparticles. Toxicol. Sci. 2009, 107, 404–415. [Google Scholar] [CrossRef]
- Xin, Q.; Rotchell, J.M.; Cheng, J.; Yi, J.; Zhang, Q. Silver nanoparticles affect the neural development of zebrafish embryos. J. Appl. Toxicol. 2015, 35, 1481–1492. [Google Scholar] [CrossRef] [PubMed]
- Osborne, O.J.; Lin, S.; Chang, C.H.; Ji, Z.; Yu, X.; Wang, X.; Lin, S.; Xia, T.; Nel, A.E. Organ-specific and size-dependent Ag nanoparticle toxicity in gills and intestines of adult zebrafish. Acs Nano 2015, 9, 9573–9584. [Google Scholar] [CrossRef] [PubMed]
- Osborne, O.J.; Johnston, B.D.; Moger, J.; Balousha, M.; Lead, J.R.; Kudoh, T.; Tyler, C.R. Effects of particle size and coating on nanoscale Ag and TiO2 exposure in zebrafish (Danio rerio) embryos. Nanotoxicology 2013, 7, 1315–1324. [Google Scholar] [CrossRef] [PubMed]
- Morgan, T.P.; Wood, C.M. A relationship between gill silver accumulation and acute silver toxicity in the freshwater rainbow trout: support for the acute silver biotic ligand model. Environ. Toxicol. Chem. Int. J. 2004, 23, 1261–1267. [Google Scholar] [CrossRef]
- Bar-Ilan, O.; Albrecht, R.M.; Fako, V.E.; Furgeson, D.Y. Toxicity assessments of multisized gold and silver nanoparticles in zebrafish embryos. Small 2009, 5, 1897–1910. [Google Scholar] [CrossRef]
- Olasagasti, M.; Gatti, A.M.; Capitani, F.; Barranco, A.; Pardo, M.A.; Escuredo, K.; Rainieri, S. Toxic effects of colloidal nanosilver in zebrafish embryos. J. Appl. Toxicol. 2014, 34, 562–575. [Google Scholar] [CrossRef]
- Muth-Köhne, E.; Sonnack, L.; Schlich, K.; Hischen, F.; Baumgartner, W.; Hund-Rinke, K.; Schäfers, C.; Fenske, M. The toxicity of silver nanoparticles to zebrafish embryos increases through sewage treatment processes. Ecotoxicology 2013, 22, 1264–1277. [Google Scholar] [CrossRef]
- Böhme, S.; Baccaro, M.; Schmidt, M.; Potthoff, A.; Stärk, H.-J.; Reemtsma, T.; Kühnel, D. Metal uptake and distribution in the zebrafish (Danio rerio) embryo: differences between nanoparticles and metal ions. Environ. Sci. Nano 2017, 4, 1005–1015. [Google Scholar] [CrossRef]
- Al-Awady, M.J.; Greenway, G.M.; Paunov, V.N. Nanotoxicity of polyelectrolyte-functionalized titania nanoparticles towards microalgae and yeast: role of the particle concentration, size and surface charge. Rsc. Adv. 2015, 5, 37044–37059. [Google Scholar] [CrossRef]
- Lee, K.J.; Browning, L.M.; Nallathamby, P.D.; Osgood, C.J.; Xu, X.-H.N. Silver nanoparticles induce developmental stage-specific embryonic phenotypes in zebrafish. Nanoscale 2013, 5, 11625–11636. [Google Scholar] [CrossRef]
- Cheng, J.; Flahaut, E.; Cheng, S.H. Effect of carbon nanotubes on developing zebrafish (Danio rerio) embryos. Environ. Toxicol. Chem. Int. J. 2007, 26, 708–716. [Google Scholar] [CrossRef] [PubMed]
- Abramenko, N.B.; Demidova, T.B.; Abkhalimov, E.V.; Ershov, B.G.; Krysanov, E.Y.; Kustov, L.M. Ecotoxicity of different-shaped silver nanoparticles: Case of zebrafish embryos. J. Hazard. Mater. 2018, 347, 89–94. [Google Scholar] [CrossRef]
- Rajasekaran, A.K.; Rajasekaran, S.A. Role of Na-K-ATPase in the assembly of tight junctions. Am. J. Physiol. Renal. Physiol. 2003, 285, F388–F396. [Google Scholar] [CrossRef] [PubMed]
- FRÖMTER, E.; DIAMOND, J. Route of passive ion permeation in epithelia. Nat. New Biol. 1972, 235, 9–13. [Google Scholar] [CrossRef] [PubMed]
- Scown, T.M.; Santos, E.M.; Johnston, B.D.; Gaiser, B.; Baalousha, M.; Mitov, S.; Lead, J.R.; Stone, V.; Fernandes, T.F.; Jepson, M. Effects of aqueous exposure to silver nanoparticles of different sizes in rainbow trout. Toxicol. Sci. 2010, 115, 521–534. [Google Scholar] [CrossRef] [PubMed]
- Park, H.-G.; Yeo, M.-K. Metabolic gene expression profiling of Zebrafish embryos exposed to silver nanocolloids and nanotubes. Mol. Cell. Toxicol. 2014, 10, 401–409. [Google Scholar] [CrossRef]
- Garcia-Reyero, N.l.; Kennedy, A.J.; Escalon, B.L.; Habib, T.; Laird, J.G.; Rawat, A.; Wiseman, S.; Hecker, M.; Denslow, N.; Steevens, J.A. Differential effects and potential adverse outcomes of ionic silver and silver nanoparticles in vivo and in vitro. Environ. Sci. Technol. 2014, 48, 4546–4555. [Google Scholar] [CrossRef]
- Rokka, A.; Antonenkov, V.D.; Soininen, R.; Immonen, H.L.; Pirilä, P.L.; Bergmann, U.; Sormunen, R.T.; Weckström, M.; Benz, R.; Hiltunen, J.K. Pxmp2 is a channel-forming protein in mammalian peroxisomal membrane. PLoS ONE 2009, 4, e5090. [Google Scholar] [CrossRef]
- Schrader, M.; Fahimi, H.D. Peroxisomes and oxidative stress. Biochim. Biophys. Acta 2006, 1763, 1755–1766. [Google Scholar] [CrossRef]
- Lee, B.; Duong, C.N.; Cho, J.; Lee, J.; Kim, K.; Seo, Y.; Kim, P.; Choi, K.; Yoon, J. Toxicity of citrate-capped silver nanoparticles in common carp (Cyprinus carpio). Biomed. Res. Int. 2012, 2012. [Google Scholar] [CrossRef]
- Jevtov, I.; Samuelsson, T.; Yao, G.; Amsterdam, A.; Ribbeck, K. Zebrafish as a model to study live mucus physiology. Sci. Rep. 2014, 4, 6653. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, F.; Gallego-Urrea, J.A.; Jurkschat, K.; Crossley, A.; Hassellöv, M.; Taylor, C.; Soares, A.M.; Loureiro, S. Silver nanoparticles and silver nitrate induce high toxicity to Pseudokirchneriella subcapitata, Daphnia magna and Danio rerio. Sci. Total Environ. 2014, 466, 232–241. [Google Scholar] [CrossRef] [PubMed]
- Osborne, O.J.; Mukaigasa, K.; Nakajima, H.; Stolpe, B.; Romer, I.; Philips, U.; Lynch, I.; Mourabit, S.; Hirose, S.; Lead, J.R. Sensory systems and ionocytes are targets for silver nanoparticle effects in fish. Nanotoxicology 2016, 10, 1276–1286. [Google Scholar] [CrossRef] [PubMed]
- Clark, E.S.; Pompini, M.; Uppal, A.; Wedekind, C. Genetic correlations and little genetic variance for reaction norms may limit potential for adaptation to pollution by ionic and nanoparticulate silver in a whitefish (Salmonidae). Ecol. Evol. 2016, 6, 2751–2762. [Google Scholar] [CrossRef] [PubMed]
- Kwok, K.W.; Auffan, M.; Badireddy, A.R.; Nelson, C.M.; Wiesner, M.R.; Chilkoti, A.; Liu, J.; Marinakos, S.M.; Hinton, D.E. Uptake of silver nanoparticles and toxicity to early life stages of Japanese medaka (Oryzias latipes): effect of coating materials. Aquat. Toxicol. 2012, 120, 59–66. [Google Scholar] [CrossRef]
- Shoults-Wilson, W.A.; Reinsch, B.C.; Tsyusko, O.V.; Bertsch, P.M.; Lowry, G.V.; Unrine, J.M. Effect of silver nanoparticle surface coating on bioaccumulation and reproductive toxicity in earthworms (Eisenia fetida). Nanotoxicology 2011, 5, 432–444. [Google Scholar] [CrossRef]
- Yi, J.; Cheng, J. Effects of water chemistry and surface contact on the toxicity of silver nanoparticles to Bacillus subtilis. Ecotoxicology 2017, 26, 639–647. [Google Scholar] [CrossRef]
- Zhang, Q.; Cheng, J.; Xin, Q. Effects of tetracycline on developmental toxicity and molecular responses in zebrafish (Danio rerio) embryos. Ecotoxicology 2015, 24, 707–719. [Google Scholar] [CrossRef]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of embryonic development of the zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef]
- Griffitt, R.J.; Lavelle, C.M.; Kane, A.S.; Denslow, N.D.; Barber, D.S. Chronic nanoparticulate silver exposure results in tissue accumulation and transcriptomic changes in zebrafish. Aquat. Toxicol. 2013, 130, 192–200. [Google Scholar] [CrossRef]
- Jia, Y.; Fang, M.; Wu, Y.; Liu, H.; Miao, Y.; Wang, X.; Lin, W.; Tong, X. Pollution characteristics and potential ecological risk of heavy metals in river sediments of Shanghai. China Environ. Sci. 2013, 33, 147–153. [Google Scholar]
- Fung, C.; Lam, J.; Zheng, G.; Connell, D.; Monirith, I.; Tanabe, S.; Richardson, B.; Lam, P. Mussel-based monitoring of trace metal and organic contaminants along the east coast of China using Perna viridis and Mytilus edulis. Environ. Pollut. 2004, 127, 203–216. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organism | Source | Exposure Period | NP Sizes | Exposure Concentration | Ag Uptake of Negative Control | Ag Uptake | Biological Effects | References |
|---|---|---|---|---|---|---|---|---|
| Zebrafish Embryos | AgNPs | 0–4 days | 3–8 nm | 0.1 and 0.125 mg/L | — | — | Induced delayed hatching and formation of oedema. | [43] |
| AgNO3 | — | 0.04 and 0.08 mg/L | Ag+ decreased absorption of yolk sac and resulted in pericardial oedema. | |||||
| Zebrafish Embryos | AgNPs | 6–120 hpf | 10 nm | 1.7 and 5.1 mg/L | — | — | AgNPs induced slowed swim bladder inflation. | [20] |
| AgNO3 | — | 0.51, 1.7 and 5.1 mg/L | Ag+ slowed swim bladder inflation. | |||||
| Zebrafish Embryos | AgNPs | 4–24 and 4–48 hpf | 10 nm | 0.005 mg/L | — | — | AgNPs altered gene expression associated with oxidative phosphorylation and protein synthesis. | [44] |
| AgNO3 | 4–24 hpf | — | 0.00025 mg/L | Ag+ altered oxidative phosphorylation and protein synthesis gene expression. | ||||
| Adult Zebrafish | AgNPs | 0–4 days | 20 nm | 1 mg/L | Gill and intestine: No uptake detected | Gill: 5.11 μg/adult Intestine: 0.68 μg/adult | AgNPs decreased ATPase enzyme activity. | [23] |
| AgNO3 | — | Gill: 1.66 μg/adult Intestine: 2.29 μg/adult | Ag+ decreased ATPase enzyme activity. | |||||
| Zebrafish Embryos | AgNPs | 4–96 hpf | 10 nm | 1.925 mg/L | Head and trunk: No uptake detected | Head: 0.03 ng/embryo Trunk: 0.05 ng/embryo | AgNPs induced small head, small eye and cardiac defects, and altered the gene expression of neural development-related, metal-sensitive metallothioneins, and detoxification. | [22] |
| Zebrafish Embryos | AgNPs | 4–96 hpf | 4 nm | 1.925 mg/L | Whole body tissue: no uptake detected | Whole body tissue: 21.47 ng/g | AgNPs decreased body length, delayed yolk sac absorption, and altered oxidative stress and channel-forming protein gene expression. | This study |
| Adult Zebrafish | AgNPs | 0–48 h | 26.6 ± 8.8 nm | 1 mg/L | Carcass: 0.02 ng/g | Carcass: 0.47 ng/g | AgNPs altered signal transducer activity, enzyme activity and membrane function gene expression. | [21] |
| Zebrafis Embryos | AgNO3 | 1–24, 1–48 or 1–96 hpf | — | 0.02 mg/L | — | — | Ag+ altered detoxifying processes and oxidative stress gene expression. | [44] |
| Whitefis Embryos | AgNO3 | hatch | — | 0.1 mg/L | — | — | Ag+ accelerated hatching and decreased absorption of yolk sac. | [46] |
| Targeted Transcript | Forward Primer (5′-3′) | Reverse Primer (5′-3′) | Amplicon Size (bp) |
|---|---|---|---|
| 18S | TAGAGGGACAAGTGGCGTTC | CCTCGTTGATGGGAAACAGT | 195 |
| EF1 | GATGCACCACGAGTCTCTGA | TGATGACCTGAGCGTTGAAG | 158 |
| MUC | TGGGATCGCAAAACCACTGT | GTTGTGCATCAGGGCAAGTG | 187 |
| PXMP2 | AGTTACTGGCCTGCGATGAA | CACTGCAGTAAGGCACAACC | 157 |
| HIF | AGACTGCCACGGAAAAGCTA | CCAGAGGCAGAAGAGCAGTT | 161 |
| CAT | GCGGATACCAGAGAGAGTCG | ATCTGATGACCCAGCCTCAC | 172 |
| SOD2 | AGCGTGACTTTGGCTCATTT | ATGAGACCTGTGGTCCCTTG | 166 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qiang, L.; Arabeyyat, Z.H.; Xin, Q.; Paunov, V.N.; Dale, I.J.F.; Lloyd Mills, R.I.; Rotchell, J.M.; Cheng, J. Silver Nanoparticles in Zebrafish (Danio rerio) Embryos: Uptake, Growth and Molecular Responses. Int. J. Mol. Sci. 2020, 21, 1876. https://doi.org/10.3390/ijms21051876
Qiang L, Arabeyyat ZH, Xin Q, Paunov VN, Dale IJF, Lloyd Mills RI, Rotchell JM, Cheng J. Silver Nanoparticles in Zebrafish (Danio rerio) Embryos: Uptake, Growth and Molecular Responses. International Journal of Molecular Sciences. 2020; 21(5):1876. https://doi.org/10.3390/ijms21051876
Chicago/Turabian StyleQiang, Liyuan, Zeinab H. Arabeyyat, Qi Xin, Vesselin N. Paunov, Imogen J. F. Dale, Richard I. Lloyd Mills, Jeanette M. Rotchell, and Jinping Cheng. 2020. "Silver Nanoparticles in Zebrafish (Danio rerio) Embryos: Uptake, Growth and Molecular Responses" International Journal of Molecular Sciences 21, no. 5: 1876. https://doi.org/10.3390/ijms21051876
APA StyleQiang, L., Arabeyyat, Z. H., Xin, Q., Paunov, V. N., Dale, I. J. F., Lloyd Mills, R. I., Rotchell, J. M., & Cheng, J. (2020). Silver Nanoparticles in Zebrafish (Danio rerio) Embryos: Uptake, Growth and Molecular Responses. International Journal of Molecular Sciences, 21(5), 1876. https://doi.org/10.3390/ijms21051876