Development of Physiologically Responsive Human iPSC-Derived Intestinal Epithelium to Study Barrier Dysfunction in IBD

 , ,
, ,
Abstract
1. Introduction
2. Results
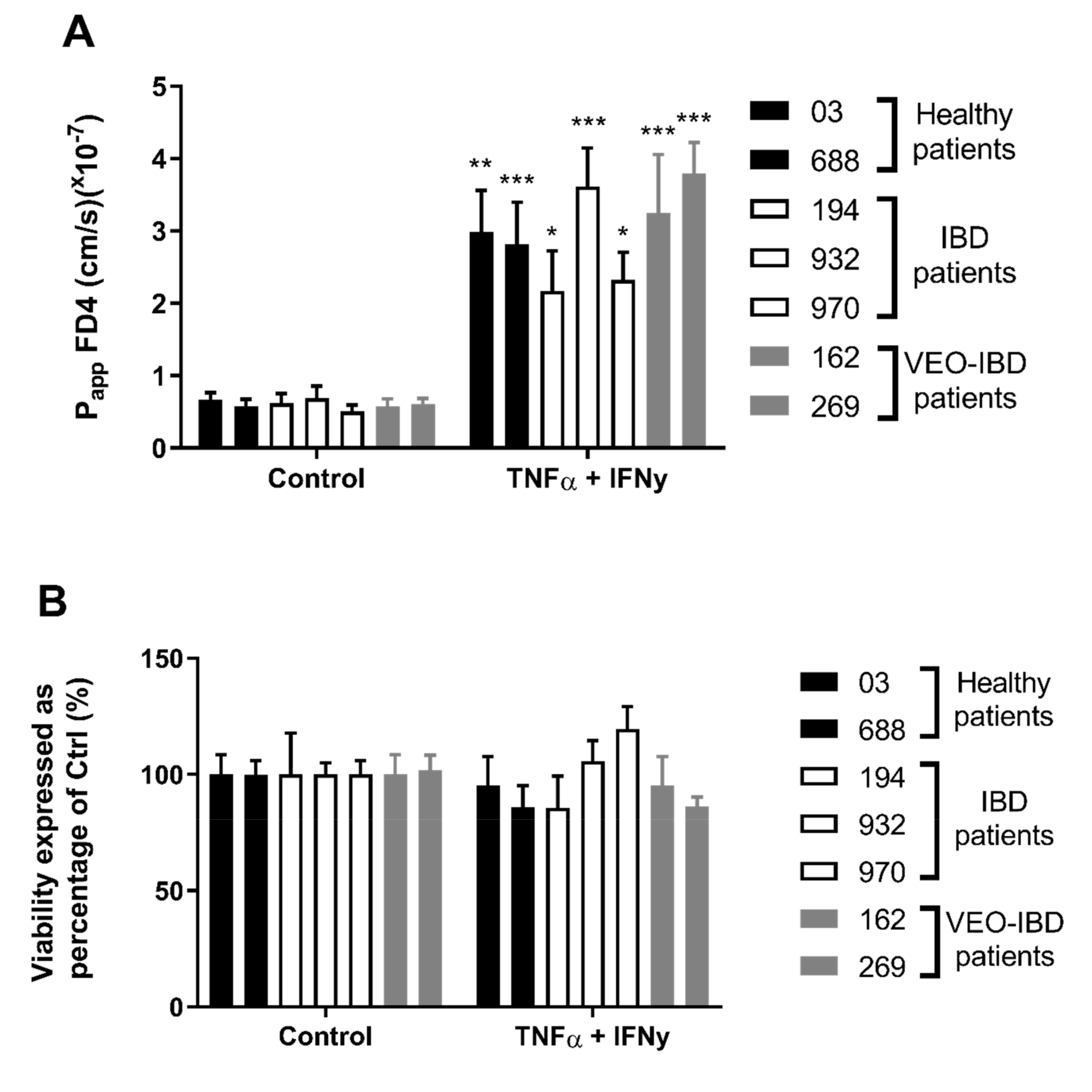
2.1. HIO-Derived Epithelium Becomes More Permeable When Exposed to Pro-Inflammatory Cytokines
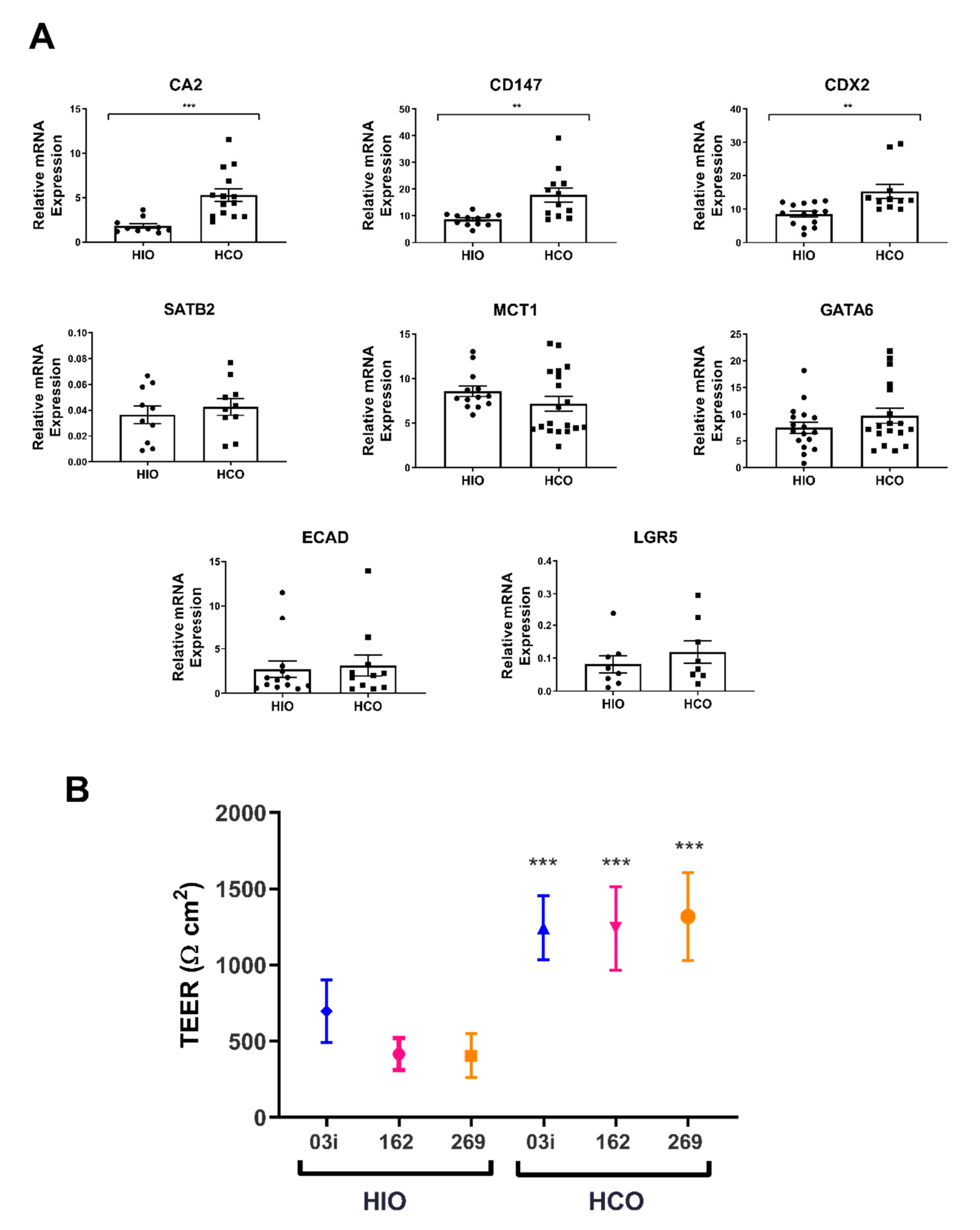
2.2. HCO-Derived Epithelium Reflects a Colonic Phenotype in Both Gene Expression and TEER
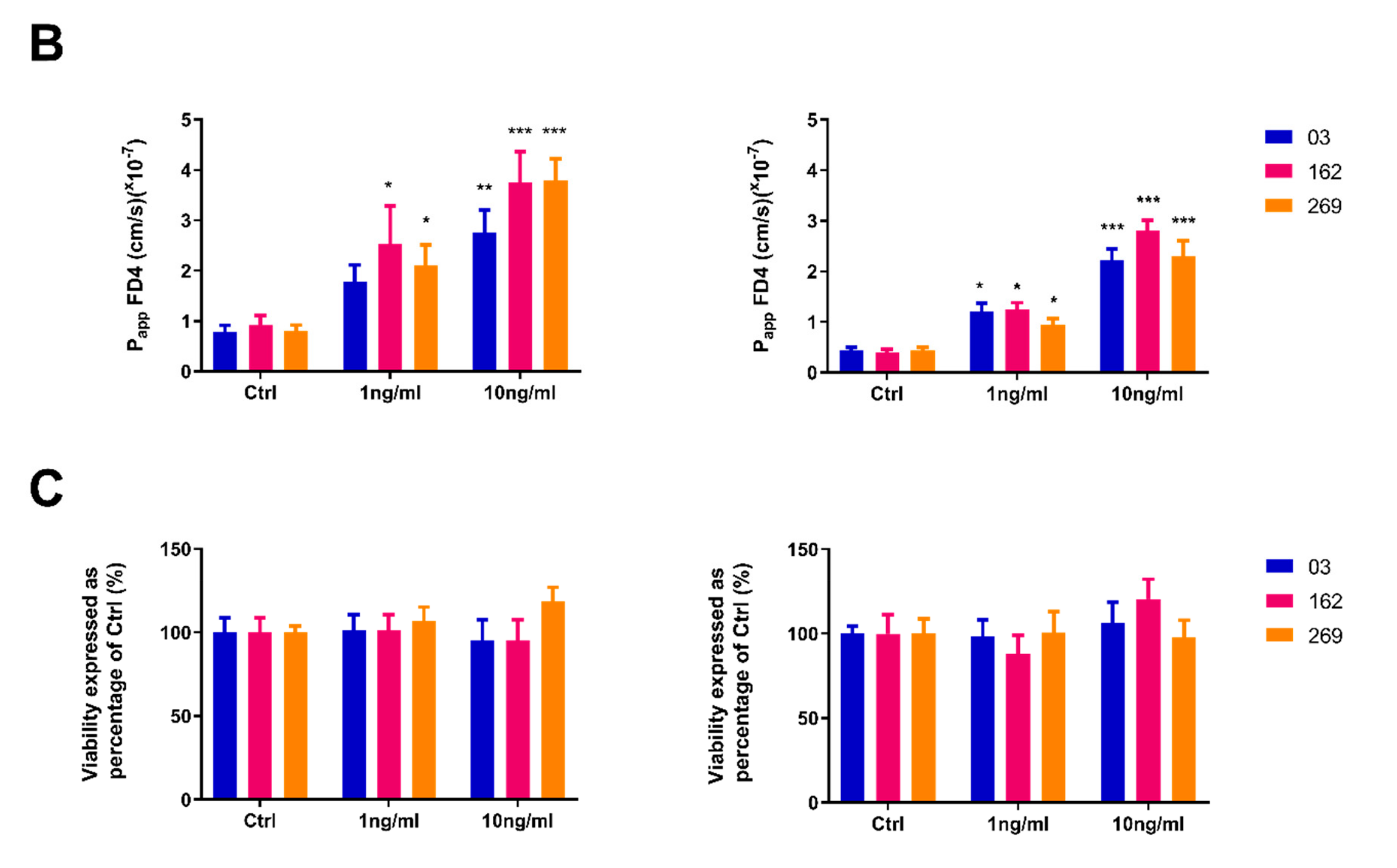
2.3. Pro-Inflammatory Cytokines Induce a Concentration-Dependent Increase in Permeability in both HIO and HCO Epithelium
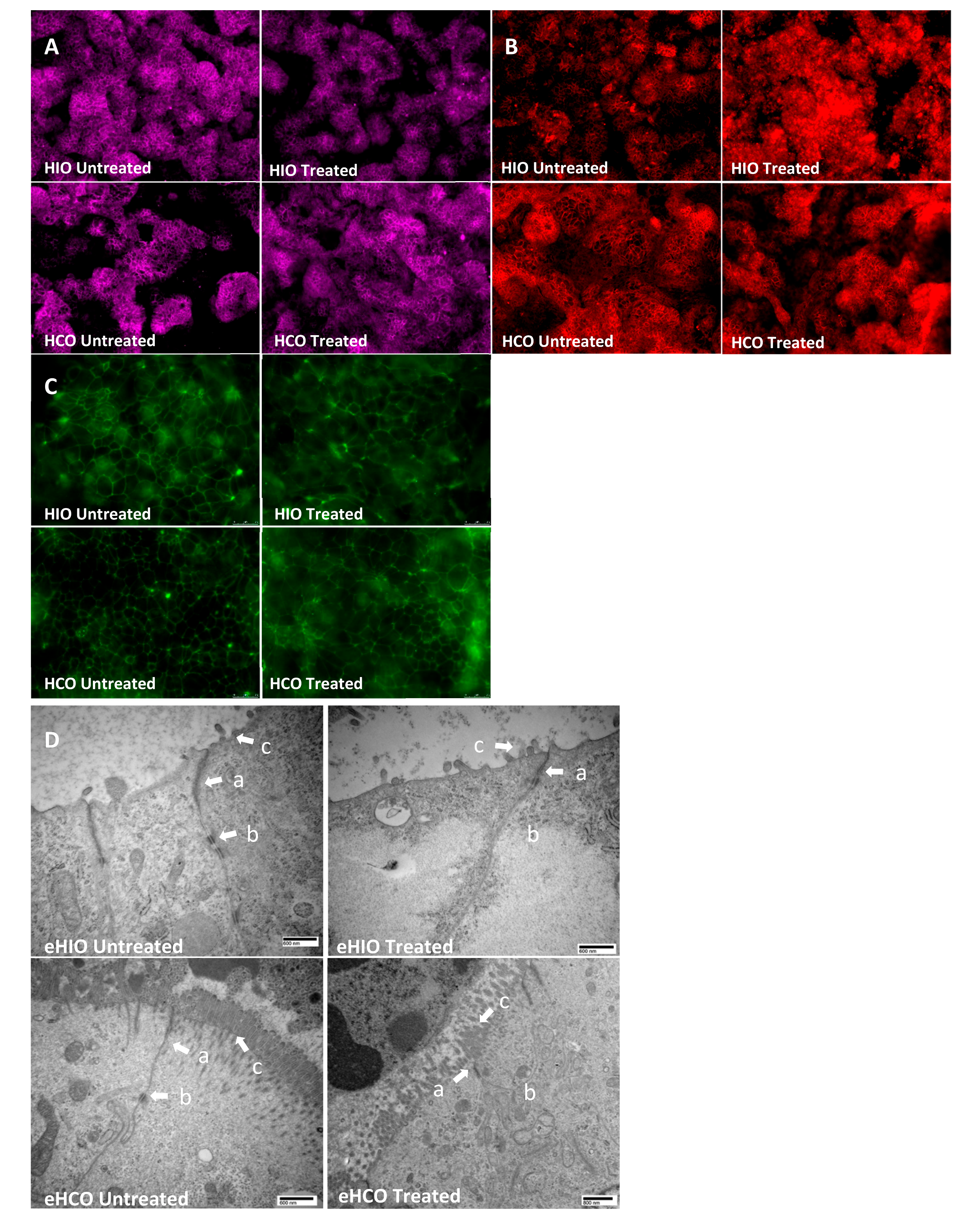
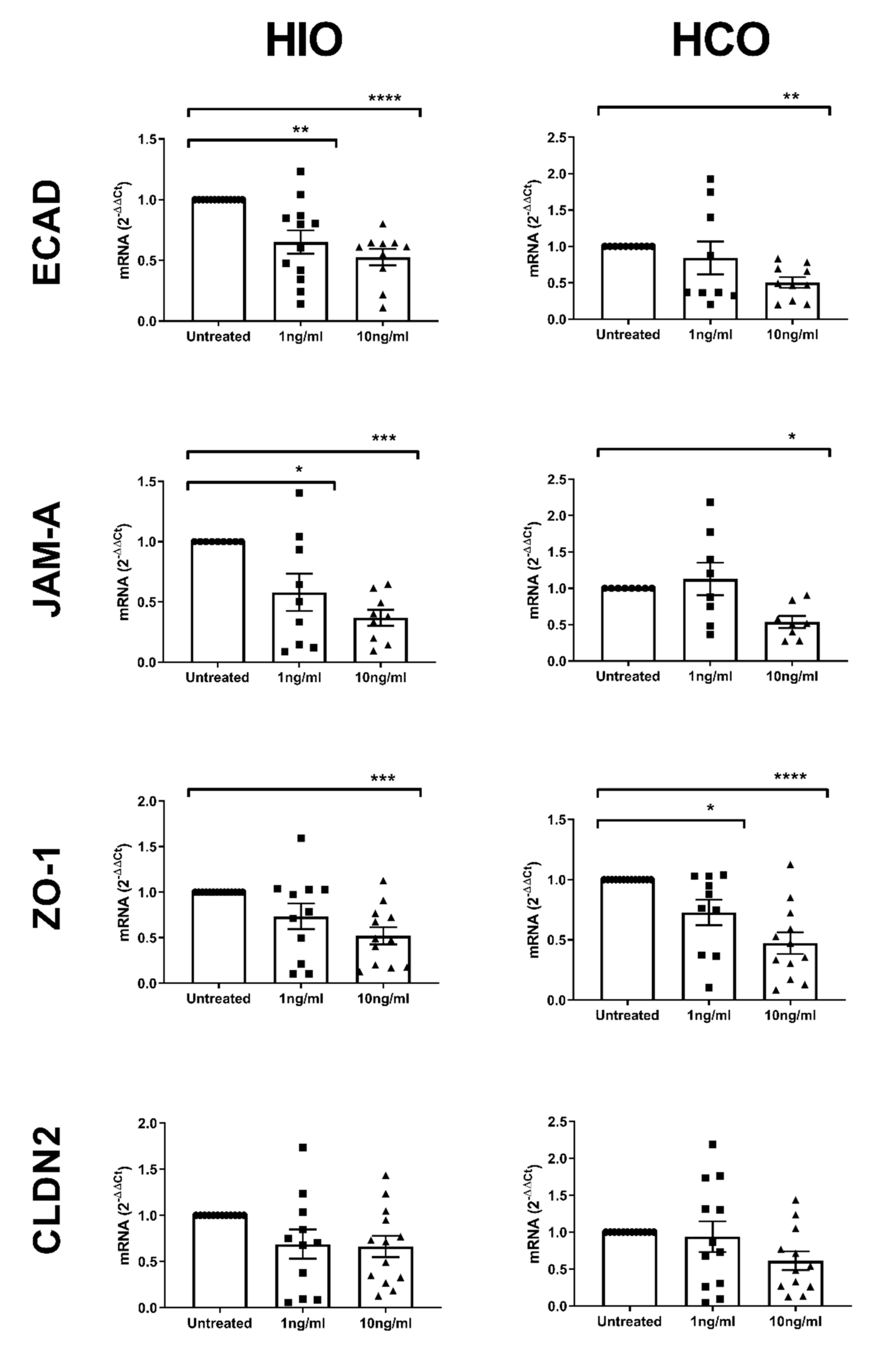
2.4. Tight and Adherens Junction Proteins are Downregulated and Mislocalized in HIO- and HCO-Epithelium Treated with Pro-Inflammatory Cytokines
3. Discussion
4. Materials and Methods
4.1. Ethical Statement
4.2. Cell Lines and Culturing
4.3. Organoid Differentiation From iPSCs
4.4. Cell Sorting and Transwell Seeding and Monolayer Maintenance
4.5. Transwell Cytokine Challenge: Permeability and Cytotoxicity Assay
4.6. Epithelial only-HIO/HCO Analysis: Regional Markers and Cytokine Challenge
4.7. Image Analysis: Immunofluorescence and Transmission Electron Microscopy
4.8. Statistical Analyses
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Welcker, K.; Martin, A.; Kolle, P.; Siebeck, M.; Gross, M. Increased intestinal permeability in patients with inflammatory bowel disease. Eur. J. Med. Res. 2004, 9, 456–460. [Google Scholar] [PubMed]
- Wyatt, J.; Vogelsang, H.; Hubl, W.; Waldhoer, T.; Lochs, H. Intestinal permeability and the prediction of relapse in Crohn’s disease. Lancet 1993, 341, 1437–1439. [Google Scholar] [CrossRef]
- Vivinus-Nebot, M.; Frin-Mathy, G.; Bzioueche, H.; Dainese, R.; Bernard, G.; Anty, R.; Filippi, J.; Saint-Paul, M.C.; Tulic, M.K.; Verhasselt, V.; et al. Functional bowel symptoms in quiescent inflammatory bowel diseases: Role of epithelial barrier disruption and low-grade inflammation. Gut 2014, 63, 744–752. [Google Scholar] [CrossRef] [PubMed]
- Hollander, D.; Vadheim, C.M.; Brettholz, E.; Petersen, G.M.; Delahunty, T.; Rotter, J.I. Increased intestinal permeability in patients with Crohn’ s disease and their relatives. A possible etiologic factor. Ann. Intern. Med. 1986, 105, 883–885. [Google Scholar] [CrossRef] [PubMed]
- Camoglio, L.; Te Velde, A.A.; Tigges, A.J.; Das, P.K.; Van Deventer, S.J. Altered expression of interferon-gamma and interleukin-4 in inflammatory bowel disease. Inflamm. Bowel Dis. 1998, 4, 285–290. [Google Scholar] [CrossRef]
- Suzuki, T. Regulation of intestinal epithelial permeability by tight junctions. Cell. Mol. Life Sci. 2013, 70, 631–659. [Google Scholar] [CrossRef]
- Hill, D.R.; Huang, S.; Tsai, Y.H.; Spence, J.R.; Young, V.B. Real-time Measurement of Epithelial Barrier Permeability in Human Intestinal Organoids. J. Vis. Exp. 2017, e56960. [Google Scholar] [CrossRef]
- Leslie, J.L.; Huang, S.; Opp, J.S.; Nagy, M.S.; Kobayashi, M.; Young, V.B.; Spence, J.R. Persistence and toxin production by Clostridium difficile within human intestinal organoids result in disruption of epithelial paracellular barrier function. Infect. Immun. 2015, 83, 138–145. [Google Scholar] [CrossRef]
- Spence, J.R.; Mayhew, C.N.; Rankin, S.A.; Kuhar, M.F.; Vallance, J.E.; Tolle, K.; Hoskins, E.E.; Kalinichenko, V.V.; Wells, S.I.; Zorn, A.M.; et al. Directed differentiation of human pluripotent stem cells into intestinal tissue in vitro. Nature 2011, 470, 105–109. [Google Scholar] [CrossRef] [PubMed]
- Barrett, R.; Ornelas, L.; Yeager, N.; Mandefro, B.; Sahabian, A.; Lenaeus, L.; Targan, S.R.; Svendsen, C.N.; Sareen, D. Reliable generation of induced pluripotent stem cells from human lymphoblastoid cell lines. Stem Cells Transl. Med. 2014, 3, 1429–1434. [Google Scholar] [CrossRef]
- Sato, T.; Stange, D.E.; Ferrante, M.; Vries, R.G.; Van Es, J.H.; Van den Brink, S.; Van Houdt, W.J.; Pronk, A.; Van Gorp, J.; Siersema, P.D.; et al. Long-term expansion of epithelial organoids from human colon, adenoma, adenocarcinoma, and Barrett’s epithelium. Gastroenterology 2011, 141, 1762–1772. [Google Scholar] [CrossRef] [PubMed]
- VanDussen, K.L.; Marinshaw, J.M.; Shaikh, N.; Miyoshi, H.; Moon, C.; Tarr, P.I.; Ciorba, M.A.; Stappenbeck, T.S. Development of an enhanced human gastrointestinal epithelial culture system to facilitate patient-based assays. Gut 2015, 64, 911–920. [Google Scholar] [CrossRef] [PubMed]
- Finkbeiner, S.R.; Hill, D.R.; Altheim, C.H.; Dedhia, P.H.; Taylor, M.J.; Tsai, Y.H.; Chin, A.M.; Mahe, M.M.; Watson, C.L.; Freeman, J.J.; et al. Transcriptome-wide Analysis Reveals Hallmarks of Human Intestine Development and Maturation In Vitro and In Vivo. Stem Cell Rep. 2015, 4, 1140–1155. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Inoue, H.; Wu, J.C.; Yamanaka, S. Induced pluripotent stem cell technology: A decade of progress. Nat. Rev. Drug Discov. 2017, 16, 115–130. [Google Scholar] [CrossRef] [PubMed]
- Munera, J.O.; Sundaram, N.; Rankin, S.A.; Hill, D.; Watson, C.; Mahe, M.; Vallance, J.E.; Shroyer, N.F.; Sinagoga, K.L.; Zarzoso-Lacoste, A.; et al. Differentiation of Human Pluripotent Stem Cells into Colonic Organoids via Transient Activation of BMP Signaling. Cell Stem Cell 2017, 21, 51–64. [Google Scholar] [CrossRef]
- Crespo, M.; Vilar, E.; Tsai, S.Y.; Chang, K.; Amin, S.; Srinivasan, T.; Zhang, T.; Pipalia, N.H.; Chen, H.J.; Witherspoon, M.; et al. Colonic organoids derived from human induced pluripotent stem cells for modeling colorectal cancer and drug testing. Nat. Med. 2017, 23, 878–884. [Google Scholar] [CrossRef]
- Jostins, L.; Ripke, S.; Weersma, R.K.; Duerr, R.H.; McGovern, D.P.; Hui, K.Y.; Lee, J.C.; Schumm, L.P.; Sharma, Y.; Anderson, C.A.; et al. Host-microbe interactions have shaped the genetic architecture of inflammatory bowel disease. Nature 2012, 491, 119–124. [Google Scholar] [CrossRef]
- Uhlig, H.H. Monogenic diseases associated with intestinal inflammation: Implications for the understanding of inflammatory bowel disease. Gut 2013, 62, 1795–1805. [Google Scholar] [CrossRef]
- Uhlig, H.H.; Schwerd, T.; Koletzko, S.; Shah, N.; Kammermeier, J.; Elkadri, A.; Ouahed, J.; Wilson, D.C.; Travis, S.P.; Turner, D.; et al. The diagnostic approach to monogenic very early onset inflammatory bowel disease. Gastroenterology 2014, 147, 990–1007. [Google Scholar] [CrossRef]
- Sjoberg, A.; Lutz, M.; Tannergren, C.; Wingolf, C.; Borde, A.; Ungell, A.L. Comprehensive study on regional human intestinal permeability and prediction of fraction absorbed of drugs using the Ussing chamber technique. Eur. J. Pharm. Sci. 2013, 48, 166–180. [Google Scholar] [CrossRef]
- Soderholm, J.D.; Hedman, L.; Artursson, P.; Franzen, L.; Larsson, J.; Pantzar, N.; Permert, J.; Olaison, G. Integrity and metabolism of human ileal mucosa in vitro in the Ussing chamber. Acta Physiol. Scand. 1998, 162, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Haslam, I.S.; O’Reilly, D.A.; Sherlock, D.J.; Kauser, A.; Womack, C.; Coleman, T. Pancreatoduodenectomy as a source of human small intestine for Ussing chamber investigations and comparative studies with rat tissue. Biopharm. Drug Dispos. 2011, 32, 210–221. [Google Scholar] [CrossRef] [PubMed]
- Mardones, P.; Andrinolo, D.; Csendes, A.; Lagos, N. Permeability of human jejunal segments to gonyautoxins measured by the Ussing chamber technique. Toxicon 2004, 44, 521–528. [Google Scholar] [CrossRef] [PubMed]
- Rogers, A.C.; McDermott, F.D.; Mohan, H.M.; O’Connell, P.R.; Winter, D.C.; Baird, A.W. The effects of polyamines on human colonic mucosal function. Eur. J. Pharmacol. 2015, 764, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Riegler, M.; Castagliuolo, I.; So, P.T.; Lotz, M.; Wang, C.; Wlk, M.; Sogukoglu, T.; Cosentini, E.; Bischof, G.; Hamilton, G.; et al. Effects of substance P on human colonic mucosa In Vitro. Am. J. Physiol. 1999, 276, G1473–G1483. [Google Scholar] [CrossRef] [PubMed]
- Riegler, M.; Sedivy, R.; Pothoulakis, C.; Hamilton, G.; Zacherl, J.; Bischof, G.; Cosentini, E.; Feil, W.; Schiessel, R.; LaMont, J.T.; et al. Clostridium difficile toxin B is more potent than toxin A in damaging human colonic epithelium In Vitro. J. Clin. Invest. 1995, 95, 2004–2011. [Google Scholar] [CrossRef] [PubMed]
- Maher, S.; Kennelly, R.; Bzik, V.A.; Baird, A.W.; Wang, X.; Winter, D.; Brayden, D.J. Evaluation of intestinal absorption enhancement and local mucosal toxicity of two promoters. I. Studies in isolated rat and human colonic mucosae. Eur. J. Pharm. Sci. 2009, 38, 291–300. [Google Scholar] [CrossRef] [PubMed]
- Sjogren, E.; Eriksson, J.; Vedin, C.; Breitholtz, K.; Hilgendorf, C. Excised segments of rat small intestine in Ussing chamber studies: A comparison of native and stripped tissue viability and permeability to drugs. Int. J. Pharm. 2016, 505, 361–368. [Google Scholar] [CrossRef]
- Gleeson, J.P.; Brayden, D.J.; Ryan, S.M. Evaluation of PepT1 transport of food-derived antihypertensive peptides, Ile-Pro-Pro and Leu-Lys-Pro using in vitro, ex vivo and in vivo transport models. Eur. J. Pharm. Biopharm. 2017, 115, 276–284. [Google Scholar] [CrossRef]
- Petersen, S.B.; Nolan, G.; Maher, S.; Rahbek, U.L.; Guldbrandt, M.; Brayden, D.J. Evaluation of alkylmaltosides as intestinal permeation enhancers: Comparison between rat intestinal mucosal sheets and Caco-2 monolayers. Eur. J. Pharm. Sci. Off. J. Eur. Fed. Pharm. Sci. 2012, 47, 701–712. [Google Scholar] [CrossRef]
- Gleeson, J.P.; Heade, J.; Ryan, S.M.; Brayden, D.J. Stability, toxicity and intestinal permeation enhancement of two food-derived antihypertensive tripeptides, Ile-Pro-Pro and Leu-Lys-Pro. Peptides 2015, 71, 1–7. [Google Scholar] [CrossRef]
- Feighery, L.; Smyth, A.; Keely, S.; Baird, A.W.; O’Connor, W.T.; Callanan, J.J.; Brayden, D.J. Increased intestinal permeability in rats subjected to traumatic frontal lobe percussion brain injury. J. Trauma 2008, 64, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Bjorkman, E.; Casselbrant, A.; Lundberg, S.; Fandriks, L. In Vitro assessment of epithelial electrical resistance in human esophageal and jejunal mucosae and in Caco-2 cell layers. Scand. J. Gastroenterol. 2012, 47, 1321–1333. [Google Scholar] [CrossRef] [PubMed]
- Hubbard, D.; Enda, M.; Bond, T.; Moghaddam, S.P.; Conarton, J.; Scaife, C.; Volckmann, E.; Ghandehari, H. Transepithelial Transport of PAMAM Dendrimers Across Isolated Human Intestinal Tissue. Mol. Pharm. 2015, 12, 4099–4107. [Google Scholar] [CrossRef] [PubMed]
- Takayama, K.; Negoro, R.; Yamashita, T.; Kawai, K.; Ichikawa, M.; Mori, T.; Nakatsu, N.; Harada, K.; Ito, S.; Yamada, H.; et al. Generation of Human iPSC-Derived Intestinal Epithelial Cell Monolayers by CDX2 Transduction. Cell. Mol. Gastroenterol. Hepatol. 2019, 8, 513–526. [Google Scholar] [CrossRef]
- Workman, M.J.; Gleeson, J.P.; Troisi, E.J.; Estrada, H.Q.; Kerns, S.J.; Hinojosa, C.D.; Hamilton, G.A.; Targan, S.R.; Svendsen, C.N.; Barrett, R.J. Enhanced Utilization of Induced Pluripotent Stem Cell-Derived Human Intestinal Organoids Using Microengineered Chips. Cell. Mol. Gastroenterol. Hepatol. 2018, 5, 669–677. [Google Scholar] [CrossRef]
- Wang, F.; Graham, W.V.; Wang, Y.; Witkowski, E.D.; Schwarz, B.T.; Turner, J.R. Interferon-gamma and tumor necrosis factor-alpha synergize to induce intestinal epithelial barrier dysfunction by up-regulating myosin light chain kinase expression. Am. J. Pathol. 2005, 166, 409–419. [Google Scholar] [CrossRef]
- Cao, M.; Wang, P.; Sun, C.; He, W.; Wang, F. Amelioration of IFN-gamma and TNF-alpha-induced intestinal epithelial barrier dysfunction by berberine via suppression of MLCK-MLC phosphorylation signaling pathway. PLoS ONE 2013, 8, e61944. [Google Scholar] [CrossRef]
- Wang, F.; Schwarz, B.T.; Graham, W.V.; Wang, Y.; Su, L.; Clayburgh, D.R.; Abraham, C.; Turner, J.R. IFN-gamma-induced TNFR2 expression is required for TNF-dependent intestinal epithelial barrier dysfunction. Gastroenterology 2006, 131, 1153–1163. [Google Scholar] [CrossRef]
- Till, A.; Rosenstiel, P.; Brautigam, K.; Sina, C.; Jacobs, G.; Oberg, H.H.; Seegert, D.; Chakraborty, T.; Schreiber, S. A role for membrane-bound CD147 in NOD2-mediated recognition of bacterial cytoinvasion. J. Cell Sci. 2008, 121, 487–495. [Google Scholar] [CrossRef]
- Mizoshita, T.; Inada, K.-I.; Tsukamoto, T.; Kodera, Y.; Yamamura, Y.; Hirai, T.; Kato, T.; Joh, T.; Itoh, M.; Tatematsu, M. Expression of Cdx1 and Cdx2 mRNAs and Relevance of This Expression to Differentiation in Human Gastrointestinal Mucosa--With Special Emphasis on Participation in Intestinal Metaplasia of the Human Stomach. Gastric Cancer 2001, 4, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Englund, G.; Rorsman, F.; Ronnblom, A.; Karlbom, U.; Lazorova, L.; Grasjo, J.; Kindmark, A.; Artursson, P. Regional levels of drug transporters along the human intestinal tract: Co-expression of ABC and SLC transporters and comparison with Caco-2 cells. Eur. J. Pharm. Sci. 2006, 29, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Al-Mosauwi, H.; Ryan, E.; McGrane, A.; Riveros-Beltran, S.; Walpole, C.; Dempsey, E.; Courtney, D.; Fearon, N.; Winter, D.; Baird, A.; et al. Differential protein abundance of a basolateral MCT1 transporter in the human gastrointestinal tract. Cell Biol. Int. 2016, 40, 1303–1312. [Google Scholar] [CrossRef] [PubMed]
- Haveri, H.; Westerholm-Ormio, M.; Lindfors, K.; Maki, M.; Savilahti, E.; Andersson, L.C.; Heikinheimo, M. Transcription factors GATA-4 and GATA-6 in normal and neoplastic human gastrointestinal mucosa. BMC Gastroenterol. 2008, 8, 9. [Google Scholar] [CrossRef]
- Dogan, A.; Wang, Z.D.; Spencer, J. E-cadherin expression in intestinal epithelium. J. Clin. Pathol. 1995, 48, 143–146. [Google Scholar] [CrossRef]
- Nejdfors, P.; Ekelund, M.; Jeppsson, B.; Westrom, B.R. Mucosal in vitro permeability in the intestinal tract of the pig, the rat, and man: Species- and region-related differences. Scand. J. Gastroenterol. 2000, 35, 501–507. [Google Scholar] [CrossRef]
- Mukherjee, T.M.; Williams, A.W. A comparative study of the ultrastructure of microvilli in the epithelium of small and large intestine of mice. J. Cell Biol. 1967, 34, 447–461. [Google Scholar] [CrossRef]
- Bruewer, M.; Luegering, A.; Kucharzik, T.; Parkos, C.A.; Madara, J.L.; Hopkins, A.M.; Nusrat, A. Proinflammatory cytokines disrupt epithelial barrier function by apoptosis-independent mechanisms. J. Immunol. 2003, 171, 6164–6172. [Google Scholar] [CrossRef]
- Al-Sadi, R.M.; Ma, T.Y. IL-1beta causes an increase in intestinal epithelial tight junction permeability. J. Immunol. 2007, 178, 4641–4649. [Google Scholar] [CrossRef]
- Ma, T.Y.; Iwamoto, G.K.; Hoa, N.T.; Akotia, V.; Pedram, A.; Boivin, M.A.; Said, H.M. TNF-alpha-induced increase in intestinal epithelial tight junction permeability requires NF-kappa B activation. Am. J. Physiol. Gastrointest Liver Physiol. 2004, 286, G367–G376. [Google Scholar] [CrossRef]
- Haas, M.; Mirocha, J. Early ultrastructural changes in renal allografts: Correlation with antibody-mediated rejection and transplant glomerulopathy. Am. J. Transpl. 2011, 11, 2123–2131. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tissue Source | TEER Values (Ω.cm2) | Fold Difference | References |
|---|---|---|---|
| Human small intestine | 23–45 | - | [20,21,22,23] |
| Human colon | 103–174 | 4-fold | [23,24,25,26,27] |
| Rat small intestine | 30–37 | - | [22,28,29] |
| Rat colon | 84–116 | 3-fold | [27,30,31,32] |
| HIO | 478 ± 24 | - | |
| HCO | 1268 ± 50 | 3-fold |
| Gene | Forward | Reverse |
|---|---|---|
| CD147 | ACTCCTCACCTGCTCCTTGA | GTCCACCTTGAACTCCGTTTTC |
| CDX2 | TTCCTCTCCTTTGCTCTGCG | AGTCGCTACATCACCATCCG |
| CA2 | AAACACAACGGACCTGAGCA | GGAAACAGACAGGGGCTTCA |
| SATB2 | GCAAAGCCAAGCACCAGAAA | TAAAACGCACAGGGACTGCT |
| GATA6 | TGCTATTACCAGAGCAAGTCTTTG | TGTGCAATGCTTGTGGACTC |
| MCT1 | CCACCACTTTTAGGTCGGCT | TGCCCATGCCAATGAAGAGA |
| LGR5 | GAACACTGACTCTGAATGGTGC | ACGGTTTGAGGAAGAGATGAGA |
| ZO-1 | ACAGCAATGGAGGAAACAGC | CCCCACTCTGAAAATGAGGA |
| JAM-A | TCATATTGGCGATCCTGTTG | AGGCACAGGACAACTTCACA |
| ECAD | GCCGAGAGCTACACGTTCAC | GTCGAGGGAAAAATAGGCTG |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gleeson, J.P.; Estrada, H.Q.; Yamashita, M.; Svendsen, C.N.; Targan, S.R.; Barrett, R.J. Development of Physiologically Responsive Human iPSC-Derived Intestinal Epithelium to Study Barrier Dysfunction in IBD. Int. J. Mol. Sci. 2020, 21, 1438. https://doi.org/10.3390/ijms21041438
Gleeson JP, Estrada HQ, Yamashita M, Svendsen CN, Targan SR, Barrett RJ. Development of Physiologically Responsive Human iPSC-Derived Intestinal Epithelium to Study Barrier Dysfunction in IBD. International Journal of Molecular Sciences. 2020; 21(4):1438. https://doi.org/10.3390/ijms21041438
Chicago/Turabian StyleGleeson, John P., Hannah Q. Estrada, Michifumi Yamashita, Clive N. Svendsen, Stephan R. Targan, and Robert J. Barrett. 2020. "Development of Physiologically Responsive Human iPSC-Derived Intestinal Epithelium to Study Barrier Dysfunction in IBD" International Journal of Molecular Sciences 21, no. 4: 1438. https://doi.org/10.3390/ijms21041438
APA StyleGleeson, J. P., Estrada, H. Q., Yamashita, M., Svendsen, C. N., Targan, S. R., & Barrett, R. J. (2020). Development of Physiologically Responsive Human iPSC-Derived Intestinal Epithelium to Study Barrier Dysfunction in IBD. International Journal of Molecular Sciences, 21(4), 1438. https://doi.org/10.3390/ijms21041438